Embed Size (px)
Citation preview

1
Towards the Rational Design of a Malaria Vaccine Construct; Example of 1
the MSP3 Family, Part II: Antigenicity Studies 2
3
Corine G. Demanga1, Lena-Juliette Daher
1,2, Eric Prieur
1, Catherine Blanc
1, Jean-Louis 4
Pérignon1, Hasnaa Bouharoun-Tayoun
2, Pierre Druilhe
1 5
6
Biomedical Parasitology Unit, Institut Pasteur1, Paris, France; Laboratory of Immunology, 7
Faculty of Public Health, Lebanese University2, Fanar, Lebanon 8
9
10
Running Title: From MSP3 antigens family to tailored malaria vaccine 11
12
13
Corresponding author
14
Pierre Druilhe 15
Biomedical Parasitology Unit, Pasteur Institute, 16
25 rue du Dr Roux, 75015 Paris, France. 17
Phone: +33145688578 18
Fax: +33145688640 19
Mail: [email protected] 20
21
22
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.01359-08 IAI Accepts, published online ahead of print on 2 November 2009
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

2
ABSTRACT 1
Plasmodium falciparum MSP3 is a main target of protective immunity against malaria, 2
currently undergoing vaccine development. It was shown recently to belong, together with 3
MSP6, to a new multigene family, which C-terminal regions have a similar organisation, 4
contain both homologous and divergent regions, and are highly conserved across isolates. In 5
an attempt to rationally design novel vaccine constructs, we extended the analysis of 6
antigenicity and function of region-specific antibodies, previously performed with MSP3 and 7
MSP6, to the remaining 4 proteins of the MSP3 family using 4 recombinant proteins and 24 8
synthetic peptides. Antibodies to each MSP3 family antigen were found highly prevalent 9
among malaria exposed individuals from the village of Dielmo (Senegal). Each of the 24 10
peptides was antigenic, defining at least one epitope mimicking that of the native proteins, 11
with a distinct IgG isotype pattern for each, though with an overall predominance of the IgG3 12
subclass. Human antibodies affinity-purified upon each of the 24 peptides exerted an anti-13
parasite ADCI effect, which in most cases was as strong as that of IgG from protected African 14
adults. The two regions with high homology were found to generate a broad network of cross-15
reactive antibodies with various avidities. A first multigenic construct was designed using the 16
above findings and those from related immunogenicity studies in mice, and was shown to 17
have valuable immunological properties. These results indicate that numerous regions from 18
the MSP3 family play a role in protection and provide a rationale for the tailoring of new 19
MSP3 derived malaria vaccines. 20
21
Key words: P.falciparum malaria, Multi-subunit vaccine, Vaccine tailoring, MSP3, Antigens 22
family, ADCI. 23
24
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

3
Abbreviations list: MSP, Merozoite Surface Protein; LSP, Long Synthetic Peptide; ADCI, 1
Antibody dependent cellular inhibition; PIAG, Pool of Immune African globulins. 2
3
INTRODUCTION. 4
Malaria vaccine development is both urgently needed and remarkably challenging in 5
scientific terms, particularly in the choice of concepts determining which path(s) should be 6
followed to achieve this end. The criteria employed in the decision-making process affect 7
obviously the choice of a candidate among the 5300 Plasmodium falciparum proteins, i.e. the 8
vaccine discovery process, but they also affect markedly the type of construct, expression 9
system or formulation chosen, as well as the selection of candidates to be included in vaccine 10
combinations (18). 11
Given the uncertainties related to experimental animal models, particularly their 12
relevance to human malaria (10), we have made the choice to base our research orientations 13
on clinical observations, as we trust they are the most reliable. According to this strategy, 14
animal models come into play only in a second step, mainly for detailed immunogenicity 15
studies that obviously cannot be conducted initially in humans. For vaccine discovery, 16
immunity was passively transferred into P.falciparum infected recipients (3, 31) by human 17
IgG from protected African adults. This led us to compare non-protective with protective 18
antibodies upon each parasite isolate, and to identify a novel defence mechanism which had 19
not been foreseen by research in models, in which the effective antibodies act in a monocyte 20
dependent manner. This antibody dependent cellular inhibition (ADCI) of parasite 21
multiplication (3, 5) was in turn employed to identifiy merozoite surface protein 3 (MSP3) as 22
a main target of protective antibodies (25). Its C-terminal region was found to be fully 23
conserved across P.falciparum isolates, in contrast to most other vaccine candidates (30). 24
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

4
Studies in exposed populations show that MSP3 induces antibody responses that are 1
strongly associated with clinical protection against disease (28, 30, 36). A detailed antigenic 2
analysis of the C-terminal portion of MSP3 highlighted the importance of 3 antigenic 3
determinants targeted by cytophilic antibodies induced by natural infection and able to inhibit 4
parasite growth in functional in vitro ADCI assays (32). This information was instrumental in 5
the design of the MSP3-LSP (long synthetic peptide) construct that was taken into the clinics 6
(1) and which induced in human volunteers antibodies that inhibited P.falciparum 7
multiplication both in vitro and in vivo (12). 8
We have recently reported that msp3 belongs to a multigene family, differing in its 9
features from other P. falciparum gene families (34). Indeed, the six msp3 genes are 10
transcribed simultaneously, the corresponding proteins are expressed in all isolates 11
investigated, at late schizont stage. Their C-terminal portions share a similar organization and 12
include regions with sequence homologies flanked by regions that markedly differ from one 13
protein to the other, yet are very well conserved among isolates. The original msp3 was 14
renamed as msp3.1, msp6 (41) as msp3.2, and the remaining msp3.3, msp3.4, msp3.7 and 15
msp3.8 (SI-figure 1). In view of the unique features of this MSP3 family, particularly the high 16
degree of conservation of homologous and divergent sequences implying that they had a role 17
for parasite survival, we thought it valuable to develop a polyantigenic construct combining 18
carefully chosen regions among the most immunologically relevant from these six proteins. 19
A detailed immunological study combining immunogenicity and antigenicity data 20
provides a means to design malaria vaccines based on the new knowledge resulting from this 21
analysis, where immunogenicity and antigenicity studies are complementary. Indeed, the 22
former analyse the responses induced by a given vaccine formulation in experimental models, 23
however have the great limitation to do so in animals whose relevance to humans is not 24
granted (10). In contrast the latter have the paramount advantage to identify immune 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

5
responses developed by the target population, human beings, although these responses have 1
been induced by exposure to the whole parasite. However, our first generation MSP3-LSP 2
vaccine readily induced in naïve European volunteers an immune response having the same 3
characteristics (cytophilicity, titre, fine epitope specificity) as the immune response observed 4
in exposed populations having acquired protection (10). 5
In the present detailed antigenicity study we characterized the antibody responses 6
from humans naturally exposed to malaria directed to recombinant antigens and to a series of 7
overlapping peptides spanning the C-terminal portions of MSP3.3, MSP3.4, MSP3.7 and 8
MSP3.8, and assessed the biological anti-parasite activity of the corresponding antibodies,. 9
The data obtained were compared to the results previously obtained with MSP3.1 and 10
MSP3.2 (32, 33). The antigenicity data of all six members of the MSP3 family combined with 11
the detailed study of the immunogenicity of the C-terminal portion of MSP3.1 (7), led to the 12
design of the first multi-antigenic vaccine construct based on the MSP3 family of proteins, 13
which was shown to have valuable immunological features. 14
15
16
17
18
19
20
21
22
23
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

6
MATERIAL AND METHODS. 1
Recombinant antigens 2
The current Gene IDs in the database PlasmoDB release 5.5 (www.plasmodb.org) of 3
the P. falciparum MSP3 family proteins are: PF10_0345, PF10_0346, PF10_0347, 4
PF10_0348, PF10_0352 and PF10_0355 for MSP3.1, MSP3.2, MSP3.3, MSP3.4, MSP3.7 5
and MSP3.8, respectively. The set of six recombinant hexahistidine-tagged proteins that were 6
designed to cover the related C-terminal regions of these MSP3 proteins are: MSP3.1191-354, 7
MSP3.2161-371, MSP3.3228-424, MSP3.4508-697, MSP3.7214-405 and MSP3.8537-762 (34). The overall 8
percent homology of the MSP3 family proteins were analysed with EMBOSS pairwise 9
sequence alignments (www.ebi.ac.uk/Tools/emboss/align/), 10
11
Peptide design 12
Multiple sequence alignments for the MSP3 antigens were generated through 13
ClustalW2 (www.ebi.ac.uk/Tools/clustalw2) and DIALIGN2 (mobyle.pasteur.fr). The scheme 14
of alignments and further manual refinements were used to direct the design of six 15
overlapping peptides (SI-Fig 2), from 21-61 amino acids in length, covering the C-terminal 16
portion (excluding the glutamic acid rich region) for each of the four MSP3 proteins MSP3.3, 17
MSP3.4, MSP3.7 and MSP3.8. They were designed in order to fit those previously 18
synthesized for MSP3.1 and MSP3.2 (32), (33), and were named “a,b,c,d,e, f”, as was the 19
case for the latter. The position and the sequence of the peptides are detailed in 20
Supplementary Information (Table 1 and figure 2). Peptide synthesis was carried out through 21
standard stepwise solid phase peptide synthesis (GenScript Corp., Piscataway, NJ). 22
23
24
25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

7
ELISA assays 1
The immunoepidemiological studies aimed at determining the prevalences and IgG 2
subclass distributions were performed using serum samples from the inhabitants of Dielmo, a 3
village located in an area of Senegal, West Africa, where malaria is holoendemic (40). These 4
samples were collected during a cohort prospective follow-up study designed to identify and 5
analyse all malaria-associated episodes of morbidity (40). The detection of total IgG and IgG 6
subclasses was performed by ELISA as described previously (4, 9), using secondary 7
monoclonal antibodies originally selected as reacting with African subclass dominant 8
allotypes. Mouse monoclonal antibodies specific for human IgG subclasses (clones NL16 = 9
IgG1 [Boehringer Mannheim, Meylan, France], HP6002 = IgG2 [Sigma-Aldrich, Steinheim, 10
Germany], Zg4 = IgG3, and RJ4 = IgG4 [both from Immunotech, Marseille, France] were 11
used at a final dilution of 1/2,000; 1/10,000; 1/10,000; and 1/1,000, respectively). For the total 12
IgG detection, the horseradish peroxidase-conjugated goat anti-human (Caltag-Invitrogen, 13
Carlsbad, CA) was used at a 1/3,000 dilution. For each antigen (peptides or recombinant 14
proteins), the results were expressed as OD ratios, or Arbitrary Units, calculated by dividing 15
the optical density (OD) of each serum sample over the mean OD + 3 standard deviations 16
(SD) of 5 control serum samples (30). These control sera were randomly selected among 17
French blood donors without exposure to malaria and their antibody reactivity was assumed 18
to be normally distributed. Therefore the mean OD + 3xSD covered all the negative controls 19
responses. 20
21
Affinity Purification of Specific Antibodies. 22
The antibodies specific for the 24 overlapping peptides covering the C-terminal 23
sequence of MSP3.3, MSP3.4, MSP3.7 and MSP3.8 were affinity-purified from a pool of sera 24
derived from 333 rural African adults from Ivory Coast, West Africa, who had reached a state 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

8
of premunition, i.e. a pool similar to that employed to prepare the IgG used in the passive 1
transfer experiment (31). 2
Antibodies specific to either peptides or recombinant antigens were prepared as 3
described previously (6, 32). Briefly, plain polystyrene beads (mean diameter, 10 m; 4
Polysciences, Eppelheim, Germany) were coated with antigens, in a 5% (w/v) PBS 5
suspension, and washed thoroughly in PBS. The coated beads were incubated for 2 hours at 6
room temperature with the pool of African sera mentioned above diluted 1/10, then washed 7
twice in PBS-0.01% Tween-20. The antibodies captured on the beads were eluted using a 8
Glycine buffer 0.2M, pH 2.5, and immediately neutralised with the required volume of Tris 9
buffer 2M, pH 11.2. The antibodies eluted were dialysed extensively against PBS, followed 10
by RPMI, and then concentrated by centrifugation using the molecular sieve Vivaspin2 11
(30,000 MWCO PES, Sartorius Stedim, Göttingen, Germany). They were stored at 4°C after 12
addition of 1% AlbumaxI (Gibco Invitrogen, Carlsbad, CA). The specificity of affinity-13
purified antibodies was ascertained by ELISA on each peptide, using bovine serum albumin 14
and the peptide SALSA-1 from the Sporozoite and Liver Stage Antigen-1 (2), as negative 15
controls. The purity of antibodies preparation was ascertained in SDS-PAGE using a 10% 16
polyacrylamide gel. 17
Indirect Immunofluorescence assays (IFA) were performed by previously described 18
techniques (3) in order to assess recognition of the native parasite proteins by affinity-purified 19
antibodies. Air-dried acetone-fixed thin smears from the 3D7 P. falciparum clone at mature 20
schizont stage were incubated with serial dilutions of affinity-purified antibodies, and using at 21
a 1/200 dilution a fluorescein labelled Goat anti-human IgG (Alexa Fluor 488, Invitrogen, 22
Carlsbad, CA) as second antibody. 23
24
25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

9
Functional ADCI assays. 1
The P. falciparum 3D7 clone (MRA-102, MR4, ATCC, Manassas, VA) was cultivated 2
in complete medium, i.e. RPMI-1640 supplemented with 0.5% AlbumaxI (both from Gibco 3
Invitrogen, Carlsbad, CA), as described earlier (21). Schizonts enrichment by floatation on 4
1% (w/v) gelatin (Sigma-Aldrich, Steinheim, Germany) (21) was performed 48 hours before 5
initiating the ADCI-based functional assays. 6
IgG was extracted from the same pool of sera derived from Ivory Coast adults used for 7
affinity purification, by ion exchange chromatography on DEAE Ceramic HyperD F (Pall 8
BioSepra, Cergy-Saint-Christophe, France) and named PIAG (Pool of Immune African 9
Globulins). In the ADCI assays performed to evaluate the biological activity of antibodies 10
specific to each antigen, two positive controls were used: the PIAG, and antibodies affinity-11
purified on MSP3.1-Cterm. The negative control was IgG from a pool of French donors with 12
no malaria history (N-IgG). Peripheral blood mononuclear cells (PBMC) from healthy French 13
donors were prepared and cryopreserved as previously described (20). For the ADCI assay, 14
PBMC were rapidly thawed at 37°C and assessed for viability by trypan blue exclusion. Cell 15
suspensions were adjusted to a concentration of 10x106 cells per ml with RPMI-1640, and 16
200 l/well distributed in flat bottom 96-well plates (TPP, Trasadingen, Switzerland) were 17
incubated for 1 hour at 37°C, 5% CO2. The non-adherent cells were removed by two washing 18
steps with RPMI-1640, then the cell appearance and the relative homogeneity of distribution 19
of the adherent cells in the different wells, were controlled by observation under an inverted 20
microscope. Antibodies affinity purified upon the different peptides or recombinant antigens 21
were tested at a final concentration yielding a 1/20 IFA titre, the N-IgG and PIAG were used 22
at 2mg/ml (about 10% the IgG concentration in African adults sera). To each well were added 23
50 l of the test antibody and 50 l of a schizont infected red blood cells suspension at a final 24
hematocrit of 2.5% and 0.5% parasitemia, both in complete medium. The control wells 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

10
consisted of either monocytes alone, or monocytes with N-IgG, or parasites with antibodies 1
but without monocytes or parasites alone (8, 25). At 48 and 72h, 50#l of complete medium 2
were added to each well. Following cultivation for 96 h, the level of parasitemia was 3
determined on Giemsa-stained thin smears from each well by microscopic examination of at 4
least 50,000 erythrocytes. The Specific Growth Inhibitory index (SGI), which takes into 5
account the possible inhibition induced by monocytes or antibodies alone, was calculated as 6
follows: SGI= 100x[1-(parasitemia with monocytes and test IgG/parasitemia with test 7
IgG)/(parasitemia with monocytes and N-IgG/parasitemia with N-IgG)], where parasitemias 8
are expressed as percentage of infected red blood cells. The ADCI assays were performed in 9
duplicate for each affinity-purified antibody, using monocytes showing less than 10% direct 10
non-specific inhibition of parasite growth. Based on previous experience, the threshold of 11
positivity of the ADCI assay was considered to be 30% or greater of the positive control (12). 12
13
Cross-reactivity studies. 14
In a first set of experiments, the cross-reactivity of antibodies was assessed towards 15
the sequence of the peptides “b” which are quasi-identical for MSP3.1, MSP3.2 and MSP3.7. 16
The competition ELISA was performed as described elsewhere (14), with competing antigen 17
at concentrations ranging from 0 to 800#g/ml. The results expressed as percentage reactivity 18
were calculated as follows: [(mean O.D values with the competing antigen at a given 19
concentration)/ (mean O.D values without the competing antigen)] 100. To control the 20
specificity of the competition between antigen and antibody, the non-relevant antigen 21
SALSA-1 was added at the same concentration as the competing antigen. 22
The second set of experiments, dedicated to characterise the overall cross-reactivity 23
network among the MSP3 family, were based on the reactivity of antibodies specific to the 24
peptide “d”. Indeed the sequences of the peptides “d” are highly homologous among the 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

11
MSP3 family. The ELISA reactivity of antibodies specific to each peptide “d” upon all the six 1
recombinant MSP3-Cterm antigens was determined. 2
3
Avidity studies. 4
The binding strength of antibodies induced in mice by a first MSP3 polyantigenic 5
construct (7), was evaluated by ELISA towards each MSP3 recombinant C-terminus protein 6
as described elsewhere (29). Briefly, antibodies affinity purified upon the polyantigenic 7
construct from a pool of sera from immunized mice were tested on microplates coated with 8
each of the six MSP3 recombinant proteins. After allowing the formation of antigen-antibody 9
complexes, the latter were dissociated with increasing concentrations of the chaotropic ion 10
ammonium thiocyanate (NH4SCN), from 0.25 to 4 molar. After a wash step the remaining 11
antibodies were quantified. The percentage reactivity was calculated as follows: [(mean O.D 12
values with a given concentration of NH4SCN)/ (mean O.D values without NH4SCN)] 100. 13
The avidity index was estimated as the molar concentration of NH4SCN required to reduce 14
the initial optical density by 50%. 15
16
17
18
19
20
21
22
23
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

12
RESULTS 1
Antibodies specific for the six MSP3 antigens are highly prevalent in subjects from malaria 2
endemic area. 3
IgG antibodies specific for all the six MSP3 antigens were detected in serum samples 4
from adults living in the village of Dielmo, Senegal. Prevalences, i.e. percentages of 5
individuals with a positive response, were very high, ranging from 89% to 95%, even though 6
IgG levels varied from one antigen to the other (Table 1). The high antibody reactivity 7
obtained with MSP3.2 was similar to that obtained with MSP3.4 or with MSP3.1. The lowest 8
one was with MSP3.7, which was in the same range as that of MSP3.8 and MSP3.3. Given 9
the role of cytophilic antibodies in parasite killing and their association with protection 10
against malaria, we performed IgG cytophilic subclass analysis (IgG1 and IgG3) of responses 11
towards the six MSP3 antigens using sera from a cohort of 76 malaria exposed individuals of 12
all ages (0.9 -80.4 years) from the same village (figure 1). The results confirm the high 13
prevalence of responses, since for each antigen, except MSP3.8, the prevalences were 80% 14
for IgG1 or IgG3. While IgG3 antibodies were dominant in the case of MSP3.2, MSP3.4, and 15
MSP3.7 the responses to the three remaining antigens were balanced between IgG1 and IgG3. 16
17
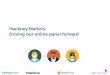
The C-terminal regions of the six MSP3 antigens contain highly homologous sequences 18
flanked by non-homologous ones. 19
In addition to their overall structural homology, the proteins of the MSP3 family share 20
sequence similarities in their C-terminal portions. The overall percent homology, which is 21
between 32-49% for the full-length protein, increases to 43- 57% for the C-terminal portions 22
(figure 2-A). The first conserved motif, made of eleven amino acid residues centred on three 23
glycine residues (figure 2-B box I), is included in the peptides called “b”. The sequences 24
upstream and downstream are unique for each antigen, which directed the design of 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

13
“peptides a” and “peptides c”, respectively (SI-table 1). Noticeably, the MSP3.1a sequence 1
has the unique feature of heptad repeats. For each MSP3 antigen the “peptide d” located 2
upstream of the glutamic acid rich region is characterised by a stretch of 20 amino acids with 3
high position specific scoring i.e. identical or homologous residues being conserved for a 4
given position (figure2-B, box II). The “peptides e" are located downstream of the Glu-rich 5
region and finally, the homologous leucine-zipper like pattern is displayed in the “peptides f” 6
(figure2-B box III). 7
8
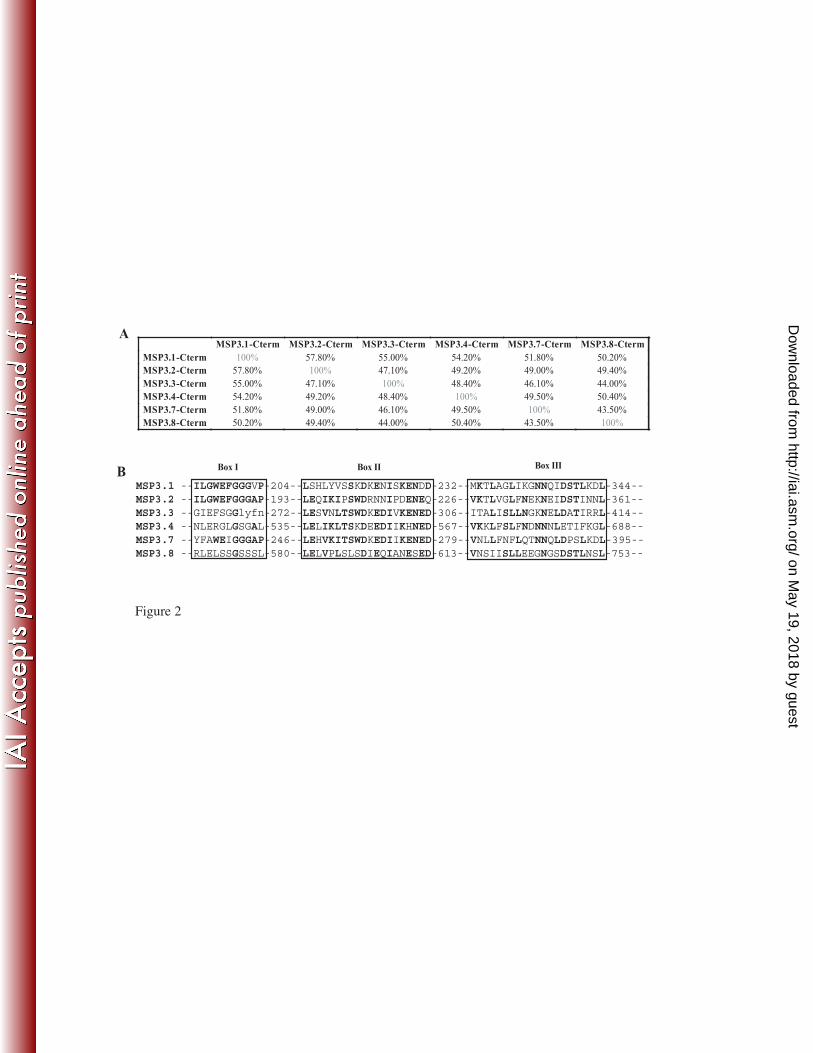
The 24 peptides from MSP3.3, MSP3.4, MSP3.7 and MSP3.8 are all antigenic and 9
predominantly targeted by cytophilic antibodies. 10
The detailed analysis of the IgG subclass distribution among a cohort of malaria 11
exposed individuals of all age groups, against the twenty-four peptides covering the C-12
terminal regions of these four antigens, showed that each of them was recognized by 13
antibodies from endemic area sera, i.e. all were antigenic. The prevalence of specific 14
antibodies was high; indeed more than 60% of the individuals had antibodies reacting with 15
two thirds of the peptides (figure 3). For all of them the antibodies were predominantly of 16
IgG1 and IgG3 subclasses, and only one (MSP3.8b) was targeted by a non-cytophilic IgG2 17
response. In most cases, the IgG1 and IgG3 levels were of similar magnitude, but a trend, 18
although non-significant, towards a predominance of IgG3 was observed against some 19
peptides (MSP3.3a, MSP3.4b and MSP3.7e), or of IgG1 for others (e.g. MSP3.3b and 20
MSP3.7d) (figure 3). 21
The present results together with those previously reported for MSP3.1 and MSP3.2 22
(32, 33), show that the subclass patterns of responses to the homologous peptides b, d or f 23
from the six antigens of the MSP3 family differ markedly. This suggests that the flanking 24
regions, defining T-helper cell epitopes (31), may play an important role in the modulation of 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

14
the IgG1 to IgG3 balance. Additional differences in antigenicty emerge for the various 1
peptides “a”: in contrast to the low prevalence and low levels of antibodies previously 2
observed to peptide MSP3-1a, high prevalences and levels are observed for MSP3-3a and 3
MSP3-8a. Conversely responses to the various peptides “b”, which is a major target of ADCI 4
for MSP3-1, are high in 5 out of 6 proteins (i.e. all except MSP3-7). 5
6
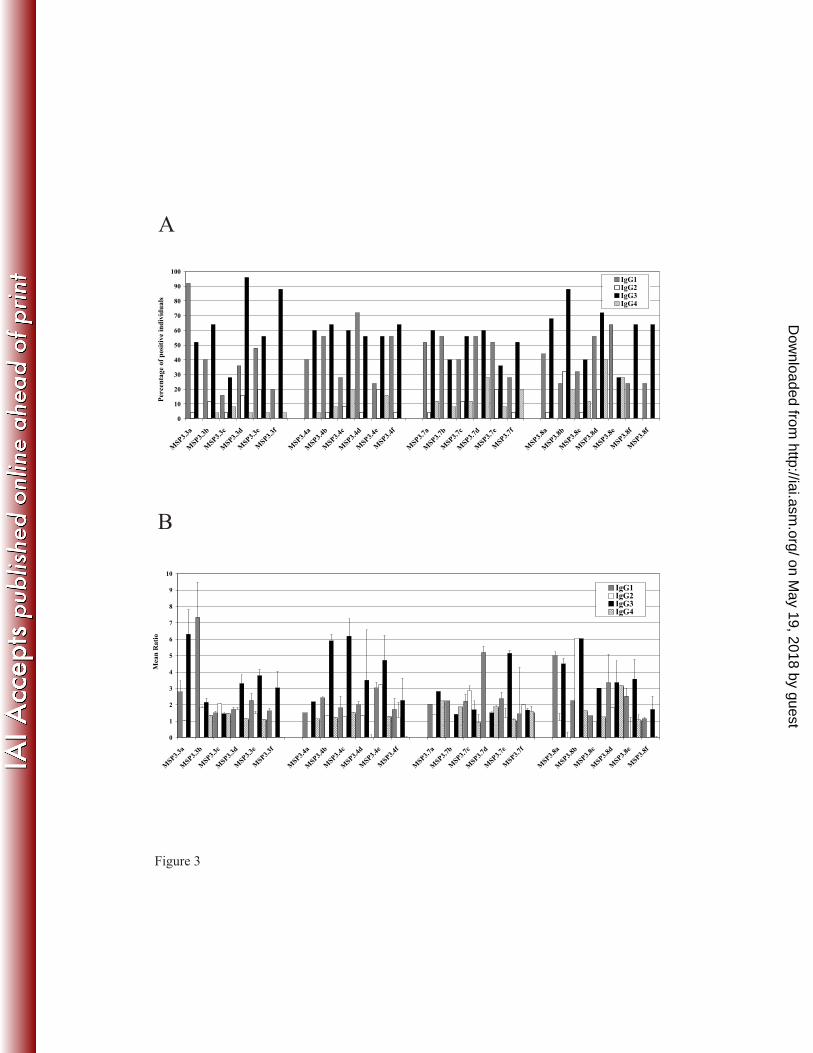
Multiple targets of antibodies triggering ADCI are found throughout the MSP3 family. 7
Human antibodies affinity-purified upon each of the 24 peptides of MSP3.3, MSP3.4, 8
MSP3.7 and MSP3.8, recognized both the corresponding peptide and the recombinant C-9
terminus region (not shown). More importantly, these affinity-purified antibodies were all 10
able to recognize the parasite native proteins by IFA, indicating that each synthetic peptide 11
mimicked at least one natural parasite epitope. 12
In order to allow comparisons in ADCI assays, these antibodies were adjusted 13
according to their IFA titre. The ADCI assay measures the intra-erythrocytic killing effect of 14
mediators released by monocytes, and the resulting reduction of parasite density. As shown in 15
figure 4, each of the 24 peptides from the four antigens was found to be a target of human 16
antibodies with potent monocyte-dependent antiplasmodial activity in the ADCI assay. For a 17
few of them (anti-3.3c, anti-3.3f, anti-3.4c and anti-3.7c) the ADCI effect was only marginal, 18
yet significant, whereas for the majority it was comparable to that of the PIAG, i.e. IgG of 19
protected African adults. The anti-plasmodial effect mediated by some of them, such as anti-20
3.3e, anti-3.4b and anti-3.8d antibodies was even higher than that observed using the PIAG. 21
This strong effect is remarkable as each peptide specific antibody corresponds to a very minor 22
subset of total malarial antibodies and a small subset of antibodies directed to the 6 MSP3 23
family proteins. Although antibodies specific to the recombinant antigens were all efficient in 24
ADCI (figure 4), the antibodies specific to several peptides (MSP3.3a, MSP3.3e, MSP3.4a, 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

15
MSP3.4b, MSP3.7a, MSP3.8a, MSP3.8b, MSP3.8d, MSP3.8e, MSP3.3f) mediated an effect 1
stronger than that observed with the corresponding recombinant antigen. These results differ 2
from those obtained previously with MSP3.1 where only 3 of the 6 peptides were the target of 3
biologically active antibodies (32). They are in keeping with results obtained using MSP3.2 4
peptide-specific antibodies which all had a strong inhibitory effect (33). Thus the present data 5
complement those obtained in our two previous studies, to characterize a very large series of 6
new targets for antibodies mediating the killing of P. falciparum parasite through the ADCI 7
mechanism. 8
9
Cross-reactions among MSP3 family members. 10
As mentioned above, a stretch of eleven amino acids is quasi-identical in the peptides 11
"b" derived from MSP3.1 and MSP3.2, and highly homologous in MSP3.7b, which suggests 12
that antibodies specific to these peptides may be cross-reactive. Indeed, in competition ELISA 13
assays, the MSP3.2b peptide at 200 g/ml concentration was able to decrease by 80% the 14
binding of anti-MSP3.1b antibodies to MSP3.1-Cterm. Similarly, the binding of anti-15
MSP3.7b was decreased by 60% by the competing MSP3.1b peptide (figure 5). These 16
findings extend the cross-reactivity observed previously between MSP3.1b and MSP3.2b 17
(33). The competition was less efficient between MSP3.1b and MSP3.7b than between 18
MSP3.1b and MSP3.2b (figure 5), most probably due to the fact that, within the highly 19
conserved stretch of eleven amino acids of MSP3.7b, there are substitutions in the first three 20
positions: YFA vs. ILG, and in the 6th position F/I, whereas MSP3.1b and MSP3.2b only 21
differ by a V/A substitution in the 10th
position (figure 2-B boxI). 22
The cross reactivity was also assessed among the second group of holomogous 23
peptides, i.e. peptides “d” (figure 2-B, box II), using antibodies immunopurified on each 24
peptide. The cross-reactivity network summarized in figure 6 indicates that naturally 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

16
occurring human antibodies specific to "peptide d" of each MSP3 protein cross-reacted with 1
at least one other antigen of the MSP3 family. Anti-MSP3-8d presented cross-reactivity with 2
only one other member of the gene family, whereas anti-MSP3.1d and anti-MSP3.2d 3
presented cross-reactivities with each of the remaining five proteins, i.e. reacted with all 6 4
MSP3 proteins. On the other hand, MSP3.4-Cterm and MSP3.8-Cterm appeared as the “most 5
antigenic”, as they were recognized by antibodies immuno-purified on each of the six “d” 6
peptides. Conversely, these results indicate that the limited point mutations occurring in the 7
highly conserved motif of the peptides "d" can also induce true antigenic divergence despite a 8
high level of sequence homology. In this manner the various homologous sequences generate 9
a large number of distinct antibody species with a wide pattern of reactivity to the 6 proteins. 10
11
A MSP3 multigenic construct induces in mice antibodies cross-reacting with each antigen 12
of the MSP3 family. 13
The above studies show that sequence homologies among MSP3 proteins generate a 14
large network of cross-reactions, despite sequence divergences particularly among the 15
flanking regions. Based on the combination of data from immunogenicity studies (7) and the 16
present antigenicity data, a first MSP3 polyantigenic construct covering the “b-c-d” peptide 17
sequences was designed and used to immunize mice. The mouse antibodies affinity-purified 18
upon the synthetic MSP3s-multigenic protein reacted with each of the six antigens of the 19
MSP3 family. However they differently recognized each MSP3 antigen as avidity indexes 20
were estimated as 0.4M, 0.5M, 0.7M, 0.9M, 2.6M and 2.8M for MSP3.1, MSP3.4, MSP3.7, 21
MSP3.8, MSP3.3 and MSP3.2, respectively. Therefore avidity of these antibodies was high 22
for MSP3.3 and MSP3.2, medium for MSP3.8, MSP3.7 and low for MSP3.4 and MSP3.1 23
(figure 7). 24
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

17
These results, and those above, indicate that the antigens of the MSP3 family are the targets of 1
a family of cross-reactive antibodies, with wide differences in terms of avidity and specificity. 2
3
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

18
DISCUSSION. 1
In order to design a multi-subunit MSP3 vaccine based on a sound immunological 2
rationale, we performed a detailed immunological study of the remaining four members of the 3
MSP3 family of proteins, to complement the data previously obtained on the other two, 4
MSP3.1 and MSP3.2 (32, 33). As a complement to the immunogenicity study reported 5
elsewhere (7), we performed in the present work a detailed antigenicity analysis of the 6
homologous C-terminal portions of these proteins, and evaluated the biological activity of the 7
corresponding antibodies. As in our previous studies with MSP3.1 and MSP3.2, we used both 8
recombinant proteins and peptides corresponding to homologous or divergent regions, in 9
order to select, based on immune responses from exposed individuals, those to include in 10
vaccine constructs. 11
Using sera from subjects spanning all ages living in a holoendemic region of Senegal, 12
we found that the conserved C-terminal regions of the six MSP3 proteins were all antigenic, 13
and that a large majority of subjects harboured antibodies to each of them. A more detailed 14
analysis of MSP3.3, MSP3.4, MSP3.7 and MSP3.8, was performed using for each of them six 15
overlapping peptides covering their C-terminal portion. Each of these 24 peptides was 16
antigenic, in accordance with what had previously been observed for the 12 peptides from 17
MSP3.1 and MSP3.2. This shows that each synthetic peptide defines at least one B-cell 18
epitope in the correct conformation. The recognition of the parasite native protein by anti-19
peptides antibodies, demonstrated by IFA assays, and indirectly confirmed by ADCI assays, 20
is an additional proof of the similarity of the peptides’ epitopes to those of the native proteins. 21
Besides their overall structural homology, the six C-terminal MSP3 proteins share 22
highly homologous sequences displayed on peptides "b", "d" (and "f"). The sequence 23
“ILGWEFGGGAP” from peptides "b" is well conserved among MSP3.1, MSP3.2 and 24
MSP3.7, and was therefore found to be targeted by antibodies that are fully cross-reactive to 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

19
these 3 antigens. The antibodies directed to the various peptides "d" presented a broad range 1
of cross-reactivities on each of the six MSP3 proteins, consistent with the occurrence of single 2
point amino acid changes among the homologous sequences. A malaria vaccine combining 3
these antigens would thus not only have the advantage of inducing antibodies to an increased 4
number of targets of ADCI, but presumably also to broaden the types of antibody species 5
raised against these proteins. 6
Twenty of the twenty-four peptides analysed reacted with antibodies which displayed 7
an IgG subclass pattern with dominant cytophilic responses. This was also found to be the 8
case for 4 of the 6 MSP3.1 peptides and all the 6 MSP3.2 peptides tested previously (32, 33). 9
The IgG3 type of responses was more prevalent than IgG1 responses whereas the levels of 10
both were equivalent. The importance of this finding is related to previous studies showing 11
that IgG antibodies either mediating parasite killing in vitro, or associated with protection in 12
vivo, are restricted to IgG1 and IgG3 subclasses (4, 16, 20, 30). In agreement with this 13
characteristic, all twenty-four peptides were indeed found to define antigenic determinants 14
able to trigger the antiparasitic ADCI mechanism. The ADCI anti-parasite activity of peptide-15
specific antibodies was, for more than half of them, as much or more effective as total IgG 16
from protected African adults. Therefore they constitute a large series of novel valuable 17
targets able to trigger the ADCI mechanism in addition to the nine targets previously 18
described in the other two members of the MSP3 family (32, 33). They also add to epitopes 19
identified in unrelated proteins such as GLURP, SERP (35, 39) and recently MSP1-Block2 20
(15). 21
The multiple targets of in vitro inhibitory antibodies, found in all six members of the 22
MSP3 family of proteins, suggests that they may play, in addition to MSP3.1, a role in 23
protection. The additional, and most unusual, full sequence conservation among several field 24
isolates of the C-terminal portion of each MSP3 gene (34) suggests that the conservation of 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

20
differences in both homologous and non-homologous regions has a critical function. Within 1
the regions of highest homology, the cross-reactive B-cell epitopes generate a wider diversity 2
in the affinity and fine-specificity, of the antibody repertoire, than would a single antigen. 3
Conversely, the conservation of these regions suggests a minimal selective advantage to 4
random mutations. Antibodies elicited by any member could still cross-react with other non-5
mutated members and mediate parasite inhibition. Within non-homologous regions, sequence 6
differences from one gene to the other, may be related to T-helper function, which was 7
documented in MSP3.1 (7, 32), each sequence being better fitted to a given MHC class-II 8
subset. The conservation of the diversity would provide improved T-cell help in individuals 9
with diverse MHC class-II genetic background so as to generate the same essential antibodies 10
directed to the conserved cross-reactive B-cell epitopes. These features open new perspectives 11
on the role of MSP3 antigens in the natural host-parasite interaction. Their function might be 12
to maintain the homeostasis between P. falciparum and human beings, by contributing to 13
control parasite densities through an indirect, non-sterilizing, defence mechanism (11). 14
Our findings have also practical consequences for vaccine development. Indeed, they 15
pave the way towards the rational design of a multi-subunit vaccine construct based on a 16
family of proteins having both homologous and divergent sequences that are highly 17
conserved. This combination strategy differs from constructs combining different 18
polymorphic antigens (13, 17, 19, 23, 24, 27, 38). The C-terminal sequences being conserved, 19
a polyantigenic MSP3 vaccine avoids antigen polymorphism, a recognized bottleneck for 20
vaccine development (18, 22, 26, 37).. 21
Finally the present antigenicity data has to be combined with results from 22
immunogenicty studies (7) to design improved constructs. Indeed, the latter suggest that 23
certain regions are involved in the regulation of the intensity of immune responses. This was 24
expected, as one of the remarkable characteristics of malaria infection in African children is 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

21
the overwhelming number of parasites harboured over childhood and adolescence. The 1
corresponding massive amounts of malarial antigens repeatedly stimulate their immune 2
system for years. Since the antibody titres to MSP3 and other antigens, remain very moderate 3
in children as in adults, it implies that malarial antigens must contain regulatory regions to 4
prevent the induction of high immune responses that could otherwise eradicate the parasite. 5
Such regions were identified in the immunogenicity study (7) where it was found that 6
removing the “e-f” region from MSP3.1 resulted in an increase by two orders of magnitude of 7
antibodies to the ADCI-relevant “b-d” region. In this manner the immunity induced by a 8
carefully designed immunogen should be more effective than natural exposure to the parasite. 9
Our first polyantigenic construct indeed capitalizes on both sets of available data to 10
include only the regions "b-d" of the six MSP3 antigens. The resulting multi-component 11
fusion protein was strongly immunogenic in mice, inducing biologically active antibodies, 12
cross-reacting with the 6 MSP3 family proteins with various avidities (the present study, and 13
(7)). However, the combination of data emerging from detailed antigenicity and 14
immunogenicity studies can be exploited in diverse manners, i.e. to design various constructs, 15
so as to best fit the general strategy of a multi-component vaccine based on the conserved 16
MSP3 family antigens aimed at triggering more efficiently the ADCI mechanism on diverse 17
human genetic backgrounds. 18
19
20
21
22
23
24
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

22
TABLE AND FIGURES LEGENDS. 1
TABLE 1: Total IgG responses upon the C-terminal portion of the six MSP3 family proteins 2
in hyperimmune serum samples from adult individuals (n=45) from Dielmo, Senegal. To 3
calculate the prevalence values, antibody responses were considered positive when the net 4
OD value of each serum sample was > the mean OD+ 3SD of control serum samples. The 5
prevalence values were not statistically different (Pearson’s 2 test). The mean of net OD 6
values represents the mean responses of the positive serum samples. 7
8
FIGURE 1: Cytophilic IgG subclasses responses to the C-terminal portion of the six MSP3 9
antigens in serum samples of individuals of all age groups (n=76, 0.9 -80.4 years, mean age 10
=14.4 years) from Dielmo, Senegal. To calculate the prevalence values, antibody responses 11
were considered positive when the net OD value of each serum sample was > the mean OD+ 12
3SD of control serum samples. The mean of net OD values represents the mean responses of 13
the positive serum samples. Bars are standard errors of the mean. 14
15
FIGURE 2: Panel A, Matrix of overall percent homology (identity and similarity) of the C-16
terminal sequences of the six MSP3 proteins obtained with EMBOSS pairwise alignments; 17
Panel B, Truncated multiple sequence alignment of the MSP3 proteins family showing the 18
three regions with common linear motifs present in the C-terminal portion. Conserved 19
residues are highlighted in bold. 20
FIGURE 3: Prevalences (A) and Titers (B) of IgG subclasses distribution in 24 peptides 21
derived from MSP3.3, MSP3.4, MSP3.7 and MSP3.8 in serum samples of malaria exposed 22
individuals (n=25; age range 2-54 years, mean age 16.86) from Dielmo village. Bars are 23
standard errors of the mean. 24
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

23
FIGURE 4: Antibody-dependent cellular inhibitory effect of human antibodies affinity 1
purified upon the 24 peptides and the 4 recombinant antigens covering the C-terminal regions 2
of MSP3 proteins. In black positive controls, gray for anti-MSP3.3 peptides, White for anti-3
MSP3.4 peptides, Hatched for anti-MSP3.7 peptides and dotted for anti-MSP3.8 peptides. 4
PIAG, Pool of African Immune Globulins. Each of the affinity-purified antibodies was tested 5
in duplicate at a concentration corresponding to a 1/20 final IFA titer. The PIAG and 6
antibodies affinity purified upon recombinant MSP3.1-Cterm were used as positive controls 7
in each experiment. The SGI values are expressed as a % of the effect of PIAG (100%); 8
values > 30% are considered significant. 9
10
FIGURE 5: Antibodies cross-reactivity between peptides "b" of the MSP3 proteins family. 11
Open squares: binding of anti-MSP3.1b specific antibodies upon the recombinant antigen 12
MSP3.1-Cterm in the presence of increasing concentrations of the MSP3.2b peptide. Plain 13
squares: binding of anti-MSP3.7b specific antibodies upon the recombinant antigen MSP3.7-14
Cterm in the presence of increasing concentrations of the MSP3.1b peptide. Affinity purified 15
antibodies were preincubated with different amounts of peptides before being tested in 16
ELISA. 17
18
FIGURE 6: Pattern of cross-reactivity of antibodies to peptides "d" from the MSP3 proteins 19
family. Binding pattern of antibodies specific to the peptides "d" of the six MSP3 proteins 20
were deduced from their ELISA reactivity against the different recombinant antigens. Note 21
that the reactivity to the homologous protein (e.g. Anti-MSP3-8d peptide with MSP3-8 C-22
term protein) was present for each of them, however is not shown for clarity reasons. A 23
significant cross-reactivity corresponds to OD values 0.2. 24
25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

24
FIGURE 7: Avidity of antibodies induced in mice after immunisation with a MSP3- family 1
multigenic construct. The curves represent the reactivity on the MSP3 multigenic construct 2
itself and on the different MSP3 antigens in the presence of increasing concentrations of 3
chaotropic ion, ammonium thiocyanate (NH4SCN). 4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

26
Acknowledgments: 1
We are grateful to Christian Roussilhon for helpful discussions and Nicolas Puchot for help 2
with illustrations. 3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

27
REFERENCES 1
1. Audran, R., M. Cachat, F. Lurati, S. Soe, O. Leroy, G. Corradin, P. Druilhe, and 2
F. Spertini. 2005. Phase I malaria vaccine trial with a long synthetic peptide derived 3
from the merozoite surface protein 3 antigen. Infect Immun 73:8017-8026. 4
2. Bottius, E., L. BenMohamed, K. Brahimi, H. Gras, J. P. Lepers, L. Raharimalala, 5
M. Aikawa, J. Meis, B. Slierendregt, A. Tartar, A. Thomas, and P. Druilhe. 1996. 6
A novel Plasmodium falciparum sporozoite and liver stage antigen (SALSA) defines 7
major B, T helper, and CTL epitopes. J Immunol 156:2874-2884. 8
3. Bouharoun-Tayoun, H., P. Attanath, A. Sabchareon, T. Chongsuphajaisiddhi, 9
and P. Druilhe. 1990. Antibodies that protect humans against Plasmodium falciparum 10
blood stages do not on their own inhibit parasite growth and invasion in vitro, but act 11
in cooperation with monocytes. J Exp Med 172:1633-1641. 12
4. Bouharoun-Tayoun, H., and P. Druilhe. 1992. Plasmodium falciparum malaria: 13
evidence for an isotype imbalance which may be responsible for delayed acquisition 14
of protective immunity. Infect Immun 60:1473-1481. 15
5. Bouharoun-Tayoun, H., C. Oeuvray, F. Lunel, and P. Druilhe. 1995. Mechanisms 16
underlying the monocyte-mediated antibody-dependent killing of Plasmodium 17
falciparum asexual blood stages. J Exp Med 182:409-418. 18
6. Brahimi, K., J. L. Perignon, M. Bossus, H. Gras, A. Tartar, and P. Druilhe. 1993. 19
Fast immunopurification of small amounts of specific antibodies on peptides bound to 20
ELISA plates. J Immunol Methods 162:69-75. 21
7. Daher, L. J., C. G. Demanga, E. Prieur, J. L. Perignon, H. Bouharoun-Tayoun, 22
and P. Druilhe. 2009. Towards the rational design of a malaria vaccine construct: 23
example of the MSP3 family. Part I: Immunogenicity studies in models. Infect 24
Immun, in press. 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

28
8. Druilhe, P., and H. Bouharoun-Tayoun. 2002. Antibody-dependent cellular 1
inhibition assay. Methods Mol Med 72:529-534. 2
9. Druilhe, P., and H. Bouharoun-Tayoun. 2002. Human antibody subclass ELISA. 3
Methods Mol Med 72:457-459. 4
10. Druilhe, P., P. Hagan, and G. A. Rook. 2002. The importance of models of infection 5
in the study of disease resistance. Trends Microbiol 10:S38-46. 6
11. Druilhe, P., and J. L. Perignon. 1997. A hypothesis about the chronicity of malaria 7
infection. Parasitol Today 13:353-357. 8
12. Druilhe, P., F. Spertini, D. Soesoe, G. Corradin, P. Mejia, S. Singh, R. Audran, A. 9
Bouzidi, C. Oeuvray, and C. Roussilhon. 2005. A malaria vaccine that elicits in 10
humans antibodies able to kill Plasmodium falciparum. PLoS Med 2:e344. 11
13. Fluck, C., S. Schopflin, T. Smith, B. Genton, M. P. Alpers, H. P. Beck, and I. 12
Felger. 2007. Effect of the malaria vaccine Combination B on merozoite surface 13
antigen 2 diversity. Infect Genet Evol 7:44-51. 14
14. Friguet, B., A. F. Chaffotte, L. Djavadi-Ohaniance, and M. E. Goldberg. 1995. 15
Under proper experimental conditions the solid-phase antigen does not disrupt the 16
liquid phase equilibrium when measuring dissociation constants by competition 17
ELISA. J Immunol Methods 182:145-150. 18
15. Galamo, C. D., A. Jafarshad, C. Blanc, and P. Druilhe. 2009. Anti-MSP1 Block 2 19
Antibodies Are Effective at Parasite Killing in an Allele-Specific Manner by 20
Monocyte-Mediated Antibody-Dependent Cellular Inhibition. J Infect Dis 199:1151-21
1154. 22
16. Garraud, O., S. Mahanty, and R. Perraut. 2003. Malaria-specific antibody 23
subclasses in immune individuals: a key source of information for vaccine design. 24
Trends Immunol 24:30-35. 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

29
17. Genton, B., I. Betuela, I. Felger, F. Al-Yaman, R. F. Anders, A. Saul, L. Rare, M. 1
Baisor, K. Lorry, G. V. Brown, D. Pye, D. O. Irving, T. A. Smith, H. P. Beck, and 2
M. P. Alpers. 2002. A recombinant blood-stage malaria vaccine reduces Plasmodium 3
falciparum density and exerts selective pressure on parasite populations in a phase 1-4
2b trial in Papua New Guinea. J Infect Dis 185:820-827. 5
18. Genton, B., and Z. H. Reed. 2007. Asexual blood-stage malaria vaccine 6
development: facing the challenges. Curr Opin Infect Dis 20:467-475. 7
19. Hu, J., Z. Chen, J. Gu, M. Wan, Q. Shen, M. P. Kieny, J. He, Z. Li, Q. Zhang, Z. 8
H. Reed, Y. Zhu, W. Li, Y. Cao, L. Qu, Z. Cao, Q. Wang, H. Liu, X. Pan, X. 9
Huang, D. Zhang, X. Xue, and W. Pan. 2008. Safety and immunogenicity of a 10
malaria vaccine, Plasmodium falciparum AMA-1/MSP-1 chimeric protein formulated 11
in montanide ISA 720 in healthy adults. PLoS ONE 3:e1952. 12
20. Jafarshad, A., M. H. Dziegiel, R. Lundquist, L. K. Nielsen, S. Singh, and P. L. 13
Druilhe. 2007. A novel antibody-dependent cellular cytotoxicity mechanism involved 14
in defense against malaria requires costimulation of monocytes FcgammaRII and 15
FcgammaRIII. J Immunol 178:3099-3106. 16
21. Jensen, J. B. 2002. In vitro culture of Plasmodium parasites. Methods Mol Med 17
72:477-488. 18
22. Lalitha, P. V., P. Malhotra, R. Chattopadhyay, and V. S. Chauhan. 1999. 19
Plasmodium falciparum: variations in the C-terminal cysteine-rich region of the 20
merozoite surface protein-1 in field samples among Indian isolates. Exp Parasitol 21
92:12-18. 22
23. Malkin, E., J. Hu, Z. Li, Z. Chen, X. Bi, Z. Reed, F. Dubovsky, J. Liu, Q. Wang, 23
X. Pan, T. Chen, B. Giersing, Y. Xu, X. Kang, J. Gu, Q. Shen, K. Tucker, E. 24
Tierney, W. Pan, C. Long, and Z. Cao. 2008. A Phase 1 trial of PfCP2.9: An 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

30
AMA1/MSP1 chimeric recombinant protein vaccine for Plasmodium falciparum 1
malaria. Vaccine. 2
24. Miller, L. H., T. Roberts, M. Shahabuddin, and T. F. McCutchan. 1993. Analysis 3
of sequence diversity in the Plasmodium falciparum merozoite surface protein-1 4
(MSP-1). Mol Biochem Parasitol 59:1-14. 5
25. Oeuvray, C., H. Bouharoun-Tayoun, H. Gras-Masse, E. Bottius, T. Kaidoh, M. 6
Aikawa, M. C. Filgueira, A. Tartar, and P. Druilhe. 1994. Merozoite surface 7
protein-3: a malaria protein inducing antibodies that promote Plasmodium falciparum 8
killing by cooperation with blood monocytes. Blood 84:1594-1602. 9
26. Pan, W., D. Huang, Q. Zhang, L. Qu, D. Zhang, X. Zhang, X. Xue, and F. Qian. 10
2004. Fusion of two malaria vaccine candidate antigens enhances product yield, 11
immunogenicity, and antibody-mediated inhibition of parasite growth in vitro. J 12
Immunol 172:6167-6174. 13
27. Polley, S. D., W. Chokejindachai, and D. J. Conway. 2003. Allele frequency-based 14
analyses robustly map sequence sites under balancing selection in a malaria vaccine 15
candidate antigen. Genetics 165:555-561. 16
28. Polley, S. D., K. K. Tetteh, J. M. Lloyd, O. J. Akpogheneta, B. M. Greenwood, K. 17
A. Bojang, and D. J. Conway. 2007. Plasmodium falciparum merozoite surface 18
protein 3 is a target of allele-specific immunity and alleles are maintained by natural 19
selection. J Infect Dis 195:279-287. 20
29. Pullen, G. R., M. G. Fitzgerald, and C. S. Hosking. 1986. Antibody avidity 21
determination by ELISA using thiocyanate elution. J Immunol Methods 86:83-87. 22
30. Roussilhon, C., C. Oeuvray, C. Muller-Graf, A. Tall, C. Rogier, J. F. Trape, M. 23
Theisen, A. Balde, J. L. Perignon, and P. Druilhe. 2007. Long-term clinical 24
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

31
protection from falciparum malaria is strongly associated with IgG3 antibodies to 1
merozoite surface protein 3. PLoS Med 4:e320. 2
31. Sabchareon, A., T. Burnouf, D. Ouattara, P. Attanath, H. Bouharoun-Tayoun, P. 3
Chantavanich, C. Foucault, T. Chongsuphajaisiddhi, and P. Druilhe. 1991. 4
Parasitologic and clinical human response to immunoglobulin administration in 5
falciparum malaria. Am J Trop Med Hyg 45:297-308. 6
32. Singh, S., S. Soe, J. P. Mejia, C. Roussilhon, M. Theisen, G. Corradin, and P. 7
Druilhe. 2004. Identification of a conserved region of Plasmodium falciparum MSP3 8
targeted by biologically active antibodies to improve vaccine design. J Infect Dis 9
190:1010-1018. 10
33. Singh, S., S. Soe, C. Roussilhon, G. Corradin, and P. Druilhe. 2005. Plasmodium 11
falciparum merozoite surface protein 6 displays multiple targets for naturally 12
occurring antibodies that mediate monocyte-dependent parasite killing. Infect Immun 13
73:1235-1238. 14
34. Singh S., S. S., Barnwell J. and Druilhe P. in press 2009. A merozoite surface multi 15
antigen family of P. falciparum : a possible means to ensure parasite survival. 16
PlosOne. 17
35. Soe, S., S. Singh, D. Camus, T. Horii, and P. Druilhe. 2002. Plasmodium 18
falciparum serine repeat protein, a new target of monocyte-dependent antibody-19
mediated parasite killing. Infect Immun 70:7182-7184. 20
36. Soe, S., M. Theisen, C. Roussilhon, K. S. Aye, and P. Druilhe. 2004. Association 21
between protection against clinical malaria and antibodies to merozoite surface 22
antigens in an area of hyperendemicity in Myanmar: complementarity between 23
responses to merozoite surface protein 3 and the 220-kilodalton glutamate-rich 24
protein. Infect Immun 72:247-252. 25
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

32
37. Stowers, A. W., V. Cioce, R. L. Shimp, M. Lawson, G. Hui, O. Muratova, D. C. 1
Kaslow, R. Robinson, C. A. Long, and L. H. Miller. 2001. Efficacy of two alternate 2
vaccines based on Plasmodium falciparum merozoite surface protein 1 in an Aotus 3
challenge trial. Infect Immun 69:1536-1546. 4
38. Takala, S. L., D. Coulibaly, M. A. Thera, A. Dicko, D. L. Smith, A. B. Guindo, A. 5
K. Kone, K. Traore, A. Ouattara, A. A. Djimde, P. S. Sehdev, K. E. Lyke, D. A. 6
Diallo, O. K. Doumbo, and C. V. Plowe. 2007. Dynamics of polymorphism in a 7
malaria vaccine antigen at a vaccine-testing site in Mali. PLoS Med 4:e93. 8
39. Theisen, M., S. Soe, S. G. Jessing, L. M. Okkels, S. Danielsen, C. Oeuvray, P. 9
Druilhe, and S. Jepsen. 2000. Identification of a major B-cell epitope of the 10
Plasmodium falciparum glutamate-rich protein (GLURP), targeted by human 11
antibodies mediating parasite killing. Vaccine 19:204-212. 12
40. Trape, J. F., C. Rogier, L. Konate, N. Diagne, H. Bouganali, B. Canque, F. 13
Legros, A. Badji, G. Ndiaye, P. Ndiaye, and et al. 1994. The Dielmo project: a 14
longitudinal study of natural malaria infection and the mechanisms of protective 15
immunity in a community living in a holoendemic area of Senegal. Am J Trop Med 16
Hyg 51:123-137. 17
41. Trucco, C., D. Fernandez-Reyes, S. Howell, W. H. Stafford, T. J. Scott-Finnigan, 18
M. Grainger, S. A. Ogun, W. R. Taylor, and A. A. Holder. 2001. The merozoite 19
surface protein 6 gene codes for a 36 kDa protein associated with the Plasmodium 20
falciparum merozoite surface protein-1 complex. Mol Biochem Parasitol 112:91-101. 21
22
23
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Mean OD
Value
Range
[]
Prevalence
(%)
MSP3.1-Cterm 2.2±1.2 [0.2-3.8] 91.1
MSP3.2-Cterm 2.5±1.0 [0.2-3.8] 88.9
MSP3.3-Cterm 1.9 ±1.0 [0.1-3.6] 93.3
MSP3.4-Cterm 2.3 ±1.1 [0.2-4.1] 95.6
MSP3.7-Cterm 1.3 ±0.8 [0.1-3.4] 91.1
MSP3.8-Cterm 1.8 ±0.9 [0.2-3.6] 91.1
TABLE 1
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Figure 1
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
MSP3.1-
Cterm
MSP3.2-
Cterm
MSP3.3-
Cterm
MSP3.4-
Cterm
MSP3.7-
Cterm
MSP3.8-
Cterm
IgG1
IgG3
87%
86%
72%
86%
82%
84%
83%
93%63%
84%
68%
79%
Mea
n O
.D v
alu
es
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
MSP3.1-Cterm MSP3.2-Cterm MSP3.3-Cterm MSP3.4-Cterm MSP3.7-Cterm MSP3.8-Cterm
MSP3.1-Cterm 100% 57.80% 55.00% 54.20% 51.80% 50.20%
MSP3.2-Cterm 57.80% 100% 47.10% 49.20% 49.00% 49.40%
MSP3.3-Cterm 55.00% 47.10% 100% 48.40% 46.10% 44.00%
MSP3.4-Cterm 54.20% 49.20% 48.40% 100% 49.50% 50.40%
MSP3.7-Cterm 51.80% 49.00% 46.10% 49.50% 100% 43.50%
MSP3.8-Cterm 50.20% 49.40% 44.00% 50.40% 43.50% 100%
A
BMSP3.1 --ILGWEFGGGVP-204--LSHLYVSSKDKENISKENDD-232--MKTLAGLIKGNNQIDSTLKDL-344--
MSP3.2 --ILGWEFGGGAP-193--LEQIKIPSWDRNNIPDENEQ-226--VKTLVGLFNEKNEIDSTINNL-361--
MSP3.3 --GIEFSGGlyfn-272--LESVNLTSWDKEDIVKENED-306--ITALISLLNGKNELDATIRRL-414--
MSP3.4 --NLERGLGSGAL-535--LELIKLTSKDEEDIIKHNED-567--VKKLFSLFNDNNNLETIFKGL-688--
MSP3.7 --YFAWEIGGGAP-246--LEHVKITSWDKEDIIKENED-279--VNLLFNFLQTNNQLDPSLKDL-395--
MSP3.8 --RLELSSGSSSL-580--LELVPLSLSDIEQIANESED-613--VNSIISLLEEGNGSDSTLNSL-753--
Box I Box II Box III
Figure 2
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
2
3
4
5
6
7
8
9
:
;
32
OUR505c
OUR505d
OUR505e
OUR505f
OUR505g
OUR505h
OUR506c
OUR506d
OUR506e
OUR506f
OUR506g
OUR506h
OUR509c
OUR509d
OUR509e
OUR509f
OUR509g
OUR509h
OUR50:c
OUR50:d
OUR50:e
OUR50:f
OUR50:g
OUR50:h
Ogcp"Tcvkq
KiI3KiI4KiI5KiI6
2
32
42
52
62
72
82
92
:2
;2
322
OUR505c
OUR505d
OUR505e
OUR505f
OUR505g
OUR505h
OUR506c
OUR506d
OUR506e
OUR506f
OUR506g
OUR506h
OUR509c
OUR509d
OUR509e
OUR509f
OUR509g
OUR509h
OUR50:c
OUR50:d
OUR50:e
OUR50:f
OUR50:g
OUR50:h
OUR50:h
Rgtegp
vcig"qh"r
qukvkxg"kp
fkxkfw
cnu
KiI3KiI4KiI5KiI6
A
B
Figure 3
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
0
10
20
30
40
50
60
70
80
90
100
110
120
PIAG
anti-
MSP3.
3a
anti-
MSP3.
3b
anti-
MSP3.
3c
anti-
MSP3.
3d
anti-
MSP3.
3e
anti-
MSP3.
3f
anti-
MSP3.
3-C
term
anti-
MSP3.
4a
anti-
MSP3.
4b
anti-
MSP3.
4c
anti-
MSP3.
4d
anti-
MSP3.
4e
anti-
MSP3.
4f
anti-
MSP3.
4-C
term
anti-
MSP3.
7a
anti-
MSP3.
7.b
anti-
MSP3.
7c
anti-
MSP3.
7d
anti-
MSP3.
7e
anti-
MSP3.
7f
anti-
MSP3.
7-C
term
anti-
MSP3.
8a
anti-
MSP3.
8b
anti-
MSP3.
8c
anti-
MSP3.
8d
anti-
MSP3.
8e
anti-
MSP3.
8f
anti-
MSP3.
8-ct
erm
anti-
MSP3.
1-C
term
% S
pec
ific
Gro
wth
In
hib
itory
In
dex
(S
GI)
Figure 4
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
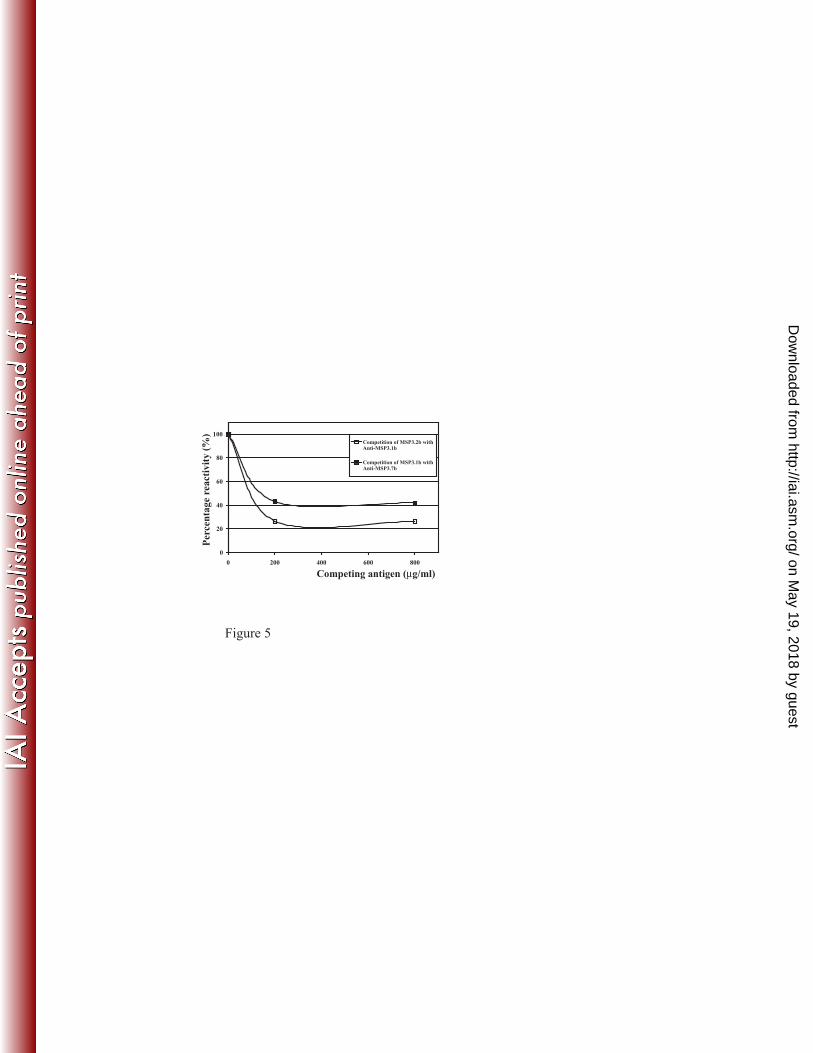
Figure 5
0
20
40
60
80
100
0 200 400 600 800
Competition of MSP3.2b withAnti-MSP3.1b
Competition of MSP3.1b with Anti-MSP3.7b
Per
cen
tage
react
ivit
y (
%)
Competing antigen (µg/ml)
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

Anti-
MSP3.1d
Anti-
MSP3.7d
Anti-
MSP3.2d
Anti-
MSP3.3d
Anti-
MSP3.4d
Anti-
MSP3.8d
MSP3.1
-Cterm
MSP3.2-Cterm
MSP3.3
-Cterm
MSP3.4
-Cterm
MSP3.7-Cterm
MSP3.8-Cterm
Figure 6
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
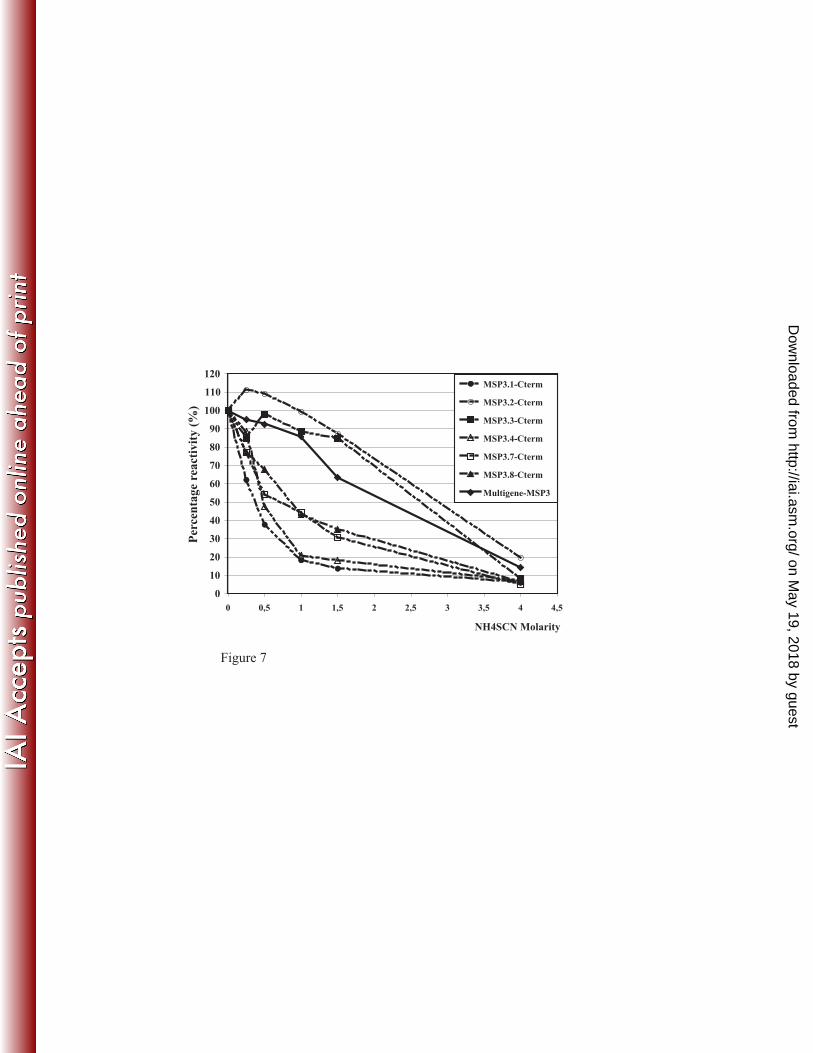
Figure 7
0
10
20
30
40
50
60
70
80
90
100
110
120
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5
NH4SCN Molarity
MSP3.1-Cterm
MSP3.2-Cterm
MSP3.3-Cterm
MSP3.4-Cterm
MSP3.7-Cterm
MSP3.8-Cterm
Multigene-MSP3
Per
cen
tage
react
ivit
y (
%)
on May 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from