Embed Size (px)
Citation preview

LAB INVESTIGATION-HUMAN/ANIMAL TISSUE
Type I collagen is overexpressed in medulloblastomaas a component of tumor microenvironment
Yu Liang Æ Maximilian Diehn Æ Andrew W. Bollen ÆMark A. Israel Æ Nalin Gupta
Received: 27 May 2007 / Accepted: 25 June 2007 / Published online: 25 July 2007
� Springer Science+Business Media B.V. 2007
Abstract Medulloblastoma is the most common malig-
nant brain tumor of children, and more specific and
effective therapeutic management needs to be developed to
improve upon existing survival rates and to avoid side-
effects from current treatment. Gain of chromosome seven
is the most frequent chromosome copy number aberration
in medulloblastoma, suggesting that overexpression of
genes on chromosome seven might be important for the
pathogenesis of medulloblastoma. We used microarrays to
identify chromosome seven genes overexpressed in
medulloblastoma specimens, and validated using data from
published gene expression datasets. The gene encoding the
alpha 2 subunit of type I collagen, COL1A2, was overex-
pressed in all three datasets. Immunohistochemistry of
tumor tissues revealed type I collagen in the leptomenin-
ges, and in the extracellular matrix surrounding blood
vessels and medulloblastoma cells. Expression of both type
I collagen and the b1 subunit of integrin, a subunit of a
known type I collagen receptor, localized to the same area
of medulloblastoma. Adherence of D283 medulloblastoma
cells to type I collagen matrix in vitro depends on the b1
subunit of integrin. Because medulloblastoma is charac-
teristic of high vascularity, and because inhibition of type I
collagen synthesis has been shown to suppress angiogen-
esis and tumor growth, our data suggest that type I collagen
might be a potential therapeutic target for treating medul-
loblastoma.
Keywords Medulloblastoma � Microarray � Extracellular
matrix � Type I collagen � Adhesion
Introduction
Brain tumors are the primary cause of non-traumatic death
in children and young adults under the age of 20 years.
Primitive neuroectodermal tumors (PNETs) are the most
common malignant brain tumor in children and account for
25% of all pediatric brain tumors [1]. PNETs can arise in a
supratentorial (cerebral) or infratentorial (cerebellar) loca-
tion but most PNETs occurring in children arise within the
cerebellum. These cerebellar tumors are also called
medulloblastoma. The standard treatment for medulloblas-
toma is surgical resection followed by fractionated external
beam radiation and/or chemotherapy [2]. Although medul-
loblastoma is sensitive to both therapeutic modalities,
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11060-007-9457-5) contains supplementarymaterial, which is available to authorized users.
Y. Liang � N. Gupta
Department of Neurological Surgery, Brain Tumor Research
Center, University of California, San Francisco, CA 94143, USA
Y. Liang (&)
Division of Molecular Cell Biology, Applied Biosystems, 850
Lincoln Centre Drive, Foster City, CA 94404, USA
e-mail: [email protected]
M. Diehn
Department of Radiation Oncology, Stanford University School
of Medicine, Stanford, CA 94305, USA
M. Diehn
Department of Biochemistry, Stanford University School
of Medicine, Stanford, CA 94305, USA
A. W. Bollen
Department of Pathology, University of California,
San Francisco, CA 94143, USA
M. A. Israel
Departments of Pediatrics and Genetics, Norris Cotton Cancer
Center, Dartmouth Medical School, Lebanon, NH 03756, USA
123
J Neurooncol (2008) 86:133–141
DOI 10.1007/s11060-007-9457-5

irradiation of the central nervous system (CNS) in young
children often results in serious long-term side effects, such
as deafness, cognitive decline, and neuroendocrine insuffi-
ciency. For this reason, a variety of chemotherapy regimens
are used as initial treatment in the very young patients to
delay the use of radiation therapy. Early diagnosis and
improved treatment have increased the 5 year survival rate
of patients with localized medulloblastoma to greater than
60% [3, 4].
Invasion of the leptomeninges by medulloblastoma cells
and their dissemination via the subarachnoid spaces
throughout the neuraxis to distant sites can occur either at
presentation or at relapse and is associated with poor sur-
vival [5]. Genetic studies and the development of trans-
genic mouse models have provided important information
about the cellular origin and oncogenic pathways under-
lying medulloblastoma [6]. The molecular mechanisms
responsible for invasion into and dissemination within the
neuraxis remain largely unknown. A recent study using
DNA microarrays suggested that expression of platelet-
derived growth factor a and the RAS/mitogen-activated
protein kinase signal transduction pathway might be
up-regulated in disseminated medulloblastoma and could
be targets for more effective treatments [7].
Both medulloblastoma and supratentorial PNET are
malignant and invasive embryonal tumors that have similar
histological features, but the survival for children with
supratentorial PNET is much poorer, only 20–30% at
5 years [8–10]. The degree of surgical resection may play a
role in the improved survival of medulloblastoma, since
gross total resection of supratentorial PNET is usually
more difficult to achieve. PNET arise from both locations
exhibit high microvascular density and express wide range
of angiogenic factors [11, 12]. Despite these similarities,
recent evidence has shown that medulloblastoma and
supratentorial PNET are two genetically distinct tumor
types, primarily based upon gene expression profiling [13]
and the patterns of chromosome copy number aberrations
from comparative genomic hybridization [14–17]. One
important observation is that majority of these studies also
demonstrated that gain of chromosome seven is the most
frequent copy number alteration shared by both medullo-
blastoma and supratentorial PNET [14–18]. For example,
gain of chromosome seven was detected in 44–57% of
medulloblastoma and in 66% of supratentorial PNET [17,
18]. For this reason, we sought to identify genes located on
the chromosome seven which are consistently overex-
pressed in both tumor types might provide the genetic basis
for shared features that characterize these tumors, such as
tumoral vascularity.
In the present study, we used cDNA microarrays to
examine the expression profiles of chromosome seven
genes in three medulloblastoma and three supratentorial
PNET specimens, and in normal brain tissue. We validated
the results with published datasets and confirmed overex-
pression of COL1A2 in both these tumor types. This gene
and COL1A1, each encodes a distinctive subunit that
together heterodimerizes forming type I collagen. We also
detected increased accumulation of type I collagen in
medulloblastoma by immunohistochemistry.
Materials and methods
Cell culture
D283 medulloblastoma cell line was obtained from the
Brain Tumor Research Center Tissue Bank at UCSF. All
cells were maintained in Eagle’s minimal essential medium
with 15% fetal bovine serum and 5% CO2.
Tissue specimens
Medulloblastoma and normal brain specimens were
obtained from the Brain Tumor Research Center Tissue
Bank at UCSF after approval by the Committee on Human
Research.
Microarray and bioinformatic analyses
The detailed microarray methods were published at the
website http://microarray-pubs.stanford.edu/gbm/. Briefly,
total RNA was extracted using Trizol followed by mRNA
purification using FastTrack (Invitrogen). mRNA was
reverse transcribed to cDNA and directly labeled with Cy
dyes (Amersham Biosciences) before hybridization. The
raw data of 22,636 features on the array were extracted
from the Stanford Microarray Database using the ScanA-
lyze-featured extraction software with the following
settings: regression correlation >0.6, channel 1 mean
intensity/median background intensity ‡1.5, and channel
two normalized mean intensity/median background inten-
sity ‡1.5. The DNA sequences of individual clones from
the extracted raw data were examined for chromosomal
locations with the ‘‘Table Browser’’ function at the UCSC
Genome Bioinformatics website (the Human May 2004
assembly at http://genome.ucsc.edu), and those localized
on chromosome seven (1,116 features) were identified.
Among these 1,116 features, only those with available data
in three or more tumor specimens and in one or more
normal brain specimens were selected for final analyses
(757 features).
Gene expression data of three independent published
datasets (four groups of tumor samples in total) for veri-
fication of expression of the COL1A1 and COL1A2 genes
were extracted from the Broad Institute Cancer Program
134 J Neurooncol (2008) 86:133–141
123

Datasets (http://www.broad.mit.edu/cancer/datasets.html)
and Serial Analysis of Gene Expression (SAGE) Anatomic
Viewer (http://cgap.nci.nih.gov/SAGE/AnatomicViewer).
Antibodies
Anti-b1 subunit of integrin (ITGB1), anti-type I collagen,
and anti-integrin linked kinase polyclonal antibodies were
from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-
ITGB1 monoclonal antibodies AIIB2 and P4G11 were
from Developmental Studies Hybridoma Bank (Iowa
City, IA). Anti-CD31 and anti-fibronectin antibodies were
from Novocastro Laboratories (United Kingdom) and
GIBCO-BRL (Gaithersburg MD), respectively. Peroxi-
dase-conjugated and biotinylated secondary antibodies
were from Vector Laboratories (Burlingame, CA). Fluo-
rescine-conjugated and Rhodamine-conjugated secondary
antibodies and normal serum were from Jackson Immu-
noResearch Laboratories (West Grove, PA).
Immunohistochemistry
Frozen tissue sections used for immunohistochemistry
were fixed in 4% formaldehyde, treated with H2O2,
blocked with normal serum, incubated with primary anti-
bodies at 4�C overnight or room temperature (RT) 2 h,
incubated with biotinylated secondary antibody and per-
oxidase-labeled streptavidin at RT for 30 min, and visu-
alized using the DAB Reagent kit (KPL; Gaithersburg,
MA). Some frozen sections were visualized using FITC- or
Texas Red-conjugated secondary antibodies (Vector Lab-
oratories). Staining of paraffin-embedded sections followed
the same protocol, except for prior de-waxing and antigen
retrieval by microwave heating.
Adhesion assay
Wells of 96-well tissue culture plates (Corning, Corning,
NY) were coated with 100 lg/ml of rat-tail type I collagen
(BD Biosciences, Bedford, MA) for 1 hour at room tem-
perature, followed by two PBS washes. 1 · 105 D283 cells
resuspended in serum-free medium were plated into either
uncoated or type I collagen-coated wells for 1 hour. Non-
adhered cells were removed by lightly tapping the plates
and the number of cells remaining adherent was counted.
Monoclonal antibodies AIIB2 and P4G11 were diluted to
1:20 into the serum-free medium during a 1-hour incuba-
tion to inhibit cell adhesion.
Data analysis
All statistical analyses used the Student’s t test (2-tailed
and equal variance not assumed) and Pearson correlation in
SPSS for Windows (Release 11.5.0). A P value £ 0.05 was
considered statistically significant.
Immunoprecipitation and immunoblotting
D283 cells were lysed in RIPA buffer (in 1% NP-40, 0.1%
SDS, 0.5% Na deoxycholate, 50 mM Tris pH 8.0, and
150 mM NaCl) supplemented with 1 mM NaF, 1 mM
Na3VO4, and Complete protease inhibitor cocktail tablets
(Roche, Basel, Switzerland). After high-speed centrifuga-
tion, the lysate was pre-cleared twice with protein A
Sepharose beads (Zymed Laboratories, South San Fran-
cisco) followed by mixing with 1:100 dilution of anti-
bodies at 4�C for 1 h. Immunoprecipitated proteins were
captured by protein G Sepharose beads (Zymed Labora-
tories) and released by sample buffer. Samples were sep-
arated by SDS-PAGE and transferred to nitrocellulose
membranes, followed by 10% skim milk blocking and
antibody incubation, and visualized using the Super Signal
West Pico Chemiluminescent Substrate (Pierce, Rockford,
IL).
Results
Gene expression profiling identifies genes on
chromosome seven overexpressed in primitive
neuroectodermal tumors compared to normal brain
We performed gene expression profiling of three medul-
loblastoma and three supratentorial PNETs using cDNA
microarrays that consisted of ~23,000 features (represent-
ing ~18,000 unique UniGene clusters), and compared the
results to the expression patterns we observed for two
specimens derived from normal cerebrum and one from
normal cerebellum. After data filtering, we selected
approximately 750 genes located on chromosome seven
(see Methods and Additional file 1 for data), and computed
the difference of their expression between all six tumors
and the three normal brain specimens. Six UniGene clones
representing five different genes, ETV1, CDK6, COL1A2,
TAC1, and CAV2, had average expression in tumors at least
4-fold higher when compared to normal tissues (Fig. 1).
Differences in gene expression between tumor and normal
brain specimens reached significance in three genes using
the Student’s t test (ETV1, P = 0.006; CDK6, P = 0.0005;
COL1A2, P = 0.009).
COL1A2 mRNA is overexpressed in independent
groups of primitive neuroectodermal tumors
We verified the increased expression of ETV1, CDK6, and
COL1A2 using other gene expression datasets. In a recent
J Neurooncol (2008) 86:133–141 135
123

study, oligonucleotide microarrays were used to charac-
terize gene expression from embryonal tumors of the CNS,
and found a correlation between gene expression and the
clinical outcome of patients with medulloblastoma [13].
Their study included two groups of medulloblastoma and
supratentorial PNET specimens: the first group had 10
medulloblastoma and eight supratentorial PNET (Fig. 2a),
and the second group consisted of 60 medulloblastoma and
six supratentorial PNET (Fig. 2b). We reviewed the
expression of AEBP1, CDK6, and COL1A2 in these two
groups, and we found that COL1A2 but not the other two
genes demonstrated higher expression in tumors than in
normal cerebellum with a P value of less than 0.05 (see
Additional file 2 for data). We also identified increased
expression of COL1A2 in an additional published dataset
[19] that included 10 medulloblastomas (Fig. 2c).
Finally, we sought to validate this finding using gene
expression data from the SAGE Anatomic Viewer [20] that
displays gene expression in human normal and malignant
tissues based on the number of SAGE tags corresponding
to individual mRNAs [21]. In this database, expression of
COL1A2 in 20 medulloblastoma specimens was signifi-
cantly higher when compared to two normal cerebellum
specimens (Fig. 2d, and see Additional file 3 for data).
Type I collagen is overexpressed in medulloblastoma
COL1A2 gene encodes the alpha 2 subunit of type I col-
lagen. Unlike other types of collagen in which the subunits
of the collagen fibrils heterodimerize in several possible
combinations, type I collagen is always composed of one
alpha 2 chain and two alpha 1 chains (encoded by the
COL1A1 gene). Expression of type I collagen is regulated
primarily at the transcriptional level, and common cis-
acting regulatory elements are present in the promoters of
both COL1A1 and COL1A2 genes [22, 23] This is consis-
tent with the coherent expression patterns between
COL1A1 and COL1A2 in the two groups of normal,
medulloblastoma, and supratentorial PNET specimens used
by Pomeroy et al. [13] in which we verified COL1A2
expression in Figs. 2a,b (group 1, N = 20, P = 1 · 10–8;
group 2, N = 68, P = 2.4 · 10–15; both by Pearson corre-
lation and see Additional file 2 for data). As expected,
COL1A1 mRNA was also significantly higher in these two
sets of medulloblastoma and supratentorial PNET than in
normal cerebellum (Figs. 2ab, and Additional file 2 for
data), so was the number of SAGE tags corresponding to
COL1A1 significantly higher in medulloblastoma than in
normal cerebellum (Fig. 2d).
Overexpression of both the COL1A1 and COL1A2 genes
in both medulloblastoma and supratentorial PNET strongly
suggests increased production of type I collagen in their
tumor microenvironment. We examined the expression of
type I collagen protein in a panel of 17 human medullo-
blastoma specimens using immunohistochemistry. In nor-
mal brain, type I collagen was detected in the
leptomeninges and basement membrane associated with
blood vessels but not in brain parenchyma (Fig. 3a). Two
patterns of type I collagen expression were observed in the
tumor specimens. The first pattern consisted of type I
collagen immunoreactivity surrounding blood vessels,
while the ECM between neoplastic cells was essential
negative (Fig. 3b). The boundaries of the type I collagen-
positive basement membrane were sharply demarcated.
The second pattern of type I collagen immunostaining was
not restricted in the ECM of blood vessels but was dif-
fusely associated with thickened and distended basement
membrane of the tumor vasculature and throughout the
ECM within the tumor (Fig. 3c, e, g). We noticed that both
immunostaining patterns occurred together in some tumors
(data not shown).
b1 integrin subunit is detected in type I collagen-
surrounded medulloblastoma cells and mediates their
in vitro adhesion to type I collagen matrix
Our immunohistochemical data indicate that type I colla-
gen is present in the basement membrane associated with
intra-tumoral vasculature as well as in the ECM sur-
rounding neoplastic cells. This suggests that type I colla-
gen-containing ECM interacts with at least a subset of
neoplastic cells. The b1 subunit of integrin (ITGB1) is a
known constituent of receptors for type I collagen, and
Fig 1 Expression of genes located on chromosome seven in
medulloblastoma and supratentorial PNET specimens compared to
normal brain tissue. The average expression of each gene in three
normal brain specimens was subtracted from its average expression in
all six tumor specimens, and plotted against the base position of this
gene on chromosome seven; the value of gene expression (Y-axis) is
in log2 scale. Five genes with the values greater than two (4-fold
higher expression in tumor than in normal) were selected for further
consideration: from left to right ETV1, CDK6 (two clones), COL1A2,
TAC1, and CAV2. The Student’s t test showed that expression of
ETV1, CDK6, and COL1A2 was significantly greater in tumors than in
normal brains, while expression of TAC1 and CAV2 was not
136 J Neurooncol (2008) 86:133–141
123
could be a candidate for mediating the cellular receptor to
type I collagen. We investigated this possibility by first
examining localization of the ITGB1 in medulloblastoma
tumor tissues. We contrasted the CD31 immunoreactivity
that stains endothelial cells (Fig. 3f) to the type I collagen
immunoreactivity that stains basement membrane of the
vessels and tumoral ECM (Fig. 3g). ITGB1 was detected in
endothelial cells as well as neoplastic cells in regions
where the ECM contained type I collagen (Fig. 3h). Dual
immunofluorescence microscopy also showed that type I
collagen-positive tumoral ECM overlapped with ITGB1
immunoreactivity (Figs. 3j–l).
We then characterized whether type I collagen mediates
medulloblastoma cell adhesion by examining the adhesion
of D283 medulloblastoma cells to type I collagen matrix
in vitro. One hour after plating, D283 cells started to
adhere to a type I collagen-coated surface whereas they
could be easily removed from an uncoated surface by
lightly tapping a culture dish (Fig. 4). Most D283 cells
detached from either coated or uncoated surface after
vigorous shaking (data not shown). AIIB2 and P4G11 are
two monoclonal antibodies that have been previously
characterized for their opposite effects on the ITGB1-
mediated functions, in that AIIB2 blocks binding of b1
integrin subunit to its ligand, [24] whereas P4G11 stimu-
lates the binding [25]. Adherence of D283 cells to type I
collagen is ITGB1-dependent, as AIIB2 abolished the
attachment of cells to type I collagen, while P4G11 had no
such effect (Fig. 4). Our preliminary experiments demon-
strated that the integrin-linked kinase (ILK), a kinase
downstream of ITGB1, was detectable only in neoplastic
cells positive for ITGB1 (Fig. 3i). Anti-type I collagen
polyclonal antibodies co-immunoprecipitated ILK from
D283 cells (Additional file 4) suggesting that medullo-
blastoma cells interacting with type I collagen ECM via
ITGB1 might trigger signaling pathways involving ILK.
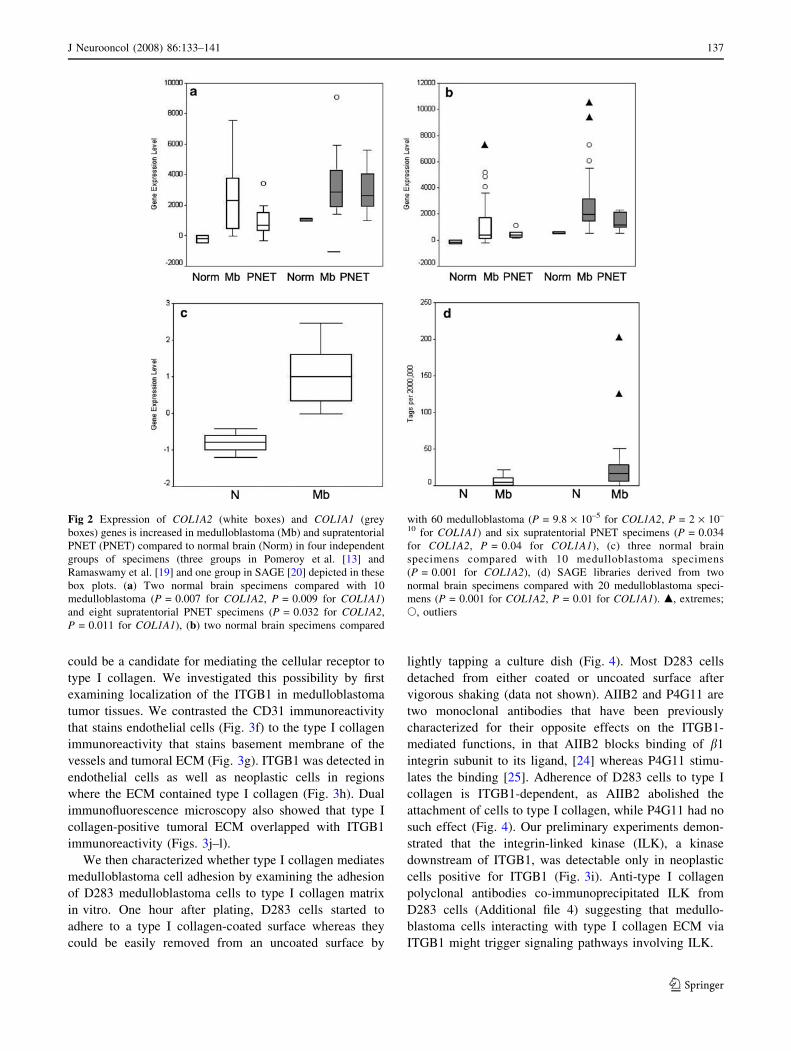
Fig 2 Expression of COL1A2 (white boxes) and COL1A1 (grey
boxes) genes is increased in medulloblastoma (Mb) and supratentorial
PNET (PNET) compared to normal brain (Norm) in four independent
groups of specimens (three groups in Pomeroy et al. [13] and
Ramaswamy et al. [19] and one group in SAGE [20] depicted in these
box plots. (a) Two normal brain specimens compared with 10
medulloblastoma (P = 0.007 for COL1A2, P = 0.009 for COL1A1)
and eight supratentorial PNET specimens (P = 0.032 for COL1A2,
P = 0.011 for COL1A1), (b) two normal brain specimens compared
with 60 medulloblastoma (P = 9.8 · 10–5 for COL1A2, P = 2 · 10–
10 for COL1A1) and six supratentorial PNET specimens (P = 0.034
for COL1A2, P = 0.04 for COL1A1), (c) three normal brain
specimens compared with 10 medulloblastoma specimens
(P = 0.001 for COL1A2), (d) SAGE libraries derived from two
normal brain specimens compared with 20 medulloblastoma speci-
mens (P = 0.001 for COL1A2, P = 0.01 for COL1A1). m, extremes;
s, outliers
J Neurooncol (2008) 86:133–141 137
123

Discussion
Microarrays provide a high throughput platform to char-
acterize gene expression profiles of tumor specimens and to
identify clusters of differentially expressed genes that may
mediate specific biological functions or clinical phenotypes
or mark specific cell lineages from which tumors are de-
rived [26]. For example, we have previously used cDNA
microarrays and sequential supervised and unsupervised
algorithmic analyses to identify a group of genes highly
expressed in rapidly progressing glioblastoma tumors. We
validated the prognostic value of the expression of one of
the genes, FABP7, in two independent sets of specimens
[27]. FABP7 was found to be preferentially expressed in
Fig 3 Type I collagen is overexpressed in medulloblastoma and co-
distributed with ITGB1. In normal brain, expression of type I collagen
is associated with the basement membrane of blood vessels (arrow in
a). Two patterns of type I collagen expression were observed in the
medulloblastoma specimens. The first pattern consisted of type I
collagen immunoreactivity in the basement membrane surrounding
blood vessels (arrow in b), but not in the ECM between neoplastic
cells (* in b). The second pattern of type I collagen immunostaining
was diffusely associated with the ECM of the tumor (* in c, e, g) as
well as the vasculature indicated by arrows in from d to i (endothelial
cells were highlighted by CD31 staining in d and f). Tumor cells
surrounded by type I collagen-positive ECM also expressed ITGB1
(h) and ILK (i). Panels d/e and f–i were consecutive sections from the
same specimen. Dual immunofluorescence microscopy showed that
ITGB1 (j) and type I collagen (k) were co-distributed (yellow color in
l) in the ECM surrounding tumor cells counterstained by DAPI (blue
color in l). Bar in a represents 45 lm for from a to i. Bar in jrepresents 15 lm for from j to l
Fig 4 D283 medulloblastoma cells adhered to type I collagen matrix
via the ITGB1 in vitro. The inhibitory monoclonal antibody, AIIB2,
blocked this interaction, whereas the activating monoclonal antibody,
P4G11, did not. The data were plotted from a representative
experiment with quadruplicates, and * indicates a P value of 0.0017
138 J Neurooncol (2008) 86:133–141
123

glioma cells with astrocytic features, [28] and its prog-
nostic value for glioblastoma was associated with EGFR
overexpression of the tumor cells [29]. Because the avail-
ability of only a limited number of medulloblastoma
specimens might easily confound the reliability of clus-
tering analysis, we utilized a different strategy in this
investigation to identify candidate genes of pathological
importance. We built upon recognition of a frequently
occurring chromosomal aberration in medulloblastoma,
gain of chromosome seven, that is shared by supratentorial
PNET, and sought to identify chromosome seven genes
that were overexpressed in both tumor types using cDNA
microarrays. We validated our finding in published data-
bases that used distinctive assay platforms, and repeatedly
found increased expression of the genes encoding the two
subunits of type I collagen. Immunohistochemistry further
confirmed increased deposition of type I collagen protein in
the ECM of medulloblastoma. These findings provide
evidence for roles of type I collagen in the pathogenesis of
medulloblastoma.
ECM components and the cellular machinery that
mediates the interaction of tumor cells with the ECM are of
great importance in efforts to understand the molecular
mechanisms regulating angiogenesis of malignant cells.
We found high levels of expression of type I collagen in
medulloblastoma specimens associated with tumor vascu-
lature. In normal tissues, type I collagen is a major com-
ponent of basement membranes associated with blood
vessels, and it is required for maintaining the integrity of
the vasculature and for regulating angiogenesis, [30, 31]
but type I collagen is not expressed in normal brain
parenchyma. Type I collagen is believed to directly mod-
ulate the behavior of tumor cells mainly based on data from
studies using halofuginone, a low molecular weight qui-
nazolinone alkaloid that inhibits synthesis of type I colla-
gen. While halofuginone inhibits neovascularization by
inhibiting vascular sprouting, tubular formation, and ECM
deposition by endothelial cells both in vitro and in vivo,
[32] it also inhibits tumor invasion and tumor growth in an
animal model for glioma and chemically induced mouse
bladder carcinoma [33–35].
The best-characterized cellular receptors for type I col-
lagen are a family of integrin membrane proteins. Integrins
are heterodimeric glycoproteins composed of one a chain
and one b chain. All type I collagen-binding integrins share
a common b1 integrin subunit [36]. Anti-a2b1 integrin
antibodies inhibit the attachment of endothelial cells to
type I collagen matrix but not to fibronectin or laminin that
are also the components of vasculature ECM [37]. ITGB1
plays central roles in tumor progression, ECM remodeling,
and angiogenesis by transmitting mechanical signals as it
mediates cell adhesion and migration by interacting with
the cytoskeleton proteins, [38] and it also conveys chemical
signals to control cell survival and proliferation by acti-
vating a spectrum of kinases and downstream adaptor
proteins [38]. ILK is a serine-threonine protein kinase that
binds to the cytoplasmic domain of ITGB1 and regulates
ITGB1-mediated signaling following growth factor stimu-
lation [39]. Identification of ILK-binding proteins has
recently demonstrated that the functions of ILK in ITGB1
signaling pathways include anchoring actin filaments and
activating signaling molecules downstream of growth fac-
tor receptors [40].
Type 1 collagen may be of importance in the pathobi-
ology of medulloblastoma because of its role in blood
vessel formation. Medulloblastoma is a highly vascular
tumor and several studies have suggested that anti-angio-
genesis-directed therapy may be a potential strategy for
future approaches to the treatment of medulloblastoma. For
example, an integrin av antagonist peptide successfully
inhibited angiogenesis induced by human DAOY medul-
loblastoma cells growing in chicken chorioallantoic mem-
brane [41] and also reduced growth of DAOY cells
implanted into nude mice [42]. Whether inhibitors of type I
collagen, such as halofuginone, can suppress the spread and
growth of medulloblastoma cells via inhibition of angio-
genesis or by a direct effect on neoplastic cells deserves
further investigation. Type I collagen is also a major
constituent of leptomeninges, [43] and invasion of the
leptomeninges is a frequent route over which medullo-
blastoma cells spread throughout the neuraxis and establish
systemic metastatic disease [44]. However, expression of
type I collagen might not be involved in medulloblastoma
invasion into adjacent brain structures or metastasis through
cerebrospinal fluid, as analyses using the published dataset
did not show correlation between type I collagen expression
and tumor/metastasis stages (data not shown). This is con-
sistent with the findings that the vascularity of medullo-
blastoma is not associated with either metastasis or patient
survival [12, 45].
We demonstrated that type I collagen is also present in
the ECM associated with medulloblastoma cells in vivo.
The type I collagen-positive interstitial matrix may be
essential for the angiogenic behavior of medulloblastoma
cells. Co-distribution of ITGB1 with type I collagen also
suggests the possibility that by binding to its ligand, ITGB1
triggers intracellular signaling pathways that might regu-
late the pathophysiology of medulloblastoma cells, proba-
bly through ILK. In one study, ILK expression was not
seen in normal brain but was detected in all three medul-
loblastoma and four supratentorial PNET specimens
examined [46]. We had preliminary data suggesting that
ILK might be recruited to the type I collagen/ITGB1
complex in D283 cells. These findings support the notion
that the type I collagen/ITGB1 interaction in medullo-
blastoma may activate a signaling cascade via ILK. It
J Neurooncol (2008) 86:133–141 139
123

would be interesting to investigate whether interaction of
type I collagen and ITGB1 relates to angiogenic activities
of medulloblastoma cells.
Our findings suggest the possibility of a role for the
inhibition of type I collagen and its signaling pathways in
the treatment of medulloblastoma. The therapeutic poten-
tial of inhibiting type I collagen synthesis or downstream
signaling events in medulloblastoma might have a two-
prong effect: inhibition of angiogenesis and the tumoral
vasculature in which type I collagen is actively synthe-
sized, and blocking type I collagen-mediated tumor cell
interaction by reducing the activated integrin signaling.
Such therapeutic activities seem very likely to affect the
pathobiology of this tumor in a manner that would have a
significant impact on the clinical behavior of the tumor and
thereby contribute to an improved outcome for patients.
Acknowledgements We thank the Brain Tumor Research Center
Tissue Bank of UCSF for contributing tissue specimens in this study.
This work was supported by funding from the Department of Neu-
rological Surgery at UCSF, and by National Institute of General
Medical Sciences training grant GM07365 (M.D.), and the Theodora
B. Betz Foundation and Kyra Memorial Fund (M.A.I.). UCSF is an
NCI-designated Specialized Program of Research Excellence for
Brain Tumors.
References
1. Jemal A, Murray T, Samuels A (2003) Cancer Statistics. CA
Cancer J Clin 53:5–26
2. Rutkowski S (2006) Current treatment approaches to early child-
hood medulloblastoma. Expert Rev Neurother 6(8):1211–1221
3. Evans AE, Jenkin RD, Sposto R et al (1990) The treatment of
medulloblastoma. Results of a prospective randomized trial of
radiation therapy with and without CCNU, vincristine, and
prednisone. J Neurosurg 72(4):572–582
4. Packer RJ, Sutton LN, Elterman R et al (1994) Outcome for
children with medulloblastoma treated with radiation and cis-
platin, CCNU, and vincristine chemotherapy. J Neurosurg
81(5):690–698
5. Zeltzer PM, Boyett JM, Finlay JL et al (1999) Metastasis stage,
adjuvant treatment, and residual tumor are prognostic factors for
medulloblastoma in children: conclusions from the Children’s
Cancer Group 921 randomized phase III study. J Clin Oncol
17(3):832–845
6. Marino S (2005) Medulloblastoma: developmental mechanisms
out of control. Trends Mol Med 11(1):17–22
7. MacDonald TJ, Brown KM, LaFleur B et al (2001) Expression
profiling of medulloblastoma: PDGFRA and the RAS/MAPK
pathway as therapeutic targets for metastatic disease. Nat Genet
29(2):143–152
8. Dirks PB, Harris L, Hoffman HJ et al (1996) Supratentorial
primitive neuroectodermal tumors in children. J Neurooncol
29(1):75–84
9. Albright AL, Wisoff JH, Zeltzer P et al (1995) Prognostic factors
in children with supratentorial (nonpineal) primitive neuroecto-
dermal tumors. A neurosurgical perspective from the Children’s
Cancer Group. Pediatr Neurosurg 22(1):1–7
10. Cohen BH, Zeltzer PM, Boyett JM et al (1995) Prognostic factors
and treatment results for supratentorial primitive neuroectodermal
tumors in children using radiation and chemotherapy: a Childrens
Cancer Group randomized trial. J Clin Oncol 13(7):1687–1696
11. Huber H, Eggert A, Janss AJ et al (2001) Angiogenic profile of
childhood primitive neuroectodermal brain tumours/medullo-
blastomas. Eur J Cancer 37(16):2064–2072
12. Grotzer MA, Wiewrodt R, Janss AJ et al (2001) High micro-
vessel density in primitive neuroectodermal brain tumors of
childhood. Neuropediatrics 32(2):75–79
13. Pomeroy SL, Tamayo P, Gaasenbeek M et al (2002) Prediction of
central nervous system embryonal tumour outcome based on gene
expression. Nature 415(6870):436–442
14. Russo C, Pellarin M, Tingby O et al (1999) Comparative geno-
mic hybridization in patients with supratentorial and infratento-
rial primitive neuroectodermal tumors. Cancer 86(2):331–339
15. Burnett ME, White EC, Sih S, von Haken MS, Cogen PH (1997)
Chromosome arm 17p deletion analysis reveals molecular genetic
heterogeneity in supratentorial and infratentorial primitive neu-
roectodermal tumors of the central nervous system. Cancer Genet
Cytogenet 97(1):25–31
16. McCabe MG, Ichimura K, Liu L et al (2006) High-resolution
array-based comparative genomic hybridization of medulloblas-
tomas and supratentorial primitive neuroectodermal tumors. J
Neuropathol Exp Neurol 65(6):549–561
17. Bayani J, Zielenska M, Marrano P et al (2000) Molecular cyto-
genetic analysis of medulloblastomas and supratentorial primitive
neuroectodermal tumors by using conventional banding, com-
parative genomic hybridization, and spectral karyotyping. J
Neurosurg 93(3):437–448
18. Reardon DA, Michalkiewicz E, Boyett JM et al (1997) Extensive
genomic abnormalities in childhood medulloblastoma by com-
parative genomic hybridization. Cancer Res 57(18):4042–4047
19. Ramaswamy S, Tamayo P, Rifkin R et al (2001) Multiclass
cancer diagnosis using tumor gene expression signatures. Proc
Natl Acad Sci U S A 98(26):15149–15154
20. Boon K, Osorio EC, Greenhut SF et al (2002) An anatomy of
normal and malignant gene expression. Proc Natl Acad Sci U S A
99(17):11287–11292
21. Velculescu VE, Zhang L, Vogelstein B, Kinzler KW (1995)
Serial analysis of gene expression. Science 270(5235):484–487
22. Karsenty G, Park RW (1995) Regulation of type I collagen genes
expression. Int Rev Immunol 12(2–4):177–185
23. Karsenty G, de Crombrugghe B (1991) Conservation of binding
sites for regulatory factors in the coordinately expressed alpha 1
(I) and alpha 2 (I) collagen promoters. Biochem Biophys Res
Commun 177(1):538–544
24. Werb Z, Tremble PM, Behrendtsen O, Crowley E, Damsky CH
(1989) Signal transduction through the fibronectin receptor in-
duces collagenase and stromelysin gene expression. J Cell Biol
109(2):877–889
25. Wayner EA, Gil SG, Murphy GF, Wilke MS, Carter WG (1993)
Epiligrin, a component of epithelial basement membranes, is an
adhesive ligand for alpha 3 beta 1 positive T lymphocytes. J Cell
Biol 121(5):1141–1152
26. Quackenbush J (2006) Microarray analysis and tumor classifi-
cation. N Engl J Med 354(23):2463–2472
27. Liang Y, Diehn M, Watson N et al (2005) Gene expression
profiling reveals clinically distinct subtypes of glioblastoma
multiforme. Proc Natl Acad Sci U S A 102(16):5814–5819
28. Liang Y, Bollen AW, Nicholas MK, Gupta N (2005) Id4 and
FABP7 are preferentially expressed in cells with astrocytic fea-
tures in oligodendrogliomas and oligoastrocytomas. BMC Clin
Pathol 5:6
29. Liang Y, Bollen AW, Aldape KD, Gupta N (2006) Nuclear
FABP7 immunoreactivity is preferentially expressed in infiltra-
tive glioma and is associated with poor prognosis in EGFR-
overexpressing glioblastoma. BMC Cancer 6:97
140 J Neurooncol (2008) 86:133–141
123

30. Montesano R, Orci L, Vassalli P (1983) In vitro rapid organiza-
tion of endothelial cells into capillary-like networks is promoted
by collagen matrices. J Cell Biol 97(5 Pt 1):1648–1652
31. Jackson CJ, Jenkins KL (1991) Type I collagen fibrils promote
rapid vascular tube formation upon contact with the apical side of
cultured endothelium. Exp Cell Res 192(1):319–323
32. Elkin M, Miao HQ, Nagler A et al (2000) Halofuginone: a potent
inhibitor of critical steps in angiogenesis progression. Faseb J
14(15):2477–2485
33. Abramovitch R, Dafni H, Neeman M, Nagler A, Pines M (1999)
Inhibition of neovascularization and tumor growth, and facilita-
tion of wound repair, by halofuginone, an inhibitor of collagen
type I synthesis. Neoplasia 1(4):321–329
34. Elkin M, Ariel I, Miao HQ et al (1999) Inhibition of bladder
carcinoma angiogenesis, stromal support, and tumor growth by
halofuginone. Cancer Res 59(16):4111–4118
35. Gross DJ, Reibstein I, Weiss L et al (2003) Treatment with
halofuginone results in marked growth inhibition of a von Hippel-
Lindau pheochromocytoma in vivo. Clin Cancer Res 9(10 Pt
1):3788–3793
36. White DJ, Puranen S, Johnson MS, Heino J (2004) The collagen
receptor subfamily of the integrins. Int J Biochem Cell Biol
36(8):1405–1410
37. Gamble JR, Matthias LJ, Meyer G et al (1993) Regulation of
in vitro capillary tube formation by anti-integrin antibodies. J
Cell Biol 121(4):931–943
38. Guo W, Giancotti FG (2004) Integrin signalling during tumour
progression. Nat Rev Mol Cell Biol 5(10):816–826
39. Hannigan GE, Leung-Hagesteijn C, Fitz-Gibbon L et al (1996)
Regulation of cell adhesion and anchorage-dependent growth by
a new beta 1-integrin-linked protein kinase. Nature
379(6560):91–96
40. Wu C, Dedhar S (2001) Integrin-linked kinase (ILK) and its in-
teractors: a new paradigm for the coupling of extracellular matrix
to actin cytoskeleton and signaling complexes. J Cell Biol
155(4):505–510
41. MacDonald TJ, Taga T, Shimada H et al (2001) Preferential
susceptibility of brain tumors to the antiangiogenic effects of an
alpha(v) integrin antagonist. Neurosurgery 48(1):151–157
42. Taga T, Suzuki A, Gonzalez-Gomez I et al (2002) alpha v-Inte-
grin antagonist EMD 121974 induces apoptosis in brain tumor
cells growing on vitronectin and tenascin. Int J Cancer
98(5):690–697
43. Rutka JT, Giblin J, Dougherty DV et al (1986) An ultrastructural
and immunocytochemical analysis of leptomeningeal and
meningioma cultures. J Neuropathol Exp Neurol 45(3):285–303
44. Rutka JT (1997) Medulloblastoma. Clin Neurosurg 44:571–585
45. Ozer E, Sarialioglu F, Cetingoz R et al (2004) Prognostic sig-
nificance of anaplasia and angiogenesis in childhood medullo-
blastoma: a pediatric oncology group study. Pathol Res Pract
200(7–8):501–509
46. Chung DH, Lee JI, Kook MC et al (1998) ILK (beta1-integrin-
linked protein kinase): a novel immunohistochemical marker for
Ewing’s sarcoma and primitive neuroectodermal tumour. Virch-
ows Arch 433(2):113–117
J Neurooncol (2008) 86:133–141 141
123




![Medulloblastoma: [Print] - eMedicine Neurology · emedicine.medscape.com eMedicine Specialties > Neurology > Pediatric Neurology Medulloblastoma George I Jallo, MD, Associate Professor](https://img.pdfslide.us/doc/110x75/5d472c3c88c993527c8b60e5/medulloblastoma-print-emedicine-neurology-emedicinemedscapecom-emedicine.jpg)
























