Embed Size (px)
Citation preview

MADM, a Novel Adaptor Protein That Mediates Phosphorylation ofthe 14-3-3 Binding Site of Myeloid Leukemia Factor 1*
Received for publication, June 18, 2002, and in revised form, July 30, 2002Published, JBC Papers in Press, August 9, 2002, DOI 10.1074/jbc.M206041200
Raelene Lim‡§, Louise N. Winteringham‡¶, James H. Williams‡¶, Ross K. McCulloch‡,Evan Ingley‡¶, Jim Y-H. Tiao‡¶, Jean-Philippe Lalonde‡¶, Schickwann Tsai�, Peta A. Tilbrook‡¶,Yi Sun**, Xiaohua Wu**, Stephan W. Morris**‡‡, and S. Peter Klinken‡¶§§
From the ‡Laboratory for Cancer Medicine, Medical Research Foundation, Royal Perth Hospital, Western AustralianInstitute for Medical Research, Perth, Western Australia 6000, the §School of Biomedical Sciences, Curtin University ofTechnology, Bentley, Western Australia 6102, the ¶Department of Biochemistry, The University of Western Australia,Nedlands, Australia 6009, the Departments of **Pathology and ‡‡Hematology-Oncology, St. Jude Children’s ResearchHospital, Memphis, Tennessee 38105, and �The Institute for Gene Therapy and Molecular Medicine, The Mount SinaiSchool of Medicine, New York, New York 10029
A yeast two-hybrid screen was conducted to identifybinding partners of Mlf1, an oncoprotein recently iden-tified in a translocation with nucleophosmin that causesacute myeloid leukemia. Two proteins isolated in thisscreen were 14-3-3� and a novel adaptor, Madm. Mlf1contains a classic RSXSXP sequence for 14-3-3 bindingand is associated with 14-3-3� via this phosphorylatedmotif. Madm co-immunoprecipitated with Mlf1 and co-localized in the cytoplasm. In addition, Madm recruiteda serine kinase, which phosphorylated both Madm andMlf1 including the RSXSXP motif. In contrast to wild-type Mlf1, the oncogenic fusion protein nucleophosmin(NPM)-MLF1 did not bind 14-3-3�, had altered Madmbinding, and localized exclusively in the nucleus.Ectopic expression of Madm in M1 myeloid cells sup-pressed cytokine-induced differentiation unlike Mlf1,which promotes maturation. Because the Mlf1 bindingregion of Madm and its own dimerization domain over-lapped, the levels of Madm and Mlf1 may affect complexformation and regulate differentiation. In summary,this study has identified two partner proteins of Mlf1that may influence its subcellular localization and bio-logical function.
Myeloid leukemia factor 1 (MLF1)1 is a recently describedoncogene involved in acute myeloid leukemia and myelodys-
plastic syndrome (1). It was initially identified in a t(3;5)(q25.1;q34) translocation between MLF1 on chromosome 3 and nu-cleophosmin (NPM) on chromosome 5, which generates thechimeric molecule NPM-MLF1 (1). Significantly, overexpres-sion of wild-type MLF1 in acute myeloid leukemias, not involv-ing the t(3;5), is associated with progression toward a malig-nant phenotype (2).
The murine homologue of MLF1 (Mlf1) (3) was isolated in-dependently as a gene expressed when J2E erythroleukemiacells (4) undergo a spontaneous phenotypic change to displaymorphological and biochemical features of monocytoid cells (5);therefore, the gene was termed Hls7 for hemopoietic lineageswitch gene 7 (6). Significantly, enforced expression of Mlf1/Hls7 in the parental erythroleukemia cells recapitulated theoriginal phenotypic change and prevented erythropoietin-in-duced maturation (6). In addition, ectopic expression of Mlf1/Hls7 in normal hemopoietic progenitors enhanced myeloid col-ony formation at the expense of erythroid colonies. From thesestudies, it was concluded that Mlf1 is normally involved in thephenotypic determination of hemopoietic cells and that its dys-regulation is important in leukemogenesis.
Analysis of the Mlf1 sequence identified no recognizablemotifs or domains, except for a characteristic RSXSXP bindingsite for 14-3-3 proteins (3, 6). Proteins that associate with14-3-3 family members are phosphorylated on serine residueswithin the RSXSXP motif (7, 8). This serine phosphorylationappears essential for 14-3-3 binding, although other non-canonical binding sites have also been identified in moleculessuch as Cbl (9) and p53 (10). The 14-3-3 family of proteins isexpressed in a broad range of organisms and is highly conservedthroughout evolution. These molecules bind and influence theactivity of numerous diverse proteins, including Raf, BAD,Cdc25c, p53, Cbl, Bcr, and Bcr-Abl (10–15); as a consequence,they have been implicated in regulating the onset of cell division,apoptosis, and differentiation. 14-3-3 proteins control the activityof their partner molecules, in part, by sequestering them fromtheir site of action and by serving as bridging molecules thatpromote dimerization of associated proteins (16).
In an attempt to decipher the biological function of Mlf1, ayeast two-hybrid screen was conducted to identify moleculesthat associate with the protein. Predictably, a 14-3-3 familymember (14-3-3�) was shown to associate with Mlf1 in thisscreen. In addition, a novel adaptor molecule (Madm, MLF1-adaptor molecule) was isolated. Madm associates with a kinasethat phosphorylates serine residues in Madm itself and the
* This work was supported in part by National Health and MedicalResearch Council (NHMRC) Grant 110298 (to S. P. K.), Cancer Foun-dation of Western Australia, Medical Research Foundation at RoyalPerth Hospital, NCI National Institutes of Health Grant CA76301 (toS. W. M.), CORE Grant CA21765, and by the American Lebanese Syr-ian Associated Charities (ALSAC), St. Jude Children’s Research Hos-pital. The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
The nucleotide sequences reported in this paper for the Mlf1 adaptormolecule (Madm) gene and mRNA have been submitted to the Gen-BankTM Data Bank with accession numbers AF302139 and AF302138,respectively.
The amino acid sequence of this protein can be accessed through NCBIProtein Database under NCBI accession number AAK97260.
§§ To whom correspondence should be addressed: Laboratory for Can-cer Medicine, WAIMR, Level 6 MRF Bldg., Rear 50 Murray St., PerthWA 6000, Australia. Tel.: 61-8-9224-0333; Fax: 61-8-9224-0322; E-mail:[email protected].
1 The abbreviations used are: MLF, myeloid leukemia factor; NPM,nucleophosmin; Madm, Mlf1 adaptor molecule; LIF, leukemia inhibi-tory factor; NLS, nuclear localization signal; NES, nuclear localizationsignal; SH, Src homology.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 43, Issue of October 25, pp. 40997–41008, 2002© 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 40997
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

RSXSXP motif of Mlf1. These data indicate that Madm isinvolved in 14-3-3-mediated regulation of Mlf1.
EXPERIMENTAL PROCEDURES
Yeast Two-hybrid Analysis and Isolation of Madm cDNA Clones—The Saccharomyces cerevisiae strain, L40 (MATa,his3�200,trp1–901,leu2–3,112,ade2,LYS2::(lexAop)4-HIS3,URA::(lexAop)8-lacZ,GAL4) was used in a yeast two-hybrid procedure essentially asdescribed by Vojtek et al. (17). The complete Mlf1 open reading framewas subcloned into pBTM116 (17) to generate a LexA DNA-bindingdomain fusion. The L40 strain was transformed with pBTM116-Mlf1and used to screen a pVP16 cDNA library made from mRNA preparedfrom the lymphohemopoietic progenitor cell line, EML C.1 (18). AnEML C.1 cDNA library in �zapII (Stratagene, La Jolla, CA) wasscreened for full-length Madm using the 220-bp Madm yeast two-hybridfragment as a probe. After a full-length clone was isolated (GenBankTM
accession number AF302138), databases (GenBank/EMBL/DDBJ) weresearched using FASTA and BLAST programs of the Genetics ComputerGroup (Madison, WI) software package.
Immunoprecipitation and Immunoblotting—A glutathione S-trans-ferase fusion protein consisting of the first 55 amino acids of Madm wasexpressed in Escherichia coli using pGEX-2T (19). The fusion proteinwas purified, and antisera were raised in rabbits. COS cells weretransiently transfected with pcDNA3(�) (Invitrogen, Carlsbad, CA)constructs expressing Mlf1 and/or Madm using LipofectAMINE 2000(Invitrogen, Carlsbad, CA) and harvested 24 h posttransfection. Proteinlysates were prepared using 1% Nonidet P-40, 0.5% deoxycholic acid,150 mM sodium chloride, 50 mM Tris-HCl (pH 8.0), 2 mM EDTA, 2 mM
EGTA, 10 mM sodium fluoride, 1 mM sodium vanadate, 1 mM benzami-dine, 1 mM phenylmethylsulfonyl fluoride, and 1 �g/ml aprotinin. Im-munoprecipitations were performed with 0.5 mg of protein lysate withthe indicated antibodies and protein G-Sepharose (Sigma). Immunopre-cipitated proteins were separated by SDS-PAGE, transferred to nitro-cellulose membranes, and probed with specific antibodies before detec-tion using enhanced chemiluminescence (Amersham Biosciences).Subcellular fractionation studies were performed essentially as de-scribed by Zhang et al. (20).
Peptide Binding Assay—Synthetic peptides corresponding to theRSXSXP motif of MLF1 were synthesized (26MRQMIRSFSEPF-GRDL41) with the second serine residue (Ser34) either phosphorylatedor nonphosphorylated. Peptides conjugated to Sepharose beads wereincubated with cell lysates before separation by SDS-PAGE andimmunoblotting.
Confocal Microscopy—Indirect immunofluorescence was performedon transiently transfected COS cells fixed with acetone:methanol (1:1).Protein localization was detected using antibodies against Madm, Mlf1,or a Myc epitope tag, followed by an anti-IgG Alexa-Fluor 488 secondaryantibody or an anti-IgG Alexa-Fluor 546 secondary antibody (MolecularProbes, Eugene, OR). Alternatively, cells were transfected with apEGFP-C2 plasmid containing Madm (Clontech, Palo Alto, CA), whichexpressed Madm with green fluorescent protein fused at the N terminusof the molecule. DNA was counterstained with Hoechst 33258 andvisualized on an MRC 1024 UV laser scanning confocal microscope(Bio-Rad, Hercules, CA).
In Vitro Kinase Assay, in Vivo Phosphorylation, and [32P]Phos-phoamino Acid Analysis—Kinase assays were performed on Madmimmunoprecipitates essentially as described previously (21). Sampleswere incubated for 30 min at 37 °C in the presence of 0.4 �Ci/�l[�-32P]ATP (3000 Ci/mmol), separated by SDS-PAGE, and transferredto polyvinylidene difluoride membranes, and radioactivity was detectedusing a PhosphorImager (Molecular Dynamics, Sunnyvale, CA). Phos-phorylation of the MLF1-S34 peptide (MRQMIRSFSEPFGRDL) byMadm immunoprecipitates was detected as described above. To deter-
mine phosphorylation status in vivo, transiently transfected COS cellswere incubated with 100 �Ci/well of phosphoric acid (H3
32PO4). Thecells were lysed, and protein immunoprecipitated using specific anti-bodies. For [32P]phosphoamino acid analysis, in vitro kinase assayswere performed, and proteins transferred to polyvinylidene difluorideafter separation by SDS-PAGE. Radiolabeled Madm or phosphorylatedsubstrates were excised, hydrolyzed, and analyzed by one-dimensionalthin-layer chromatography (22).
Infection and Analysis of M1 Cell Differentiation—The PA317 pack-aging cell line (23) was transfected with linearized constructs of theMSCV-neo 2.2 vector (24) alone or MSCV expressing Madm with aC-terminal Myc tag. M1 cells were then infected with the amphotrophicviral supernatants as described previously (21). Expression of exoge-nous Madm transcripts was determined by Northern blotting, whileimmunoblotting was employed to detect Myc-tagged Madm. Differenti-ation of M1 cells was initiated by addition of leukemia inhibitory factor(LIF) and monitored for Cd11b surface expression by flow cytometry(Beckman-Coulter Epics XL/MCL) using a fluorescein isothiocyanate-conjugated antibody (Chemi-Con, Temecula, CA).
RESULTS
Mlf1 Associates with 14-3-3�—In an attempt to identify pro-teins that interact with Mlf1, a yeast two-hybrid screen wasperformed. Table I shows that Mlf1 interacted with numerousmolecules, including a member of the 14-3-3 family of proteinsviz 14-3-3�, which was isolated as two separate clones. Othermolecules that associated with Mlf1 in this screen includedeukaryotic translation initiation factor 3 (eIF3, p42 subunit),protein inhibitor of neuronal nitric-oxide synthase (PIN, alsoknown as dynein light chain, LC8), calcyclin-binding protein,nucleolin, and several uncharacterized proteins; each of thesewere identified once in the screen.
To confirm the interaction between Mlf1 and 14-3-3� bio-chemically, COS cells were transfected with an Mlf1-express-ing construct since endogenous Mlf1 protein expression is be-low the limits of detection. Fig. 1A shows that when lysateswere immunoprecipitated using anti-14-3-3� antibodies, Mlf1specifically co-purified with endogenous 14-3-3�. In addition,Mlf1 co-immunoprecipitated with 14-3-3� in J2E erythroleuke-mia cells expressing retrovirally driven Mlf1 (data not shown).These results corroborated the interaction between Mlf1 and14-3-3� in the yeast two-hybrid screen.
Both murine and human MLF1 contain a classic RSXSXPmotif for 14-3-3 binding (1, 3, 6). To determine if this regionmediates the interaction between MLF1 and 14-3-3�, two pep-tides spanning this motif were generated (26MRQMIRSFSEPF-GRDL41) with serine 34 either phosphorylated or non-phospho-rylated. Data displayed in Fig. 1B show that endogenous andrecombinant 14-3-3� associated with the phosphorylated pep-tide only. Moreover, only the phosphorylated peptide was ableto disrupt the MLF1/14-3-3 complex (Fig. 1C). Taken together,these results demonstrate that MLF1 and 14-3-3� associate viathe RSXSXP motif of MLF1 and that phosphorylation is essen-tial for this interaction.
The binding of 14-3-3� to the fusion protein NPM-MLF1 wasalso investigated. Immunoprecipitation experiments from COScells expressing FLAG-tagged NPM-MLF1 demonstrated that
TABLE IMlf1-interacting proteins identified in a yeast two-hybrid screen
Clone Homologous to: GenBank™
M51 14-3-3� (100%) BAA11751M4 Dynein light chain 1 (LC8), cytoplasmic protein inhibitor of neuronal
nitric oxide synthase (PIN) (100%)BAB27516
M28 Eukaryotic translation initiation factor 3 (p42 subunit) (100%) AAF14221M86 Calcyclin-binding protein (100%) AAC16757M62 Nucleolin (100%) NP-035010M58 Similar to heterogenous nuclear ribonucleoprotein U; scaffold attachment
factor A; nuclear matrix protein (82%)XP-058496
M2, M44, M70, M79, M86 Novel
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs40998
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

NPM-MLF1 did not bind 14-3-3� (Fig. 1D) or that the interac-tion was extremely weak. Therefore, despite retaining the RSX-SXP motif, the leukemogenic fusion protein NPM-MLF1 dif-fered biochemically from the wild-type MLF1. These datasuggest that the inability of NPM-MLF1 to associate with14-3-3 may occur due to conformational changes produced bythe fusion of NPM to MLF1.
Mlf1 Associates with a Novel Adaptor Protein—One of thenovel clones that associated with Mlf1 in the yeast two-hybridscreen (Clone M44) contained a 220-bp cDNA fragment, which
had no homology to sequences in the database at the time ofisolation and was named Madm for Mlf1-adaptor molecule.This fragment was used to screen an EML C.1 cDNA library,and a 2153-bp cDNA was isolated. An open reading frame of1608 bp was identified, encoding a putative 535-amino acidprotein with a predicted molecular mass of 60 kDa (Fig. 2A).The probable initiating ATG was in the characteristic contextfor a translation start site (25).
A comparison of the amino acid sequence encoded by Madmusing the Swiss-Prot data base revealed similarities with a
FIG. 1. MLF1 binds to 14-3-3�. A, ly-sates from COS cells transfected withMlf1 were immunoprecipitated (IP) withanti-14-3-3� and immunoblotted (IB) withanti-Mlf1 and anti-14-3-3�. B, phospho-rylated (RSFSPEP) and non-phosphoryl-ated (RSFSEP) peptides were immobi-lized on beads and incubated with lysatesfrom 32D cells expressing MLF1 or NPM-MLF1, COS cells, or recombinant 14-3-3protein (rec. 14-3-3). C, MLF1/14-3-3 com-plexes were immunoprecipitated fromCOS cells transfected with MLF1 usingan anti-MLF1 antibody, incubated in thepresence of phosphorylated or non-phos-phorylated peptide, and immunoblottedusing an anti-14-3-3� antibody. D, immu-noprecipitations using either anti-14-3-3or anti-FLAG antibodies were performedon COS cells transfected with FLAG-tagged NPM-MLF1 and then immuno-blotted with antibodies to MLF1 or14-3-3�.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs 40999
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

FIG. 2. M44 cDNA and protein sequence. A, the nucleotide sequence of murine Madm cDNA is shown with the predicted amino acidsindicated above. Nucleotides are numbered on the left and amino acids on the right. The in-frame termination codons in the 5� untranslated regionare underlined. The putative kinase domain is in black and the nuclear export sequence (NES, amino acids 121–129) and nuclear localizationsequence (NLS, amino acids 163–181) are light gray. The boxed sequence indicates the region identified in the Mlf1 yeast two-hybrid screen.Dashed lines indicate the two putative nuclear receptor-binding motifs (consensus, LXXLL). B, schematic representation of the Madm proteinshows the potential SH2-binding region, the kinase-like domain, and the Mlf1-binding domain. The genomic organization of the Madm gene isshown below with 18 exons identified by boxes.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs41000
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

variety of protein kinases, and a consensus kinase domain wasidentified (26). However, the conserved ATP-binding motif(GXGXXG), found in conventional protein kinases (27), was notpresent in this protein (Fig. 2). Intriguingly, a bipartite nuclearlocalization signal (NLS) was present within the kinase do-main, as well as a potential nuclear export signal (NES) (Fig.2), indicating that the protein may shuttle between the nucleusand cytoplasm. Moreover, Madm contains several potentialphosphorylation sites in the vicinity of both the NLS and NES.These include protein kinase C phosphorylation sites at Ser102,Ser157, and Thr165, as well as a casein kinase II phosphoryla-tion site at Thr171, suggesting that phosphorylation may play arole in regulating Madm subcellular localization. In addition,the N-terminal region of Madm is rich in glutamic acid andserine residues, which could potentially bind SH2 modules in aphosphotyrosine-independent manner (28).
Northern blotting analysis and whole mount in situ hybrid-
ization demonstrated that Madm was expressed in all murinetissues and hemopoietic cell lines studied; moreover, the levelsof mRNA were comparable in all samples analyzed (data notshown). Isolation of murine genomic clones revealed that thegene contains 18 exons, spanning 11 kb (Fig. 2B). Examinationof the promoter region identified possible transcription factorbinding sites typical of a ubiquitously expressed gene (data notshown).
Sequence comparisons of Madm shown in Fig. 3 revealedthat it had 51% amino acid identity with an uncharacterizedprotein from Drosophila melanogaster (AF145690) and 36%identity with a predicted protein from Caenorhabditis elegans(CAA99887), suggesting evolutionary conservation. Recently, ahuman homologue of Madm was submitted to GenBank asnuclear receptor binding protein (NM-013392) because it con-tained two LXXLL putative nuclear receptor binding motifs(29). The human clone contains an open reading frame of iden-
FIG. 3. Alignment of the Madm protein sequence from different species. The GenBank accession numbers are: Homo sapiens,NM-013392; M. Mus musculus, AF302138; D. melanogaster, AF145690; C. elegans, CAA99887.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs 41001
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

FIG. 4. Madm and Mlf1 co-immunoprecipitate and co-localize. A, lysates from COS cells transfected with Mlf1 and/or Myc-tagged Madmwere immunoprecipitated (IP) with anti-Myc antibodies and immunoblotted (IB) with antibodies to Madm or Mlf1. B, nuclear and cytoplasmiclocalization of Madm in transfected COS cells expressing green fluorescent protein-Madm (green). Nuclei (blue) were stained with Hoeschst 33258.C, COS cells transfected with Madm were separated into nuclear and cytoplasmic fractions and then immunoblotted (IB) with antibodies againstMadm, nucleolin (nuclear marker), and 14-3-3� (cytoplasmic marker). D, COS cells were co-transfected with Madm and Mlf1. Madm was detectedusing anti-Myc antibodies and a green fluorescent anti-mouse secondary antibody, while Mlf1 was detected with anti-Mlf1 and a red fluorescentanti-rabbit secondary antibody. Nuclei are shown in blue.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs41002
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

tical size to Madm and shares 98% amino acid identity (Fig. 3).Madm and Mlf1 Co-immunoprecipitate and Co-localize in
Cells—To investigate the interaction between Madm and Mlf1further, COS cells were transfected with plasmids expressingfull-length Mlf1 and Myc-tagged Madm. Fig. 4A shows a 65-kDa band for tagged Madm, which is close to the predictedmolecular mass of 60 kDa. Significantly, transfection of bothMadm and Mlf1 resulted in co-immunoprecipitation of the twoproteins. A reciprocal co-immunoprecipitation in cells express-ing HA-tagged Mlf1 and full-length Madm supported theseobservations (data not shown). These data confirm the associ-ation between Madm and Mlf1 identified by yeast two-hybridanalysis.
Mlf1 has been shown previously to be localized primarily inthe cytoplasm, but also within nuclear spots (1, 6). Confocalmicroscopy was employed to determine the subcellular local-ization of Madm and whether this overlapped with Mlf1. Fig.4B shows that Madm could be detected mainly in the cytoplasmof transfected COS cells, with some expression in the nucleus.Biochemical analyses of nuclear and cytosolic fractions con-firmed that Madm was present in both compartments (Fig. 4C).Co-localization with Mlf1 was observed primarily in the cyto-plasm, especially in the perinuclear region including centro-
somes (Fig. 4D). Therefore, Madm and Mlf1 associate in theyeast two-hybrid system, co-immunoprecipitate, and sharesubcellular locations.
Madm and Mlf1 Binding Domains—To characterize the do-mains required for the Madm and Mlf1 interaction, a series ofdeletion mutants was generated and tested in both yeast two-hybrid and co-immunoprecipitation studies. Fig. 5A summa-rizes the data from yeast two-hybrid experiments and showsthat amino acids 406–479 of Madm, encoded by the cDNAfragment originally identified in the yeast two-hybrid screen,were able to bind Mlf1. Similarly, deletion mutants of Mlf1were examined for their Madm-binding ability in yeast. Wild-type Mlf1 and a C-terminal deletion to residue 227, boundMadm (Fig. 5B). The inability of the 1–163 mutant to bindMadm suggested that amino acids between 163 and 227 wereimportant for this association. However, a mutant consisting ofamino acids 122–267 did not bind Madm, indicating that re-gions in the N terminus of Mlf1 were also required for interac-tion. Furthermore, the 39–267 mutant was able to bind Madm,whereas the 82–267 mutant did not. Collectively these dataindicate that a combination of residues between amino acids39–82 and 163–227 are involved in the Madm association.These findings were confirmed by co-immunoprecipitation from
FIG. 5. Interaction domains between Madm, Mlf1, and NPM-MLF1. A, deletion mutants of Madm were used to identify domains thatassociated with full-length Mlf1. B, Mlf1 deletion mutants were used to identify Madm-binding regions. C, deletion mutants of NPM-MLF1 weretested for their ability to associate with Madm.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs 41003
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

FIG. 6. Madm, Mlf1, and NPM-MLF1are phosphorylated on serine resi-dues. A, full-length Madm (Madm FL)and deletion mutants lacking the kinase-like domain (Madm�kinase), the 57, 129,or 209 C-terminal residues (Madm�C57,Madm�C129, or Madm�C209) were im-munoprecipitated (IP) and then subjectedto in vitro kinase assays. The fold in-crease in myelin basic protein (MBP; 1�g) phosphorylation is shown below. B,COS cells transfected with Madm and/orMlf1 were lysed, Madm immunoprecipi-tated (IP) with anti-Myc antibodies, andthen subjected to a kinase assay. C, COScells transfected with Madm, Mlf1, orNPM-MLF1 were phosphorylated in vivowith H3
32PO4 and protein immunopre-cipitated as shown. D, phosphoamino acidanalysis of in vivo phosphorylated Madm,Mlf1, and NPM-MLF1. The position ofninhydrin-stained amino acid standardsis shown by circles. E, peptides bearingthe 14-3-3-binding motif of Mlf1 were in-cubated with Myc-tagged Madm/kinasecomplex immunoprecipitated with anti-Myc antibodies before being subjected to akinase assay.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs41004
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

COS cells where expression of the mutant proteins wasdemonstrated.
Binding of the fusion molecule NPM-MLF1 to Madm wasalso ascertained using co-immunoprecipitation experimentsfrom COS cells. The full-length NPM-MLF1 and a C-terminal-truncated protein were able to interact with Madm (Fig. 5C). Inaddition, deletion of amino acids 281–362 of NPM-MLF1 (cor-responding to amino acids 121–202 of MLF1) retained Madmbinding; however, removal of the N-terminal residues of MLF1present in the fusion protein (corresponding to amino acids17–120 in the normal MLF1 protein) prevented binding. Fromthese data, it appears that Mlf1 and NPM-MLF1 share anN-terminal region that interacts with Madm, but that thesecond, C-terminal, region identified in MLF1 may be dispen-sable for NPM-MLF1 interaction with Madm.
Madm Associates with a Serine Kinase That PhosphorylatesMadm and Mlf1—Although Madm has a consensus kinasedomain, it lacks a typical ATP-binding motif (Fig. 2); it was,therefore, important to determine whether Madm was a genu-ine kinase. When bacterially produced Madm was subjected toan autokinase assay there was no protein phosphorylation,indicating an absence of catalytic activity (data not shown).This observation was also supported by the inability of mam-malian-expressed Madm to autophosphorylate under the re-ducing conditions of in-gel kinase assays. However, when im-munoprecipitated from transfected COS cells, full-lengthMadm and several deletion mutants, including a Madm con-struct lacking the kinase-like domain, were phosphorylated in
in vitro kinase assays (Fig. 6A). These data suggested that aseparate kinase was precipitating with, and phosphorylating,Madm. Indeed, the phosphorylation of myelin basic protein wasincreased 3–7-fold when added to kinase assays containingMadm immunoprecipitates (Fig. 6A). In addition, the kinaserecruited by Madm was able to phosphorylate co-precipitatedMlf1 (Fig. 6B).
The in vivo phosphorylation status of Madm, Mlf1, and thefusion NPM-MLF1 were investigated in transfected COS cells.When the cells were labeled with [32P]phosphoric acid, all threeproteins were phosphorylated (Fig. 6C). Phosphoamino acidanalysis of Madm, Mlf1, and NPM-MLF1, following in vivophosphorylation, revealed that these proteins were phospho-rylated on serine residues (Fig. 6D). Preliminary studies haveexcluded several well characterized kinases as the Madm-associated kinase, including protein kinase A, protein kinase C,Akt, and calcium/calmodulin-dependent kinase. It was con-cluded from these experiments that Madm recruits a serinekinase that phosphorylates both Madm and Mlf1.
Madm-associated Kinase Phosphorylates 14-3-3 Binding Mo-tifs—As MLF1 contains a consensus 14-3-3 binding site andMadm binds a serine kinase, we postulated that Madm maymediate phosphorylation of the RSXSXP motif in MLF1. Todetermine whether the Madm/kinase complex was capable ofphosphorylating this domain, the peptide 26MRQMIRSFSEPF-GRDL41 was used in an in vitro kinase assay. Fig. 6E showsthat in addition to the phosphorylation of Madm seen in trans-fected COS cells, a concentration-dependent phosphorylation of
FIG. 7. Madm forms dimers via a C-terminal region. A, COS cells transfected with vector, Myc-tagged and/or HA-tagged Madm were lysedand immunoblotted (IB) with anti-Myc or anti-HA antibodies (upper panels). Dimerization was demonstrated by Madm immunoprecipitation (IP)using anti-Myc antibodies, followed by immunoblotting with anti-HA antibodies (lower panel). B, deletion mutants of Madm were analyzed for theirability to bind full-length Madm by reporter gene activation in yeast.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs 41005
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from
the MLF1 peptide bearing the 14-3-3 binding site occurred.Although a limited amount of peptide phosphorylation by en-dogenous kinase(s) was observed in non-transfected COS cells,phosphorylated peptide levels were 4–6-fold higher with theexogenous Madm complex. These data demonstrate that Madmbinds a kinase capable of phosphorylating the RSXSXP motif ofMLF1. Phosphorylation of the RSXSXP motif of Raf1 (8) wasalso observed with the Madm complex but not the non-canon-ical 14-3-3 binding site of Cbl (14) (data not shown).
Madm Dimerizes and Influences Myeloid Differentiation—
Preliminary experiments with the yeast two-hybrid systemindicated that Madm was capable of homodimerization (datanot shown). To confirm that Madm could also dimerize inmammalian cells, Madm constructs tagged with either Myc orHA were co-transfected into COS cells. Co-immunoprecipita-tion with anti-Myc or anti-HA antibodies, followed by appro-priate immunoblotting, confirmed that Madm was indeed ableto self-associate (Fig. 7A). Data presented in Fig. 7B indicatethat the region of Madm involved in homodimerization over-lapped with the Mlf1-binding domain. This observation sug-
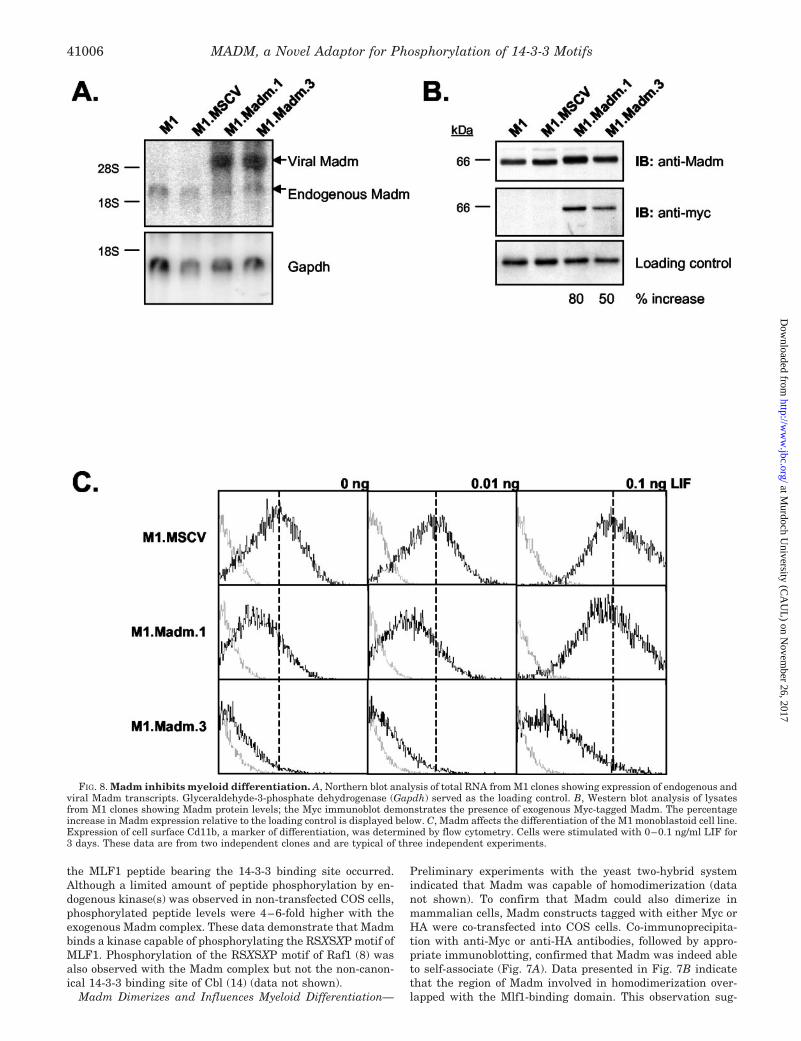
FIG. 8. Madm inhibits myeloid differentiation. A, Northern blot analysis of total RNA from M1 clones showing expression of endogenous andviral Madm transcripts. Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) served as the loading control. B, Western blot analysis of lysatesfrom M1 clones showing Madm protein levels; the Myc immunoblot demonstrates the presence of exogenous Myc-tagged Madm. The percentageincrease in Madm expression relative to the loading control is displayed below. C, Madm affects the differentiation of the M1 monoblastoid cell line.Expression of cell surface Cd11b, a marker of differentiation, was determined by flow cytometry. Cells were stimulated with 0–0.1 ng/ml LIF for3 days. These data are from two independent clones and are typical of three independent experiments.
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs41006
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

gests that Madm and Mlf1 may form multimeric complexes, oralternatively, compete for binding sites.
We have previously shown that Mlf1 overexpression in M1monoblastoid cells enhances differentiation induced by LIF, asmeasured by surface Cd11b expression (6). To determine theimpact of Madm on myeloid maturation, the full-length cDNAwas introduced into M1 cells using the MSCV retroviral vector.Despite large amounts of viral RNA being generated, Madmprotein levels increased no more than 50–80% (Fig. 8, A andB). However, in marked contrast with M1 cells overexpressingMlf1 (Ref. 6 and data not shown), all clones expressing exoge-nous Madm displayed lower Cd11b levels in the absence of LIFor when exposed to suboptimal concentrations of the cytokine(Fig. 8C). Some clones persisted with lower Cd11b expressioneven at higher concentrations of LIF. These results demon-strate that modest alterations to the concentration of Madmcan influence the maturation of M1 cells and that increasingthe intracellular content of Madm had the opposite effect toMlf1 on differentiation.
DISCUSSION
Mlf1 is a recently described oncoprotein (1). It is able toinfluence hemopoietic lineage commitment and reprogramerythroleukemic cells to develop a monoblastoid phenotype (6).The importance of Mlf1 is highlighted by (i) the t(3;5) translo-cation, which exposes the leukemogenic potential of the mole-cule, and (ii) overexpression of wild-type Mlf1 in patients withacute myeloid leukemia, which correlates with poor prognosisand reduced survival (1, 2).
In this manuscript we describe two molecules that interactwith Mlf1, viz. 14-3-3� and Madm. The identification of 14-3-3�as an Mlf1 partner protein in a yeast two-hybrid screen andconfirmation by co-immunoprecipitation was not surprising asthe only recognizable domain in Mlf1 is the 14-3-3 bindingmotif, RSXSXP (3, 6). It is noteworthy that Mlf1 is serine-phosphorylated in vivo and the phosphorylated motif associ-ated with 14-3-3�, but not the unphosphorylated form. 14-3-3proteins have been implicated in many diverse functions, andtheir role in promoting cytoplasmic localization of some partnerproteins is well documented, e.g. phosphorylation of cdc25c byChk1 results in 14-3-3 binding, which masks an NLS andretains cdc25c in the cytoplasm (30, 31). It is possible, likewise,that phosphorylation of the RSXSXP motif in Mlf1 promotes14-3-3 binding, thereby sequestering the molecule in the cyto-plasm and restricting its access to the nucleus. This model isconsistent with primary localization of wild-type Mlf1 in thecytoplasm, with small amounts detected in punctate nuclearbodies (1, 6, 32). Importantly, although the NPM-MLF1 fusionprotein is phosphorylated in vivo, it no longer binds 14-3-3�.Because the fusion junction is only 13 residues from the RSX-SXP motif, it is conceivable that structural alterations mayinhibit the binding of 14-3-3�.
Madm is a novel adaptor molecule for Mlf1. The co-localiza-tion of Madm and Mlf1 suggests that these two proteins areintimately associated, especially in the perinuclear area. Al-though Madm has a clearly defined kinase domain, it does notcontain a characteristic ATP-binding region (29). While a num-ber of kinases that lack this domain have been identified,including Chk1 and cdc7 (33, 34), our data indicate that Madmdoes not possess the catalytic activity of a kinase. Indeed,Hooper et al. considered that the human homologue of Madmwas unlikely to possess functional kinase activity because itlacked 7 of the 15 highly conserved kinase domain residues(29). However, Madm does recruit a serine kinase that phos-phorylates both Madm and Mlf1. Specifically, the Madm-asso-ciated kinase was able to phosphorylate the RSXSXP motif ofMlf1. Phosphorylation of the comparable motif in Raf1 by the
Madm-kinase complex indicates that Madm may act as a moreglobal adaptor involved in phosphorylating other 14-3-3 inter-acting proteins. The extremely high degree of Madm conserva-tion between mouse and human (98% identity) is noteworthy.In addition, Mlf1 shares 79% identity with the human homolog,and 14-3-3 proteins are highly conserved among all species(35). This may indicate that Madm-Mlf1–14-3-3� complexesserve evolutionarily conserved functions.
The opposing effects of Madm and Mlf1 on M1 cell differen-tiation are significant. Whereas expression of exogenous Mlf1potentiated M1 maturation (6), differentiation was inhibitedwhen the level of Madm was raised only 50–80%. This modestincrease in the concentration of Madm had a marked impact onthe capacity of myeloid cells to mature and was contrary to theeffects of Mlf1. As the dimerization domain of Madm over-lapped with the Mlf1-binding region, altering the Madm/Mlf1ratio is likely to affect complex formation between these pro-teins. The composition of these complexes may, in turn, dictatethe progress of terminal differentiation. It is unlikely thatthese effects were solely due to gross mRNA overexpressionbecause equivalent amounts of retroviral transcripts were de-tected in M1 cells expressing the vector alone (data not shown).
Madm contains a bipartite NLS, as well as an NES. Confocalmicroscopy and biochemical analyses revealed that Madm waspresent in both the cytoplasm and nucleus, indicating it mayshuttle between the compartments. It is interesting to notethat Madm has a potential phosphorylation site within theNLS, which could possibly regulate transport to the nucleus asseen with IFI-16 (36) and APC (20). The presence of two nu-clear receptor binding motifs introduces the tantalizing pros-pect of specific functions for Madm within the nucleus.
There are several possible reasons for the leukemogenic ac-tions of NPM-MLF1. One likely explanation is the inappropri-ate cellular compartment occupied by the fusion protein (1, 32).Unlike wild-type Mlf1, NPM-MLF1 is found almost exclusivelyin the nucleus. Data presented in this manuscript demonstratethat NPM-MLF1 is unable to associate with 14-3-3� and thatthe C-terminal Madm-binding domain is compromised. Theseobservations suggest that the NPM portion of the fusion pro-tein sterically hinders 14-3-3 binding and interferes with theMadm interaction. As a consequence, NPM-MLF1 may not besequestered in the cytoplasm and is transported into the nu-cleus by the NLS within NPM.
We postulate that the two partner proteins of Mlf1 describedin this study play a pivotal role in regulating its subcellularlocalization and potentially its function. Madm recruits a ser-ine kinase that phosphorylates the RSXSXP motif of Mlf1,thereby promoting 14-3-3� binding. Complexes involving Mlf1,Madm, and 14-3-3� are likely to influence the distribution ofthese proteins in the cell and affect their ability to differentiate.
Acknowledgments—We are grateful to Professor M. Berndt (BakerMedical Research Institute, Melbourne, Australia) for the generous giftof the anti-14-3-3� antibody.
REFERENCES
1. Yoneda-Kato, N., Look, A. T., Kirstein, M. N., Valentine, M. B., Raimondi,S. C., Cohen, K. J., Carroll, A. J., and Morris, S. W. (1996) Oncogene 12,265–275
2. Matsumoto, N., Yoneda-Kato, N., Iguchi, T., Kishimoto, Y., Kyo, T., Sawada,H., Tatsumi, E., and Fukuhara, S. (2000) Leukemia 14, 1757–1765
3. Hitzler, J. K., Witte, D. P., Jenkins, N. A., Copeland, N. G., Gilbert, D. J.,Naeve, C. W., Look, A. T., and Morris, S. W. (1999) Am. J. Pathol. 155,53–59
4. Klinken, S. P., Nicola, N. A., and Johnson, G. R. (1988) Proc. Natl. Acad. Sci.U. S. A. 85, 8506–8510
5. Keil, U., Busfield, S. J., Farr, T. J., Papadimitriou, J., Green, A. R., Begley,C. G., and Klinken, S. P. (1995) Cell Growth Differ. 6, 439–448
6. Williams, J. H., Daly, L. N., Ingley, E., Beaumont, J. G., Tilbrook, P. A.,Lalonde, J.-P., Stillitano, J. P., and Klinken, S. P. (1999) EMBO J. 18,5559–5566
7. Yaffe, M. B., Rittinger, K., Volinia, S., Caron, P. R., Aitken, A., Leffers, H.,
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs 41007
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

Gamblin, S. J., Smerdon, S. J., and Cantley, L. C. (1997) Cell 91, 961–9718. Muslin, A. J., Tanner, J. W., Allen, P. M., and Shaw, A. S. (1996) Cell 84,
889–8979. Liu, Y. C., Liu, Y., Elly, C., Yoshida, H., Lipkowitz, S., and Altman, A. (1997)
J. Biol. Chem. 272, 9979–998510. Waterman, M. J. F., Stavridi, E. S., Waterman, J. L. F., and Halazonetis, T. D.
(1998) Nat. Genet. 19, 175–17811. Freed, E., Symons, M., Macdonald, S. G., McCormick, F., and Ruggieri, R.
(1994) Science 265, 1713–171612. Zha, J., Harada, H., Yang, E., Jockel, J., and Korsmeyer, S. J. (1996) Cell 87,
619–62813. Peng, C. Y., Graves, P. R., Thoma, R. S., Wu, Z., Shaw, A. S., and Piwnica-
Worms, H. (1997) Science 277, 1501–150514. Robertson, H., Langdon, W. L., Thien, C. B. F., and Bowtell, D. D. L. (1997)
Biochem. Biophys. Res. Comm. 240, 46–5015. Reuther, G. W., Fu, H., Cripe, L. D., Collier, R. J., and Pendergast, A.-M.
(1994) Science 266, 129–13316. Muslin, A. J., and Xing, H. (2000) Cell. Signal. 12, 703–70917. Vojtek, A. B., Hollenburg, S. M., and Cooper, J. A. (1993) Cell 1993, 205–21418. Tsai, S., Bartelmez, S., Sitnicka, E., and Collins, S. (1994) Genes Dev. 8,
2831–284119. Smith, D. B., and Johnson, K. S. (1988) Gene 67, 31–4020. Zhang, F., White, R. L., and Neufeld, K. L. (2000) Proc. Natl. Acad. Sci. U. S. A.
97, 12577–1258221. Tilbrook, P. A., Ingley, E., Williams, J. H., Hibbs, M. L., and Klinken, S. P.
(1997) EMBO J. 16, 1610–1619
22. Hardin, S. C., and Wolniak, S. M. (1998) BioTechniques 24, 344–34623. Miller, A. D., and Buttimore, C. (1986) Mol. Cell. Biol. 6, 2895–290224. Hawley, R. G., Lieu, F. H., Fong, A. Z., and Hawley, T. S. (1994) Gene Ther. 1,
136–13825. Kozak, M. (1987) Nucleic Acids Res. 15, 8125–813126. Schultz, J., Milpetz, F., Bork, P., and Ponting, C. P. (1998) Proc. Natl. Acad.
Sci. U. S. A. 95, 5857–586427. Hanks, S. K., and Hunter, T. (1995) FASEB J. 9, 576–59628. Joung, I., Strominger, J. L., and Shin, J. (1996) Proc. Natl. Acad. Sci. U. S. A.
93, 5991–599529. Hooper, J. D., Baker, E., Ogbourne, S. M., Sutherland, G. R., and Antalis, T. M.
(2000) Genomics 66, 113–11830. Peng, C.-Y., Graves, P. R., Ogg, S., Thoma, R. S., Byrnes, M. J., Wu, Z.,
Stephenson, M. T., and Piwnica-Worms, H. (1998) Cell Growth Diff. 9,197–208
31. Lopez-Girona, A., Furnari, B., Mondesert, O., and Russell, P. (1999) Nature397, 172–175
32. Yoneda-Kato, N., Fukuhara, S., and Kato, J.-Y. (1999) Oncogene 18,3716–3724
33. Walworth, N., Davey, S., and Beach, D. (1993) Nature 363, 368–37134. Patterson, M., Sclafani, R. A., Fangman, W. L., and Rosamond, J. (1986) Mol.
Cell. Biol. 6, 1590–159835. Aitken, A., Collinge, D. B., van Heusden, B. P., Isobe, T., Roseboom, P. H.,
Rosenfeld, G., and Soll, J. (1992) Trends Biochem. Sci. 17, 198–50136. Briggs, L. J., Johnstone, R. W., Elliot, R. M., Xiao, C. Y., Dawson, M., Trapani,
J. A., and Jans, D. A. (2001) Biochem. J. 353, 69–77
MADM, a Novel Adaptor for Phosphorylation of 14-3-3 Motifs41008
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from

Sun, Xiaohua Wu, Stephan W. Morris and S. Peter KlinkenIngley, Jim Y-H. Tiao, Jean-Philippe Lalonde, Schickwann Tsai, Peta A. Tilbrook, Yi Raelene Lim, Louise N. Winteringham, James H. Williams, Ross K. McCulloch, Evan
Binding Site of Myeloid Leukemia Factor 1MADM, a Novel Adaptor Protein That Mediates Phosphorylation of the 14-3-3
doi: 10.1074/jbc.M206041200 originally published online August 9, 20022002, 277:40997-41008.J. Biol. Chem.
10.1074/jbc.M206041200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/277/43/40997.full.html#ref-list-1
This article cites 36 references, 16 of which can be accessed free at
at Murdoch U
niversity (CA
UL
) on Novem
ber 26, 2017http://w
ww
.jbc.org/D
ownloaded from





























