Embed Size (px)
Citation preview

lable at ScienceDirect
Soil Biology & Biochemistry 43 (2011) 351e358
Contents lists avai
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lb io
The oribatid mite Scheloribates moestus (Acari: Oribatida) alters litterchemistry and nutrient cycling during decomposition
Kyle Wickings*, A. Stuart GrandyDepartment of Crop and Soil Sciences, Michigan State University, East Lansing, MI 48824, USA
a r t i c l e i n f o
Article history:Received 3 May 2010Received in revised form27 October 2010Accepted 28 October 2010Available online 13 November 2010
Keywords:Oribatid mitesLitter decompositionExtracellular enzymesNutrient cycling
* Corresponding author. Tel.: þ1 517 355 0271x124E-mail address: [email protected] (K. Wickings).
0038-0717/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.soilbio.2010.10.023
a b s t r a c t
It is widely accepted that microarthropods influence decomposition dynamics but we know relativelylittle about their effects on litter chemistry, extracellular enzyme activities, and other finer-scaledecomposition processes. Further, few studies have investigated the role of individual microarthropodspecies in litter decomposition. The oribatid mite Scheloribates moestus Banks (Acari: Oribatida) isabundant in many U.S. ecosystems. We examined the potential effects of S. moestus on litter decom-position dynamics and chemical transformations, and whether these effects are influenced by variationin initial litter quality. We collected corn and oak litter from habitats with large populations of S. moestusand in microcosms with and without mites measured respiration rates, nitrogen availability, enzymeactivities, and molecular-scale changes in litter chemistry. Mites stimulated extracellular enzymeactivities, enhanced microbial respiration rates by 19% in corn litter and 17% in oak litter over 62 days,and increased water-extractable organic C and N. Mites decreased the relative abundance of poly-saccharides in decomposing corn litter but had no effect on oak litter chemistry, suggesting that theeffects of S. moestus on litter chemistry are constrained by initial litter quality. We also compared thechemistry of mite feces to unprocessed corn litter and found that feces had a higher relative abundanceof polysaccharides and phenols and a lower relative abundance of lignin. Our study establishes thatS. moestus substantially changes litter chemistry during decomposition, but specific effects vary withinitial litter quality. These chemical transformations, coupled with other observed changes in decom-position rates and nutrient cycling, indicate that S. moestus could play a key role in soil C cyclingdynamics.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The soil fauna consists of an array of species that collectivelyhave a profound influence on ecosystem processes, includingdecomposition and nutrient cycling (Wolters, 2000; Coleman,2008; Ekschmitt et al., 2008). Soil animals affect litter decomposi-tion directly by ingesting and fragmenting litter and indirectly byaltering the biomass and community structure of microbialdecomposers (Hanlon and Anderson, 1979; Cortet et al., 2003;Coleman et al., 2004). However, faunal contributions to decompo-sition and nutrient cycling are variable and depend on the identity,density and activity of the resident species, as well as on theirinteractions with environmental conditions and substrate quality(Anderson et al., 1985; Bardgett and Chan, 1999; Lenoir et al., 2007;Ayres et al., 2010). Thus, predicting how changes in soil faunal
7; fax: þ1 517 355 0270.
All rights reserved.
communities will influence ecosystem function may requireknowledge of individual species, their behavior, and their effect onsoil processes under different environmental conditions.
One potentially important but largely unexplored way that soilanimals could affect decomposition dynamics is by altering litterchemistry during decomposition. As plant litter passes through theanimal digestive tract its chemical composition may be altered byenzymatic activity, organic matter mineralization and nutrientabsorption (Gasdorf and Goodnight, 1963; Ziechman, 1994;Guggenberger et al., 1996; Hopkins et al., 1998; Gillon and David,2001; Rawlins et al., 2006; Hedde et al., 2007; Filley et al., 2008;Geib et al., 2008). These changes in organic matter chemistry mayhave implications for soil aggregation and sorption dynamics, aswell as for other biogeochemical processes that influence long-term C stabilization in soil (Golchin et al., 1994; Kleber et al., 2007).However, chemical transformations by soil animals have only beenshown in a limited number of studies, have rarely used modernspectroscopic chemical approaches, and have focused mostly onmacrofauna such as earthworms, millipedes and isopods (Teuben

K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358352
and Verhoef, 1992; Wolters, 2000; Rawlins et al., 2006; Filley et al.,2008; Geib et al., 2008).
Oribatid mites are among the most abundant soil mesofauna,ranging from 25,000 to 500,000 individuals m�2 in terrestrialecosystems (Coleman et al., 2004). Studies have shown that thepresence of oribatid mites can increase litter mass loss, stimulatemicrobial respiration rates and decrease fungal biomass (Siepel andMaaskamp, 1994; Kaneko, 1998; Hansen, 1999; Heneghan et al.,1999). However, little is known about their effects on other aspectsof decomposition including the alteration of microbial extracellularenzymes and transformations of litter chemistry. Further, predictingthe effects of oribatids on decomposition dynamics requires a betterunderstanding of the ways inwhich individual species interact withlitter quality (Hansen, 1999; Gergócs and Hufnagel, 2009).
Our recent experiments show that the oribatid mite Schelor-ibates moestus Banks (Scheloribatidae) is found in many Michiganecosystems at densities approaching 200 individuals g�1 dry litter(Wickings, Unpublished results). Both adults and juveniles maycontribute significantly to decomposition through feeding on litterand microbes as well as through the production of feces. Given itspopulation size and potential activity, S. moestusmay play a key rolein regulating decomposition in cultivated and native ecosystems.Our overall objective was to evaluate the potential effects of S.moestus on corn and oak litter decomposition dynamics and, morespecifically, to determine whether S. moestus changes C and Ncycling rates, enzyme activities, and chemical transformations indecomposing litter.
2. Methods
2.1. Litter and mite collection
Red oak (Quercus rubra L.) and corn (Zea mays L.) leaf litter werecollected from the Michigan State University W.K. Kellogg Biolog-ical Station Long-Term Ecological Research Site (KBS, LTER) inOctober 2008. Micro-invertebrates were extracted from litter usingBerlese funnels (Bioquip, Rancho Dominguez, CA) over a 3 d periodinto liter-sized plastic boxes lined with a moistened plaster-char-coal mixture (10:1 by weight). S. moestus was identified fromsamples and compared with type specimens from the Museum ofComparative Zoology, Harvard University, Cambridge, MA.Approximately 4000 S. moestus adults were collected in total(approximately 3000 individuals from corn, 1000 individuals fromoak) and all individuals were transferred to separate live boxes toavoid predation by other arthropods. Approximately 2 g of eithercorn or oak leaf litter, moistened with deionized water, were placedin each box to provide food and habitat for S. moestus. After micro-invertebrates were extracted, corn and oak litter were removedfrom Berlese funnels and dried completely at 60 �C. All litter wascut and sieved to fragments ranging 2e5 mm after drying andfurther defaunated using two successive freezeeheat cycles (�80�
and þ80 �C) similar to those used by Bardgett et al. (1998).
2.2. Microcosm setup
Microcosms consisted of a 70 mL polypropylene bottle (litterarena), from which the bottom was removed and replaced with38 mm stainless steel mesh, contained within a 120 mL glass jar.Before being placed in jars, 120 litter arenas received 1 g ofdefaunated corn or oak litter (n ¼ 60 corn and 60 oak arenas). Oncelitter was arranged in an arena, it was soaked in a litter homogenatefor 2 h to inoculate the microcosms with microbes. Homogenateswere created by agitating 500 g of field-moist corn or oak litter in4 L of deionized water for approximately 15 m. Suspensions werethen filtered through a paper filter (Whatman, grade 1, particle
retention 11 mm) to remove most particulate organic matter whileretaining bacteria or fungal spores. After soaking, arenas wereallowed to drain for 5 h before incubating in the dark at 22 �C for 5 dto allow microbial activity to increase. At the end of the 5-d incu-bation period, all arenas were placed into glass jars. In an effort tomaintain adequate litter moisture contents, we placed 10 mLof water in the bottom of each jar. The litter arenas wereelevated w5 mm above the water using polypropylene stands.Twenty adult individuals of S. moestus were added to each of 60arenas (n ¼ 30 corn and 30 oak) with the remaining 60 arenasreceiving no mites. Mite numbers were chosen to be consistentwith densities of S. moestusmeasured in field sites at KBS (Wickingset al., Unpublished results). All microcosms were covered withplastic wrap to retain moisture but still allow for some gasexchange. Microcosms were kept at approximately 21 �C on a 12 hlightedark cycle for the duration of the experiment (62 d).
2.3. Microcosm analysis
At 6, 13, 22, 34, 48 and 62 d of incubation, 20 microcosms weredestructively harvested for the measurement of enzyme activity,inorganic N, and dissolved organic carbon (DOC) and nitrogen(DON). Numbers of live mites were recorded at each harvest date at400� magnification.
2.3.1. Litter C min and compositionAt 3-d intervals, the plastic covering was removed from 20 litter
arenas, which were then transferred to dry 120 mL jars and cappedwith lids fitted with rubber septa to quantify microbial respirationrate. The arenas used for CO2 measurements were randomlyselected at the beginning of the experiment. These same arenaswere used for all CO2 measurements and were subsequently thelast to be destructively harvested. CO2 concentration of the jarheadspace was measured using an LI-820 infrared CO2 analyzer(Licor Biosciences, Lincoln, NE) at 30-m intervals over the course of1 h (0, 30 and 60 m) and used to determine production rate(mg CO2eC g�1 litter d�1).
Chemical composition of litter from time zero and after 62 d ofdecomposition was determined using pyrolysis gas chromatogra-phyemass spectroscopy (py-gc/ms) using previously describedmethods (Grandy et al., 2009; Wickings et al., 2010). Briefly,samples were pulse-pyrolyzed in quartz tubes on a Pyroprobe 5150(CDS Analytical Inc., Oxford, PA) using a pyrolysis temperature of600 �C. Pyrolysis products were transferred onto a gas chromato-graph (Trace GC Ultra, Thermo Scientific, West Palm Beach, FL)where they were further separated upon passage through a heated,fused silica capillary column (60 m � 0.25 mm i.d.). GC oventemperature was increased from 40 to 270 �C at a rate of 5 �C m�1
with a final temperature ramp to 310 �C at 30 �C m�1. Finally,compounds were transferred to a mass spectrometer (Polaris Q,Thermo Scientific,West Palm Beach, FL) via a 270 �C heated transferline. The abundance of all ions was scaled relative to the largestpeak. Peaks were compared to reference spectra using the Auto-mated Mass Spectral Deconvolution and Identification program(AMDIS V 2.65) and the National Institute of Standards and Tech-nology library (NIST).
In order to better understand the direct effects of S. moestus onlitter chemistry we also analyzed the chemical composition of itsfeces. Two additional S. moestus live boxes, containing approxi-mately 1000 adult mites each, were also maintained for the dura-tion of the study. Four nylon filter disks were placed in each live boxand each disk received approximately 1 g of litter (one box con-tained corn and one contained oak). Live boxes were kept atapproximately 21 �C and deionized water was added to the plasterbase of each box as needed to maintain adequate moisture. At

K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358 353
2e3 d intervals, mite feces were collected from the surface of eachfilter disk using a modified vacuum apparatus consisting of a 70 mLquartz tube filled half way with glass wool and attached toa vacuum pump. All feces were stored at �80 �C prior to analysis.Oven-dried (60 �C) and pulverized samples of mite feces wereanalyzed using py-gc/ms following the procedures outlined above.Analyzing fecal material chemistry allowed us to compare chem-istries among the following: 1) initial undecomposed litter; 2) litterplus microbes after decomposition for 62 d; 3) litter plus microbesplus mites after 62 d of decomposition (mites were removed fromlitter prior to chemical analysis); and 4) mite feces after feeding onlitter. Juvenile S. moestus were present in live boxes after approxi-mately one month and wewere unable to separate their feces fromthat of the adults although their numbers were low relative toadults. We attempted to collect mite feces from oak for threemonths but were unable to obtain enough material for analysis.
2.3.2. Extracellular enzyme activityThe activity of seven extracellular enzymes was analyzed using
procedures outlined in Saiya-Cork et al. (2002) and Grandy et al.(2007). Briefly, the activities of N-acetyl-b-D-glucosaminidase(chitinase), b-glucosidase (cellulase), b-D-1,4-cellobiosidase (cellu-lase), tyrosine amino peptidase and acid phosphatase wereassessed in black, 96-well microplates using substrates containingthe synthetic fluorescing molecules methylumbelliferone (MUB)and methylcoumarin (MC). The activity of urease was measuredusing colorimetric methods outlined in Sinsabaugh et al. (2000)and modified for analysis in 96-well microplates. Finally, theactivity of the oxidative enzyme phenol oxidase was measuredin clear 96-well microplates using L-3,4-dihydroxyphenylalanine(L-DOPA). Litter suspensions were prepared using 0.25 g subsam-ples from eachmicrocosm in a kitchen blender with 50mM sodiumacetate buffer at the initial pH (6.5) of the litters used for theexperiment. After incubation at 14 �C (approximately 6 h for allhydrolases and 24 h for oxidases), enzyme activity was determinedusing a flourometer (355 nm excitation and 460 nm emission for allfluorescent substrates) and spectrophotometer (phenol oxidase450 nm, and urease 620 nm) (Thermo Fisher Flouroskan andMultiskan, Thermo Scientific, Hudson, NH). All data are presentedas potential activity (mmol of substrate g�1 of dry litter h�1).
We additionally examined the potential enzymatic activity ofS. moestus collected from corn litter during our live Berleseextractions to better understand chemical transformations of litterresulting from gut passage. Prior to conducting the enzyme assay,100 mites were washed to remove associated external microor-ganisms following a modification of the procedure outlined inSiepel (1990). Briefly, mites were soaked for 10 min in 3% H2O2,followed by 10 m in a 1:5 w/v solution of MnO2 and deionizedwater. Finally, all mites were rinsed in deionized water for 10 m.Twenty adult mites were crushed in 200 mL of sodium acetatebuffer solution using a glass pestle. Mite slurries were diluted to20 mL with additional buffer solution. All enzymes were assayedfollowing the same 96-well plate methods described for litter.Fluorometric assays, including N-acetyl-b-D-glucosaminidase, b-glucosidase, tyrosine amino peptidase and acid phosphatase, wereincubated for approximately 6 h and photometric assays wereincubated for 18 (urease) and 24 h (phenol oxidase). Five replicatesamples (each containing 20 randomly selected adult mites) wererun for all gut enzymes measured.
2.3.3. Aqueous C and NAt 13, 34 and 62 d of incubation, 0.5 g subsamples of litter from
the destructively harvestedmicrocosmswere shakenwith 50mL ofdeionized water for 1 h on a bench-top shaker (200 rpm). Allsamples were then passed through a Whatman GF/A filter and
filtrates were stored at �20 �C until further analysis. Bioavailablenitrate and ammonium were quantified in litter filtrates using photo-metric procedures (Sinsabaugh et al., 2000; Doane and Horwath,2003). Total organic C and N were also measured on filtratesusing a Shimadzu TOC-TN combustion analyzer (Shimadzu Scien-tific Instruments, Columbia, MD) and leaf litter C and N values weredetermined using an elemental analyzer (Costech ECS 4010, Cost-ech Analytical Technologies, Inc., Valencia, CA). Total organic N wasdetermined by subtracting nitrate and ammonium from total N.
2.4. Statistical analysis
Microbial respiration data were analyzed using mixed modelanalysis for repeated measures in SAS (PROC MIXED) with mitepresence/absence and time as fixed effects and block as a randomeffect. Extracellular enzymes were analyzed using two-way anal-ysis of variance in SAS (ANOVA) with mite presence/absence andtime as treatment factors. Nitrate, ammonium, total organicnitrogen, total organic carbon andmolecular chemical datawere allanalyzed using one-way analysis of variance in SAS (ANOVA). Littertypes were analyzed independently for both one- and two-wayanalyses.
3. Results
3.1. Mite numbers and observations of feeding activity
The number of mites per microcosm dropped from 20 to a meanof 15 in corn and 13 in oak within one week of the start of theexperiment. This number was maintained throughout the experi-ment in corn microcosms, however, the number of live mites in oaklitter dropped to a mean of 4.5 individuals g�1 of litter from 48 to62 d of incubation. Juvenile mites were observed in both corn andoak microcosms beginning on day 34 and were present in lownumbers for the remainder of the incubation. Daily observationsconfirmed that both adults and juveniles fed throughout theexperiment and the majority of individuals examined containedmultiple fecal boluses. Additionally, slide mounts of fecal materialduring the course of the experiment showed that boluses containedboth plant and microbial material.
3.2. Litter C and N dynamics
Corn and oak litter differed qualitatively at the start of theexperiment. Although corn litter had a higher proportion of ligninthan oak (38 vs. 14% of sample composition) it also had higherproportions of nitrogen bearing compounds (13 vs. 5%) and poly-saccharides (20 vs. 9%). Additionally, oak litter had a higherproportion of lipids (50 vs. 11%) and a higher C/N ratio than corn.CO2 flux was higher in corn than in oak microcosms for the dura-tion of the incubation (Fig. 1). Mites increased cumulative CO2production by 19% in corn (F1, 168 ¼ 23.00, P < 0.0001) and 17% inoak (F1, 168 ¼ 51.72, P < 0.0001). The C/N ratio of oak increasedthrough time regardless of whether or not S. moestus was present.In contrast, corn C/N ratios only increased during decomposition inthe absence of mites (Table 1).
Polysaccharide relative abundance increased in corn over 62 d ofdecomposition in the absence of mites (F2, 6¼ 11.01, P< 0.01) (Fig. 2top). This increasewas due primarily to the compound 2,3-dihydro-benzofuran and was not observed when mites were present(Appendix Table 1). Oak litter was dominated by lipids whichcomprised 40e50% of all identified compounds. Of the lipidspresent, 60e90% were long-chain (>C20) compounds. Unlike corn,the chemistry of oak litter was not affected by the presence of mites(Fig. 2 bottom).

0 6 12 18 24 30 36 42 48 54 600
5
10
15
20
25
days of incubation
Cum
ulat
ive
CO
2(m
g C
O2
g-1dr
y lit
ter)
corn oakw/ S. moestus
w/o S. moestus
Cumulative CO2
Fig. 1. Cumulative CO2 production from litter over the 62 d incubation. Data representmeans � SE.
K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358354
3.3. Fecal material chemistry
Mites that were fed corn litter generated feces that werechemically different from the original litter (Fig. 2 top). While therelative abundance of lignin was lower in feces than litter, that ofphenol was higher (Fig. 2 top). In addition, polysaccharide relativeabundance was significantly higher in feces than in both the initiallitter and in litter decomposed for 62 d with microbes and mites.Ratios of lignin to nitrogen, and lignin to polysaccharides, were alsolower in feces than in corn litter by approximately 88 and 87%respectively. Further, the relative abundance of five compoundsincluding one polysaccharide, a phenol, a compound of unknownorigin and two lignin derivatives differed between feces and litter(Appendix Table 2). We planned to also include feces from oak-fedmites in our analyses but were unable to gather an amount suffi-cient for analysis.
3.4. Enzymes e extracellular enzymes within litter
The presence of mites altered the activity of both hydrolytic andoxidative extracellular enzymes. In corn microcosms, the activitiesof urease (32 d, F5, 48 ¼ 2.88, P < 0.05), glucosidase (48 d, F5,48 ¼ 2.49, P < 0.05) and acid phosphatase (48 d F4, 40 ¼ 7.32,P < 0.05) were greater in the presence than absence of mites(Fig. 3). The presence of mites also resulted in a 30% increase in theactivity of phenol oxidase averaged across sample dates (F1,48 ¼ 8.20, P < 0.01) (Fig. 3). The most marked increase occurred inweek 3, when oxidase activity was approximately 2.5 times higherin microcosms with than without mites. Chitinase, cellobiosidaseand tyrosine amino peptidase activities in cornwere not affected bymites. In oak litter, the presence of mites stimulated glucosidase (F5,48 ¼ 3.48, P < 0.01) and chitinase (F5, 48 ¼ 3.64, P < 0.01) activity at48 and 62 d but suppressed chitinase, glucosidase and phosphataseactivity during the initial week of the experiment (Fig. 4). Phenol
Table 1Carbon (C) and nitrogen (N) content (mg g�1 dry mass) of corn and oak litter at timezero and after 62 d of incubation with and without mites (P<0.05).
62 days
Time zero w/ S. moestus w/o S. moestus
Corn N (mg) 14.33a 11.29b 10.43bC (mg) 312.39 323.38 327.14C:N 21.80a 29.10ab 31.56b
Oak N (mg) 7.48a 6.73b 6.85bC (mg) 352.68 343.15 349.33C:N 47.15a 50.97b 51.04b
oxidase activity was also marginally suppressed by the presence ofmites during sampling week 1 but was greater in microcosms withthan without mites during the following week (F5, 48 ¼ 2.50,P < 0.05). Urease, cellobiosidase and tyrosine amino peptidaseactivities were not affected by mites in oak litter.
3.5. Enzymes e enzymatic potential of S. moestus
Chitinase was the most active enzyme detected within S.moestus averaging 62.7 mmol h�1 g�1 dry mite biomass followed bycellulase, phosphatase and peptidase (44.8, 17 and 0.13 mmolh�1 g�1 dry mite biomass, respectively). Urease and phenol oxidaseactivities were not detected in S. moestus.
3.6. Aqueous C and N in microcosms
The presence of mites led to a significant increase in availablenitrate, ammonium and both dissolved organic C and N (Tables 2and 3). The effects of mites on ammonium concentrations peakedin corn litter at 34 d (4.9 fold increase) and in oak litter at 13 d(12.26 fold increase). In contrast, ammonium concentrations weregreatest in corn microcosms without mites at 13 d. Mites increasednitrate concentrations in corn litter at 34 d (35.19 times more thanin litter without mites), as well as dissolved organic C and N (1.3and 4.2 times more, respectively) (Table 3).
4. Discussion
The soil fauna strongly influences ecosystem processes but theeffects of individual species on C and N cycling and microbialactivity during decomposition remain contentious (Ayres et al.,2010). Further, the effects of soil animals on changes in litterchemistry, extracellular enzyme activities and other key ecosystemprocesses during decomposition are poorly understood but couldhave a substantial, long-term effect on soil C cycling dynamics. Wefound that the widely distributed and often abundant oribatid miteS. moestus: 1) increased the mineralization of C and N; 2) acceler-ated extracellular enzyme activities; 3) altered the molecularchemistry of decomposing litter; and 4) interacted with variation ininitial litter chemistry, indicating that the overall effect of S. moestuson ecosystem processes depends upon substrate quality.
4.1. C and N cycling and enzyme activities
Some microarthropods have been shown to stimulate Cmineralization (Kaneda and Kaneko, 2008) but studies focusing onoribatids are rare (c.f. Kaneko 1998). We observed that C mineral-ization in the presence of S. moestus increased by 19% in corn and17% in oak litter. Previous estimates suggest that the entire meso-faunal community directly accounts for approximately 5% of theCO2 released from soil during decomposition and mathematicalmodels predict that oribatids should have little impact onmicrobialC mineralization (Berg et al., 2001). Although our microcosms donot simulate field conditions with fluctuating environmentalconditions and substrate availability, our results indicate that somespecies of oribatid mites can strongly enhance mineralization rates(Siepel and Maaskamp, 1994). We also show that S. moestusincreases concentrations of both nitrate and ammonium indecomposing litter. The increases in inorganic N that we observedwere temporally variable but generally similar to or greater thanthose found in studies using collembolans or other soil animals(Ineson et al., 1982; Anderson et al., 1985; Bardgett and Chan,1999).The observed changes in C and N cycling are likely due to the directeffects of S. moestus on organic matter mineralization as well as toenhanced microbial enzyme activities and decomposition rates
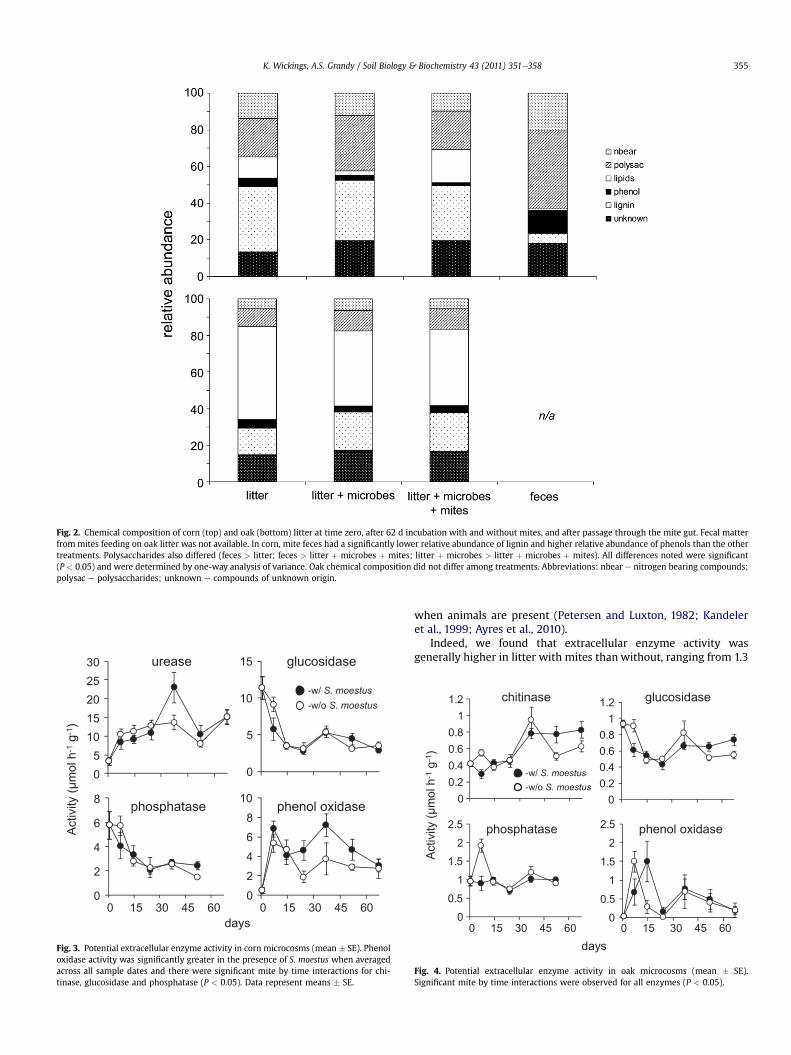
Fig. 2. Chemical composition of corn (top) and oak (bottom) litter at time zero, after 62 d incubation with and without mites, and after passage through the mite gut. Fecal matterfrom mites feeding on oak litter was not available. In corn, mite feces had a significantly lower relative abundance of lignin and higher relative abundance of phenols than the othertreatments. Polysaccharides also differed (feces > litter; feces > litter þ microbes þ mites; litter þ microbes > litter þ microbes þ mites). All differences noted were significant(P < 0.05) and were determined by one-way analysis of variance. Oak chemical composition did not differ among treatments. Abbreviations: nbear e nitrogen bearing compounds;polysac e polysaccharides; unknown e compounds of unknown origin.
0
5
10
15
0
2
4
6
8
05
1015202530
02468
10
urease glucosidase
phosphatase phenol oxidase
0 15 30 45 60 0 15 30 45 60
Activ
ity (µ
mol
h-1
g-1 )
days
-w/ S. moestus
-w/o S. moestus
Fig. 3. Potential extracellular enzyme activity in corn microcosms (mean � SE). Phenoloxidase activity was significantly greater in the presence of S. moestus when averagedacross all sample dates and there were significant mite by time interactions for chi-tinase, glucosidase and phosphatase (P < 0.05). Data represent means � SE.
K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358 355
when animals are present (Petersen and Luxton, 1982; Kandeleret al., 1999; Ayres et al., 2010).
Indeed, we found that extracellular enzyme activity wasgenerally higher in litter with mites than without, ranging from 1.3
00.20.40.60.8
11.2
00.20.40.60.8
11.2
00.5
11.5
22.5
00.5
11.5
22.5
chitinase glucosidase
phosphatase phenol oxidase
Activ
ity (µ
mol
h-1
g-1 )
days0 15 30 45 60 0 15 30 45 60
-w/ S. moestus
-w/o S. moestus
Fig. 4. Potential extracellular enzyme activity in oak microcosms (mean � SE).Significant mite by time interactions were observed for all enzymes (P < 0.05).
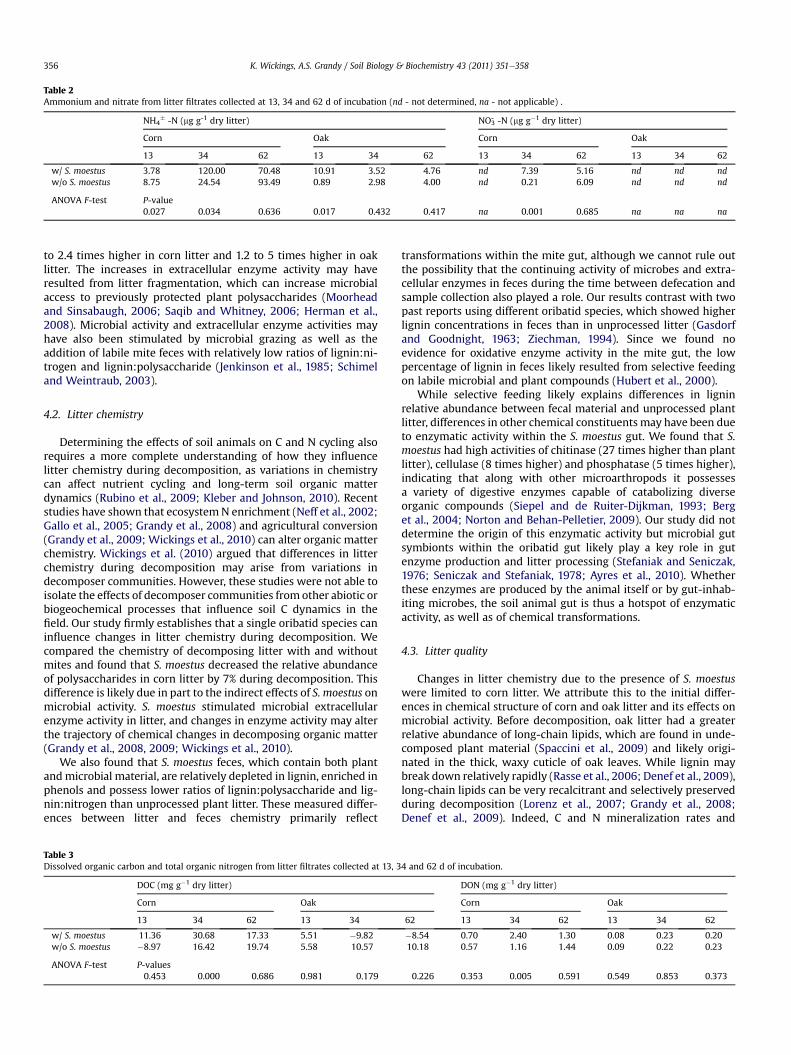
Table 2Ammonium and nitrate from litter filtrates collected at 13, 34 and 62 d of incubation (nd - not determined, na - not applicable) .
NH4� -N (mg g-1 dry litter) NO3
- -N (mg g�1 dry litter)
Corn Oak Corn Oak
13 34 62 13 34 62 13 34 62 13 34 62
w/ S. moestus 3.78 120.00 70.48 10.91 3.52 4.76 nd 7.39 5.16 nd nd ndw/o S. moestus 8.75 24.54 93.49 0.89 2.98 4.00 nd 0.21 6.09 nd nd nd
ANOVA F-test P-value0.027 0.034 0.636 0.017 0.432 0.417 na 0.001 0.685 na na na
K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358356
to 2.4 times higher in corn litter and 1.2 to 5 times higher in oaklitter. The increases in extracellular enzyme activity may haveresulted from litter fragmentation, which can increase microbialaccess to previously protected plant polysaccharides (Moorheadand Sinsabaugh, 2006; Saqib and Whitney, 2006; Herman et al.,2008). Microbial activity and extracellular enzyme activities mayhave also been stimulated by microbial grazing as well as theaddition of labile mite feces with relatively low ratios of lignin:ni-trogen and lignin:polysaccharide (Jenkinson et al., 1985; Schimeland Weintraub, 2003).
4.2. Litter chemistry
Determining the effects of soil animals on C and N cycling alsorequires a more complete understanding of how they influencelitter chemistry during decomposition, as variations in chemistrycan affect nutrient cycling and long-term soil organic matterdynamics (Rubino et al., 2009; Kleber and Johnson, 2010). Recentstudies have shown that ecosystemN enrichment (Neff et al., 2002;Gallo et al., 2005; Grandy et al., 2008) and agricultural conversion(Grandy et al., 2009; Wickings et al., 2010) can alter organic matterchemistry. Wickings et al. (2010) argued that differences in litterchemistry during decomposition may arise from variations indecomposer communities. However, these studies were not able toisolate the effects of decomposer communities from other abiotic orbiogeochemical processes that influence soil C dynamics in thefield. Our study firmly establishes that a single oribatid species caninfluence changes in litter chemistry during decomposition. Wecompared the chemistry of decomposing litter with and withoutmites and found that S. moestus decreased the relative abundanceof polysaccharides in corn litter by 7% during decomposition. Thisdifference is likely due in part to the indirect effects of S. moestus onmicrobial activity. S. moestus stimulated microbial extracellularenzyme activity in litter, and changes in enzyme activity may alterthe trajectory of chemical changes in decomposing organic matter(Grandy et al., 2008, 2009; Wickings et al., 2010).
We also found that S. moestus feces, which contain both plantandmicrobial material, are relatively depleted in lignin, enriched inphenols and possess lower ratios of lignin:polysaccharide and lig-nin:nitrogen than unprocessed plant litter. These measured differ-ences between litter and feces chemistry primarily reflect
Table 3Dissolved organic carbon and total organic nitrogen from litter filtrates collected at 13, 3
DOC (mg g�1 dry litter)
Corn Oak
13 34 62 13 34
w/ S. moestus 11.36 30.68 17.33 5.51 �9.82w/o S. moestus �8.97 16.42 19.74 5.58 10.57
ANOVA F-test P-values0.453 0.000 0.686 0.981 0.179
transformations within the mite gut, although we cannot rule outthe possibility that the continuing activity of microbes and extra-cellular enzymes in feces during the time between defecation andsample collection also played a role. Our results contrast with twopast reports using different oribatid species, which showed higherlignin concentrations in feces than in unprocessed litter (Gasdorfand Goodnight, 1963; Ziechman, 1994). Since we found noevidence for oxidative enzyme activity in the mite gut, the lowpercentage of lignin in feces likely resulted from selective feedingon labile microbial and plant compounds (Hubert et al., 2000).
While selective feeding likely explains differences in ligninrelative abundance between fecal material and unprocessed plantlitter, differences in other chemical constituents may have been dueto enzymatic activity within the S. moestus gut. We found that S.moestus had high activities of chitinase (27 times higher than plantlitter), cellulase (8 times higher) and phosphatase (5 times higher),indicating that along with other microarthropods it possessesa variety of digestive enzymes capable of catabolizing diverseorganic compounds (Siepel and de Ruiter-Dijkman, 1993; Berget al., 2004; Norton and Behan-Pelletier, 2009). Our study did notdetermine the origin of this enzymatic activity but microbial gutsymbionts within the oribatid gut likely play a key role in gutenzyme production and litter processing (Stefaniak and Seniczak,1976; Seniczak and Stefaniak, 1978; Ayres et al., 2010). Whetherthese enzymes are produced by the animal itself or by gut-inhab-iting microbes, the soil animal gut is thus a hotspot of enzymaticactivity, as well as of chemical transformations.
4.3. Litter quality
Changes in litter chemistry due to the presence of S. moestuswere limited to corn litter. We attribute this to the initial differ-ences in chemical structure of corn and oak litter and its effects onmicrobial activity. Before decomposition, oak litter had a greaterrelative abundance of long-chain lipids, which are found in unde-composed plant material (Spaccini et al., 2009) and likely origi-nated in the thick, waxy cuticle of oak leaves. While lignin maybreak down relatively rapidly (Rasse et al., 2006; Denef et al., 2009),long-chain lipids can be very recalcitrant and selectively preservedduring decomposition (Lorenz et al., 2007; Grandy et al., 2008;Denef et al., 2009). Indeed, C and N mineralization rates and
4 and 62 d of incubation.
DON (mg g�1 dry litter)
Corn Oak
62 13 34 62 13 34 62
�8.54 0.70 2.40 1.30 0.08 0.23 0.2010.18 0.57 1.16 1.44 0.09 0.22 0.23
0.226 0.353 0.005 0.591 0.549 0.853 0.373

K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358 357
enzyme activities, all indicators of microbial activity, were lower inoak litter. This provides additional evidence that oak litter wasmore recalcitrant than corn litter, but also points to the probabilitythat the effects of decomposer communities on litter chemicaltransformations may depend upon the quality of the substratebeing decomposed (Kampichler and Bruckner, 2009; Wickingset al., 2010).
4.4. Summary
Soil animals are broadly known to have a range of effects ondecomposition dynamics. However, at a finer-scale there remainmany key uncertainties, including the effects of individual specieson decomposition processes, and the degree to which animals canalter the chemistry of litter during decomposition (Ayres et al.,2010). We have demonstrated that the presence of a single orib-atid mite species can enhance C mineralization rates by almost 20%,stimulate hydrolytic and oxidative enzyme activities and changelitter chemistry during decomposition. No other studies have cometo our attention that show molecular-scale differences in thechemistry of oribatid feces compared to litter, or of litter decom-posing with and without mites. These changes in litter chemistryduring decomposition varied with initial substrate quality andcould have long-term implications for soil C cycling. Future studiescomparing different oribatid mite species, different groups ofmicroarthropods and different trophic levels under more realisticconditions will help elucidate the effects of individual species andfunctional groups on decomposition.
Acknowledgements
The authors thank Roy Norton, SUNY Environmental Scienceand Forestry, Syracuse, NY for assistance in identification of Sche-loribates moestus to species and for advice on the maintenance ofmite populations in the laboratory. We also thank David Weed andSteve Hamilton at the Kellogg Biological Station LTER, HickoryCorners, MI for their assistance in the measurement of dissolvedorganic carbon and nitrogen from litter filtrates. Roy Norton andDeborah Neher provided insightful comments on earlier drafts ofthis manuscript.
Appendix. Supplementary data
Supplementary data related to this article can be found online atdoi:10.1016/j.soilbio.2010.10.023.
References
Anderson, J.M., Leonard, M.A., Ineson, P., Huish, S.A., 1985. Faunal biomass: a keycomponent of a general model of nitrogen mineralization. Soil Biology &Biochemistry 17, 735e737.
Ayres, E., Wall, D.H., Bardgett, R.D., 2010. Trophic interactions and their implicationsfor soil carbon fluxes. In: Kutsch, W.L., Bahn, M., Heinemeyer, A. (Eds.), SoilCarbon Dynamics: an Integrated Methodology. Cambridge University Press,Cambridge UK, pp. 187e206.
Bardgett, R.D., Chan, K.F., 1999. Experimental evidence that soil fauna enhancenutrient mineralization and plant nutrient uptake in montane grasslandecosystems. Soil Biology & Biochemistry 31, 1007e1014.
Bardgett, R.D., Keiller, S., Cook, R., Gilburn, A.S., 1998. Dynamic interactions betweensoil animals and microorganisms in upland grassland soils amended with sheepdung: a microcosm experiment. Soil Biology & Biochemistry 30, 531e539.
Berg, M.P., de Ruiter, P., Didden, W., Janssen, M., Schouten, T., Verhoef, H., 2001.Community food web, decomposition and nitrogen mineralisation in a strati-fied Scots pine forest soil. Oikos 94, 130e142.
Berg, M.P., Stoffer, M., van den Heuvel, H.H., 2004. Feeding guilds of Collembolabased on digestive enzymes. Pedobiologia 48, 589e601.
Coleman, D.C., 2008. From peds to paradoxes: linkages between soil biota and theirinfluences on ecological processes. Soil Biology & Biochemistry 40, 271e289.
Coleman, D.C., Crossley Jr., D.A., Hendrix, P.F., 2004. Fundamentals of Soil Ecology,second ed. Elsevier Academic Press, Burlington, MA.
Cortet, J., Joffre, R., Elmholt, S., Krogh, P.H., 2003. Increasing species and trophicdiversity of mesofauna affects fungal biomass, mesofauna community structureand organic matter decomposition processes. Biology and Fertility of Soils 37,302e312.
Denef, K., Plante, A.F., Six, J., 2009. Characterization of soil organic matter. In:Kutsch, W.L., Bahn, M., Heinemeyer, A. (Eds.), Soil Carbon Dynamics: anIntegrated Methodology. Cambridge University Press, Cambridge UK, pp.91e126.
Doane, T.A., Horwath, W.A., 2003. Spectrophotometric determination of nitrate witha single reagent. Analytical Letters 36, 2713e2722.
Ekschmitt, K., Liu, M., Vetter, S., Fox, O., Wolters, V., 2008. Strategies used by soilbiota to overcome soil organic matter stability ewhy is dead organic matter leftover in the soil? Geoderma 128, 167e176.
Filley, T.R., McCormick, M.K., Crow, S.E., Szlaveck, K., Whigham, D.F., Johnston, C.T.,van den Heuvel, R.N., 2008. Comparison of the chemical alteration trajectory ofLiriodendron tulipifera L. leaf litter among forests with different earthwormabundance. Journal of Geophysical Research 113, 1e14.
Gallo, M.E., Lauber, C.L., Cabaniss, S.E., Waldrop, M.P., Sinsabaugh, R.L., Zak, D.R.,2005. Soil organic matter and litter chemistry response to experimental Ndeposition in northern temperate deciduous forest ecosystems. Global ChangeBiology 11, 1514e1521.
Gasdorf, E.C., Goodnight, C.J., 1963. Studies on the ecology of soil arachnids. Ecology44, 261e268.
Geib, S.M., Filley, T.R., Hatcher, P.G., Hoover, K., Carlson, J.E., Jiminez-Gasco, M.,Nakagawa-Izumi, A., Sleighter, R.L., Tien, M., 2008. Lignin degradation in wood-feeding insects. Proceedings of the National Academy of Sciences 105,12932e12937.
Gergócs, V., Hufnagel, L., 2009. Application of oribatid mites as indicators (review).Applied Ecology and Environmental Research 7, 79e98.
Gillon, D., David, J.F., 2001. The use of near infrared reflectance spectroscopy tostudy chemical changes in the leaf litter consumed by saprophagous inverte-brates. Soil Biology & Biochemistry 33, 2159e2161.
Golchin, A., Oades, J.M., Skjemstad, J.O., Clarke, P., 1994. Study of free and occludedparticulate organic-matter in soils by solid-state C-13 CP/MAS NMR-spectros-copy and scanning electron-microscopy. Australian Journal of Soil Research 32,285e309.
Grandy, A.S., Sinsabaugh, R., Neff, J., Stursova, M., Zak, D., 2008. Nitrogen depositioneffects on soil organic matter chemistry are linked to variation in enzymes,ecosystems and size fractions. Biogeochemistry 91, 37e49.
Grandy, A.S., Neff, J.C., Weintraub, M.N., 2007. Carbon structure and enzymeactivities in alpine and forest ecosystems. Soil Biology & Biochemistry 39,2701e2711.
Grandy, A.S., Strickland, M.S., Lauber, C.L., Bradford, M.A., Fierer, N., 2009. Theinfluence of microbial communities, management, and soil texture on soilorganic matter chemistry. Geoderma 150, 278e286.
Guggenberger, G., Thomas, R.J., Zech, W., 1996. Soil organic matter within earth-worm casts of an anecic-endogeic tropical pasture community. ColombiaApplied Soil Ecology 3, 263e274.
Hanlon, R.D.G., Anderson, J.M., 1979. The effects of Collembola grazing on microbialactivity in decomposing leaf litter. Oecologia 38, 93e99.
Hansen, R.A., 1999. Red oak litter promotes a microarthropod functional group thataccelerates its decomposition. Plant and Soil 209, 37e45.
Hedde, M., Bureau, F., Akpa-Vinceslas, M., Aubert, M., Decaens, T., 2007. Beech leafdegradation in laboratory experiments: effects of eight detritivorous inverte-brate species. Applied Soil Ecology 35, 291e301.
Heneghan, L., Coleman, D.C., Zou, X., Crossley, D.A., Haines, B.L., 1999. Soil micro-arthropod contributions to decomposition dynamics: tropical-temperatecomparisons of a single substrate. Ecology 80, 1873e1882.
Herman, J., Moorhead, D., Berg, B., 2008. The relationship between rates of ligninand cellulose decay in aboveground forest litter. Soil Biology & Biochemistry 40,2620e2626.
Hopkins, D.W., Chudek, J.A., Bigness, D.E., Frouz, J., Webster, E.A., Lawson, T., 1998.Application of 13C NMR to investigate the transformations and biodegradationof organic materials by wood- and soil-feeding termites, and a coprophagouslitter-dwelling dipteran larva. Biodegradation 9, 423e431.
Hubert, J., Kubatova, A., Sarova, J., 2000. Feeding of Scheloribates laevigatus (Acari:Oribatida) on different stadia of decomposing grass litter (Holcus lanatus).Pedobiologia 44, 627e639.
Ineson, P., Leonard, M.A., Anderson, J.M., 1982. Effect of collembolan grazing uponnitrogen and cation leaching from decomposing leaf litter. Soil Biology andBiochemistry 14, 601e605.
Jenkinson, D.S., Fox, R.H., Rayner, J.H., 1985. Interactions between fertilizer nitrogenand soil-nitrogen e the so-called priming effect. Journal of Soil Science 36,425e444.
Kampichler, C., Bruckner, A., 2009. The role of microarthropods in terrestrialdecomposition: a meta-analysis of 40 years of litterbag studies. BiologicalReviews 84, 375e389.
Kaneda, S., Kaneko, N., 2008. Collembolans feeding on soil affect carbon andnitrogen mineralization by their influence on microbial and nematode activi-ties. Biology and Fertility of Soils 44, 435e442.
Kandeler, E., Kampichler, C., Joergensen, R.G., Mölter, K., 1999. Effects of mesofaunain a spruce forest on soil microbial communities and N cycling in field meso-cosms. Soil Biology and Biochemistry 31, 1783e1792.
Kaneko, N., 1998. Do mites and Collembola affect pine litter fungal biomass andmicrobial respiration? Applied Soil Ecology 9, 209e213.

K. Wickings, A.S. Grandy / Soil Biology & Biochemistry 43 (2011) 351e358358
Kleber, M., Johnson, M.G., 2010. Advances in understanding the molecular structureof soil organic matter: implications for interactions in the environment.Advances in Agronomy 106, 77e142.
Kleber, M., Sollins, P., Sutton, R., 2007. A conceptual model of organo-mineralinteractions in soils: self-assembly of organic molecular fragments into zonalstructures on mineral surfaces. Biogeochemistry 85, 9e24.
Lenoir, L., Persson, T., Bengtsson, J., Wallander, H., Wiren, A., 2007. Bottom-upand top-down control in forest soil microcosms? Effects of soil fauna onfungal biomass and C/N mineralization. Biology and Fertility of Soils 43,281e294.
Lorenz, K., Lal, R., Preston, C.M., Nierop, K.G.J., 2007. Strengthening the soil organiccarbon pool by increasing contributions from recalcitrant aliphatic bio (macro)molecules. Geoderma 142, 1e10.
Moorhead, D.L., Sinsabaugh, R.L., 2006. A theoretical model of litter decay andmicrobial interaction. Ecological Monographs 76, 151e174.
Neff, J.C., Townsend, A.R., Gleixner, G., Lehman, S.J., Turnbull, J., Bowman, W.D.,2002. Variable effects of nitrogen additions on the stability and turnover of soilcarbon. Nature 419, 915e917.
Norton, R.A., Behan-Pelletier, V.M., 2009. Oribatida. In: Krantz, G.W., Walter, D.E.(Eds.), A Manual of Acarology. Texas Tech Univ. Press, Lubbock, TX, pp. 430e564.
Petersen, H., Luxton, M., 1982. A Comparative analysis of soil fauna populations andtheir role in decomposition processes. Oikos 39, 287e388.
Rasse, D.P., Dignac, M.F., Bahri, H., Rumpel, C., Mariotti, A., Chenu, C., 2006. Ligninturnover in an agricultural field: from plant residues to soil-protected fractions.European Journal of Soil Science 57, 530e538.
Rawlins, A.J., Bull, I.D., Poirier, N., Ineson, P., Evershed, R.P., 2006. The biochemicaltransformation of oak (Quercus robur) leaf litter consumed by the pill millipede(Glomeris marginata). Soil Biology & Biochemistry 38, 1063e1076.
Rubino, M., Lubritto, C., D’Onofrio, A., Terrasi, F., Kramer, C., Gleixner, G.,Cotrufo, M.F., 2009. Isotopic evidences for microbiologically mediated anddirect C input to soil compounds from three different leaf litters during theirdecomposition. Environmental Chemistry Letters 7, 85e95.
Saiya-Cork, K.R., Sinsabaugh, R.L., Zak, D.R., 2002. The effects of long term nitrogendeposition on extracellular enzyme activity in an Acer saccharum forest soil.Soil Biology & Biochemistry 34, 1309e1315.
Saqib, A.A.N., Whitney, P.J., 2006. Role of fragmentation activity in cellulosehydrolysis. International Biodeterioration and Biodegradation 58, 180e185.
Schimel, J.P., Weintraub, M.N., 2003. The implications of exoenzyme activity onmicrobial carbon and nitrogen limitation in soil: a theoretical model. SoilBiology & Biochemistry 35, 1e15.
Seniczak, S., Stefaniak, O., 1978. The microflora of the alimentary canal of Oppianitens (Acarina, Oribatei). Pedobiologia 18, 110e119.
Siepel, H., 1990. Niche relationships between two panphytophagous soil mites,Nothrus silvestris Nicolet (Acari, Oribatida, Nothridae) and Platynothrus peltifer(Koch) (Acari, Oribatida, Camisiidae)*. Biology and Fertility of Soils 9, 139e144.
Siepel, H., de Ruiter-Dijkman, E.M., 1993. Feeding guilds of oribatid mites based ontheir carbohydrase activities. Soil Biology & Biochemistry 25, 1491e1497.
Siepel, M., Maaskamp, F., 1994. Mites of different feeding guilds affect decomposi-tion of organic matter. Soil Biology & Biochemistry 26, 1389e1394.
Sinsabaugh,R.L., Reynolds, H., Long, T.M., 2000. Rapidassay for amidohydrolase (urease)activity in environmental samples. Soil Biology & Biochemistry 32, 2095e2097.
Spaccini, R., Sannino, D., Piccolo, A., Fagnano, M., 2009. Molecular changes in organicmatter of a compost-amended soil. European Journal of Soil Science 60, 287e296.
Stefaniak, O., Seniczak, S., 1976. The microflora of the alimentary canal of Achipteriacoleoptrata (Acarina, Oribatei). Pedobiologia 16, 185e194.
Teuben, A., Verhoef, H.A., 1992. Direct contribution by soil arthropods to nutrientavailability through body and fecal nutrient content. Biology and Fertility ofSoils 14, 71e75.
Wickings, K., Grandy, A.S., Reed, S.C., Cleveland, C.C., 2010. Management intensityalters decomposition via biological pathways. Biogeochemistry. doi:10.1007/s10533-010-9510-x.
Wolters, V., 2000. Invertebrate control of soil organic matter stability. Biology andFertility of Soils 31, 1e19.
Ziechman, W., 1994. Humic Substances. Wissenschaftsverlag, Mannheim.





























