Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR BIOLOGY, Jan. 2007, p. 120–134 Vol. 27, No. 10270-7306/07/$08.00�0 doi:10.1128/MCB.00815-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
The Activity and Stability of the Transcriptional Coactivator p/CIP/SRC-3Are Regulated by CARM1-Dependent Methylation�
Hina Naeem,1,2 Donghang Cheng,4 Qingshi Zhao,1 Caroline Underhill,1 Marc Tini,2,3
Marc T. Bedford,4 and Joseph Torchia1,2*Department of Oncology, London Regional Cancer Program,1 Department of Physiology and Pharmacology,2 and Department of
Microbiology and Immunology,3 The University of Western Ontario, London, Ontario, Canada, and University ofTexas M. D. Anderson Cancer Center, Department of Carcinogenesis, Smithville, Texas 78957 4
Received 8 May 2006/Returned for modification 18 June 2006/Accepted 5 October 2006
The transcriptional coactivator p/CIP(SRC-3/AIB1/ACTR/RAC3) binds liganded nuclear hormone receptorsand facilitates transcription by directly recruiting accessory factors such as acetyltransferase CBP/p300 andthe coactivator arginine methyltransferase CARM1. In the present study, we have established that recombinantp/CIP (p300/CBP interacting protein) is robustly methylated by CARM1 in vitro but not by other proteinarginine methyltransferase family members. Metabolic labeling of MCF-7 breast cancer cells with S-adenosyl-L-[methyl-3H]methionine and immunoblotting using dimethyl arginine-specific antibodies demonstrated thatp/CIP is specifically methylated in intact cells. In addition, methylation of full-length p/CIP is not supportedby extracts derived from CARM1�/� mouse embryo fibroblasts, indicating that CARM1 is required for p/CIPmethylation. Using mass spectrometry, we have identified three CARM1-dependent methylation sites locatedin a glutamine-rich region within the carboxy terminus of p/CIP which are conserved among all steroidreceptor coactivator proteins. These results were confirmed by in vitro methylation of p/CIP using carboxy-terminal truncation mutants and synthetic peptides as substrates for CARM1. Analysis of methylation sitemutants revealed that arginine methylation causes an increase in full-length p/CIP turnover as a result ofenhanced degradation. Additionally, methylation negatively impacts transcription via a second mechanism byimpairing the ability of p/CIP to associate with CBP. Collectively, our data highlight coactivator methylationas an important regulatory mechanism in hormonal signaling.
Nuclear hormone receptors are a superfamily of ligand-dependent transcription factors that mediate the effects ofsmall lipophilic hormones such as estrogen and retinoic acid.Chromatin-based assays examining promoter occupancy of nu-clear receptor (NR)-dependent genes have shown that tran-scriptional activation involves recruitment of a large repertoireof coactivators that interact with liganded NRs in a coordi-nated fashion (42). Among the first coactivators recruited inresponse to hormone are the steroid receptor coactivator(SRC) proteins that include the p300/CBP interacting protein(p/CIP), also known in human form as SRC-3/AIB1/ACTR/RAC3/TRAM1 (2, 8, 32, 51, 53), steroid receptor coactivator 1(SRC-1) (23, 41), and steroid receptor coactivator 2 (SRC-2),also known as GRIP1/TIF2 (22, 56). p/CIP is a well-establishedNR coactivator which has been shown to mediate the tran-scriptional effects of liganded NRs in a variety of contexts invitro as well as in vivo (34, 53). Downregulation of p/CIPexpression in intact cells has been shown to inhibit cell growth(35, 52, 65, 66), and the targeted disruption of the p/CIP genein mice has demonstrated a requirement for p/CIP in mam-mary gland development as well as several nuclear receptor-dependent and -independent pathways (4, 30, 59, 61). Impor-tantly, the human homologue of p/CIP is amplified in apercentage of breast and ovarian cancers (2), and overexpres-
sion of p/CIP has been shown to initiate tumor development inmice (54).
Biochemical and functional studies have shown that p/CIPfunctions as a bridging factor that promotes the assembly oftranscriptional complexes essential for NR-mediated transac-tivation. This assembly is mediated by several domains withinthe p/CIP protein that are conserved in SRC-1 and GRIP/TIF2(17). The amino terminus contains a highly conserved basichelix-loop-helix and Per/ARNT/SIM domain which functionsas an interaction surface for a wide variety of proteins (24).The central region of the SRC proteins encodes the receptorinteracting domain consisting of three leucine-rich motifs (alsoknown as NR boxes) with the consensus amino acid sequenceLXXLL (where X is any amino acid and L is leucine). TheLXXLL motifs are responsible for making direct contact withliganded NRs (20, 37, 53, 55). The carboxy terminus of p/CIPcontains two transcriptional activation domains (AD1 andAD2) which serve as interaction surfaces for the recruitment oftwo major classes of coactivators. AD1 interacts directly withthe acetyltransferases CBP and p300 (8, 53) and with GCN5/PCAF(1, 5, 27, 38). AD2 serves as an interaction surface for asecond class of enzymatic proteins, known as protein argininemethyltransferases (PRMTs) (3). Eight mammalian PRMTshave been identified, although the coactivator arginine meth-yltransferase (CARM1/PRMT4), PRMT1, and, more recently,PRMT2 appear to be the most relevant with respect to signal-ing by NRs (28, 43). Cotransfection assays using transient orstably integrated reporter genes have shown that individualSRC proteins can synergize with histone acetyltransferases andmethyltransferases to activate NR-dependent transcription
* Corresponding author. Mailing address: Cancer Research Labo-ratories, London Regional Cancer Program, London, Ontario, CanadaN6A 4L6. Phone: (519) 685-8692. Fax: (519) 685-8673. E-mail: [email protected].
� Published ahead of print on 16 October 2006.
120
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

(26, 50, 57). CARM1 and PRMT1 can methylate specific ar-ginines on histones H3 and H4, respectively, and several stud-ies have demonstrated a correlation between methylation ofhistone H3, recruitment of CARM1, and activation of severalsteroid-responsive genes (11). Disruption of the CARM1 genein mice results in embryonic lethality (64), and estrogen recep-tor (ER)-dependent signaling is compromised in the CARM1null cells. Collectively, these studies suggest that recruitment ofCARM1 represents an essential activating step in mammaliangene transcription. The recent discoveries of lysine and argi-nine demethylases indicates that methylation, similar toacetylation (49, 58), represents a dynamic and reversible mod-ification and adds another level of complexity to our under-standing of transcriptional regulation. Importantly, argininemethylation is not restricted to posttranslational modificationof histones. There is mounting evidence supporting the hy-pothesis that in addition to methylating histone H3, the phys-iological role of CARM1 may be dependent on methylation ofnonhistone proteins. For example, CARM1 specifically meth-ylates CBP/p300, and mutation of the methylation sites in CBPabolishes steroid-dependent transcriptional activation (10, 31,62). In addition to histone H3 and p300/CBP, CARM1 meth-ylates proteins involved at several other critical points in thetranscriptional process including the poly(A)-binding protein(PABP1) (29), RNA binding molecules involved in transcriptstability (HuR and HuD) (15), and the core splicing factorSmB (12).
In the present study we have characterized the associationbetween p/CIP and the methyltransferase protein CARM1.We demonstrate that the association between p/CIP andCARM1 occurs on target genes in response to hormone and isnot detectable in solution following immunopurification ofp/CIP. In addition, we show that p/CIP is a substrate forCARM1 in vitro and in vivo. Using a combination of massspectrometry and molecular mapping, we have identified sev-eral CARM1-dependent methylation sites in p/CIP. At leastthree sites are found within a highly conserved glutamine-richregion of the carboxy-terminus of p/CIP, and additional sitesare also located in the central region of p/CIP. Our resultssuggest that p/CIP methylation by CARM1 may have a dualrole in regulating p/CIP function. First, arginine methylationappears to reduce protein stability and causes an increase inp/CIP turnover. Second, we demonstrate that arginine meth-ylation represents a critical regulatory signal involved in coor-dinating the dissociation of CBP and p/CIP. Collectively, theresults of our study suggest that arginine methylation repre-sents an important posttranslational modification which maybe important for regulating coregulator stability as well as thedynamics of nuclear receptor-mediated transcription.
MATERIALS AND METHODS
Plasmids and reagents. The cDNAs encoding full-length CARM1 were kindlyprovided by Michael Stallcup (University of California-Los Angeles), andPRMT1 cDNA was provided by Harvey Herschman (University of California-Los Angeles). The baculovirus vectors for p/CIP, p/CIP deletion mutants,CARM1, CBP, and PRMT1 were subcloned into a Fastbac vector using standardcloning techniques. For the Gal4 transfection assays, p/CIP fragments weregenerated by PCR using PCR Supermix High Fidelity (Invitrogen) and sub-cloned in frame into a PMgal (Stratagene) expression vector. Antibodies againstCARM1 and CBP were purchased from Santa Cruz Biotechnologies. Antibodyagainst human ER� was obtained from Bethyl Laboratories. Anti-p/CIP anti-
body was generated as previously described (44). The experiments shown in Fig.3C and Fig. 7 were performed using an anti-SRC-3 antibody obtained fromBethyl Laboratories. �mR17 antibodies were purchased from UpState. Thep/CIPR3A cDNA (containing substitutions of arginine to alanine at amino acids1178, 1184, and 1195) and the p/CIPR6A (containing substitutions of arginine toalanine at amino acids 1163,1168, 1170, 1178,1184, and 1195) were generated bysite-directed mutagenesis of p/CIP using a Quickchange kit (Stratagene) accord-ing to the manufacturer’s instructions. S-adenosyl-L-[methyl-3H]methionine([3H]SAM) (80 Ci/mmol) was purchased from Amersham and [35S]methionine(419 mBq/ml) was purchased from Perkin Elmer. Biotin-labeled peptides wereobtained commercially from Biosynthesis Incorporated.
Cell lines and transfections. HeLa, COS-1, CV-1, MCF-7, U2OS, CARM1�/�,and CARM1�/� mouse embryonic fibroblasts (MEFs) were maintained in Dul-becco modified Eagle medium (DMEM) supplemented with 10% fetal bovineserum (FBS), 0.1 U/ml penicillin G, and 0.1 �g/ml streptomycin sulfate in 5%CO2 at 37°C. To assess transcriptional activity of Gal4-p/CIP fusion proteins,transient transfections were performed using Lipofectamine 2000 according tothe manufacturer’s instructions (Invitrogen). COS-1 cells were seeded at approx-imately 180,000 cells/well in 24-well plates. A total of 100 ng of the pMGal vector(cytomegalovirus) directing the expression of either the Gal4 DNA bindingdomain (DBD) alone and either p/CIP deletions or mutants fused in frame withGal4 DBD were cotransfected with 250 ng of luciferase reporter plasmid con-taining five binding sites for Gal4 upstream of a minimal promoter. Renillaluciferase (0.5 ng) was used to monitor transfection efficiency, and total DNAwas adjusted to 500 ng per well with pBKS vector. Forty-eight hours aftertransfection, cells were harvested, and cell extracts were prepared in 75 �l of 1�reporter lysis buffer (Promega). Ten-microliter aliquots of the extract were usedfor subsequent determination of luciferase and Renilla activity according to themanufacturer’s instructions (Promega). Luciferase values were normalized to theoutput of the internal Renilla control plasmid, and the relative activation (n-fold)was calculated relative to the level of pMGal alone, which was arbitrarily set to1. Luciferase activities are shown as the means and standard errors of at leastthree independent experiments performed in triplicates.
To analyze the expression levels of Gal4 fusion proteins, 4 �g of each proteinwas transfected with Lipofectamine 2000 into COS-1 cells according to themanufacturer’s protocol (Invitrogen). Prior to transfection, COS-1 cells wereseeded at 90% confluence in six-well plates. Forty-eight hours posttransfection,cells were harvested and lysed with radioimmunoprecipitation assay lysis buffer(50 mM Tris, pH 8.0, 1 mM EDTA pH 8.0, 150 mM NaCl, 0.1% sodium dodecylsulfate [SDS], 1.0% NP-40, 0.5% sodium deoxycholate, and protease inhibitors).Bradford protein assays were performed to determine the concentration ofwhole-cell extracts. Twenty micrograms of each extract was subjected to SDS-polyacrylamide gel electrophoresis (PAGE) followed by Western blotting withanti-Gal antibody (Santa Cruz). Antitubulin antibody (Sigma) was used as load-ing control.
To assess the significance of methylation in estrogen signaling, cells weremaintained in phenol red-free DMEM supplemented with 10% charcoal-stripped FBS for 48 h. A day before transfection, cells were seeded at �75,000cells/well in 24-well plates. Twenty-five nanograms of wild-type p/CIP and ofp/CIPR6A cDNA was cotransfected with 200 ng of a luciferase reporter con-taining three copies of an estrogen response element (ERE) upstream of aminimal promoter and 10 ng of ER. For some experiments, increasing amountsof wild-type p/CIP or p/CIPR6A (50, 100, and 200 ng) were cotransfected with200 ng of a pS2 luciferase reporter and 10 ng of ER. Total DNA was adjusted to800 ng/well with empty vector, and transfections were performed using Lipo-fectamine 2000 according to the manufacturer’s protocol (Invitrogen).
Twenty-four hours after transfection, cells were stimulated with 1 �M �-estradiol for 16 h. Cells were lysed with 75 �l of 1� reporter lysis buffer (Promega),and a 20-�l aliquot of the whole-cell extract was used for luciferase activityanalysis. Luciferase activities are shown as the means � standard errors of threeindependent samples.
Immunoprecipitation, Western blot analysis, and fluorography. Cells werewashed twice in phosphate-buffered saline (PBS), harvested, and lysed in lysisbuffer (�300 �l/10-cm plate) consisting of 20 mM Tris (pH 7.9), 300 mM KCl,0.1% NP-40, 10% glycerol, 0.1 mM dithiothreitol (DTT), 0.5 mM EDTA, 0.5mM EGTA, and protease inhibitor cocktail. Extracts were then centrifuged for10 min at 16,000 � g at 4 C, and the soluble extracts were retained. Samples werenormalized for protein content, followed by the addition of the appropriateantibody for 2 h on ice. Samples were then diluted to 500 �l with lysis buffer, and50 �l of a 50% slurry (vol/vol) of protein A/G agarose was added overnight withrocking. The antibody complexes were washed extensively with lysis buffer, andthe bound complexes were separated by SDS-PAGE, transferred to nitrocellu-lose membrane, and blocked overnight in PBS containing 0.1% Tween 20 and
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 121
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

5% nonfat dried milk. The appropriate antibodies were then diluted in blockingbuffer, and the membrane was probed for 2 h at room temperature with rocking,followed by the appropriate secondary antibody for 1 h. Proteins were detectedusing enhanced chemiluminescence according to the manufacturer’s recommen-dations (Amersham). For experiments involving the analysis of 35S-labeled pro-teins, immunoprecipitated proteins were separated by SDS-PAGE and analyzedby fluorography. Gels were then treated with ENHANCE, dried, and exposed forseveral days to film. For the experiments shown in Fig. 3C and 7, cells were grownfor 8 days in the presence of AdOx prior to immunoprecipitation of p/CIP.
Purification and analysis of p/CIP. Nuclear extracts were dialyzed againstbuffer A (20 mM Tris [pH 7.9], 0.5 mM EDTA, 0.5 mM EGTA, 10% glycerol, 0.5mM DTT, 0.2 mM phenylmethylsulfonyl fluoride, and 5 �g/ml of leupeptin,aprotinin, and pepstatin) containing 100 mM KCl. To purify p/CIP, the nuclearextract was loaded onto a P11 phosphocellulose column preequilibrated in thesame buffer. The flowthrough was collected, and the column was washed sequen-tially with buffer A containing increasing concentrations of KCl. The 0.1 Mfraction containing p/CIP was precipitated with 20 to 60% ammonium sulfate,and the precipitated proteins were resuspended in 4 ml of buffer A containing100 mM KCl. This was then dialyzed against the same buffer to remove residualammonium sulfate before the sample was applied to a Sephacryl S300 gel filtra-tion column. The column was washed with buffer A at a flow rate of 0.4 ml/min.Fractions were collected, pooled, and analyzed for p/CIP by Western blotting.The p/CIP-containing fractions were pooled and dialyzed against buffer A con-taining 100 mM KCl. For immunoaffinity purification of p/CIP, affinity-purifiedp/CIP antibody was cross-linked to protein A-Sepharose using dimethylpalmili-date according to standard procedures (19). Fractions from the gel filtration stepwere pooled and precleared by passing the pooled fractions through a controlaffinity column containing anti-rabbit immunoglobulin G. The eluant was thenloaded onto the anti-p/CIP affinity column at a flow rate of 0.2 to 0.5 ml/min. Theflowthrough was collected and reloaded on the column five times prior to elutionof the bound proteins with 100 mM glycine (pH 3.0). For mock-purificationexperiments, samples from the gel filtration step were loaded onto proteinA-Sepharose cross-linked to an irrelevant antibody. Normally, protein sampleswere fractionated by SDS-PAGE and transferred to nitrocellulose membrane,and specific proteins were analyzed by Western blotting with the appropriateantibodies as indicated. For the purification of p/CIP from MEFs, cells weregrown to confluence on 10- by 150-mm plates and harvested, and nuclear extractswere prepared as described above. Extracts were then passed directly throughthe anti-p/CIP immunoaffinity column as described above and then subjected toSDS-PAGE and Western blotting.
Expression and purification of recombinant proteins. FLAG- or hemaggluti-nin-tagged proteins were generated using the Bac-to-Bac baculovirus expressionsystem according to the manufacturer’s instructions (Invitrogen). Proteins weresubcloned into the Fastbac expression vectors and transformed into DH10 bac-teria. The resulting bacmids were transfected into Sf9 cells to produce recom-binant baculovirus, which was amplified and used to infect Sf9 cells at a multi-plicity of infection of 5. After 48 h, cells were lysed, and recombinant proteinswere purified by immunoaffinity with anti-Flag M2 affinity resin essentially asdescribed previously. After extensive washing, proteins were eluted with 20 mMTris buffer, pH 7.9, 100 mM KCl, 10% glycerol, 0.5 mM EDTA, and 0.2 mg/mlof the appropriate peptide competitor. Proteins were then dialyzed against elu-tion buffer without peptide, frozen, and stored at �80 C.
Glutathione S-transferase (GST) fusion proteins containing various regions ofp/CIP were purified on glutathione agarose beads following standard procedures.Regions of p/CIP were subcloned into the pGEX bacterial expression vector inframe with the GST moiety, and the resulting plasmid was transformed into theEscherichia coli strain BL-21. The overnight culture was diluted 1:100 in 100 to300 ml of selection medium and grown at 37°C with shaking until an opticaldensity at 600 nm of �0.5 was reached, at which time protein expression wasinduced with 1.0 mM isopropyl-�-D-thiogalactopyranoside (final concentration)for 5 h at room temperature. Cells were pelleted by centrifugation at 4°C for 10min at 10,000 � g and resuspended in 10 ml of resuspension buffer per 100 ml ofstarting culture volume prior to incubation on ice for 30 min with occasionalmixing. The resuspended cells were then lysed by sonication prior to centrif-ugation of the samples at 4°C for 1 h at 30,000 � g. A total of 500 �l ofGST-Sepharose beads prewashed in resuspension buffer was added to the clar-ified supernatant, and the samples were incubated at 4°C with rocking for 1 h.The beads were pelleted by centrifugation at 4°C for 2 min at 1000 rpm andsubsequently washed five times with 12 ml of resuspension buffer. The beadswere resuspended in 1 ml of resuspension buffer containing 10 mM reducedglutathione and incubated for 5 min with rocking. The supernatants containingthe eluted proteins were dialyzed against a buffer consisting of 20 mM Tris, pH
7.9, 100 mM KCl, 10% glycerol, 0.5 mM EDTA, and 0.5 mM DTT and stored at�80°C.
Methylation assays. In vitro methylation reactions were performed using 0.2 to1 �g of either purified proteins or biotinylated peptides as substrates and 100 ngof purified CARM1 protein. Reactions were performed at 30°C for 1 h in 20 �l(final volume) of reaction buffer consisting of 50 mM Tris-HCl, pH 8.5, 20 mMKCl, 10 mM MgCl2, 1 mM �-mercaptoethanol, and 0.1 M sucrose and 5 �Ci of[3H]SAM. Upon completion of the methylation reactions, biotin-labeled pep-tides were purified with MagnaSphere streptavidin beads (Promega) for 1 h at4°C with rotation. Beads were washed three times with wash buffer (50 mM Tris,pH 8, 150 mM NaCl, 5 mM EDTA, 0.5% NP-40), and radioactivity was measuredby scintillation counting.
For in vivo labeling of methylated proteins, MCF-7 cells were grown for 24 hin DMEM without methionine and containing stripped serum, followed by theaddition of L-[methyl-3H]methionine (50 �Ci/ml) for 1 h. Cells were then har-vested, lysed, and p/CIP immunopurified as previously described (36).
To analyze methylation of p/CIP by mass spectrometry, a 100-ml culture of Sf9cells (1.5 � 106 cells/ml) was infected with FLAG-p/CIP baculovirus alone orcoinfected with FLAG-CARM1. At 48 h postinfection, the cells were harvestedand resuspended in a 50 mM Tris buffer (pH 7.9) containing 300 mM KCl and0.1% NP-40 and disrupted by 10 strokes with a dounce homogenizer; the celllysate was then clarified by centrifugation at 100,000 � g for 30 min. To each ofthe cell extracts, anti-FLAG-agarose was added and incubated for 2 h at 4 C;cells were washed extensively with lysis buffer, with a final wash with buffercontaining 100 mM KCl and 0.1% NP-40. The bound proteins were eluted withbuffer containing 1.0 mg/ml FLAG peptide, and the coimmunopurified com-plexes were separated by SDS-PAGE. The 160-kDa band corresponding top/CIP was excised and analyzed by mass spectrometry (MS) performed at theSouthern Alberta Mass Spectrometry Center (Calgary, Alberta, Canada).
Chromatin immunoprecipitation (ChIP). MCF-7 cells grown in phenol red-free DMEM containing 10% charcoal-stripped FBS for 48 h prior to stimulationwith 10�7 M �-estradiol for the time periods indicated in Fig. 1C. Cells were thencross-linked with 1% formaldehyde at room temperature for 10 min. Cross-linking was terminated by adding glycine to a final concentration of 125 mM andincubating for 5 min. Cells were washed twice with ice-cold PBS containing 0.5mM EDTA and harvested. Cells pellets were lysed in 0.3 ml of cell lysis buffer (50mM Tris-HCl [pH 8.1], 10 mM EDTA, 1% SDS, and protease inhibitors) andincubated on ice for 10 min. Cell lysates were sonicated to yield DNA fragmentsranging in size from 300 to 1,000 bp. Supernatants were diluted 10-fold indilution buffer (20 mM Tris-HCl [pH 8.1], 1% Triton X-100, 2 mM EDTA, 150mM NaCl, and protease inhibitors) and precleared with 60 �l of a 50% slurry ofprotein A-Sepharose containing 2.5 �g of sheared salmon sperm DNA for 2 h at4°C. Immunoprecipitation was performed overnight at 4°C with 1.5 to 4 �g of theantibodies. A total of 60 �l of protein A-Sepharose containing 2.5 �g of salmonsperm DNA per ml was added to the solution and incubated for 1 h at 4°C. Thebeads were washed one time with buffer A (20 mM Tris-HCl [pH 8.1], 150 mMNaCl, 2 mM EDTA, 0.1% SDS, 1% Triton X-100), four times with cold buffer B(20 mM Tris-HCl [pH 8.1], 500 mM NaCl, 2 mM EDTA, 0.1% SDS, 1% TritonX-100), and one time with LiCl-detergent buffer (10 mM Tris-HCl [pH 8.1], 0.25M LiCl, 1% NP-40, 1% deoxycholate, 1 mM EDTA). Finally, the samples werewashed three times with Tris-EDTA buffer. Immunocomplexes were extractedtwice with 200 �l of elution buffer (1% SDS–0.1 M NaHCO3). NaCl was addedto a final concentration of 200 mM, and the cross-linking was reversed by heatingat 65°C overnight. The protein was digested by adding Tris (pH 6.5) to a finalconcentration of 40 mM and 20 �g of proteinase K and incubating at 45°C for1 h. The DNA was purified using QIAGEN PCR purification spin columns. PCRwas performed using primers specific to the human pS2 promoter consisting ofthe forward primer 5-GGCCATCTCTCACTATGAATCATTCTGC-3 and thereverse primer 5-GGCAGGCTCTGTTTGCTTAAAGAGCG-3. PCR condi-tions were as follows: initial denaturing cycle of at 94°C for 3 min, followed by 35cycles of 94°C for 30 s, 52°C for 30 s, and 72°C for 1 min, with a final elongationstep of 72°C for 10 min. The PCRs were quantified by scanning densitometry.
Pulse-chase labeling experiments. MEFs were grown to approximately 80%confluence and grown in DMEM methionine-cysteine-free medium for 1 h andthen replaced by medium containing methionine for 40 min. Cells were thenwashed twice in PBS and incubated with medium containing 2 mM cold methi-onine-cysteine. Cells were harvested at the indicated times (see Fig. 6C and D),and p/CIP was immunoprecipitated from the cell lysates using p/CIP antibody.For cells overexpressing p/CIP, 2- by 15-cm plates were transfected with 10 �g ofcDNA for 24 h. The transfected cells corresponding to each cDNA were pooled,replated (3 � 106 cells/10-cm plate), and allowed to adhere overnight, andpulse-chase experiments were performed the following day as described above.
122 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

RESULTS
Target gene assembly of p/CIP and CARM1 coactivatorcomplexes. CARM1 was initially identified based on its abilityto interact with the carboxy terminus of GRIP1 (7) and hasalso been purified as a component of a large multiproteincomplex from MCF-7 cells(63). To determine if p/CIP andCARM1 are stably associated in solution, we performed bio-chemical purification experiments of p/CIP and CARM1 usingHeLa nuclear extracts. Gel filtration chromatography of nu-clear extracts followed by Western blotting indicated that
p/CIP could be found in two chromatographically distinctpeaks, a large peak corresponding to a size of approximately 1MDa and a second, more abundant peak, migrating at approx-imately 600 kDa (Fig. 1A). Interestingly, CARM1 was identi-fied as a broad peak partially overlapping with the smaller600-kDa p/CIP component, whereas CBP was found in frac-tions corresponding to the larger 1-MDa peak. Fractions cor-responding to each peak were pooled and purified using ananti-p/CIP affinity column. Immunoblotting of the affinity-pu-rified p/CIP complexes indicated that CBP was associated with
FIG. 1. CARM1 associates with p/CIP on DNA. (A) HeLa nuclear extracts were separated by gel filtration chromatography, and fractions wereassayed by immunoblotting using the antibodies indicated on the left. (B) Fractions corresponding to peaks I and II shown in panel A were pooledand passed through an anti-p/CIP immunoaffinity column or immunoglobulin G (IgG) column as control. After extensive washing, bound proteinswere eluted using low pH buffer and assayed by immunoblotting using antibodies specific for p/CIP, CBP, or CARM1. (C) p/CIP and CARM1 bindto the endogenous PS2 promoter in vivo. ChIP analysis of the PS2 promoter in MCF-7 cells using antibodies against p/CIP and CARM1demonstrates that both p/CIP and CARM1 are recruited to the PS2 promoter in response to estradiol. MCF-7 cells were stimulated for varioustime periods. Cells were fixed, and sequential ChIPs were performed using an anti-p/CIP antibody, followed by reimmunoprecipitation (ReChIP)using a CARM1-specific antibody. Samples were analyzed by PCR using specific primers flanking the PS2 promoter, and the relative abundancewas quantified by densitometry.
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 123
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

p/CIP purified from the larger 1-MDa component (Fig. 1B). Incontrast, CARM1 was not retained by the anti-p/CIP affinitycolumn but was detected entirely in the flowthrough compo-nent of the 600-kDa (data not shown) fraction, suggesting that
CARM1 and p/CIP do not stably associate in solution in HeLacells. We also used ChIP to assess if p/CIP and CARM1 asso-ciate at the PS2 promoter in response to hormone (Fig. 1C).Following stimulation with estradiol, the cells were fixed withformaldehyde, and chromatin was immunoprecipitated with ap/CIP antibody, followed by an immunoprecipitation using aCARM1-specific antibody. These results indicate that bothp/CIP and CARM1 associate with the PS2 promoter in a hor-mone-dependent manner, with a peak occurring at approxi-mately 30 min, which is consistent with earlier observations(38, 47). Collectively, these results establish that the associa-tion between CARM1 and p/CIP occurs on target genes in thecontext of chromatin and that they most likely do not form astable complex in solution.
p/CIP is a substrate for CARM1-dependent methylation.Recent studies have shown that CARM1 is capable of meth-ylating nonhistone proteins, such as CBP, resulting in concom-itant changes in transcriptional regulation (10, 28, 62). Basedon the direct association between p/CIP and CARM1, we as-sessed whether p/CIP may also serve as a substrate forCARM1. In vitro methylation assays using purified proteins(Fig. 2A) indicated that both p/CIP and CBP are robustlymethylated by CARM1 (Fig. 2B). The p/CIP methylation isdirectly dependent on the methyltransferase activity ofCARM1 as a point mutation that abolishes methyltransferaseactivity was incapable of methylating either p/CIP or CBP (notshown). In contrast, neither p/CIP nor CBP is methylated bythe related family member PRMT1 (Fig. 2B). Furthermore,the addition of increasing amounts of free histones in themethylation reaction had no effect on p/CIP methylation, sug-gesting that p/CIP serves as a very efficient substrate forCARM1 (Fig. 2C).
To further define the specificity of the methylation reaction,we utilized whole-cell extracts from MEFs isolated fromCARM1 null mice as a source of methyltransferase activity.Purified recombinant p/CIP could not be methylated in vitrousing extracts from CARM1�/� MEFs, indicating that that noalternative pathway exists for p/CIP methylation (Fig. 3A).Experiments were also performed to assess whether p/CIP wasspecifically methylated in intact cells. First, MCF-7 cells weremetabolically labeled with [3H]SAM, followed by immunopre-cipitation using a p/CIP antibody. The results of the fluoro-graphic image indicated specific incorporation of the radioiso-tope into the immunoprecipitated p/CIP protein (Fig. 3B).Second, p/CIP was immunoprecipitated from cells grown in theabsence or presence of the methylation inhibitor periodateoxidized adenosine (AdOx). The immunoprecipitated proteinswere then immunoblotted with p/CIP antibody or with an an-tibody which specifically recognizes asymmetric dimethylargi-nines found on arginine 17 of histone H3 (�mR17). The anti-body, �mR17, has previously been shown to not only recognizethe methylated H3 but also display immunoreactivity with anumber of other proteins in a CARM1-dependent fashion(64). Both the anti-p/CIP antibody and the �mR17 antibodywere able to recognize the immunoprecipitated p/CIP protein(Fig. 3C). However, for cells grown in the presence of AdOx,a significant decrease in immunoreactivity was detected usingthe �mR17. Taken together, these results establish that p/CIPis a substrate for CARM1-dependent methylation both in vitroand in intact cells.
FIG. 2. In vitro methylation of p/CIP. (A) Epitope-tagged recombi-nant proteins were generated in Sf9 cells using baculovirus. Cells werethen harvested, and proteins were immunopurified using anti-FLAGSepharose. Approximately 500 ng of various purified proteins was thenanalyzed by SDS-PAGE and Coomassie blue staining. (B) Approximately500 ng of purified recombinant CBP, p/CIP, or specific histones wasincubated in the presence of 100 ng of either CARM1 or PRMT1 and[3H]SAM for 1 h. Proteins were then separated by SDS-PAGE andanalyzed by fluorography. (C) In vitro methylation assays were performedusing 500 ng of p/CIP, 100 ng of recombinant CARM1, and increasingconcentrations of histones as indicated at the top of the panel. Thereactions were terminated, proteins were separated by SDS-PAGE, andmethylation of p/CIP was monitored by fluorography. BSA, bovine serumalbumin.
124 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

CARM1-dependent methylation sites are found in the car-boxy terminus of p/CIP. To identify the specific amino acidsmethylated by CARM1, we overexpressed p/CIP, alone or incombination with CARM1, in Sf9 cells using a baculovirusexpression system. Forty-eight hours after infection, p/CIP wasaffinity purified, isolated by SDS-PAGE, and analyzed by ma-trix-assisted laser desorption ionization–time of flight MS. Themasses of 15 peptides were identified and found to match thepredicted peptide masses of p/CIP. Four were identified withincreased masses, consistent with the presence of dimethylgroups (Table 1). Three of the peptides were found to containlow levels of an arginine dimethylated form, regardless of the
presence of CARM1. Significantly, it was found that the rela-tive abundance of the methylated form of a peptide corre-sponding to amino acids (aa) 1176 to 1201 of p/CIP was di-rectly correlated with the coexpression of CARM1. Thispeptide was then analyzed further by liquid chromatography-tandem MS, and it was determined that the arginines locatedat positions 1178, 1184, and 1195 were all asymmetrically di-methylated in a CARM1-dependent manner.
To validate the results obtained by MS, C-terminal trunca-tion mutants of p/CIP were generated using baculovirus, puri-fied, and tested as substrates for methylation by CARM1 invitro (Fig. 4 A). Whereas a fragment spanning residues 1 to1398 was strongly methylated, deletion of residues 1085 to1398 dramatically decreased methylation. This is consistentwith the indicated sites of methylation determined by MS. Inaddition, we also employed biotinylated synthetic peptides assubstrates for in vitro methylation reactions. The peptides werebound to streptavidin-coated beads, washed, and analyzed byliquid scintillation counting. The peptide corresponding to aa1176 to 1201, which contained the CARM1-dependent meth-ylation sites identified by MS, was robustly methylated in vitro(Fig. 4B). Further inspection of this region indicated the pres-ence of three additional arginines at aa 1163, 1168, and 1170which lie immediately N-terminal to the region identified byMS. Although not identified by MS, in vitro methylation reac-tions using a synthetic peptide indicated that this region is alsomethylated by CARM1 in vitro (Fig. 4B).
Finally, mapping studies were performed using recombinantGST proteins fused to regions of p/CIP spanning the full-lengthprotein. Using the GST fusion proteins as substrates, two addi-tional regions of p/CIP, corresponding to aa 819 to 853 and 920 to968, were found to serve as highly efficient substrates for CARM1in vitro (Fig. 5A). Further mapping using biotinylated peptidesindicated that aa 844 to 865, containing arginines at aa 839 and844, was robustly methylated by CARM1 in vitro (Fig. 5B). Asecond peptide containing an arginine at aa 961 was also meth-ylated, although to a lesser degree. To determine which arginineis specifically methylated within aa 844 to 865, individual arginineswere mutated to alanine. Mutation of the amino acid at position839 completely abolished CARM1-dependent methylation,whereas mutation of arginine to alanine at aa 844 diminishedmethylation by approximately 50%, suggesting that arginine 844may be involved in substrate recognition by CARM1 (Fig. 5C,center panel). Collectively, the results of our biochemical analysisindicate two methylated regions in p/CIP: methylation domain 1(MD1) stretching from R1163 to R1195 and MD2 stretching fromR839 to R961.
Phosphorylation at S847 antagonizes p/CIP methylation.Amino acids at positions 839 and 844 are found directly adjacentto a region of p/CIP containing several conserved serines, at aa847 and 850, which have recently been identified as physiologi-cally relevant phosphorylation sites (60). The proximity of thesephosphorylation sites to R839 suggests a potential for cross talkbetween methylation and phosphorylation of p/CIP. To test thishypothesis, we generated a peptide phosphorylated at aa 847which has been shown to be phosphorylated by IB kinase inresponse to hormone treatment (60). Phosphorylation at aa 847decreased the methylation at aa 839 by approximately fivefold,indicating that prior phosphorylation antagonizes methylation ataa 839 in vitro (Fig. 5C, right panel).
FIG. 3. p/CIP is methylated in intact cells. (A) Extracts fromCARM1�/� MEFs do not support methylation of p/CIP in vitro. Ap-proximately 500 ng of purified recombinant p/CIP protein was incu-bated with extracts derived from either CARM1�/� or CARM1�/�
MEFs in the presence of [3H]SAM. Methylation reactions were thenanalyzed by SDS-PAGE and fluorography. (B) MCF-7 cells were la-beled for 1 h with L-[methyl-3H]methionine (50 �Ci/ml) for 1 h, fol-lowed by cell lysis and immunoprecipitation of p/CIP (36). The com-plexes were then separated by SDS-PAGE and analyzed byfluorography. (C) MCF-7 cells were grown in the presence and absenceof AdOx (20 �M) for 8 days, and p/CIP was immunoprecipitated. Theimmunoprecipitated complexes were separated by SDS-PAGE andprobed with either an anti-p/CIP antibody (�p/CIP) or an antibodyraised against dimethylated arginine 17 on histone H3 (�mR17).
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 125
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

p/CIP turnover is regulated by methylation. To investigatethe functional relevance of p/CIP methylation of the carboxyterminus (MD1), we generated substitutions of arginine toalanine at R1178, R1184, and R1195, yielding (p/CIPR3A, andat R1163, R1168, R1170, R1178, R1184, and R1195, yieldingp/CIPR6A. Methylation reactions using purified recombinantp/CIP3A and p/CIP6RA proteins indicated a significant de-crease in CARM1-dependent methylation in vitro (Fig. 6A).Surprisingly, transient expression of the methylation site mu-tants in COS-1 cells consistently yielded higher protein levelscompared to wild-type p/CIP, suggesting that methylation ofp/CIP may play a role in p/CIP turnover (Fig. 6B). This wasalso reflected in the amount of mutant protein immunopre-cipitated relative to wild-type p/CIP. In order to examine theeffect of CARM1-mediated methylation on p/CIP stability,we conducted pulse-chase experiments in CARM1�/� andCARM1�/� MEFs. Cells were labeled with [35S]methioninefor 40 min and then chased with cold methionine for varioustime periods, followed by immunoprecipitation of endoge-nous p/CIP and analysis by immunoblotting and fluorogra-phy. The level of radiolabeled p/CIP relative to the totalcellular pool was substantially reduced at 8 h in CARM1�/�
cells, while no noticeable reductions were observed inCARM1�/� cells (Fig. 6C). These results suggest that theturnover of p/CIP in the CARM1 wild-type cells was signif-icantly faster, consistent with the notion that methylationaffects protein stability. To determine whether this effect isdirectly dependent on the amino acid residues recognized byCARM1, pulse-chase experiments were also conducted us-ing cells transfected with p/CIP mutants. Cells were trans-fected with either p/CIP or p/CIPR6A, and after 24 h cellswere replated and labeled with [35S]methionine. In agree-ment with the above data, the stability of the mutant wassignificantly increased compared to wild-type p/CIP (Fig.6D). To examine if p/CIP turnover is directly dependent onCARM1 methylation, MCF-7 cells were grown in the pres-ence of AdOx to hypomethylate p/CIP; subsequently, p/CIPwas immunoprecipitated and incubated with extracts de-rived from CARM1�/� or CARM1�/� MEFs, in the pres-ence of SAM, for various time periods, followed by immu-noblotting with p/CIP antibody. A dramatic reduction inp/CIP protein levels was observed following incubation withextracts derived from the CARM1 wild-type MEFs (Fig. 7).In contrast, p/CIP incubated with CARM1�/� extracts wasrelatively resistant to degradation. These results suggest that
CARM1-dependent methylation may be required for p/CIPturnover.
CARM1 dependent methylation regulates CBP association.To determine a functional role of p/CIP methylation, we ex-amined the effects of p/CIP on transcriptional activity by theestrogen receptor. Transient transfection assays in COS-1 cellswere performed using a reporter vector containing three cop-ies of an ERE upstream of a minimal promoter, as well as thePS2 proximal promoter-driven luciferase gene. Transfection oflimiting amounts of wild-type p/CIP resulted in a slight in-crease in transcriptional activity of the ERE luciferase in thepresence of �-estradiol (Fig. 8A). In contrast, an equivalentconcentration of p/CIPR6A mutant resulted in a significantlylarger response in all cell types examined. An enhanced tran-scriptional response by the p/CIPR6A mutant was more evi-dent on the PS2 promoter. Overexpression of wild-type p/CIPresulted in an approximately 2.8-fold increase in activity,whereas transfection of the p/CIPR6A mutant resulted in aconcentration-dependent effect with an additional threefoldincrease in luciferase activity relative to the wild type (Fig. 8A).These results suggest that a lack of p/CIP methylation maycause p/CIP to accumulate within the cell, which in turn resultsin enhanced transcription in response to �-estradiol.
We also examined the transcriptional activity of various re-gions of p/CIP fused to the Gal4 DBD. p/CIP (aa 819 to 1160)contains the major activation domain (AD1), which overlapsdirectly with the CBP/p300 interaction domain, but does notinclude the MD1 (5). When expressed in COS-1 cells, this Galfusion produced a robust transcriptional response (Fig. 8B).Surprisingly, extension of this construct to include the MD1(aa 819 to 1215) resulted in a dramatic reduction in transcrip-tional activity, indicating that this region may impart a negativeregulatory influence on p/CIP-mediated transcriptional activa-tion. Analysis of Gal4-p/CIP (aa 1158 to 1215) encompassingthe MD1 alone revealed no significant transcriptional activity.Experiments were also performed using the entire carboxyterminus of p/CIP fused to Gal4 (aa 819 to 1398) containingpoint mutations in the MD1, which should prevent (Fig. 8B,lower panel) the effects on transcriptional activity imparted bymethylation. Remarkably, these mutants displayed increasedtranscriptional activity when overexpressed in COS-1 cells,consistent with the effects obtained on the ERE-driven re-porter constructs. However, under these conditions we did notobserve significant differences in expression of the variousGal4 fusion proteins. These results suggest that the reduced
TABLE 1. Matrix-assisted laser desorption ionization–time of flight MS analysis of p/CIPa
Peptideno.
Observedmass (Da)
Predictedmass (Da) Residues (aa) Sequence
Modification(no. of asymmetricdi-Met arginines)b
1 1,619.83 1,618.82 192–204 KAPYILIVRMLMK 12 3,687.78 3,686.78 315–346 RHQEAYVHGHAETPVYRFSLADGTIVSAQTK 23 3,124.38 3,123.37 1176–1201 QLRMQLQQRLQGQQFLNQSRQALEMK 3c
4 3,140.43 3,139.50 1202–1229 MENPAGTAVMRPMMPQAFFNAQMAAQQK 1
a These peptides in p/CIP were identified by matrix-assisted laser desorption ionization–time of flight MS analysis as showing asymmetric dimethylation. For eachpeptide, the expected mass is the predicted mass containing the corresponding arginine modification indicated in the last column. The presence of each modificationindicated was confirmed based on the similarities between the observed and predicted masses.
b di-Met, dimethylated.c Methylation of residues 1176 to 1201 in p/CIP was highly dependent on CARM1 coexpression. Asymmetric dimethylation of R1178, R1184, and R1195 within residues
1176 to 1201 was then confirmed using tandem MS.
126 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

transcriptional activity associated with methylation is not sim-ply a result of increased p/CIP degradation but that the meth-ylation may affect activity by an additional mechanism.
The effects of p/CIP methylation on transcription and theproximity of the methylation sites to the CBP/p300 interactiondomain suggested that methylation may regulate the associa-tion with CBP. To investigate this possibility, we preparednuclear extracts from both wild-type and CARM1�/� MEFs
and purified endogenous p/CIP protein complexes using animmunoaffinity column. Initial experiments indicated thatp/CIP levels were elevated in the CARM1�/� MEFs by ap-proximately twofold (Fig. 9A). In order to compensate for theincrease in p/CIP levels, the amount of starting material in theinput was normalized so that each had approximately the sameamount of p/CIP, based on Western blotting. Immunoblottingof the purified proteins with a CBP-specific antibody indicated
FIG. 4. Molecular mapping of CARM1-dependent methylation of p/CIP. p/CIP truncation mutants were generated using a baculovirus systemand purified by immunoaffinity chromatography. Approximately 500 ng of the various truncation mutants was then tested as a substrate forCARM1 by in vitro methylation assay using 100 ng of purified CARM1. On the left is the Coomassie-stained gel of the purified proteins resolvedon an 8% SDS-PAGE gel, and on the right is the corresponding fluorograph. (B) One microgram of biotinylated peptides corresponding to theregions of p/CIP indicated was used as a substrate, and the incorporation of [3H]SAM was measured by in vitro methylation in the presence ofpurified CARM1. The asterisk indicates the three dimethylated arginines identified by mass spectrometry. (C) Homology of MD1 within the SRCfamily of proteins. SRC3 corresponds to the human homologue of p/CIP. The shaded areas indicate conserved arginines, and asterisks indicatearginines identified by mass spectrometry in p/CIP.
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 127
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

that the amount of CBP stably associated with p/CIP is signif-icantly enhanced in the CARM1 knockout MEFs (Fig. 9B).Indeed, in the wild-type MEFs, CBP was not detectable in thepurified p/CIP complexes. These results indicate that in theabsence of CARM1, the association between p/CIP and CBP isfacilitated.
DISCUSSION
Previous overexpression studies in intact cells and in vitroreconstitution assays using chromatinized templates have
shown that CARM1 can interact and synergize with both p/CIPand CBP/p300 to activate NR-mediated transcription (6, 26,62). This synergy is dependent on the methyltransferase activ-ity of CARM1 and has been attributed to the ability ofCARM1 to methylate specific arginines on histone H3 tails(50). Our data suggest that recruitment of CARM1 by p/CIPmay play a more complex role in transcription than previouslyanticipated. In the present study we demonstrate that the tran-scriptional coactivator p/CIP/SRC-3 is methylated by CARM1.Using mass spectrometric analysis, we have shown that thepredominant CARM1-dependent methylation sites in p/CIP
FIG. 5. Phosphorylation at S847 antagonizes methylation of p/CIP at R839. (A) GST recombinant proteins containing various regions of p/CIPspanning the full-length protein were generated in bacteria, purified, and used as substrates (500 ng) for purified CARM1 (100 ng) by in vitromethylation. At left is a Coomassie-stained gel of the GST fusion proteins used in the methylation assay resolved on a 12% SDS-PAGE gel, andat right is the corresponding fluorograph of the in vitro methylation reactions. (B) Comparison of the MD2 domains between p/CIP and SRC3.The asterisk corresponds to the phosphorylation site at aa 847. The shaded areas contain the methylation sites defined by in vitro methylationreactions using recombinant GST proteins. (C) Biotinylated peptides corresponding to the regions P1, P2, and P3 (shaded areas in panel B) wereused as substrates, and the incorporation of [3H]SAM was measured by in vitro methylation in the presence of purified CARM1. Peptidescorresponding to P1R839A and P1R844A contain substitutions of arginine to alanine. The peptide P1S847(p) is identical to P1 but contains aphosphorylated serine at aa 847.
128 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
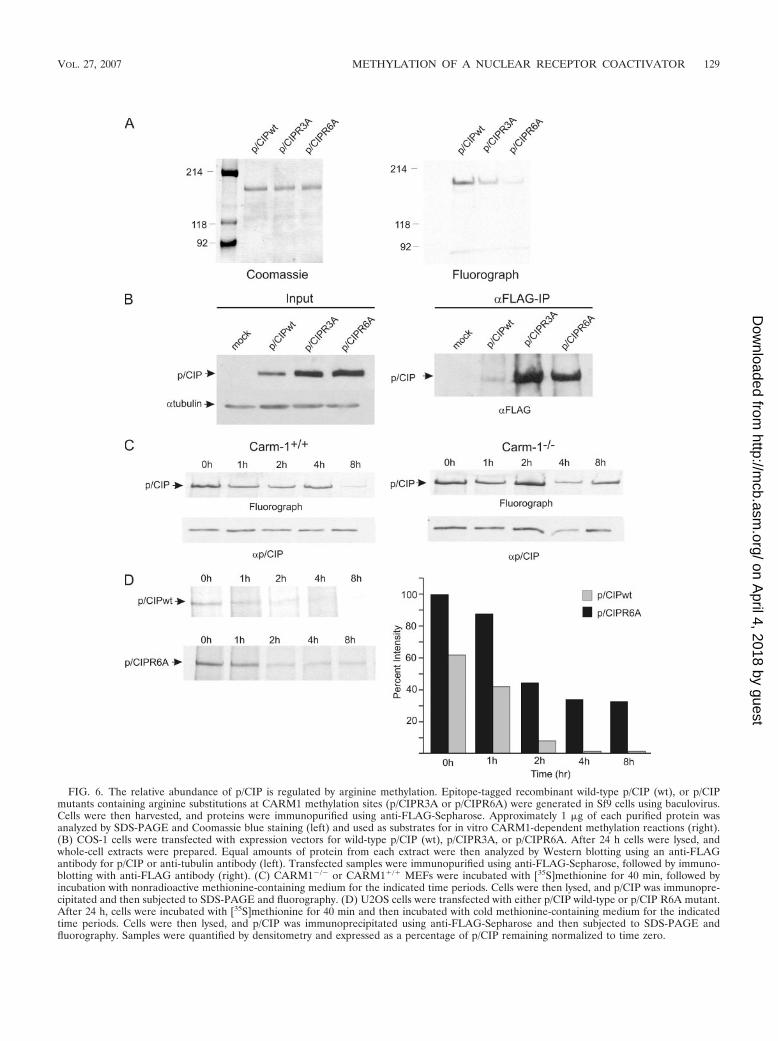
FIG. 6. The relative abundance of p/CIP is regulated by arginine methylation. Epitope-tagged recombinant wild-type p/CIP (wt), or p/CIPmutants containing arginine substitutions at CARM1 methylation sites (p/CIPR3A or p/CIPR6A) were generated in Sf9 cells using baculovirus.Cells were then harvested, and proteins were immunopurified using anti-FLAG-Sepharose. Approximately 1 �g of each purified protein wasanalyzed by SDS-PAGE and Coomassie blue staining (left) and used as substrates for in vitro CARM1-dependent methylation reactions (right).(B) COS-1 cells were transfected with expression vectors for wild-type p/CIP (wt), p/CIPR3A, or p/CIPR6A. After 24 h cells were lysed, andwhole-cell extracts were prepared. Equal amounts of protein from each extract were then analyzed by Western blotting using an anti-FLAGantibody for p/CIP or anti-tubulin antibody (left). Transfected samples were immunopurified using anti-FLAG-Sepharose, followed by immuno-blotting with anti-FLAG antibody (right). (C) CARM1�/� or CARM1�/� MEFs were incubated with [35S]methionine for 40 min, followed byincubation with nonradioactive methionine-containing medium for the indicated time periods. Cells were then lysed, and p/CIP was immunopre-cipitated and then subjected to SDS-PAGE and fluorography. (D) U2OS cells were transfected with either p/CIP wild-type or p/CIP R6A mutant.After 24 h, cells were incubated with [35S]methionine for 40 min and then incubated with cold methionine-containing medium for the indicatedtime periods. Cells were then lysed, and p/CIP was immunoprecipitated using anti-FLAG-Sepharose and then subjected to SDS-PAGE andfluorography. Samples were quantified by densitometry and expressed as a percentage of p/CIP remaining normalized to time zero.
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 129
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

are localized to a glutamine-rich region in the carboxy termi-nus, which we refer to as MD1.
PRMT1, a related methyltransferase, has also been shown tosynergize with SRC and CBP/p300 proteins to activate NRsignaling in transient transfection assays (26). The substratespecificity of CARM1 and PRMT1 is different, suggesting thatthe mechanism of coactivation of these two related methyl-transferases may also differ (50). Indeed, our data suggest thatPRMT1 is unable to methylate p/CIP in vitro, suggesting thatp/CIP is a specific substrate for CARM1. Additionally, p/CIPcould not be methylated in vitro by extracts derived fromCARM1�/� MEFs which still express PRMT1. This indicatesthat CARM1 is required for methylation of p/CIP in vivo.Importantly, immunoprecipitation of p/CIP from cell extracts,followed by Western blotting using the �R17H3 antibodywhich recognizes a dimethylated CARM1 recognition site onhistone H3, clearly recognized methylated p/CIP. Taken to-gether, these results, in conjunction with the mass spectrome-try data, demonstrate that p/CIP is a direct substrate forCARM1 in vivo.
By in vitro mapping experiments we have also identified asecond region in p/CIP that can be methylated (R839 and R961)which we refer to as MD2. Although methylation of MD2 isextremely robust using peptide substrates and recombinantGST proteins, the functional significance of this finding iscurrently unclear as we did not detect methylation at this siteusing MS. Furthermore, deletion of MD2 did not have a sig-nificant impact on overall methylation of p/CIP. Our inabilityto detect methylation at R839 through mass comparisons maybe due to the fact that R839 in Sf9 cells is already occupied bymethyl groups and would not be accessible to CARM1-depen-dent methylation in Sf9 cells or in vitro using baculovirus-generated protein. Additionally, we have shown that phosphor-ylation of S847 antagonizes methylation at R839. This finding issignificant as this site has recently been shown to be phosphor-
ylated on endogenous SRC-3 and is functionally required forcoactivation in response to hormonal signaling (60). Perhapsp/CIP produced in Sf9 cells may already be phosphorylated atS847, which may render R839 or R844 inaccessible to subsequentmethylation by CARM1.
A major finding of this study is that methylation regulatesthe stability of p/CIP. p/CIP mutants containing substitutionsof arginine to alanine accumulate to a much greater degreewhen overexpressed in various cell types compared to wild-type p/CIP. A significantly higher concentration of p/CIP wasalso detected in the CARM1�/� MEFs relative to the wild-type MEFs. This finding is also reflected in the pulse-chaseexperiments, which indicate that the methylation status ofp/CIP is an important determinant regulating its stability, bothin the CARM1�/� MEFs and in cells expressing p/CIP meth-ylation site mutants. Furthermore, we demonstrate that recon-stituting hypomethylated p/CIP with extracts derived fromwild-type MEFs resulted in a more rapid degradation of p/CIPcompared to reconstitution of p/CIP using CARM1�/� MEFextracts. These results suggest that the methylation status ofp/CIP may serve as a prerequisite for rapid degradation. Acritically unanswered question is the mechanism through whichmethylation affects p/CIP stability. Studies have shown thatarginine methylation promotes protein-protein interactions ina variety of signal transduction pathways (3). Thus, one possi-bility is that methylation of p/CIP by CARM1 allows specificproteins to interact selectively with p/CIP, which, in turn, fa-cilitates proteolytic degradation.
An additional finding in this study is that p/CIP methylationmay play a role in the transcriptional process by regulatingcomplex assembly. This assertion is based on the observationthat extension of the carboxy terminus AD1 to include theMD1 resulted in a significant decrease in transcriptional activ-ity when tethered to the Gal4 DBD. Conversely, mutation ofthe CARM1 recognition sites to nonmethylatable alanines en-
FIG. 7. Methylation of p/CIP is associated with increased degradation. HeLa cells were grown in the presence of AdOx for 8 days, followedby immunoprecipitation of p/CIP. Equal concentrations of p/CIP were then incubated with extracts from CARM1�/� or CARM1�/� MEFs in thepresence of SAM for the indicated time periods. Reactions were then terminated and analyzed by Western blotting using anti-p/CIP antibody. Thegraph on the right indicates the relative amounts of p/CIP remaining, based on densitometry.
130 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

FIG. 8. p/CIP methylation is associated with decreased transcriptional activity. (A) COS-1 cells were transfected with expression plasmidscontaining ER (10 ng), full-length wild-type p/CIP (10 ng), or p/CIPR6A (10 ng) cDNAs, along with reporter plasmids (200 ng) containing threecopies of an estrogen response element (ERE3Luc) or the PS2 proximal promoter (PS2Luc). Cells were stimulated with 10�6 M estradiol overnightand lysed on the following day, and luciferase activity was measured as described in Materials and Methods. (B) COS-1 cells were transfected withvarious regions of p/CIP fused to the Gal4 DBD along with a luciferase reporter plasmid containing five Gal4 binding sites. After 24 h, cells werelysed, and luciferase activity was measured. The corresponding Western blots shown on the right of each graph compare the relative levels ofexpression from the transfected Gal4 fusion cDNAs with the expression of antitubulin used as a loading control. NRID, nuclear receptorinteraction domain; wt, wild type.
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 131
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

hanced the transcriptional response of the carboxy terminuswhen tethered to Gal4. Under these experimental conditions,however, we did not observe a significant difference in theexpression levels of the various fusion proteins. This suggestedthat in addition to its role in protein stability, methylation maypromote a conformational change that weakens the associationof accessory proteins such as CBP/p300 and consequently con-tributes to complex dissociation. In support of this hypothesis,we observed that CBP forms a stable association with p/CIPand copurified with p/CIP from extracts derived fromCARM1�/� MEFs, compared to CARM�/� MEFs where noCBP was detected by Western blotting. Interestingly, a recentstudy has shown that CARM1-dependent methylation of theGRIP1-binding domain within CBP may also promote coacti-vator complex disassembly (31). Alternatively, methylationmay result in a conformational change in p/CIP which allowsanother protein to bind, thereby preventing interaction withCBP/p300.
The increase in p/CIP turnover and dissociation of CBP inresponse to methylation by another transcriptional coacti-vator seem paradoxical as these activities would typically beassociated with the negative control of transcription. How-
ever, they are consistent with the notion that transcriptionalactivation by nuclear hormone receptors is a dynamic pro-cess involving repetitive cycles of association and dissocia-tion of receptors and a large number of coregulators withtarget genes (38, 39). A number of mechanisms may con-tribute to this process, such as interaction with molecularchaperones which can associate with liganded nuclear re-ceptors and disassemble nuclear receptor/coregulator com-plexes (14). In addition, several studies have established alink between ligand-dependent transcriptional activation bynuclear hormone receptors and the ubiquitin-proteasomepathway (25). For example, it has been shown that thedegradation of ER�, in response to �-estradiol, is depen-dent on the presence of AIB1 (48). Coactivator turnover isalso dependent on interactions with the proteasome andmay be directly regulated by phosphorylation. The phosphor-ylation of GRIP1 by protein kinase A induces its degrada-tion via the ubiquitin-proteasome pathway, and a recentstudy has shown that SRC-3 is phosphorylated and thendegraded in response to retinoic acid (16, 21). Interestingly,Li et al. have identified a novel interaction between thecarboxy terminus of SRC-3 and Reg�, an activator of the20S proteasome. Loss of Reg� results in the accumulation ofSRC-3 protein levels (33). Studies examining promoter oc-cupancy have also established a link between proteasomalactivity and transcriptional activation (25). It has beenshown that recruitment of the proteasomal machinery isrequired for gene activation, presumably to allow cyclicclearance and continuous recycling of NR and coregulatorsat target genes (46, 48). The exact mechanism for coordi-nating the events involved in coactivator association anddissociation are poorly understood although increasing evi-dence suggests that posttranslational modifications may beinstrumental to the overall process. For example, treatmentof cells with hormones or various growth factors stimulatesthe phosphorylation of p/CIP/SRC-3 at specific sites andappears to be a prerequisite for hormone-dependent tran-scription activation that facilitates binding to CBP (13, 60).Acetylation of SRC-3 also appears to play a role in influ-encing transcription by facilitating coactivator release fromhormone-bound NR (9).
The human homologue of p/CIP was first isolated fromhuman breast tumors, and it is located within a region ofchromosome 20 that is often amplified in breast and ovariancancer (2). Several follow-up studies have confirmed thatp/CIP is amplified in a fraction of breast tumors, with ampli-fication frequencies ranging from 5 to 10% (18, 35, 40, 45).Moreover, compared to normal breast epithelium, p/CIP isoverexpressed at the RNA level in 31 to 64% of breast tumorsexamined. One question arising from the present study iswhether deregulated arginine methylation of p/CIP plays a rolein cellular transformation. Based on the data presented, wewould predict that the methylation status of p/CIP is an im-portant determinant of p/CIP transcriptional activity and mayaffect its oncogenic capacity. Alternatively, the accumulation ofhypomethylated p/CIP could also adversely affect the cyclingprocess inherent in NR-mediated transcription and impairtranscription at selective promoters.
In summary, the findings presented in this study linkCARM1-dependent methylation of p/CIP to both protein turn-
FIG. 9. The association between CBP and p/CIP is decreased inCARM1�/� MEFs. (A) Whole-cell extracts were prepared fromCARM1�/� MEFs or CARM1�/� MEFs (at a similar passage num-ber). Approximately 20 �g of protein from each extract was separatedby SDS-PAGE, and Western blotting was performed using anti-p/CIPantibody or antitubulin as a control. (B) Whole-cell extracts wereprepared from CARM1�/� MEFs or CARM1�/� MEFs, and theamount of starting material in the input was normalized so that eachhad approximately the same amount of p/CIP based on Western blot-ting. p/CIP was then immunopurified by passing the protein extractsthrough an anti-p/CIP immunoaffinity column. After extensive wash-ing, the bound proteins were eluted using 100 mM glycine (pH 3.0).Fractions were collected and assayed for p/CIP or CBP by SDS-PAGEfollowed by Western blotting.
132 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

over and coactivator complex dissociation, establishing argi-nine methylation as a critical regulatory signal in p/CIP-medi-ated transcription. In addition, this type of regulation couldserve to integrate diverse signaling pathways at the level ofcoactivator proteins and may have important implications forunderstanding the role of p/CIP in breast cancer. Further de-lineation of the role of p/CIP methylation and its downstreamphysiological effects is essential to decipher the complex reg-ulatory circuits in nuclear receptor signaling.
ACKNOWLEDGMENTS
We thank M. Stallcup for the CARM1 cDNA and H. Herschman forthe PRMTI cDNA.
J.T. is supported by National Cancer Institute of Canada grant015428. M.T.B. is supported by NIH grant DK62248 and in part by aninstitutional center grant (ES07784).
REFERENCES
1. Anafi, M., Y. F. Yang, N. A. Barlev, M. V. Govindan, S. L. Berger, T. R. Butt,and P. G. Walfish. 2000. GCN5 and ADA adaptor proteins regulate triiodo-thyronine/GRIP1 and SRC-1 coactivator-dependent gene activation by thehuman thyroid hormone receptor. Mol. Endocrinol. 14:718–732.
2. Anzick, S. L., J. Kononen, R. L. Walker, D. O. Azorsa, M. M. Tanner, X. Y.Guan, G. Sauter, O. P. Kallioniemi, J. M. Trent, and P. S. Meltzer. 1997.AIB1, a steroid receptor coactivator amplified in breast and ovarian cancer.Science 277:965–968.
3. Bedford, M. T., and S. Richard. 2005. Arginine methylation an emergingregulator of protein function. Mol. Cell 18:263–272.
4. Beischlag, T. V., S. Wang, D. W. Rose, J. Torchia, S. Reisz-Porszasz, K.Muhammad, W. E. Nelson, M. R. Probst, M. G. Rosenfeld, and O. Hankin-son. 2002. Recruitment of the NCoA/SRC-1/p160 family of transcriptionalcoactivators by the aryl hydrocarbon receptor/aryl hydrocarbon receptornuclear translocator complex. Mol. Cell. Biol. 22:4319–4333.
5. Brown, K., Y. Chen, T. M. Underhill, J. S. Mymyrk, and J. Torchia. 2003.The coactivator p/CIP/SRC-3 facilitates retinoic acid receptor signalling viarecruitment of GCN5. J. Biol. Chem. 278:39402–39412.
6. Chen, D., S. M. Huang, and M. R. Stallcup. 2000. Synergistic, p160 coacti-vator-dependent enhancement of estrogen receptor function by CARM1 andp300. J. Biol. Chem. 275:40810–40816.
7. Chen, D., H. Ma, H. Hong, S. S. Koh, S. M. Huang, B. T. Schurter, D. W.Aswad, and M. R. Stallcup. 1999. Regulation of transcription by a proteinmethyltransferase. Science 284:2174–2177.
8. Chen, H., R. J. Lin, R. L. Schiltz, D. Chakravarti, A. Nash, L. Nagy, M. L.Privalsky, Y. Nakatani, and R. M. Evans. 1997. Nuclear receptor coactivatorACTR is a novel histone acetyltransferase and forms a multimeric activationcomplex with P/CAF and CBP/p300. Cell 90:569–580.
9. Chen, H., R. J. Lin, W. Xie, D. Wilpitz, and R. M. Evans. 1999. Regulationof hormone-induced histone hyperacetylation and gene activation via acet-ylation of an acetylase. Cell 98:675–686.
10. Chevillard-Briet, M., D. Trouche, and L. Vandel. 2002. Control of CBPco-activating activity by arginine methylation. EMBO J. 21:5457–5466.
11. Daujat, S., U. M. Bauer, V. Shah, B. Turner, S. Berger, and T. Kouzarides.2002. Crosstalk between CARM1 methylation and CBP acetylation on his-tone H3. Curr. Biol. 12:2090–2097.
12. Espejo, A., J. Cote, A. Bednarek, S. Richard, and M. T. Bedford. 2002. Aprotein-domain microarray identifies novel protein-protein interactions. Bio-chem. J. 367:697–702.
13. Font de Mora, J., and M. Brown. 2000. AIB1 is a conduit for kinase-mediated growth factor signaling to the estrogen receptor. Mol. Cell. Biol.20:5041–5047.
14. Freeman, B. C., and K. R. Yamamoto. 2002. Disassembly of transcriptionalregulatory complexes by molecular chaperones. Science 296:2232–2235.
15. Fujiwara, T., Y. Mori, D. L. Chu, Y. Koyama, S. Miyata, H. Tanaka, K.Yachi, T. Kubo, H. Yoshikawa, and M. Tohyama. 2006. CARM1 regulatesproliferation of PC12 cells by methylating HuD. Mol. Cell. Biol. 26:2273–2285.
16. Gianni, M., E. Parrella, I. Raska, Jr., E. Gaillard, E. A. Nigro, C. Gaudon,E. Garattini, and C. Rochette-Egly. 2006. P38MAPK-dependent phosphor-ylation and degradation of SRC-3/AIB1 and RAR�-mediated transcription.EMBO J. 25:739–751.
17. Gill, R. K., L. M. Atkins, B. W. Hollis, and N. H. Bell. 1998. Mapping thedomains of the interaction of the vitamin D receptor and steroid receptorcoactivator-1. Mol. Endocrinol. 12:57–65.
18. Glaeser, M., T. Floetotto, B. Hanstein, M. W. Beckmann, and D. Nieder-acher. 2001. Gene amplification and expression of the steroid receptor co-activator SRC3 (AIB1) in sporadic breast and endometrial carcinomas.Horm. Metab. Res. 33:121–126.
19. Harlow, E., and D. Lane. 1999. Using antibodies: a laboratory manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
20. Heery, D. M., E. Kalkhoven, S. Hoare, and M. G. Parker. 1997. A signaturemotif in transcriptional co-activators mediates binding to nuclear receptors.Nature 387:733–736.
21. Hoang, T., I. S. Fenne, C. Cook, B. Borud, M. Bakke, E. A. Lien, and G.Mellgren. 2004. cAMP-dependent protein kinase regulates ubiquitin-protea-some-mediated degradation and subcellular localization of the nuclear re-ceptor coactivator GRIP1. J. Biol. Chem. 279:49120–49130.
22. Hong, H., K. Kohli, A. Trivedi, D. L. Johnson, and M. R. Stallcup. 1996.GRIP1, a novel mouse protein that serves as a transcriptional coactivator inyeast for the hormone binding domains of steroid receptors. Proc. Natl.Acad. Sci. USA 93:4948–4952.
23. Kamei, Y., L. Xu, T. Heinzel, J. Torchia, R. Kurokawa, B. Gloss, S. C. Lin,R. A. Heyman, D. W. Rose, C. K. Glass, and M. G. Rosenfeld. 1996. A CBPintegrator complex mediates transcriptional activation and AP-1 inhibitionby nuclear receptors. Cell 85:403–414.
24. Kim, J. H., H. Li, and M. R. Stallcup. 2003. CoCoA, a nuclear receptorcoactivator which acts through an N-terminal activation domain of p160coactivators. Mol. Cell 12:1537–1549.
25. Kinyamu, H. K., J. Chen, and T. K. Archer. 2005. Linking the ubiquitin-proteasome pathway to chromatin remodeling/modification by nuclear re-ceptors. J. Mol. Endocrinol. 34:281–297.
26. Koh, S. S., D. Chen, Y. H. Lee, and M. R. Stallcup. 2001. Synergisticenhancement of nuclear receptor function by p160 coactivators and twocoactivators with protein methyltransferase activities. J. Biol. Chem. 276:1089–1098.
27. Korzus, E., J. Torchia, D. W. Rose, L. Xu, R. Kurokawa, E. M. McInerney,T. M. Mullen, C. K. Glass, and M. G. Rosenfeld. 1998. Transcription factor-specific requirements for coactivators and their acetyltransferase functions.Science 279:703–707.
28. Lee, D. Y., C. Teyssier, B. D. Strahl, and M. R. Stallcup. 2005. Role ofprotein methylation in regulation of transcription. Endocr. Rev. 26:147–170.
29. Lee, J., and M. T. Bedford. 2002. PABP1 identified as an arginine methyl-transferase substrate using high-density protein arrays. EMBO Rep. 3:268–273.
30. Lee, S. K., H. J. Kim, J. W. Kim, and J. W. Lee. 1999. Steroid receptorcoactivator-1 and its family members differentially regulate transactivationby the tumor suppressor protein p53. Mol. Endocrinol. 13:1924–1933.
31. Lee, Y. H., S. A. Coonrod, W. L. Kraus, M. A. Jelinek, and M. R. Stallcup.2005. Regulation of coactivator complex assembly and function by proteinarginine methylation and demethylimination. Proc. Natl. Acad. Sci. USA102:3611–3616.
32. Li, H., P. J. Gomes, and J. D. Chen. 1997. RAC3, a steroid/nuclear receptor-associated coactivator that is related to SRC-1 and TIF2. Proc. Natl. Acad.Sci. USA 94:8479–8484.
33. Li, X., D. M. Lonard, S. Y. Jung, A. Malovannaya, Q. Feng, J. Qin, S. Y. Tsai,M. J. Tsai, and B. W. O’Malley. 2006. The SRC-3/AIB1 coactivator isdegraded in a ubiquitin- and ATP-independent manner by the REG� pro-teasome. Cell 124:381–392.
34. Liao, L., S. Q. Kuang, Y. Yuan, S. M. Gonzalez, B. W. O’Malley, and J. Xu.2002. Molecular structure and biological function of the cancer-amplifiednuclear receptor coactivator SRC-3/AIB1. J. Steroid Biochem. Mol. Biol.83:3–14.
35. List, A., K. Lauritson, R. Rieter, C. Powers, A. Wellstien, and A. Riegel. 2001.Ribozyme targeting demonstrates that the nuclear receptor coactivator AIB1is a rate-limiting factor for estrogen-dependent growth of human MCF-7breast cancer cells. J. Biol. Chem. 276:23763–23768.
36. Liu, Q., and G. Dreyfuss. 1995. In vivo and in vitro arginine methylation ofRNA-binding proteins. Mol. Cell. Biol. 15:2800–2808.
37. McInerney, E. M., D. W. Rose, S. E. Flynn, S. Westin, T. M. Mullen, A.Krones, J. Inostroza, J. Torchia, R. T. Nolte, N. Assa-Munt, M. V. Milburn,C. K. Glass, and M. G. Rosenfeld. 1998. Determinants of coactivatorLXXLL motif specificity in nuclear receptor transcriptional activation.Genes Dev. 12:3357–3368.
38. Metivier, R., G. Penot, M. R. Hubner, G. Reid, H. Brand, M. Kos, and F.Gannon. 2003. Estrogen receptor-alpha directs ordered, cyclical, and com-binatorial recruitment of cofactors on a natural target promoter. Cell 115:751–763.
39. Metivier, R., G. Reid, and F. Gannon. 2006. Transcription in four dimen-sions: nuclear receptor-directed initiation of gene expression. EMBO Rep.7:161–167.
40. Murphy, L. C., S. L. Simon, A. Parkes, E. Leygue, H. Dotzlaw, L. Snell, S.Troup, A. Adeyinka, and P. H. Watson. 2000. Altered expression of estrogenreceptor coregulators during human breast tumorigenesis. Cancer Res. 60:6266–6271.
41. Onate, S. A., V. Boonyaratanakornkit, T. E. Spencer, S. Y. Tsai, M. J. Tsai,D. P. Edwards, and B. W. O’Malley. 1998. The steroid receptor coactivator-1contains multiple receptor interacting and activation domains that cooper-atively enhance the activation function 1 (AF1) and AF2 domains of steroidreceptors. J. Biol. Chem. 273:12101–12108.
VOL. 27, 2007 METHYLATION OF A NUCLEAR RECEPTOR COACTIVATOR 133
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

42. Perissi, V., and M. G. Rosenfeld. 2005. Controlling nuclear receptors: thecircular logic of cofactor cycles. Nat. Rev. Mol. Cell. Biol. 6:542–554.
43. Qi, C., J. Chang, Y. Zhu, A. V. Yeldandi, S. M. Rao, and Y. J. Zhu. 2002.Identification of protein arginine methyltransferase 2 as a coactivator forestrogen receptor alpha. J. Biol. Chem. 277:28624–28630.
44. Qutob, M. S., R. N. Bhattacharjee, E. Pollari, S. P. Yee, and J. Torchia. 2002.Microtubule-dependent subcellular redistribution of the transcriptional co-activator p/CIP. Mol. Cell. Biol. 22:6611–6626.
45. Rebbeck, T. R., Y. Wang, P. W. Kantoff, K. Krithivas, S. L. Neuhausen, A. K.Godwin, M. B. Daly, S. A. Narod, J. S. Brunet, D. Vesprini, J. E. Garber,H. T. Lynch, B. L. Weber, and M. Brown. 2001. Modification of BRCA1- andBRCA2-associated breast cancer risk by AIB1 genotype and reproductivehistory. Cancer Res. 61:5420–5424.
46. Reid, G., M. R. Hubner, R. Metivier, H. Brand, S. Denger, D. Manu, J.Beaudouin, J. Ellenberg, and F. Gannon. 2003. Cyclic, proteasome-mediatedturnover of unliganded and liganded ER� on responsive promoters is anintegral feature of estrogen signaling. Mol. Cell 11:695–707.
47. Shang, Y., X. Hu, J. DiRenzo, M. A. Lazar, and M. Brown. 2000. Cofactordynamics and sufficiency in estrogen receptor-regulated transcription. Cell103:843–852.
48. Shao, W., E. K. Keeton, D. P. McDonnell, and M. Brown. 2004. CoactivatorAIB1 links estrogen receptor transcriptional activity and stability. Proc. Natl.Acad. Sci. USA 101:11599–11604.
49. Shi, Y., F. Lan, C. Matson, P. Mulligan, J. R. Whetstine, P. A. Cole, and R. A.Casero. 2004. Histone demethylation mediated by the nuclear amine oxidasehomolog LSD1. Cell 119:941–953.
50. Strahl, B. D., S. D. Briggs, C. J. Brame, J. A. Caldwell, S. S. Koh, H. Ma,R. G. Cook, J. Shabanowitz, D. F. Hunt, M. R. Stallcup, and C. D. Allis.2001. Methylation of histone H4 at arginine 3 occurs in vivo and is mediatedby the nuclear receptor coactivator PRMT1. Curr. Biol. 11:996–1000.
51. Suen, C. S., T. J. Berrodin, R. Mastroeni, B. J. Cheskis, C. R. Lyttle, andD. E. Frail. 1998. A transcriptional coactivator, steroid receptor coactiva-tor-3, selectively augments steroid receptor transcriptional activity. J. Biol.Chem. 273:27645–27653.
52. Tilli, M. T., R. Reiter, A. S. Oh, R. T. Henke, K. McDonnell, G. I. Gallicano,P. A. Furth, and A. T. Riegel. 2005. Overexpression of an N-terminallytruncated isoform of the nuclear receptor coactivator amplified in breastcancer 1 leads to altered proliferation of mammary epithelial cells in trans-genic mice. Mol. Endocrinol. 19:644–656.
53. Torchia, J., D. W. Rose, J. Inostroza, Y. Kamei, S. Westin, C. K. Glass, andM. G. Rosenfeld. 1997. The transcriptional co-activator p/CIP binds CBP andmediates nuclear-receptor function. Nature 387:677–684.
54. Torres-Arzayus, M. I., J. Font de Mora, J. Yuan, F. Vazquez, R. Bronson, M.Rue, W. R. Sellers, and M. Brown. 2004. High tumor incidence and activa-tion of the PI3K/AKT pathway in transgenic mice define AIB1 as an onco-gene. Cancer Cell 6:263–274.
55. Voegel, J. J., M. J. Heine, M. Tini, V. Vivat, P. Chambon, and H. Gronem-eyer. 1998. The coactivator TIF2 contains three nuclear receptor-bindingmotifs and mediates transactivation through CBP binding-dependent and-independent pathways. EMBO J. 17:507–519.
56. Voegel, J. J., M. J. Heine, C. Zechel, P. Chambon, and H. Gronemeyer. 1996.TIF2, a 160-kDa transcriptional mediator for the ligand-dependent activa-tion function AF-2 of nuclear receptors. EMBO J. 15:3667–3675.
57. Wang, H., Z. Q. Huang, L. Xia, Q. Feng, H. Erdjument-Bromage, B. D.Strahl, S. D. Briggs, C. D. Allis, J. Wong, P. Tempst, and Y. Zhang. 2001.Methylation of histone H4 at arginine 3 facilitating transcriptional activationby nuclear hormone receptor. Science 293:853–857.
58. Wang, Y., J. Wysocka, J. Sayegh, Y. H. Lee, J. R. Perlin, L. Leonelli, L. S.Sonbuchner, C. H. McDonald, R. G. Cook, Y. Dou, R. G. Roeder, S. Clarke,M. R. Stallcup, C. D. Allis, and S. A. Coonrod. 2004. Human PAD4 regulateshistone arginine methylation levels via demethylimination. Science 306:279–283.
59. Wu, R. C., J. Qin, Y. Hashimoto, J. Wong, J. Xu, S. Y. Tsai, M. J. Tsai, andB. W. O’Malley. 2002. Regulation of SRC-3 (pCIP/ACTR/AIB-1/RAC-3/TRAM-1) coactivator activity by IB kinase. Mol. Cell. Biol. 22:3549–3561.
60. Wu, R. C., J. Qin, P. Yi, J. Wong, S. Y. Tsai, M. J. Tsai, and B. W. O’Malley.2004. Selective phosphorylations of the SRC-3/AIB1 coactivator integrategenomic responses to multiple cellular signaling pathways. Mol. Cell 15:937–949.
61. Xu, J., L. Liao, G. Ning, H. Yoshida-Komiya, C. Deng, and B. W. O’Malley.2000. The steroid receptor coactivator SRC-3 (p/CIP/RAC3/AIB1/ACTR/TRAM-1) is required for normal growth, puberty, female reproductive func-tion, and mammary gland development. Proc. Natl. Acad. Sci. USA 97:6379–6384.
62. Xu, W., H. Chen, K. Du, H. Asahara, M. Tini, B. M. Emerson, M. Montminy,and R. M. Evans. 2001. A transcriptional switch mediated by cofactor meth-ylation. Science 294:2507–2511.
63. Xu, W., H. Cho, S. Kadam, E. M. Banayo, S. Anderson, J. R. Yates, 3rd,B. M. Emerson, and R. M. Evans. 2004. A methylation-mediator complex inhormone signaling. Genes Dev. 18:144–156.
64. Yadav, N., J. Lee, J. Kim, J. Shen, M. C. Hu, C. M. Aldaz, and M. T. Bedford.2003. Specific protein methylation defects and gene expression perturbationsin coactivator-associated arginine methyltransferase 1-deficient mice. Proc.Natl. Acad. Sci. USA 100:6464–6468.
65. Zhou, G., Y. Hashimoto, I. Kwak, S. Y. Tsai, and M. J. Tsai. 2003. Role ofthe steroid receptor coactivator SRC-3 in cell growth. Mol. Cell. Biol. 23:7742–7755.
66. Zhou, H. J., J. Yan, W. Luo, G. Ayala, S. H. Lin, H. Erdem, M. Ittmann, S. Y.Tsai, and M. J. Tsai. 2005. SRC-3 is required for prostate cancer cellproliferation and survival. Cancer Res. 65:7976–7983.
134 NAEEM ET AL. MOL. CELL. BIOL.
on April 4, 2018 by guest
http://mcb.asm
.org/D
ownloaded from





























