Embed Size (px)
Citation preview

695
Mycologia, 97(3), 2005, pp. 695–709.q 2005 by The Mycological Society of America, Lawrence, KS 66044-8897
The hyphomycete Teberdinia hygrophila gen. nov., sp. nov. and relatedanamorphs of Pseudeurotium species
M.V. SogonovSystematic Botany and Mycology Laboratory, USDA,Beltsville, Maryland
H.-J. SchroersPlant Protection Department, Agricultural Institute ofSlovenia, Ljubljana, Slovenia
W. GamsJ. DijksterhuisR.C. Summerbell1
Centraalbureau voor Schimmelcultures, Uppsalalaan8, 3584 CT Utrecht, The Netherlands
Abstract: A hyphomycetous fungus isolated frommontane fen soil in the Caucasus Mountains, Russia,had obscurely sympodial conidiogenous cells thatsuggested a link to the heterogeneous genus Lepto-dontidium. Sequence analysis of the nuclear ribosom-al small subunit and internal transcribed spacerregion, however, disclosed that the fungus was ananamorphic member of a clade containing the cleis-tothecial ascomycetous genus Pseudeurotium. Teber-dinia, gen. nov., is proposed for the blastic, generallysympodially proliferating anamorphs in this group,and Teberdinia hygrophila, sp. nov., is proposed forthe species from upland fens. Binomials are not pro-posed for the remaining Teberdinia anamorphs ofPseudeurotium species. Purely anamorphic isolates inthis clade are difficult to recognize using currentmorphological keys and might be more widely dis-tributed and ecologically significant than is currentlyevident.
Key words: fen habitat, internal transcribed spac-er regions, Leptodontidium, Pseudeurotiaceae, phy-logeny, small subunit ribosomal DNA
INTRODUCTION
The genus Pseudeurotium with one species, P. zona-tum, was established in the family Eurotiaceae by vanBeyma (1937). Thirteen additional species subse-quently were placed in the genus. They were P. ovaleStolk, P. multisporum (Saito & Minura) Stolk (Stolk1955), P. indicum (Chattop. & Das Gupta) Chattop.
Accepted for publication 15 Mar 2005.1 Corresponding author. E-mail: [email protected]
(Chattopadhyay 1957), P. bakeri C. Booth, P. rilstoniiC. Booth (Booth 1961), P. globosum J.N. Rai & Tewari(Rai and Tewari 1961), P. punctatum Panasenko(Panasenko 1964), P. irregulare Lodha, P. jaipurenseLodha (Lodha 1971), P. desertorum Mouchacca(Mouchacca 1971), P. luteolum Matsushima (Matsush-ima 1975) and P. macroglobosum Matsushima (Mat-sushima 1996). In addition two varieties were de-scribed, namely P. indicum var. oryzetorum A.M. Corte(Montemartini Corte 1968) and P. ovale var. milkoiBelyakova (Belyakova 1969). P. multisporum, P. indi-cum, P. globosum and P. rilstonii later were transferredto other genera (Cain 1961, Rai and Tewari 1963,Malloch 1974). Nothing has been published on theredisposition of P. indicum var. oryzetorum, but au-thentic culture CBS 309.72 later was found to be anisolate of Westerdykella multispora (Saito & Minoura)Cejp & Milko and it now is listed under this name inthe CBS list of cultures (2001). Among the remainingPseudeurotium taxa only P. zonatum, P. ovale, P. ovalevar. milkoi, P. bakeri and P. desertorum are still avail-able as living cultures. P. irregulare, P. jaipurense, P.luteolum and P. macroglobosum were preserved only asherbarium specimens. There is no indication of atype in the description of P. punctatum.
The position of the genus Pseudeurotium in the sys-tem of Ascomycetes was altered by Malloch and Cain(1970), who established the family Pseudeurotiaceaeincluding four existing genera, Pseudeurotium, Emer-icellopsis, Fragosphaeria and Testudina, and five newlydescribed genera, Cryptendoxyla, Hapsidospora, Leu-coneurospora, Mycoarachis and Nigrosabulum. Phylo-genetic research by Suh and Blackwell (1999)changed the concept of the family Pseudeurotiaceaeso that among Malloch’s and Cain’s genera only Pseu-deurotium and Leuconeurospora remained there whileConnersia (with only one species initially placed inPseudeurotium) and Pleuroascus were added. The datashowed a strong distinction between Pseudeurotiaceaeand Eurotiaceae. The partial 18S ribosomal subunitstudy of Gernandt et al (2001) grouped the Pseudeu-rotiaceae loosely as ‘‘incertae sedis,’’ situated ‘‘nearthe base of Helotiales’’ and at considerable distancefrom the Eurotiales.
The conidial states of Pseudeurotium species havebeen described as Sporothrix-, Beauveria- or Acremo-nium-like. No generic name has been assigned tothem.

696 MYCOLOGIA
During a synecological study on soil fungi of alpineecosystems in Teberda State Reserve (Karachai-Cher-kess Republic, Northwestern Caucasus, Russia), anumber of anamorphic strains characterized by pro-ducing a sympodial succession of blastoconidia in ob-scurely rosette-like clusters was obtained from thesoils of two alpine fens (Sogonov and Velikanov2005). The strains were identified provisionally asLeptodontidium sp. Their morphology initially ap-peared suggestive of this genus, although the isolatesclearly were not identifiable as any described mem-ber of the genus. Subsequent 18S ribosomal DNA se-quence comparisons of two representative isolatesshowed that the alpine strains were not congenericwith the type species of Leptodontidium, L. elatius, butinstead were close to P. zonatum.
The object of the current study was to investigatethese isolates in terms of their phylogenetic relation-ships as well as their anamorph morphological simi-larities to the Pseudeurotium species available in cul-ture.
MATERIALS AND METHODS
Strains studied are listed (TABLE I). For morphological ob-servation and description, strains were grown on potato-car-rot agar (PCA), oatmeal agar (OA) and malt-extract agar(MEA) (Gams et al 1998) at 18 C in the dark for 20 d.Colors of the colony obverse and reverse were determinedaccording to Kornerup and Wanscher (1978). Microscopicobservations, line drawings and photographs were made us-ing the Olympus BX 50 microscope (Olympus Optical Co.,Hamburg, Germany) with conventional bright-field or No-marski interference contrast microscopy from preparationsin lactic acid solutions with or without cotton blue (0.5 mg/mL). Drawings and light microscopy photographs weremade with a U-DA camera lucida (Olympus) and DP 10digital camera (Olympus). Photos were taken with bright-field illumination unless stated otherwise. Microscopic mea-surements were made with an ocular micrometer at 10003and are given as the size range of perceived typical struc-tures, flanked in parentheses by the extreme sizes seen inrelatively rare variant structures.
For mating trials the anamorph strains CBS 102670, CBS102671 and CBS 326.81 were paired in all combinations.Pairings were inoculated 1.5 cm apart on OA with lupinestems and incubated under combined 24 h ultraviolet light(356 nm, Sylvania Blacklight Blue, Osram Nederland, Al-phen a/d Rijn, Netherlands) and natural daylight at roomtemperature for 1 y.
For scanning electron microscopy (SEM) strains weregrown on synthetic nutrient agar (SNA) (Gams et al 1998)and OA at 18 C in the dark for 8 d. Blocks (ca. 3 3 7 mm)were cut from colonies growing on agar. They were fixedeither in glutaraldehyde and chemically dehydrated beforecritical point drying as described by Samson et al (1979) orprepared for the low temperature SEM procedure de-
scribed by Dijksterhuis et al (1991). SEM photographs weremade with a JSM-840A scanning electron microscope ( JeolLtd., Tokyo, Japan).
For molecular study the fungi were grown on liquid com-plete yeast medium (CYM, Raper and Raper 1972). DNAwas extracted with the cetyltrimethylammonium bromide(CTAB) procedure adopted from Gerrits van den Ende andde Hoog (1999) or with the FastDNAtKit (Qbiogene, Irvi-ne, California) according to the manufacturer’s instruc-tions.
The 50 mL PCR mixtures contained 1–2 mL DNA extract,0.25 mM of each dNTP (Amersham Pharmacia Biotech,Amersham Place, Little Chalfont, Buckinghamshire, UK),0.25–3.0 pM of each of primers, 2 U DNA polymerase (Su-per Taqt, HT Biotechnology Ltd., Cambridge, UK), 5 mLof the standard PCR buffer provided together with the DNApolymerase and deionized water. PCR was performed in aGenAmpt PCR System 9700 (Applied Biosystems, FosterCity, California) with this regime: 35 cycles consisting ofdenaturation for 30–35 s at 94 C, annealing for 50 s at 52–55 C and extension for 120 s at 72 C; a final extensionperiod of 5–6 min at 72 C followed by chill to 4 C. Theregion of nuclear ribosomal DNA (rDNA) spanning the in-ternal transcribed spacer 1, 5.8S RNA gene, and internaltranscribed spacer 2 regions (ITS1-5.8S-ITS2, further abbre-viated to ITS) was amplified with primer pairs ITS1/NL4(White et al 1990, O’Donnell 1993) for strains CBS 102670,CBS 102671 and CBS 326.81, and V9D/ITS4 (de Hoog andGerrits van den Ende 1998, White et al 1990) for CBS878.71, CBS 986.72 and CBS 443.78. The 18S region wasamplified with primers NS1 and NS24 (White et al 1990,Gargas and Taylor 1992). After cleanup of the PCR prod-ucts with the GFXy PCR DNA and Gel Band PurificationKit (Amersham Pharmacia) and a check on final concen-tration, the DNA amplified with primers ITS1 and ITS4(White et al 1990) for the ITS region, and with NS1, OLI1,OLI2 and OLI9 for the 18S region, was sequenced with anABI Prismt 3700 DNA sequencer (Applied Biosystems).
Data analysis.—BLAST searching in GenBank was used toobtain relevant comparison sequences (TABLE II). Sequenc-es were aligned manually in BioEdit 4.8.9 (Hall, http://www.mbio.ncsu.edu/BioEdit/bioedit.html). Phylogeneticanalysis was performed by heuristic tree searches using themaximum parsimony criterion in PAUP 4.0b4a (Swofford2000). Branch robustness was tested based on 1000 boot-strap replicates (randomly sampled datasets with replace-ments). Heuristic tree searches were performed with start-ing trees obtained via stepwise sequence addition, tree bi-section-reconnection (TBR) as swapping algorithm, mul-trees on, and using all optimal trees for the next round ofswapping. For the analysis of 18S and ITS sequences, themaximal tree number was set respectively at 10 000 (1000in bootstrap analyses) and at 1000 (10 in bootstrap analy-ses). Parsimony uninformative characters were excludedfrom the analyses. Characters were unordered and equallyweighted. Sequence addition was random and was done in100 (10 in bootstrap analyses) replications on the 18S rDNAdataset and 1000 (10 in bootstrap analyses) on the ITS da-taset. A large intron in the 18S region of Leptodontidium
697SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
TA
BL
EI.
Lis
tof
the
Pse
ude
uro
tiu
man
dan
amor
phic
Pse
ude
uro
tiu
m-li
kest
rain
sex
amin
ed
Spec
ies
Stat
usC
BS
num
ber
Oth
erre
fere
nce
Sour
ceG
eogr
aph
y
Gen
Ban
kac
cess
ion
num
bers
ITSI
-5.8
S-IT
S218
S
Pse
ude
uro
tiu
mzo
nat
um
FHB
eym
aty
pe32
9.36
BD
UN
37,
IMI
1474
45so
il,n
ear
gas
leak
age
Vers
aille
s,Fr
ance
AY1
2928
6
P.ba
keri
C.
Boo
thty
pe87
8.71
FPR
LS8
10,
IMI
0737
49,
VK
MF-
1258
Pla
typu
scy
lindr
us
tun
-n
els
inQ
uer
cus
sp.
UK
AY1
2928
7
P.ov
ale
Stol
kty
pe38
9.54
IMI
0598
42du
ng
ofbi
rdSt
eato
rnis
Cue
vade
lG
uach
aro,
Car
ipe,
Edo
Mon
agas
,Ven
ezue
laA
Y129
289
P.ov
ale
var.
milk
oiB
ely-
akov
a44
3.78
cyst
ofG
lobo
dera
rost
o-ch
ien
sis
insa
ndy
soil
AY1
2929
0
P.de
sert
oru
mM
ouch
ac-
caty
pe98
6.72
AT
CC
2453
5,IM
I17
1135
,LC
P70
.211
3,O
rrO
-117
8,V
KM
F-18
33
dese
rtso
ilO
asis
deD
akh
la,
Wes
tern
Des
ert,
Egy
ptA
Y129
288
Teb
erdi
nia
hygr
ophi
lasp
.n
ov.
type
1026
70pe
aty
soil
ofal
pin
efe
nTe
berd
aSt
ate
Res
erve
,K
ara-
chai
-Ch
erke
ssR
epub
lic,
Rus
sia
AY1
2929
1A
Y129
282
Teb
erdi
nia
hygr
ophi
lasp
.n
ov.
1026
71V
KM
FW-2
055
peat
yso
ilof
alpi
ne
fen
Tebe
rda
Stat
eR
eser
ve,
Kar
a-ch
ai-C
her
kess
Rep
ublic
,R
ussi
a
AY1
2929
2
Teb
erdi
nia
hygr
ophi
lasp
.n
ov.
326.
81po
llute
dw
ater
from
po-
tato
mea
lfa
ctor
yO
veri
jsse
lse
Vech
t,th
eN
eth
-er
lan
dsA
Y129
293
Lep
todo
nti
diu
mel
atiu
s(M
ange
not
)de
Hoo
gva
r.el
atiu
s
type
381.
59D
AO
M42
956,
MU
CL
1575
2de
cayi
ng
woo
dof
Abi
esba
lsam
eaG
reen
Riv
er,
New
foun
dlan
d,C
anad
aA
Y129
285
AY1
2928
0
Lep
todo
nti
diu
mbo
real
e(d
eH
oog)
deH
oog
type
682.
76pi
ne
woo
dpo
lein
grou
nd
con
tact
Swed
enA
Y129
284
AY1
2928
3
Lep
todo
nti
diu
mir
regu
la-
re(d
eH
oog)
deH
oog
type
963.
72w
hit
ero
tin
Not
hofa
gus
dom
beyi
Ral
un,
Ch
ileA
Y129
281

698 MYCOLOGIA
TABLE II. Additional taxa included in the analysis
Classification Taxon
Accession number
18S ITS1-5.8S-ITS2
SaccharomycetesSaccharomycetales Saccharomyces cerevisiae Meyen ex Hansen Z75578
AscomycetesEurotiales Penicillium chrysogenum Thom M55628 AJ270768Pezizales Peziza badia Pers. L37539
Morchella esculenta (L.) Pers. L36998Dothideales Dothidea insculpta Wallr. U42474Helotiales Bulgaria inquinans Pers. AJ224362
Loramyces juncicola W. Weston AF203464Sclerotinia sclerotiorum (Lib.) de Bary X69850Phacidium infestans P. Karst. AF203466
Rhytismatales Darkera parca H. S. Whitney, J. Reid & Piroz. AF203465Erysiphales Blumeria graminis (DC.) Speer L26253Hypocreales Hypocrea schweinitzii (Fr.) Sacc. L36986
Nectria cinnabarina (Tode) Fr. AB003949Xylariales Xylaria curta Fr. U32417Ophiostomatales Ophiostoma ulmi (Buisman) Nannf. M83261
Incertae cedisMyxotrichaceae Myxotrichum deflexum Berk. AB015777
Byssoascus striatosporus (G. L. Barron & C. Booth)Arx
AB015776
Pseudogymnoascus roseus Raillo AB015778 AF062819Gymnostellatospora japonica Udagawa, Uchiy. & Ka-
miyaAF062818
Pseudeurotiaceae Pseudeurotium zonatum F.H. Beyma AF096183AF0096184
Connersia rilstonii (C. Booth) Malloch AF096174Pleuroascus nicholsonii Massee & E. S. Salmon AF096182Leuconeurospora pulcherrima (G. Winter) Malloch
& CainAF096178
Anamorphic ascomycetes Beauveria bassiana (Bals.-Criv.) Vuill. AB079125Geomyces pannorum (Link) Sigler & J. W. Carm AB015785 AF307760Oidiodendron tenuissimum (Peck) S. Hughes AB015787Phialocephala fortinii C. J. K. Wang & H. E. Wilcox L76626Scytalidium dimidiatum (Penz.) B. Sutton & Dyko IP127881Sporothrix schenckii Hektoen & C. F. Perkins M85053S. inflata de Hoog M85053
boreale was not found in any other sequence and hence wasexcluded from parsimony analysis. Uninformative and con-stant characters were excluded from the bootstrap analysis.Trees were rooted with basal polytomy.
TAXONOMY
Teberdinia Sogonov, W. Gams, Summerbell etSchroers, anam. gen. nov.Coloniae albidae vel griseo-virides. Cellulae conidiogenae
hyalinae, seu solitariae seu in conidiophoris plus minusveramosis, terminales, intercalares vel laterales, cylindricae velad basim inflatae; conidia apicalia in successione sympodialie cicatricibus inconspicuis producta. Conidia levia, hyalina,ellipsoidea ad globosa.
Colonies pale, white or gray-green, moderately fastgrowing. Conidiogenous cells hyaline, arising on ae-rial or submerged mycelium, smooth walled; in dis-position highly variable (FIGS. 1–6), found as inte-grated hyphal elements giving rise to short, lateralconidiogenous outgrowths (FIGS. 1E; 2F, H, K–M;5D–F; 6A–D), or as unicellular lateral branches orterminal cells (FIGS. 1C, D; 2A, E, G; 3B; 4B, D; 5A,C, G–I), or as intercalary or terminal elements ondiscrete, unbranched or irregularly branched, mini-mally differentiated conidiophores (FIG. 1B, F, G; 2B,C; 3A, E, F; 4E); in shape generally lageniform tobroadly aculeate to undulate, often with a distinctlyinflated region near the base and with an initially

699SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
FIG. 1. Teberdinia hygrophila (CBS 102670). A–C. Conidiophores in situ. D. Conidiophores mounted in cotton blue. E.Single intercalary conidiogenous cell mounted in lactic acid, Nomarski interference contrast. F, G. Conidiophores, scanningelectron microscopy (SEM). H. Conidiogenous tip, SEM. I. Infertile cleistothecia in situ. J. Inner layer of cleistothecialperidium, lactic acid. K. Outer layer of cleistothecial peridium, lactic acid. L. Colony on potato-carrot agar. M. Colony onoatmeal agar. N. Colony on malt-extract agar.

700 MYCOLOGIA
FIG. 2. Conidiophores. A, B. Teberdinia hygrophila (CBS 102670). C. Pseudeurotium zonatum (CBS 329.36). D, E. Pseudeu-rotium bakeri (CBS 878.71). F–I. Pseudeurotium ovale var. ovale (CBS 389.54). J. Pseudeurotium ovale var. milkoi (CBS 443.78).K–M. Pseudeurotium desertorum (CBS 986.72).
701SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
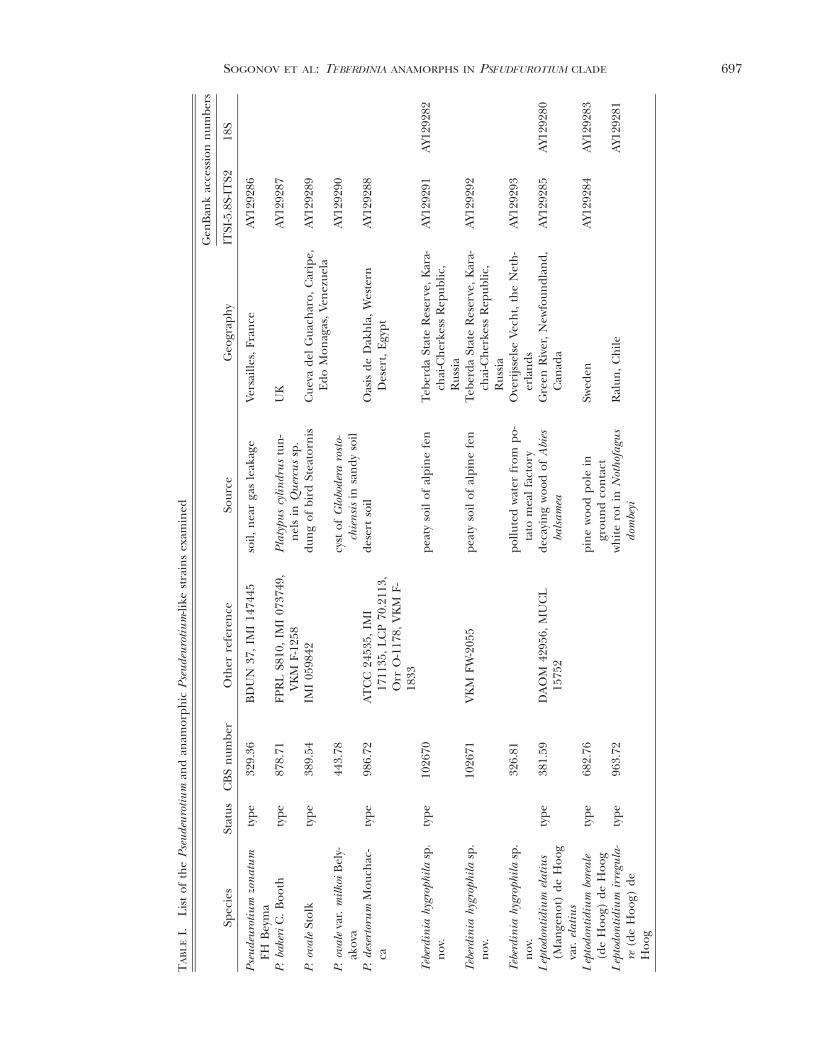
FIG. 3. Pseudeurotium zonatum (CBS 329.36). A, B. Conidiophores in situ. C. Single terminal conidiogenous cell mountedin lactic acid, Nomarski interference contrast. D. Conidiophore mounted in cotton blue. E, F. Conidiophores, scanningelectron microscopy. G. Cleistothecia in situ. H. Cleistothecium and ascospores, lactic acid-cotton blue.
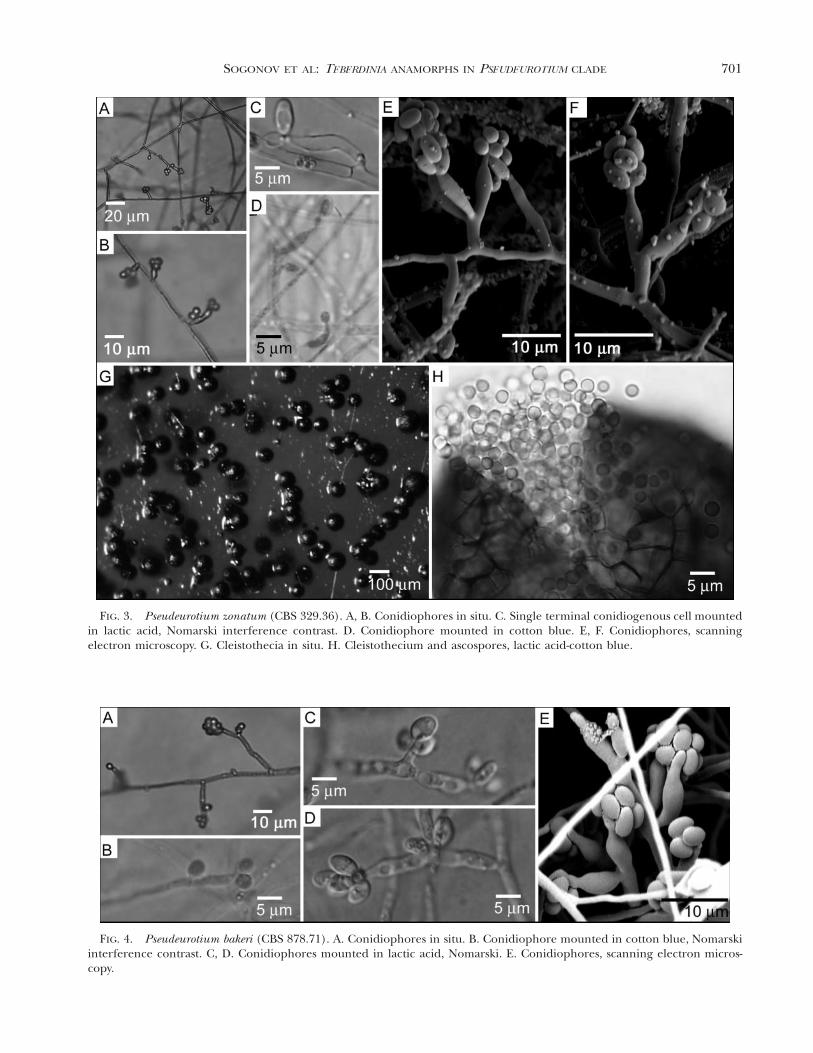
FIG. 4. Pseudeurotium bakeri (CBS 878.71). A. Conidiophores in situ. B. Conidiophore mounted in cotton blue, Nomarskiinterference contrast. C, D. Conidiophores mounted in lactic acid, Nomarski. E. Conidiophores, scanning electron micros-copy.
702 MYCOLOGIA
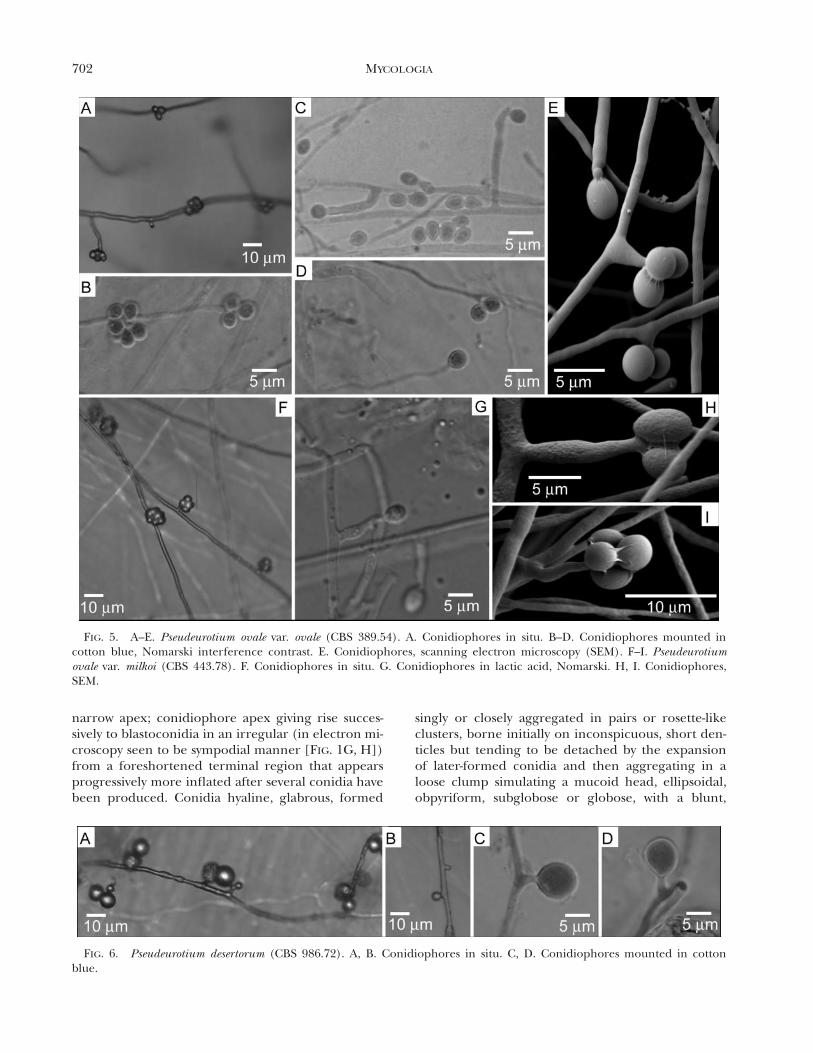
FIG. 5. A–E. Pseudeurotium ovale var. ovale (CBS 389.54). A. Conidiophores in situ. B–D. Conidiophores mounted incotton blue, Nomarski interference contrast. E. Conidiophores, scanning electron microscopy (SEM). F–I. Pseudeurotiumovale var. milkoi (CBS 443.78). F. Conidiophores in situ. G. Conidiophores in lactic acid, Nomarski. H, I. Conidiophores,SEM.
FIG. 6. Pseudeurotium desertorum (CBS 986.72). A, B. Conidiophores in situ. C, D. Conidiophores mounted in cottonblue.
narrow apex; conidiophore apex giving rise succes-sively to blastoconidia in an irregular (in electron mi-croscopy seen to be sympodial manner [FIG. 1G, H])from a foreshortened terminal region that appearsprogressively more inflated after several conidia havebeen produced. Conidia hyaline, glabrous, formed
singly or closely aggregated in pairs or rosette-likeclusters, borne initially on inconspicuous, short den-ticles but tending to be detached by the expansionof later-formed conidia and then aggregating in aloose clump simulating a mucoid head, ellipsoidal,obpyriform, subglobose or globose, with a blunt,

703SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
slightly protuberant attachment scar. Chlamydo-spores not observed. Often associated with Pseudeu-rotium teleomorphic structures.
Typus generis. Teberdinia hygrophila in opere ipsodescripta.
Etymology. Teberda refers to the place of isolationof the ex-type strain.
Teberdinia hygrophila Sogonov, W. Gams, Summer-bell et Schroers anam. sp. nov. FIG. 1Coloniae post 20 dies 18 C 20–40 mm diam, albidae vel
griseo-virides. Cellulae conidiogenae seu solitariae seu inconidiophoris plus minusve ramosis, terminales, intercala-res vel laterales; terminales 6–14 mm longae, cylindricae velad basim inflatae. Conidia late ellipsoidea vel ovoidea, 4.5–5.5 3 2.8–3.0 mm.
Description from pure culture. PCA: Colony di-ameter after 20 d at 18 C in the dark 19–42 mm.Colony surface smooth or slightly powdery at center(FIG. 1L), colorless, pale green (28A3), or greyishgreen (29E5), semitransparent. Reverse concolorouswith the surface. OA: Colony diameter 19–45 mm.Surface smooth, powdery, or loosely cottony (FIG.1M), white, or greyish green (29B4–29E5). Marginswhitish. Reverse colorless or greyish green to darkgreen (29E4–29F4). MEA: Colony diameter 25–45mm. Surface powdery, velvety or cottony (FIG. 1N),whitish, light green (27B5), greyish green (26C3–28B4), or dull green (25D3). Reverse pale to olivebrown (4E4–4F4).
Hyphae hyaline, 1.8–2.5 mm wide; conspicuous hy-phal strands not observed. Conidiophores arisingfrom prostrate aerial or substrate hyphae, consistingof single conidiogenous cells or side branches (up to100–200 mm long) that often display one or moresuccessive orders of subordinate branches disposedsingly or in whorls (FIGS. 1A–G; 2A, B). Conidiogen-ous cells terminal or intercalary; terminal conidi-ogenous cells 6–14 mm long, 2.0–2.8 mm wide at thebase, usually cylindrical or inflated in the lower part,tapering and often bent or sinuous near the tips; in-tercalary conidiogenous cells are ordinary looking,cylindrical hyphal cells bearing a single, often pro-clinally curved, laterally disposed, cytoplasmicallycontiguous conidiogenous projection near the apicalend or less commonly near the midregion (FIGS. 1E;2A, B). These conidiogenous projections are usuallysmall, cylindrical 1.5–2.5 3 0.5–0.9 mm but are vari-able and occasionally are similar to terminal conidi-ogenous cells in size and shape. Conidia seen singlyor closely aggregated in pairs or rosette-like clustersat the apices of conidiogenous cells and lateral pro-jections, borne initially on inconspicuous, short den-ticles but tending to become detached by the expan-sion of later-formed conidia and then aggregating in
a loose clump simulating a mucoid head. Conidia gla-brous, normally hyaline but becoming dark brownafter prolonged storage (1 y), globose to ellipsoidal,(2.4–)2.7–3.4(–4.1) 3 (2.1–)2.3–2.8(–3.1) mm. Basalscars are scarcely visible in light microscopy but areclear in SEM. Chlamydospores not observed.
Infertile cleistothecial structures (FIG. 1I–K; com-pare homologous structures for P. zonatum in FIG.3G, H) seen in a mating cross of CBS 102670 andCBS 326.81 superficial or immersed, dark brown, glo-bose, (90–)130–200 mm diam. Peridium consisting ofinner membranaceous layer of dark-brown, thick-walled, polygonal cells (FIG. 1J, cf. P. zonatum struc-ture in FIG. 3H) measuring 4.6–8.0 mm in face viewand 3.5–4.0 mm thick in cross section as well as anouter layer composed of loose network of dark-brown, thick-walled hyphae (FIG. 1K) 2.3–4.1 mmwide.
Specimens examined. RUSSIA. KARACHAI-CHERKESSREPUBLIC: Teberda, Teberda State Reserve (438279N,418419E) from peaty soil in an alpine fen. Feb 2000. M.V.Sogonov. HOLOTY PE: Herb. CBS 7947, ex-type culture:CBS 102670. Additional specimen from same site: CBS102671. THE NETHERLANDS: Overijsselse Vecht, pollutedwater from potato-meal factory: CBS 326.81.
Etymology. From the Latin hygrophila, refers to theapparent affinity for moist or aqueous habitats.
Habitats. Soil, industrial wastewater.Known distribution. Karachai-Cherkess Republic
(Russian Federation), the Netherlands.
RESULTS
Teleomorphic structures formed in T. hygrophila cross-es.—Infertile ascomatal structures were formed in thecross of strains CBS 102670 and CBS 326.81 after ap-proximately 1 y of coculture on oatmeal agar withlupine stem under 24 h UV combined with naturaldaylight. Single ascomata were placed on a variety ofmedia in hope that the transfer to fresh conditionsmight trigger a ripening process. Although no rip-ening occurred, the colonies initiated by this transferprocedure occasionally produced ascomata again af-ter 15–20 d. This phenomenon, remarkable in lightof the long time needed for formation of ascomatain the initial crosses, was observed only on oatmealagar with horse manure, yeast peptone-glucose agar(Y PG), soil extract agar (SEA) (Gams et al 1998) andSNA. There was no such production of infertile as-comata on potato-carrot agar (PCA), hay-decoctionagar (HAY), potato-dextrose agar (PDA) or Leonian’sagar (Gams et al 1998) under either daylight or com-bined UV/daylight. Ascospores were not formed inany subculture.

704 MYCOLOGIA
Comparative morphological studies of additional Teber-dinia anamorphic states.—The colonial macromor-phology of Pseudeurotium species generally is includ-ed in descriptions of the species known in pure cul-ture. The anamorphic structures present in these cul-tures, however, generally are described incompletely.More detailed descriptions of these micromorpholog-ical features are given below to aid morphologicalcomparison among members of this group. For quickreference a summary is provided (TABLE III). Be-cause the anamorphs are not sharply morphological-ly distinct, those wishing to identify these speciesmorphologically are strongly advised to allow suffi-cient time for formation of any teleomorphs that maydevelop.
Teberdinia state of Pseudeurotium zonatum.FIGS. 3, 2C
Conidiophores arising from aerial or substrate my-celium, consisting of single conidiogenous cells (ter-minal or intercalary) or short side branches (up to50–100 mm). The pattern of conidiophore branchingis simpler than that of T. hygrophila. Most conidio-phore side branches are single; whorls of more thantwo branches are rare. Terminal conidiogenous cells9–22 mm long, lageniform or cylindrical taperingnear their tips, usually bent or sinuous. Conidiogen-ous projections from intercalary cells usually small,cylindrical 1.0–2.0 3 0.7–1.0 mm but variable, occa-sionally similar to terminal conidiogenous cells insize and shape. Conidiogenesis similar to that of T.hygrophila. Conidia glabrous, broadly ellipsoid, ellip-soid or obovoid (4.0–)4.5–5.5(–6.1) 3 (2.4–)2.8–3.1(–3.6) mm.
Teberdinia state of Pseudeurotium bakeri.FIGS. 4; 2D, E
Conidiophores arising from aerial or substrate my-celium, mostly consisting of single conidiogenouscells (terminal or intercalary). Short side branches(up to 25–60 mm) occasionally are found. Terminalconidiogenous cells lageniform, often bent or sinu-ous 10–22 mm; intercalary conidiogenous cells givingrise to lateral conidiogenous processes similar to ter-minal cells or reduced to small denticles on interca-lary cells. Conidiogenesis similar to that of T. hygro-phila. Conidia glabrous, subglobose, ellipsoid or ob-ovoid (4.6–)5.0–5.7(–7.4) 3 (2.8–)3.2–3.5(–4.4) mm.
Teberdinia state of Pseudeurotium ovaleFIGS. 5, 2F–J
Conidiophores arising from aerial or substrate my-celium, prostrate, loosely branched. Most conidi-
ogenous cells are intercalary and bear minute or dis-tinct (up to 12 mm long, 1.5 mm diam) conidiferousprocesses. Terminal conidiogenous cells arise on sidebranches and on the main branch of conidiophore.Terminal conidiogenous cells mostly 7–12(–40) mmlong, cylindrical slightly tapering at the tips, bent orsinuous, sometimes slightly inflated in their midre-gion. Sometimes conidiogenous cells have a second,laterally situated conidiogenous locus. Conidiogene-sis similar to that of T. hygrophila. Conidia glabrous,globose, subglobose or obovoid (3.8–)4.5–5.5(–6.5)3 (2.9–)3.5–4.0(–4.4) mm. Basal scars are clearlyseen in light microscopy. Conidiogenous structuresof the Teberdinia state of CBS 443.78 (FIGS. 2J, 5F–I),the strain of P. ovale var. milkoi studied, did not showany significant difference from those of the Teberdi-nia state of P. ovale var. ovale.
Teberdinia state of Pseudeurotium desertorumFIGS. 6, 2K–M
The anamorph of P. desertorum is the most highlymorphologically reduced anamorph referable to thegenus Teberdinia. It never produces conidia in denseclusters on inflated tips of conidiogenous cells or pro-cesses. Most conidia are single, sessile or disposed onshort (2–3.5 3 1.5–1.6 mm) denticles on mycelialcells. Two conidia occasionally are produced closelytogether on separate short branches at the terminusof denticles (as shown in FIG. 5A and as depicted byMouchacca [1971]). Conidia are glabrous, broadly el-lipsoidal to subglobose, occasionally pyriform, 6.7–10.3 3 5.2–7.6(–7.9) mm.
Phylogenetic analysis.—Maximum parsimony analysisof 18S rDNA sequences (FIG. 7) yielded 524 equallyparsimonious trees based on 118 parsimony infor-mative characters, 303 steps in length with a consis-tency index (CI) of 0.518 and a retention index (RI)of 0.664. The trees showed that T. hygrophila strainCBS 102670 is related more closely to P. zonatum,including both the ex-type isolate and the rather dis-parate ATCC 62440 (5 CBS 480.86), than it is to thetype species of Leptodontidium, L. elatius. Some othertaxa possessing conidiogenesis similar to that seen inCBS 102670 (e.g. L. boreale, Sporothrix inflata) alsowere shown to be unrelated. Two relatively closelyrelated fungi on a well supported branch, Connersiarilstonii (originally Pseudeurotium rilstonii) and Pleu-roascus nicholsonii, do not have distinct conidial an-amorphs. Resolution of the phylogenetic position ofthese anomalous fungi, which appeared to makePseudeurotium paraphyletic in parsimony analysis butnot in neighbor-joining analysis (not shown), wasconsidered beyond the scope of the present study.
The branch separating T. hygrophila from the rel-

705SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
TA
BL
EII
I.D
iffe
ren
tial
char
acte
rsof
anam
orph
sin
clud
edin
the
gen
usT
eber
din
ia
Spec
ies
Dis
posi
tion
ofco
nid
ioge
nou
sce
llsTe
rmin
alco
nid
ioge
nou
sce
llsT
ypic
alco
nid
ial
size
ran
ge(m
m)
Con
idia
lm
orph
olog
y
Teb
erdi
nia
hygr
ophi
laO
ften
onel
abor
ate
side
-bra
nch
-es
bear
ing
one
orm
ore
or-
ders
ofsu
b-br
anch
es,s
ome
inw
hor
ls;
also
solit
ary
orin
terc
a-la
ry
6–14
mm
lon
g,cy
lindr
ical
orba
sally
infl
ated
4.5–
5.5
32.
8–3.
1br
oadl
yel
lipso
idal
,el-
lipso
idal
toov
oid
Teb
erdi
nia
stat
eof
Pse
ud-
euro
tiu
mzo
nat
um
Solit
ary
oron
shor
t,si
ngl
esi
debr
anch
es;
mor
eel
abor
ate
stru
ctur
es,e
spec
ially
wh
orle
dsi
de-b
ran
ches
,rar
e;in
terc
alar
yco
nid
ioge
nou
sce
llsal
sopr
e-se
nt
9–22
mm
lon
g,la
gen
ifor
mor
cylin
dric
al5.
0–5.
73
3.2–
3.5
broa
dly
ellip
soid
al,e
l-lip
soid
alor
obov
oid
Teb
erdi
nia
stat
eof
Pse
ud-
euro
tiu
mba
keri
Mos
tly
solit
ary
term
inal
orin
ter-
cala
ryce
lls;
shor
tsi
debr
anch
-es
unco
mm
on
10–2
2m
m,
lage
nif
orm
5.0–
5.7
33.
2–3.
5su
bglo
bose
,elli
psoi
dor
obov
oid
Teb
erdi
nia
stat
eof
Pse
ud-
euro
tiu
mov
ale
(bot
hva
-ri
etie
s)
Mos
tly
inte
rcal
ary
cells
;so
me
solit
ary
cells
pres
ent
asw
ell
aste
rmin
alco
nid
ioge
nou
sce
llson
shor
t,si
mpl
esi
debr
anch
es
7–12
(240
)m
m,
cylin
dric
al4.
7–5.
23
3.8–
4.2
glob
ose,
subg
lobo
seor
obov
oid
Teb
erdi
nia
stat
eof
Pse
ud-
euro
tiu
mde
sert
oru
mSh
ort
(2–3
.53
1.5–
1.6
mm
)de
nti
cles
born
eon
vege
tati
veh
yph
ae
Non
epr
esen
t6.
7–10
.33
5.2–
7.6
(27.
9)m
mgl
abro
us,b
road
lyel
lip-
soid
alto
subg
lobo
se,
occa
sion
ally
pyri
form

706 MYCOLOGIA
FIG. 7. One of 644 equally parsimonious trees obtained from analysis of 18S ribosomal DNA using PAUP. Trees were 458steps in length with CI of 0.6812 and RI of 0.3188. Percentages of 50% or higher based on 1000 bootstrap replications aregiven on tree branches accentuated as bold lines. Higher taxa next to vertical bars are given according to Kirk et al (2001).Dotted sections of the vertical bars indicate anamorph taxa considered to be related to corresponding higher taxa.
atively closely adjacent L. elatius on the 18S dendro-gram lacked strong bootstrap support. Thereforephylogenetic analysis also was performed using ITSrDNA sequences.
Maximum parsimony analysis of ITS sequences
(FIG. 8) yielded two equally parsimonious trees basedon 71 parsimony informative characters, 139 steps inlength with a CI of 0.748 and a RI of 0.767. In thesetrees Teberdinia clustered into a well supported Pseud-eurotium clade (bootstrap: 94%). The number of base

707SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
FIG. 8. One of two equally parsimonious trees obtainedfrom analysis of ITS1-5.8S-ITS2 rDNA using PAUP. Treeswere 249 steps in length with CI of 0.8594 and RI of 0.7667.Percentages of 50% or higher based on 1000 bootstrap rep-lications are given on tree branches accentuated as boldlines.
TABLE IV. Number of differing base pairs among ITS rDNA sequences of different Pseudeurotium and Teberdinia strains
Species
CBS numbers
102670 102671 326.81 329.36 878.71 389.54 443.78
T. hygrophilaT. hygrophilaT. hygrophilaP. zonatumP. bakeriP. ovaleP. ovale var. milkoiP. desertorum
102670102671
326.81329.36878.71389.54443.78986.72
004256
11
04256
11
4256
11
49
1011
78
113
14 15
pairs distinguishing members of that clade in thecomplete ITS region are provided (TABLE IV). Aweakly supported clade (bootstrap 5 59%) wasformed by the Pseudeurotium taxa and some speciesclassified in the Myxotrichaceae, including Pseudo-gymnoascus roseus, Gymnostellatospora japonica andthe anamorphic species Geomyces pannorum, all ofwhich are morphologically distinct from Pseudeuro-tium/Teberdinia. Leptodontidium elatius, the type spe-cies of Leptodontidium, and L. boreale, both forminga strongly supported (bootstrap 5 99%) clade, wererelated more distantly to Pseudeurotium than were theMyxotrichaceous taxa despite their morphologicalsimilarity to Teberdinia anamorphs. The affinity of the
pseudeurotiaceous taxa to Pseudogymnoascus also wasreflected in 18S rDNA analysis.
ITS rDNA sequences of the three T. hygrophilastrains were shown to be almost identical (except forone deletion in the sequence of CBS 102670 at po-sition 455) and to be different from all available ex-type strains of Pseudeurotium species.
DISCUSSION
The standard use of sequence information in fungalanamorph taxonomy has not obviated the require-ment for anamorph generic names, particularly whenspecies not forming a teleomorph are discovered.The common, cosmopolitan Pseudeurotium speciessuch as P. ovale and P. zonatum were long known tohave a distinctive and morphologically enigmatic an-amorph (Domsch et al 1993), but the invariable ap-pearance of the teleomorph in cultures of these spe-cies precluded any need to coin a generic name forthese anamorphs. The appearance of T. hygrophilaalters this situation. Without a teleomorph it cannotbe classified as a Pseudeurotium species and an ana-morph genus is needed.
In proposing the genus Teberdinia, we have desig-nated T. hygrophila as type species in preference tothe anamorph of one of the relatively widely distrib-uted Pseudeurotium species. This was partly becauseT. hygrophila bore the only binomial in the genus butalso because of the classical nomenclatural consid-eration that it is the most extensively developed, mor-phologically distinctive Teberdinia species. Anotherfactor was that T. hygrophila was clearly situated inthe core group of the relevant clade.
The recognition of Teberdinia does not, to ourknowledge, conflict with any existing generic con-cept. The anamorph genera heretofore described asproducing single-celled blastoconidia grouped at theapices of unswollen or minimally swollen conidiogen-ous cells arising from nonmelanized conidiophoresare Sporothrix Hektoen & C.F. Perkins, Cerinosterus

708 MYCOLOGIA
R.T. Moore, Beauveriphora Matsushima, MicrohilumYip & Rath, and Leptodontidium de Hoog. The firstfour genera are distinguished readily in that they pro-duce conidia on denticles that remain distinct longafter conidial secession (de Hoog 1974, 1993, Benadeet al 1997, Matsushima 1975, Yip and Rath 1988).The denticles of Teberdinia often are indistinct fromthe beginning; moreover, in species other than theTeberdinia anamorph of P. desertorum, they soon areeffaced through swelling of the fertile apex after sev-eral conidia have been produced. Cerinosterus hasbeen shown to be a basidiomycetous anamorph withtypical dolipore septa (Smith and Batenburg-van derVegte 1985, Moore 1987). Two species of Sporothrix,including the type, S. schenckii, can be seen in phy-logenetic analysis of 18S rDNA to be unrelated toTeberdinia (FIG. 7). S. schenckii has long been knownto be ophiostomatalean in phylogenetic affinity(Summerbell et al 1993). Leptodontidium, although ahighly heterogeneous group, is overall the genusmost similar to Teberdinia based on morphologicalcomparison. Our observations on L. elatius, the typespecies, show that it produces two types of conidio-phores; one of them is indeed similar to typical Te-berdinia conidiophores while the other features aRhinocladiella-like long, rigid, dark-walled stalk bear-ing a sympodially elongating conidiogenous rachis.Teberdinia lacks the latter type. The core group ofLeptodontidium species including L. elatius appear in18S rDNA analysis to be related to the discomyceteBulgaria inquinans; this affinity also is reflected inthe description and photograph of the anamorph ofB. inquinans published by Fenwick (1992). Most ofthe other anamorph species placed in Leptodontidiumproduce some conspicuously melanized structures;colonies generally appear black in age (de Hoog1977). Teberdinia colonies as such generally lack mel-anized structures, although the peridia of subse-quently developing Pseudeurotium ascomata areblack. In mating trials with T. hygrophila, in whichplates were held for up to 1 y under ultraviolet light,it was noted that conidia eventually became mela-nized (and thus were deceptively suggestive of liber-ated ascospores) but some months were needed forthis development. Certainly no possible confusionwith Leptodontidium species arises as a result. No de-scribed Leptodontidium species represented in CBShas been found to have rDNA sequences compatiblewith those of Teberdinia (unpubl data).
It is possible that T. hygrophila, although failing toform mature ascomata in culture, might do so in na-ture. We cannot exclude the possibility that T. hygro-phila corresponds to one of the described Pseudeu-rotium species that is unavailable in living culture.
Further efforts are needed to obtain new collectionsand isolates of these species.
T. hygrophila was relatively common within thenorth temperate montane habitat (alpine fens, Te-berda Reserve) from which it was isolated. It mightbe found to be a relatively common fungus in similarhabitats worldwide. Certainly none of the relatedPseudeurotium species to date has been shown to haveany plant host specificity or other specific relation-ships limiting its distribution within the climate zonein which it is typically found in soils. The difficulty ofinterpreting T. hygrophila’s conidiogenesis under thelight microscope makes it a singularly difficult organ-ism for users of morphological keys to deal with, andthis might have hindered its recognition in the past.It must be cautioned that relatively morphologicallycomplex Teberdinia anamorphs obtained from natu-ral substrata cannot be assumed to be T. hygrophilawhenever they fail to form Pseudeurotium cleistothe-cia in culture. Purely conidial strains suggestive ofPseudeurotium anamorphs have been isolated repeat-edly from soil (Gams unpubl) and it is not clearwhether some of these isolates correspond to unde-scribed species or to asexual variants of commonPseudeurotium species. We hope that the descriptionof T. hygrophila as well as the extended descriptionsof other Pseudeurotium anamorphs will aid recogni-tion of these species in biodiversity studies and there-fore promote further understanding of the diversityof anamorphs and teleomorphs in this group.
ACKNOWLEDGMENTS
We thank A van Iperen and M Starink for technologicalassistance. The Studienstiftung Mykologie, Koln, and theFederation of European Microbiological Societies arethanked for supporting the studies of MV Sogonov.
LITERATURE CITED
Belyakova LA. 1969. Novaya raznovidnost’ griba Pseudeuro-tium ovale Stolk, vydelennaya iz pochvy (A new soil-borne variety of Pseudeurotium ovale Stolk.) Nov SistNizh Rast 6:103–107.
Benade E, Wingfield MJ, van Wyk PS. 1997. Conidium de-velopment in Sporothrix anamorphs of Ophiostoma. My-col Res 101:1108–1112.
Booth C. 1961. Studies of Pyrenomycetes: VI. Thielavia,with notes on some allied genera. Mycol Pap 83:1–15.
Cain RF. 1961. Studies of coprophilous ascomycetes VII.Preussia. Can J Bot 39:1633–1666.
Cejp K, Milko AA. 1964. Rody celedi Eurotiaceae s 32 spor-ami ve vrecku—I. Westerdijkella (Genera of the Euroti-aceae with 32 ascospores—I. Westerdijkella). Ceska My-kol 18:82–84.
Chattopadhyay SB. 1957. Pseudeurotium indicum (Chatto-

709SOGONOV ET AL: TEBERDINIA ANAMORPHS IN PSEUDEUROTIUM CLADE
padhyay & Das Gupta) comb. nov. Trans Br Mycol Soc40:460.
, Das Gupta C. 1957. Thielavia indica sp. nov. TransBr Mycol Soc 40:277–278.
de Hoog GS. 1974. The genera Blastobotrys, Sporothrix, Cal-carisporium and Calcarisporiella gen. nov. Stud Mycol 7:1–84.
. 1977. Rhinocladiella and allied genera. Stud Mycol15:1–140.
. 1993. Sporothrix-like anamorphs of Ophiostoma spe-cies and other fungi. In: Wingfield MJ, Seifert KA, Web-ber JF, eds. Ceratocystis and Ophiostoma. St Paul, Min-nesota, USA: APS Press. p 53–60.
, Gerrits van den Ende AHG. 1998. Molecular diag-nostics of clinical strains of filamentous Basidiomycetes.Mycoses 41:183–189.
Dijksterhuis J, Harder W, Wyss U, Veenhuis M. 1991. Colo-nisation and digestion of nematodes by the endopara-sitic nematophagous fungus Drechmeria coniospora. My-col Res 95:87–878.
Domsch KH, Gams W, Anderson T-H. 1993. Compendiumof soil fungi (reprint edition). Germany, Eching: IHW-Verlag. 860 p. (Vol. 1) 1 405 p. (Vol. 2—references).
Fenwick GA. 1992. A conidial form of Bulgaria inquinans.Mycologist 6:177–179.
Gams W, Hoekstra ES, Aptroot A, van der Aa HA, BoekhoutT, de Cock AWAM, de Hoog GS, Samson RA, SmithMT, Stalpers JA, Tan CS, Verkley GJM. 1998. CBScourse of mycology. Baarn, Delft: Centraalbureau voorSchimmelcultures. 165 p.
Gargas A, Taylor JW. 1992. Polymerase chain reaction(PCR) primers for amplifying and sequencing nuclear18S rDNA from lichenized fungi. Mycologia 84:589–592.
Gernandt DS, Platt JL, Stone JK, Spatafora JW, Holst-JensenA, Hamelin RC, Kohn LM. 2001. Phylogenetics of Hel-otiales and Rhytismatales based on partial small subunitnuclear ribosomal DNA sequences. Mycologia 93:915–933.
Gerrits van den Ende AHG, de Hoog GS. 1999. Variabilityand molecular diagnostics of the neurotropic speciesCladophialophora bantiana. Stud Mycol 43:151–162.
Kirk PM, Cannon PF, David JC, Stalpers JA. Ainsworth &Bisby’s dictionary of the fungi. 9th ed. Wallingford,Oxon, UK: CAB International. 655 p.
Kornerup A, Wanscher JH. 1978. Methuen handbook ofcolour. 3rd ed. London: Eyre Methuen Ltd. 252 p.
Lodha BC. 1971. Studies on coprophilous fungi IV. Somecleistothecial ascomycetes. J Indian Bot Soc 50:196–208.
Malloch D. 1974. Connersia rilstonii. Fungi canadenses 32:1–2.
, Cain RF. 1970. Five new genera in the family Pseu-deurotiaceae. Can J Bot 48:1815–1825.
Matsushima T. 1975. Icones Microfungorum a Matsushima
lectorum. Kobe: Published by the author. 209 p 1 415pl.
. 1996. Matsushima mycological memoirs No. 9.Kobe: Published by the author. 30 p 1 70 pl.
Montemartini Corte A. 1968. Eurotiaceae trovate in risaia.Giornale Botanico Italiano 102:55–65.
Moore RT. 1987. Micromorphology of yeasts and yeast-likefungi and its taxonomic implications. Stud Mycol 30:203–226.
Mouchacca J. 1971. Pseudeurotium desertorum sp. nov. RevMycol 36:123–127.
O’Donnell K. 1993. Fusarium and its near relatives. In:Reynolds DR, Taylor JW, eds. The fungal holomorph:mitotic, meiotic and pleomorphic speciation in fungalsystematics. Wallingford: CAB International. p 225–233.
Panasenko VT. 1964. Some new species of fungi on starchfrom the Ukraine. Mycologia 56:58–63.
Rai JN, Tewari JP. 1962. Pseudeurotium globosum n. sp., anew ascomycete from Indian soils. Mycopath et MycolApplic 16:90–96.
. 1963. On some isolates of the genus Preussia Fuckelfrom Indian soils. Proc Indian Acad Sci B 57:45–55.
Raper JR, Raper CA. 1972. Genetic analysis of the life cycleof Agaricus bisporus. Mycologia 64:1088–1117.
Samson RA, Stalpers JA, Verkerke W. 1979. A simplifiedtechnique to prepare fungal specimens for scanningelectron microscopy. Cytobios 24:7–11.
Smith MT, Batenburg-van der Vegte WH. 1985. Ultrastruc-ture of septa in Blastobotrys and Sporothrix. Antonie vanLeeuwenhoek 51:121–128.
Sogonov MV, Velikanov LL. 2004. Soil microfungal com-munities. In: Onipchenko VG, ed. Alpine ecosystemsin the Northwest Caucasus. Dordrecht: Kluwer. p 271–283.
Stolk AC. 1955. The genera Anixiopsis Hansen and Pseu-deurotium van Beyma. Antonie van Leeuwenhoek J Mi-crobiol Serol 21:65–79.
Suh SO, Blackwell M. 1999. Molecular phylogeny of thecleistothecial fungi placed in Cephalothecaceae andPseudeurotiaceae. Mycologia 91:836–848.
Summerbell RC, Krajden S, Kane J, Duke EE. 1993. Medi-cally important members of the Ophiostomatales/Spo-rothrix spp. In: Wingfield MJ, Seifert KA, Webber JF,eds. Ceratocystis and Ophiostoma. Taxonomy, Ecologyand Pathogenicity. St Paul, Minnesota, USA: APS Press.p 185–192.
Swofford DL. 2000. PAUP*. Phylogenetic Analysis UsingParsimony (*and other methods). Version 4.0b4a. Sun-derland, Massachusetts: Sinauer Associates.
van Beyma thoe Kingma FH. 1937. Beschreibung einigerneuer Pilzarten aus dem Centraalbureau voor Schim-melcultures Baarn (Holland), IV. Mitteilung ZentralblBakt II Abt 96:411–432.
Yip H-Y, Rath AC. 1989. Microhilum oncoperae gen. et sp.nov. (Deuteromycotina: Hyphomycetes): The cause ofmycosis of the larvae of Oncopera intricata (Lepidop-tera: Hepialidae). J Invert Pathol 53:361–364.





























