Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/00/$04.0010
Dec. 2000, p. 9317–9330 Vol. 20, No. 24
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Functional Interaction between the Coactivator DrosophilaCREB-Binding Protein and ASH1, a Member of the
Trithorax Group of Chromatin ModifiersFREDERIC BANTIGNIES,† RICHARD H. GOODMAN, AND SARAH M. SMOLIK*
Vollum Institute and Department of Cell and Developmental Biology,Oregon Health Sciences University, Portland, Oregon 97201
Received 15 June 2000/Returned for modification 24 July 2000/Accepted 19 September 2000
CREB-binding protein (CBP) is a coactivator for multiple transcription factors that transduce a variety ofsignaling pathways. Current models propose that CBP enhances gene expression by bridging the signal-responsive transcription factors with components of the basal transcriptional machinery and by augmentingthe access of transcription factors to DNA through the acetylation of histones. To define the pathways andproteins that require CBP function in a living organism, we have begun a genetic analysis of CBP in flies. Wehave overproduced Drosophila melanogaster CBP (dCBP) in a variety of cell types and obtained distinct adultphenotypes. We used an uninflated-wing phenotype, caused by the overexpression of dCBP in specific centralnervous system cells, to screen for suppressors of dCBP overactivity. Two genes with mutant versions that actas dominant suppressors of the wing phenotype were identified: the PKA-C1/DCO gene, encoding the catalyticsubunit of cyclic AMP protein kinase, and ash1, a member of the trithorax group (trxG) of chromatin modifiers.Using immunocolocalization, we showed that the ASH1 protein is specifically expressed in the majority of thedCBP-overexpressing cells, suggesting that these proteins have the potential to interact biochemically. Thismodel was confirmed by the findings that the proteins interact strongly in vitro and colocalize at specific siteson polytene chromosomes. The trxG proteins are thought to maintain gene expression during development bycreating domains of open chromatin structure. Our results thus implicate a second class of chromatin-associated proteins in mediating dCBP function and imply that dCBP might be involved in the regulation ofhigher-order chromatin structure.
For proper cellular function and the elaboration of devel-opmental programs, gene expression must be regulated tightly.There is increasing evidence that large transcription com-plexes, composed of unique combinations of sequence-specificactivators and repressors, coactivators, and corepressors, playan important role in determining the temporal and spatialpatterns of gene expression (for review, see reference 39).
The CREB binding protein (CBP) is one of most extensivelycharacterized coactivator proteins. CBP was first identifiedthrough its ability to link the cyclic AMP protein kinase(PKA)-phosphorylated form of CREB to components of thebasal transcriptional machinery, including TFIIB (14, 34),TATA-binding protein (65), and the RNA polymerase II ho-loenzyme complex (28, 44). CBP is highly related to the ade-novirus E1A binding protein p300 (17), and CBP and p300 areconsidered to be functional homologues (4, 38), although a fewdifferences in their activities have been reported (27). CBP andp300 associate with a wide variety of transcriptional activatorsin addition to CREB, suggesting that each may serve as atranscriptional integrator of different signaling cascades (re-viewed in references 20 and 60). Thus, one model for thefunction of CBP and p300 is bridging DNA binding transcrip-tion factors to components of the basal transcriptional machin-ery.
Another function of coactivators appears to be the modifi-
cation of chromatin structure. In this regard, CBP and p300have also been proposed to mediate transcriptional activationvia intrinsic (6, 46) and associated (9, 63, 81) histone acetyl-transferase (HAT) activity. Targeted HAT activity is thoughtto facilitate the access of nuclear factors to their target sites byrelaxing the interaction between histones and the DNA (for areview, see reference 77). Moreover, recent studies suggestthat transcriptional activation mediated through CBP or p300occurs only in the context of chromatin (31, 32). Therefore,CBP and p300 may regulate gene expression by interactingwith components of the transcriptional machinery as well as byaugmenting the access of factors to DNA through their HATactivities. Acetylation of basal and sequence-specific transcrip-tional regulators may also contribute to CBP function (22, 24).
Genetic studies indicate an essential role for CBP in cellularfunction and development (reviewed in reference 21). In hu-mans, CBP loss of function is associated with Rubinstein-Taybisyndrome, a haploinsufficiency disorder characterized by men-tal retardation, developmental defects, and an increased pre-disposition to cancer (42, 48). Chromosomal translocationsthat fuse CBP with MOZ (monocytic zinc finger protein) orMLL (mixed-lineage leukemia protein, a trithorax group-likeprotein) are associated with various types of myeloid leukemia(7, 62). In addition, somatic mutations of the p300 gene havebeen detected in colorectal and gastric carcinomas (43). Geneknockouts in mice indicated that CBP and p300 are requiredfor normal embryonic development and viability (69, 82). Fi-nally, mutations in the Caenorhabditis elegans homologue ofCBP (CBP-1) affect the differentiation of several embryonictissues (59).
In Drosophila melanogaster, Drosophila CBP (dCBP) loss-of-function mutations cause embryonic lethality. Specifically,
* Corresponding author. Mailing address: Department of Cell andDevelopmental Biology L-215, Oregon Health Sciences University,3181 SW Sam Jackson Park Rd., Portland, OR 97201. Phone: (503)494-7192. Fax: (503) 494-4353. E-mail: [email protected].
† Present address: Institut de Genetique Humaine, CNRS UPR1142,Montpellier, France.
9317
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

dCBP serves as a coactivator for transcription factor Cubitusinterruptus (CI) and mediates its activity in the hedgehog path-way (2, 12). dCBP is also a coactivator of the dorsal protein(D1) and Mad, mediating dl-dependent twist expression anddpp-induced transcriptional stimulation, respectively (3, 79).However, dCBP does not always function as a coactivator.Recent studies have shown that dCBP binds to the Drosophilahomologue of the T-cell factor (dTCF) and facilitates dTCF-mediated repression in the Wnt/Wingless signaling (78).Therefore, dCBP can function as both a coactivator and acorepressor during embryogenesis.
To further define the developmental processes and the sig-naling pathways that require dCBP, we have taken advantageof the yeast GAL4 enhancer trap (8) system to generate trans-genic flies that overexpress dCBP in a variety of cell types. Thedominant overexpression adult phenotypes generated with thissystem were used to screen for suppressors of dCBP overac-tivity in specific tissues. In this report, we describe a functionaland specific interaction between transcriptional coactivatordCBP and ASH1, a member of the trithorax group (trxG) ofchromatin modifiers.
The trxG proteins are required to maintain the continuedand efficient expression of homeotic and other genes through-out development. Loss-of-function mutations in the trithoraxgroup genes cause homeotic transformations because they failto maintain the expression pattern of homeotic selector genes.While the trxG proteins function as transcriptional activators,members of Polycomb group (PcG) genes form stable com-plexes that maintain a repressed pattern of homeotic genefunction (for reviews, see references 49, 50, and 61). Currentmodels envision that trxG and PcG proteins lock in the activeor inactive state, respectively, by creating a stable chromatinorganization. trxG represents a heterogeneous family of pro-teins with diverse functions. Some of these proteins, such asTrithorax (TRX), ASH1, ASH2, GAGA, and ZESTE, areassociated with particular sites on polytene chromosomes (1,13, 33, 51, 71, 74), while others, such as Brahma (BRM) andSNR1, are found in chromatin-remodeling complexes that maynot be associated with specific chromosomal regions (16, 68).There is some evidence that one of the functions of the trxGproteins may be to recruit chromatin-remodeling complexes toDNA. GAGA is required for the function of one chromatin-remodeling complex, the Drosophila NURF complex (74), andTRX was shown to physically interact with SNR1, a componentof the Drosophila SWI/SNF complex (54). These studiesstrongly support the model that trxG proteins are importantregulators of higher-order chromatin structure. However, theprecise role of each of the diverse trxG members and thefunctional relationships that might exist among them and withother transcriptional regulatory factors are still poorly under-stood.
Our studies show that mutations in the ash1 gene suppress awing phenotype caused by the overexpression of dCBP in spe-cific central nervous system (CNS) cells. This suppression isspecific for ash1 because other members of the trithorax familydo not have the same effect. At the cellular level, ASH1 ex-pression coincides with the overexpression pattern of dCBPand, in wild-type flies, ASH1 and dCBP colocalize in the nu-cleus of the ASH1-expressing neurons. Finally, at the molec-ular level, we show that dCBP interacts strongly with ASH1and that the two proteins colocalize to specific sites on poly-tene chromosomes. Our results strongly suggest that coactiva-tor dCBP and trithorax factor ASH1 are part of a functionalcomplex in vivo. These findings implicate a new type of chro-matin-associated proteins in mediating dCBP function and im-
ply that, in addition to its HAT activity, dCBP may participatein the regulation of higher-order chromatin structure.
MATERIALS AND METHODS
Drosophila strains. All the ash1 alleles and the ash2 allele were kindly providedby Allen Shearn (Johns Hopkins University). They are described by Tripoulas etal. (70, 71) and Adamson and Shearn (1). The PKA-C1/DCO null allele, G9, (36)and the UAS-R* line containing the upstream activation sequence (UAS) dom-inant-negative catalytic subunit of PKA (37) were kindly provided by Dan Kal-deron (Columbia University). The GMR-GAL4 line was provided G. Rubin,(University of California, Berkeley). Other mutant alleles and most of the dele-tions used in this study came from the Bloomington Stock Center. UAS-dCBPtransformants (Tr21 and Tr36) were established by standard methods. The NotIdCBP fragment that includes the entire dCBP cDNA was cloned into the NotIsite of pUAST (8). yw embryos were injected with this DNA and the ppD2-3helper as described previously, and the transformants were mapped and put intostock (64). The UAS-dCBP Tr21 insert is on the fourth chromosome, and theTr36 insert is on the X. The UAS-386 and UAS-363 lines were kindly providedby C. O’Kane (Cambridge University). The balancer chromosomes MKRS,TM3,Stubble (Sb), and TM6B,Tubby (Tb), which are used in these studies, havebeen described by Lindsley and Zimm (37a). The balancer chromosome strainpk-sple33 pr cn/T(2;3)SM6.TM6B,Tb was kindly provided by J. Roote and M.Ashburner (Cambridge University). The wild-type strain used in these studies isCanton-S. All crosses were reared at 25°C on standard cornmeal yeast extractsource media.
Detection of b-galactosidase (b-GAL) activity in imaginal discs and CNS.Third-instar larvae were dissected into saline, fixed for 1 h in phosphate-bufferedsaline (PBS) and 4% formaldehyde, and rinsed several times with PBST (PBSwith 0.3% Triton X-100). Fixed samples were then placed in a 0.02% X-Gal(5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside) solution (53) at 30°C untilthe appearance of the blue color (about 1 h for the GAL4-386 CNS, no stainingafter 24 h for the GAL4-386 imaginal discs). After staining, samples were washedseveral times in PBST. Larval tissues were mounted in PBS–50% glycerol andphotographed with Echtachrome 64 film using Nomarski optics.
Screen to isolate dominant suppressors of the dCBP overexpression wingphenotype. The screen was performed at 25°C where the uninflated-wing phe-notype has a 100% penetrance. GAL4-386/MKRS; Tr21/1 females were collectedand crossed to males from the Bloomington Stock Center deficiency kit [Df(2)/Balancer or Df(3)/Balancer] (see Fig. 2). Approximately 150 different deletions,which cover more than 80% of the second and third chromosomes, were tested.From these crosses, one-fourth of the progeny were expected to be GAL4-386/1;Tr21/1 and have uninflated wings. In fact, this ratio is less and approximatesone-eighth of the total population because this genotype is weak. Although Tr21is inserted on the fourth chromosome, it is linked to a miniwhite transgene andproduces a characteristic pale-orange eye color that can be monitored in a w1
background. All of the second and third chromosome balancers used in this studywere tested and had no effect on the uninflated-wing phenotype (data notshown). When the balancer chromosome of a deficiency stock had a dominantwing marker, for example, Curly or Serrate, the balancer was exchanged with onecarrying a more convenient marker, i.e., SM6.TM6B,Tb or TM3,Sb. In this screen,a deletion was considered to have no effect on the uninflated-wing phenotypewhen the population of GAL4-386 heterozygotes had approximately the samenumber of flies with uninflated wings regardless of whether they carried thedeletion chromosome or the balancer. When the number of uninflated-wingGAL4-386/1 flies with the balancer chromosome was greater than the number ofuninflated-wing GAL4-386/1 flies with the deletion chromosome by at least afactor of two, then putative suppressed males (partially inflated or wild-typewings) were crossed to w9 females to determine the presence of the Tr21 trans-gene in the next generation. Approximately 20 to 30 putative suppressed maleswere tested for the presence of Tr21. When Df/1; GAL4-386/1; Tr21/1 maleswere obtained, the deletion was considered a potential suppressor. These dele-tions were rescreened in the same way to confirm the suppression. Only thedeletions for which the number of Df/1; GAL4-386/1; Tr21/1 flies with partiallyinflated or wild-type wings (suppressed phenotype) was at least 50% greater thanthe number of flies with the same genotype and uninflated wings were consideredsuppressors (deletions in Table 2). The same procedure was used to determinesuppression by mutations in single genes (alleles in Table 3). Each experimentwas repeated at least twice.
b-Gal expression tests. This assay allows us to measure the possible effect ofa deletion on the expression of UAS-transgene and distinguish between suppres-sors that repress the expression of the transgene and those that represent dCBP-interacting genes (see Table 2). Deficiency stocks were put over a balancerchromosome with a dominant pupal marker: SM6.TM6B,Tb or TM6b,Tb. Df(2)/SM6.TM6B,Tb or Df(3)/TM6b,Tb males were crossed to UAS-LacZ; GAL4-386homozygous females. Three-day-old pupae were collected and assayed for b-Galactivity using o-nitrophenyl-b-D-galactopyranoside (ONPG) as the substrate.Briefly, a single pupa was squashed and permeabilized in 800 ml of Z buffer(Na2HPO4 z 7H2O [16.1 g/liter], NaH2PO4 z H2O [5.5 g/liter], KCl [0.75 g/liter],MgSO4 z 7H2O [0.246 g/liter], pH 7.0) containing b-mercaptoethanol (2.7 ml/ml)plus 50 ml of 0.1% sodium dodecyl sulfate (SDS) and 50 ml of chloroform. ONPGwas used as the substrate (160 ml of a solution of 4 mg/ml in 0.1 M phosphate
9318 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

buffer, pH 7.0), and assays were performed at 30°C. After 30 min, the reactionwas stopped with 0.4 ml of 1 M Na2CO3 and the optical density at 420 nm wasdetermined. For each test, at least five pupae were analyzed separately. b-Galactivity in Tb flies was compared to b-Gal activity in the non-Tb flies thatcontained the deletion. If the activities were similar, then the deletion has noeffect and we considered the test positive. If the activity was largely reduced inthe non-Tb flies, then the deletion represses the transgene either by affectingGAL4, the UAS, or the stability of the gene product and we considered the testnegative. Using the same strategy (Tb versus non-Tb larvae), we were also ableto compare qualitatively the expression of b-Gal in fixed larval brains stainedwith X-Gal (53). In all cases, the qualitative results in whole tissues were con-sistent with the quantitative values obtained for the pupal squashes (data notshown).
Confocal analysis of larval and pupal CNS. Third-instar larvae or pharateadults were dissected into saline, and the CNSs were processed. They were fixedfor 1 h in PBS–4% paraformaldehyde, rinsed several times with PBST, andblocked at least for 1 h in PBST–10% HS. The CNSs were incubated overnightat 4°C with the primary antibody, washed several times in PBST–10% HS (atleast three times for 20 min each over 1 h), and then incubated for 1 h at roomtemperature with the secondary antibody, washed several times in PBST, andmounted in a “slow-fade” buffer specific for immunofluorescence (MolecularProbes). ASH1 was detected by a rabbit polyclonal antibody (71) (affinity puri-fied; kindly provided by Allen Shearn, Johns Hopkins University) at a dilution of1:40, and dCBP was detected by a chicken polyclonal antibody raised against theCREB binding domain (CBD) of dCBP at a dilution of 1:800. FMRFamide(PT-2) and peptidylglycine-a-hydroxylating mono-oxygenase (PHM) rabbit poly-clonal antibodies (30, 67) (kindly provided by Paul Taghert) were used at dilu-tions of 1:2,000 and 1:500, respectively. Fluorescein anti-rabbit (Vector) andrhodamine anti-chicken (Jackson) secondary antibodies were used at a dilutionof 1/200. CNSs were examined with a confocal laser scan microscope (Bio-Rad1024ES laser and Nikon Eclipse TE300 microscope).
The specificity of the dCBP antibody was determined in three ways. First, thestaining of wild-type embryos was competed with increasing dosages of dCBDantigen. Second, the antibody failed to stain embryos that have mutant dCBP andthat do not express dCBP RNAs. Third, the antibody detected a glutathioneS-transferase (GST)-CBD fusion protein made from mammalian CBP in a dos-age-sensitive manner.
Plasmid constructions. Various dCBP fragments (from pBSK-dCBP [2]) wereinserted by PCR cloning into the pGEX-KG vector (Pharmacia) to generatepGST-dCBP-825-1043, pGST-dCBP-1699-1997, and pGST-dCBP 2274-2508,corresponding, respectively, to the CBD, BrZn, and C/H3 domains. The BglIIrestriction fragment of pBKS-ASH1 (kindly provided by Allen Shearn, JohnsHopkins University) was inserted into a pCITE vector (Novagen) for in vitrotranslation of the nearly full-length ASH1 (amino acids [aa] 49 to 2011). OtherASH1 fragments were inserted by PCR cloning into the pCITE vector for in vitrotranslation of ASH1-47-456, ASH1-458-853, ASH1-855-1255, ASH1-1639-2011,and the ASH1-SET domain (aa 1245 to 1525). Two N-terminal fragments ofASH1 were inserted by PCR cloning into the pGEX-KG vector to generatepGST-ASH1-47-456 and pGST-ASH1-458-853.
In vitro binding assays. GST fusion proteins were produced in Escherichia coliBL21 and purified by affinity on GST-agarose beads according to the Pharmaciaprotocol. In vitro-translated [35S]methionine-labeled proteins were incubatedwith immobilized GST fusion proteins for 1.5 h at room temperature in Harlowbuffer (50 mM HEPES [pH 7.5], 100 mM NaCl, 0.2 mM EDTA, 0.01 mM NaF,1 mM dithiothreitol, 0.5% NP-40) with 5 mg of bovine serum albumin (BSA)/ml.After four washes in high-stringency Harlow buffer (containing 300 mM NaCl),bound proteins were eluted by boiling in SDS loading buffer, resolved by SDS-polyacrylamide gel electrophoresis, and visualized by autoradiography. Proteinsfrom Kc cell nuclear extracts (65 mg of total protein) were incubated with
immobilized GST fusion proteins using the procedure described above. In thiscase, the incubation was performed in the presence of 2.5 mg of BSA/ml insteadof 5 mg of BSA/ml. For the E1A competition assays, proteins from Kc cellnuclear extracts were incubated with immobilized GST-ASH1-47-456 in thepresence of 1, 4, or 12 mg of purified E1A or E1A-RG2 (kindly provided by J.Lundblad, Oregon Health Sciences University). This corresponds to approxi-mately 50, 200, and 500 nmol of the E1A proteins, respectively. Incubations weredone in the absence of E1A and E1A.RG2 as controls. The presence of bounddCBP from nuclear extracts was examined by Western blotting using the dCBPchicken polyclonal antibody at a dilution of 1/1,000. The Kc cell nuclear extractswere prepared as described previously (23).
Confocal analysis of polytene chromosomes. Polytene chromosomes werefixed and squashed for immunohistochemistry as described previously (83).ASH1 was detected by the ASH1 rabbit polyclonal antibody at a dilution of 1/40,and dCBP was detected by the chicken antiserum at a dilution of 1/800. Fluo-rescein anti-rabbit (Vector) and rhodamine anti-chicken (Jackson) secondaryantibodies were used at a dilution of 1/200. Polytene chromosomes were exam-ined by confocal laser scanning microscopy.
RESULTS
Dominant adult phenotypes obtained by overexpressingdCBP in specific cell types. Our goal was to obtain dominantadult phenotypes by overexpressing dCBP in specific cell typesduring development. To generate transgenic flies that overex-press dCBP in various tissues, we used the yeast GAL4 en-hancer trap system (8). We generated two independent trans-genic lines, Tr21 and Tr36, that carry the UAS-dCBP construct.These two lines are viable and show no visible phenotypes. Wecrossed the Tr21 and Tr36 transgenic lines to approximately 50different enhancer trap GAL4-expressing lines and character-ized the progeny in which the UAS-dCBP transgene is tran-scribed in a specific GAL4-dependent pattern. In most cases,overexpression of dCBP resulted in lethality at different stagesof the development. A few GAL4 strains generated visibleadult phenotypes with the Tr21-UAS-dCBP line (Table 1). Twoof the dominant phenotypes had 100% penetrance: a smooth-eye phenotype using the GMR-GAL4 driver (this phenotypewill be described in a separate paper) and an uninflated-wingphenotype using the GAL4-386 line (Fig. 1A).
To confirm that the wing phenotype resulted from the over-expression of dCBP, we determined that a strong loss-of-func-tion dCBP mutation, nej3, could suppress the phenotype. Wealso observed that the uninflated-wing phenotype is tempera-ture sensitive. At 25°C, the uninflated-wing phenotype is com-pletely penetrant, while only 40% of flies have the phenotypeat 18°C. At 28°C, most of the flies die as pharate adults.
To identify the cells in which the GAL4-386 driver activatestranscription, the GAL4-386 line was crossed to a UAS-LacZline and b-Gal activity in the progeny was analyzed. No ex-pression was detected during embryogenesis. During larval
TABLE 1. Pattern of GAL4 expression in larval and late pupal stages and the phenotypes obtained by crossing Tr21 (UAS-dCBP) withdifferent GAL4 lines
GAL4 lineExpression in:
Phenotype with Tr21 at 25°CThird-instar larva Pharate adult
386 Strong expression in specificneurons of the CNS
Strong expression in specificneurons of the CNS
Uninflated wing only; some died aspharate adults
363 Specific neurons of the CNS;similar but not identical toGAL4-386 (data not shown)
Specific neurons of the CNS Uninflated, partially unfolded, or curlywings (40%) or wild type; some diedas pharate adults
30A Wing and leg discs, CNS Strong expression throughoutwing blade
Reduced wing, some wings withblisters; Some died as pharate adults
71B Proximal eye-antennal disc,central wing disc, stalk in theleg disc, brain, ring gland
Strong expression restricted towing base
Most died as pharate adults; survivorswere minute or wild type
GMR Specific expression in the eye disc Strong expression in the eye lobes Smooth eyes only
VOL. 20, 2000 The dCBP-ASH1 INTERACTION 9319
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.
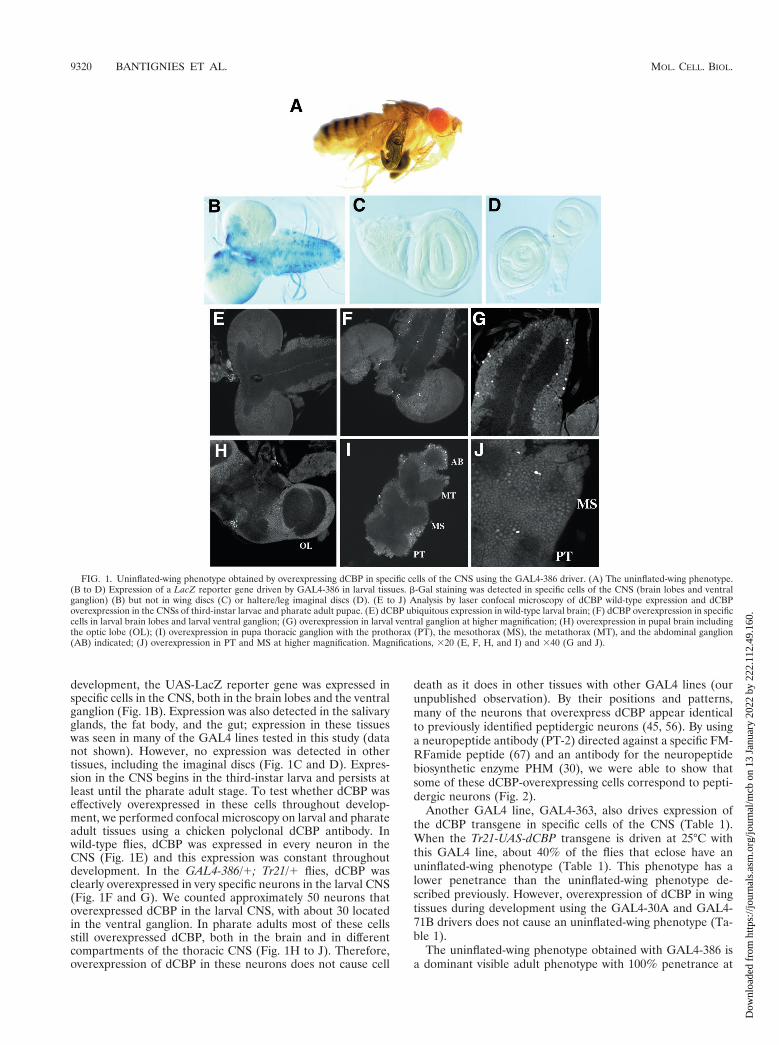
development, the UAS-LacZ reporter gene was expressed inspecific cells in the CNS, both in the brain lobes and the ventralganglion (Fig. 1B). Expression was also detected in the salivaryglands, the fat body, and the gut; expression in these tissueswas seen in many of the GAL4 lines tested in this study (datanot shown). However, no expression was detected in othertissues, including the imaginal discs (Fig. 1C and D). Expres-sion in the CNS begins in the third-instar larva and persists atleast until the pharate adult stage. To test whether dCBP waseffectively overexpressed in these cells throughout develop-ment, we performed confocal microscopy on larval and pharateadult tissues using a chicken polyclonal dCBP antibody. Inwild-type flies, dCBP was expressed in every neuron in theCNS (Fig. 1E) and this expression was constant throughoutdevelopment. In the GAL4-386/1; Tr21/1 flies, dCBP wasclearly overexpressed in very specific neurons in the larval CNS(Fig. 1F and G). We counted approximately 50 neurons thatoverexpressed dCBP in the larval CNS, with about 30 locatedin the ventral ganglion. In pharate adults most of these cellsstill overexpressed dCBP, both in the brain and in differentcompartments of the thoracic CNS (Fig. 1H to J). Therefore,overexpression of dCBP in these neurons does not cause cell
death as it does in other tissues with other GAL4 lines (ourunpublished observation). By their positions and patterns,many of the neurons that overexpress dCBP appear identicalto previously identified peptidergic neurons (45, 56). By usinga neuropeptide antibody (PT-2) directed against a specific FM-RFamide peptide (67) and an antibody for the neuropeptidebiosynthetic enzyme PHM (30), we were able to show thatsome of these dCBP-overexpressing cells correspond to pepti-dergic neurons (Fig. 2).
Another GAL4 line, GAL4-363, also drives expression ofthe dCBP transgene in specific cells of the CNS (Table 1).When the Tr21-UAS-dCBP transgene is driven at 25°C withthis GAL4 line, about 40% of the flies that eclose have anuninflated-wing phenotype (Table 1). This phenotype has alower penetrance than the uninflated-wing phenotype de-scribed previously. However, overexpression of dCBP in wingtissues during development using the GAL4-30A and GAL4-71B drivers does not cause an uninflated-wing phenotype (Ta-ble 1).
The uninflated-wing phenotype obtained with GAL4-386 isa dominant visible adult phenotype with 100% penetrance at
FIG. 1. Uninflated-wing phenotype obtained by overexpressing dCBP in specific cells of the CNS using the GAL4-386 driver. (A) The uninflated-wing phenotype.(B to D) Expression of a LacZ reporter gene driven by GAL4-386 in larval tissues. b-Gal staining was detected in specific cells of the CNS (brain lobes and ventralganglion) (B) but not in wing discs (C) or haltere/leg imaginal discs (D). (E to J) Analysis by laser confocal microscopy of dCBP wild-type expression and dCBPoverexpression in the CNSs of third-instar larvae and pharate adult pupae. (E) dCBP ubiquitous expression in wild-type larval brain; (F) dCBP overexpression in specificcells in larval brain lobes and larval ventral ganglion; (G) overexpression in larval ventral ganglion at higher magnification; (H) overexpression in pupal brain includingthe optic lobe (OL); (I) overexpression in pupa thoracic ganglion with the prothorax (PT), the mesothorax (MS), the metathorax (MT), and the abdominal ganglion(AB) indicated; (J) overexpression in PT and MS at higher magnification. Magnifications, 320 (E, F, H, and I) and 340 (G and J).
9320 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.
25°C. Therefore, this phenotype was used to screen for sup-pressors of dCBP overactivity.
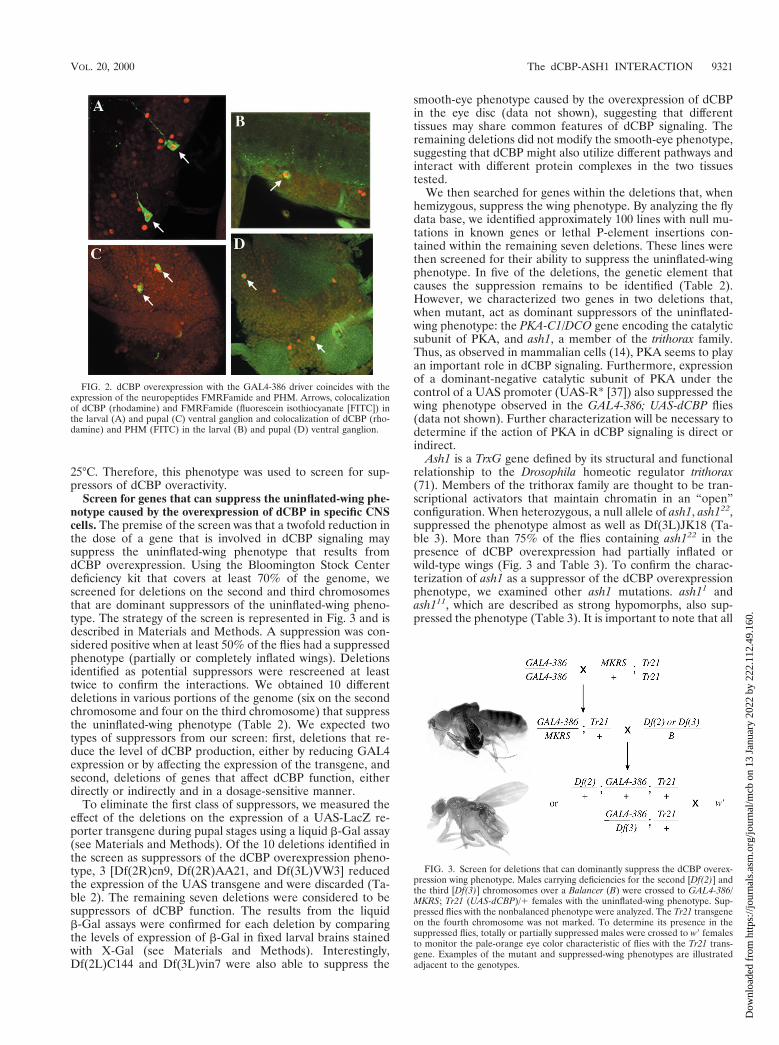
Screen for genes that can suppress the uninflated-wing phe-notype caused by the overexpression of dCBP in specific CNScells. The premise of the screen was that a twofold reduction inthe dose of a gene that is involved in dCBP signaling maysuppress the uninflated-wing phenotype that results fromdCBP overexpression. Using the Bloomington Stock Centerdeficiency kit that covers at least 70% of the genome, wescreened for deletions on the second and third chromosomesthat are dominant suppressors of the uninflated-wing pheno-type. The strategy of the screen is represented in Fig. 3 and isdescribed in Materials and Methods. A suppression was con-sidered positive when at least 50% of the flies had a suppressedphenotype (partially or completely inflated wings). Deletionsidentified as potential suppressors were rescreened at leasttwice to confirm the interactions. We obtained 10 differentdeletions in various portions of the genome (six on the secondchromosome and four on the third chromosome) that suppressthe uninflated-wing phenotype (Table 2). We expected twotypes of suppressors from our screen: first, deletions that re-duce the level of dCBP production, either by reducing GAL4expression or by affecting the expression of the transgene, andsecond, deletions of genes that affect dCBP function, eitherdirectly or indirectly and in a dosage-sensitive manner.
To eliminate the first class of suppressors, we measured theeffect of the deletions on the expression of a UAS-LacZ re-porter transgene during pupal stages using a liquid b-Gal assay(see Materials and Methods). Of the 10 deletions identified inthe screen as suppressors of the dCBP overexpression pheno-type, 3 [Df(2R)cn9, Df(2R)AA21, and Df(3L)VW3] reducedthe expression of the UAS transgene and were discarded (Ta-ble 2). The remaining seven deletions were considered to besuppressors of dCBP function. The results from the liquidb-Gal assays were confirmed for each deletion by comparingthe levels of expression of b-Gal in fixed larval brains stainedwith X-Gal (see Materials and Methods). Interestingly,Df(2L)C144 and Df(3L)vin7 were also able to suppress the
smooth-eye phenotype caused by the overexpression of dCBPin the eye disc (data not shown), suggesting that differenttissues may share common features of dCBP signaling. Theremaining deletions did not modify the smooth-eye phenotype,suggesting that dCBP might also utilize different pathways andinteract with different protein complexes in the two tissuestested.
We then searched for genes within the deletions that, whenhemizygous, suppress the wing phenotype. By analyzing the flydata base, we identified approximately 100 lines with null mu-tations in known genes or lethal P-element insertions con-tained within the remaining seven deletions. These lines werethen screened for their ability to suppress the uninflated-wingphenotype. In five of the deletions, the genetic element thatcauses the suppression remains to be identified (Table 2).However, we characterized two genes in two deletions that,when mutant, act as dominant suppressors of the uninflated-wing phenotype: the PKA-C1/DCO gene encoding the catalyticsubunit of PKA, and ash1, a member of the trithorax family.Thus, as observed in mammalian cells (14), PKA seems to playan important role in dCBP signaling. Furthermore, expressionof a dominant-negative catalytic subunit of PKA under thecontrol of a UAS promoter (UAS-R* [37]) also suppressed thewing phenotype observed in the GAL4-386; UAS-dCBP flies(data not shown). Further characterization will be necessary todetermine if the action of PKA in dCBP signaling is direct orindirect.
Ash1 is a TrxG gene defined by its structural and functionalrelationship to the Drosophila homeotic regulator trithorax(71). Members of the trithorax family are thought to be tran-scriptional activators that maintain chromatin in an “open”configuration. When heterozygous, a null allele of ash1, ash122,suppressed the phenotype almost as well as Df(3L)JK18 (Ta-ble 3). More than 75% of the flies containing ash122 in thepresence of dCBP overexpression had partially inflated orwild-type wings (Fig. 3 and Table 3). To confirm the charac-terization of ash1 as a suppressor of the dCBP overexpressionphenotype, we examined other ash1 mutations. ash11 andash111, which are described as strong hypomorphs, also sup-pressed the phenotype (Table 3). It is important to note that all
FIG. 2. dCBP overexpression with the GAL4-386 driver coincides with theexpression of the neuropeptides FMRFamide and PHM. Arrows, colocalizationof dCBP (rhodamine) and FMRFamide (fluorescein isothiocyanate [FITC]) inthe larval (A) and pupal (C) ventral ganglion and colocalization of dCBP (rho-damine) and PHM (FITC) in the larval (B) and pupal (D) ventral ganglion.
FIG. 3. Screen for deletions that can dominantly suppress the dCBP overex-pression wing phenotype. Males carrying deficiencies for the second [Df(2)] andthe third [Df(3)] chromosomes over a Balancer (B) were crossed to GAL4-386/MKRS; Tr21 (UAS-dCBP)/1 females with the uninflated-wing phenotype. Sup-pressed flies with the nonbalanced phenotype were analyzed. The Tr21 transgeneon the fourth chromosome was not marked. To determine its presence in thesuppressed flies, totally or partially suppressed males were crossed to w9 femalesto monitor the pale-orange eye color characteristic of flies with the Tr21 trans-gene. Examples of the mutant and suppressed-wing phenotypes are illustratedadjacent to the genotypes.
VOL. 20, 2000 The dCBP-ASH1 INTERACTION 9321
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

three alleles came from different sources. Thus, the observedeffect on the wing phenotype is very likely due to the loss ofash1 gene function rather than an unknown modifier on thechromosome. Two weaker alleles of ash1, ash129, a hypomor-phic allele, and ash114, a heat-sensitive hypomorph, did notsuppress the phenotype (Table 3). In these cases, the productsof both alleles probably retain some degree of function andmight still contribute to the effect of dCBP overactivity.
It is interesting that ash1 was the only trithorax member,among the ones tested, capable of suppressing the dCBP wingphenotype. The ash2, brahma, trithorax, and trithorax-like al-leles were not able to suppress the phenotype (Table 3). Asexpected, an allele of Polycomb was similarly unable to sup-press the phenotype. These results indicate that the geneticinteraction between ash1 and dCBP is very specific and wasonly apparent because we were examining dCBP signaling invery specific cells.
Specific expression of ASH1 protein in the same CNS cellsthat overexpress dCBP. To understand more precisely the re-lationship between dCBP and ASH1, we analyzed their local-ization at the cellular level. Using confocal laser microscopy,we first localized the expression of ASH1 protein in the CNSwhen dCBP is expressed under the control of GAL4-386. Forthis study, we used an ASH1 rabbit polyclonal antibody and thedCBP chicken polyclonal antibody produced in our laboratory.ASH1 was expressed in specific neurons in the larval ventralganglion (Fig. 4A) and the pupal and adult thoracic ganglion
(Fig. 4D, G, and J). No expression of ASH1 was detected in thebrain, either in third-instar larvae, late pupae, or adults (datanot shown). dCBP, which is normally expressed in every neu-ron of the CNS, was overexpressed in specific neurons whenplaced under the control of the GAL4-386 driver (Fig. 4B, E,H, and K). The overexpression of dCBP is seen both in thebrain and the ventral ganglia at larval, pupal, and adult stages.We then determined whether dCBP and ASH1 were colocal-ized. In the ventral ganglia of third-instar larvae, approxi-mately two-thirds of the overexpressing dCBP neurons wereASH1 positive (Fig. 4C). The same high level of coexpressionwas observed in the thoracic CNSs of pharate adults beforeeclosion and of adults 24 h posteclosion (Fig. 4F, I, and L). Inthe prothorax and mesothorax, 12 neurons that overexpresseddCBP expressed ASH1 as well (Fig. 4F). In the metathorax andthe abdominal ganglion, about half of the dCBP-overexpress-ing cells were ASH1 positive (Fig. 4I), representing also about12 neurons. Colocalization of ASH1 and dCBP overexpressionwas still observed in the 12 ASH1-positive cells that persisted24 h after eclosion (Fig. 4L).
It is conceivable that the overexpression of dCBP in oursystem could result in ASH1 misexpression. Therefore, wecompared the pattern and the level of CNS expression ofASH1 in dCBP-overexpressing flies with those in wild-typeflies (Fig. 5A to F). Similar patterns and levels of expressionwere obtained in larvae of both genotypes (Fig. 5, comparepanels A and D and panels C and F from merged images) in
TABLE 2. Deletions that can dominantly suppress the dCBP overexpression wing phenotype and suppressorgenes contained in these deletions
Suppressor deletion Breakpoints b-Galexpressiona Suppressor geneb
Df(2L)C144 23A1-2, 23C3-5 1 NIIn(2LR)DTD16[L]DTD42[R] 23C, 23E3-6 1 NIDf(2L)s1402 30C1-2, 30F 1 PKA-C1/DCODf(2R)cn9 42E, 44C 2 ndDf(2R)AA21 56F9-17, 57D11-12, 56D-E, 58E-F(In) 2 ndDf(2R)ES1 60E6-8, 60F1-2 1 NIDf(3L)vin7 68C8-11, 69B4-5 1 NIDf(3L)VW3 76A3, 76B2 2 ndDf(3L)JK18 76B1-2, 76D5 1 ash1In(3R)Ubx7LLatsR 96A1-7, 96A21-25 1 NI
a Effect of the deletion on the expression of a UAS-LacZ reporter gene compared to that for a balancer chromosome (see Materials and Methods). 1, no effect; 2,negative effect.
b NI, no suppressor gene identified; nd, not determined.
TABLE 3. Suppression of the uninflated-wing phenotype by the ash1 alleles and alleles of the trxG or PcG families
Allelesor deletiona
Cytological locationor breakpoints Allele class Suppression of the
wing phenotypeb
ash1 or Df(3L)JK18 76B1-2, 76D5 Deletion 111ash122 76B6 Amorph 111ash11 76B6 Strong hypomorph 111ash111 76B6 Strong hypomorph 11ash129 76B6 Hypomorph 2ash114 76B6 Weak hypomorph (heat sensitive) 2ash21 96A17 Amorph 2Df(3L)brm11 71F1-4, 72D1-10 Deletion 2brm2trxE2ca1 72A3 (brm), 88B3 (trx) Amorphs 2Df(3L)fz-M21 70D2-3, 71E4-5 Deletion including trl 2Pc1 78C9 Amorph 2
a Allele abbreviations: brm, brahma; trx, trithorax; trl, trithorax-like; Pc, polycomb.b 111, more than 75% of flies with a suppressed-wing phenotype (partial or total suppression); 11, more than 50% of flies with a suppressed-wing phenotype
(partial or total suppression); 2, no flies with a suppressed-wing phenotype.
9322 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

pupae, and in posteclosion adults (data not shown). Thus, theoverexpression of dCBP does not modify the expression ofASH1. We also examined the colocalization of the two pro-teins in a wild-type larval ganglion under higher magnification(Fig. 5G to I). Although ASH1 and dCBP proteins overlappedin expression, the two proteins did not have precisely the samelocalization pattern. dCBP was detected only in the nucleus,while ASH1 was present both within the nucleus and alsoaround the nuclear border. Importantly, overexpression ofdCBP did not modify this characteristic ASH1 distributionpattern (Fig. 5, compare panels A and D and panels C and F).
We previously used the b-Gal expression test to show thatneither the deletion containing ash1 (Table 2) nor the amor-phic ash1 mutation (data not shown) affected expression of aUAS-LacZ reporter gene. However, the regulation by proteinsinvolved in chromatin organization, such as trxG proteins, canbe sensitive to position effects. Because the UAS-LacZ andUAS-dCBP constructs are not inserted at the same position inthe genome, it was necessary to verify that an ash1 loss-of-function mutant, when heterozygous, has no effect on the levelof expression of the UAS-dCBP transgene. GAL4-386/1;Tr21/1 females with uninflated wings were crossed to ash122/
FIG. 4. ASH1 expression coincides with dCBP overexpression in the CNS of GAL4-386/1; Tr21/1 larvae and pupae. An analysis by laser confocal microscopy wasperformed. (A to C) Colocalization in the thoracic region of a third-instar larval ventral ganglion. (A) ASH1 immunostaining with fluorescein isothiocyanate (FITC);(B) dCBP immunostaining with rhodamine; (C) merged image from panels A and B. (D to F) Colocalization in the thoracic CNSs, prothoraxes (PT), and mesothoraxes(MS) of pharate adult pupae. (D) ASH1-FITC immunostaining; (E) dCBP-rhodamine immunostaining; (F) merged image from panels D and E. (G to I) Colocalizationin the thoracic CNSs metathoraxes (MT), and abdominal ganglia (AB) of pharate adult pupae. (G) ASH1-FITC immunostaining; (H) dCBP-rhodamine immuno-staining; (I) merged image from panels G and H. Magnification, 340.
VOL. 20, 2000 The dCBP-ASH1 INTERACTION 9323
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.
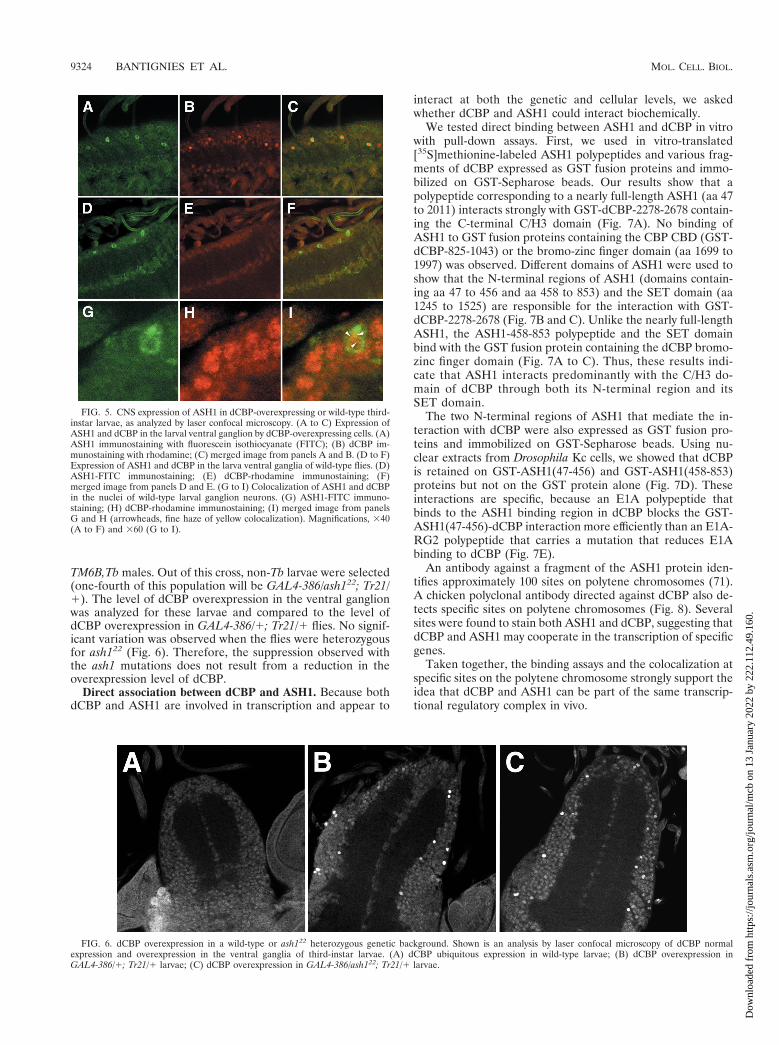
TM6B,Tb males. Out of this cross, non-Tb larvae were selected(one-fourth of this population will be GAL4-386/ash122; Tr21/1). The level of dCBP overexpression in the ventral ganglionwas analyzed for these larvae and compared to the level ofdCBP overexpression in GAL4-386/1; Tr21/1 flies. No signif-icant variation was observed when the flies were heterozygousfor ash122 (Fig. 6). Therefore, the suppression observed withthe ash1 mutations does not result from a reduction in theoverexpression level of dCBP.
Direct association between dCBP and ASH1. Because bothdCBP and ASH1 are involved in transcription and appear to
interact at both the genetic and cellular levels, we askedwhether dCBP and ASH1 could interact biochemically.
We tested direct binding between ASH1 and dCBP in vitrowith pull-down assays. First, we used in vitro-translated[35S]methionine-labeled ASH1 polypeptides and various frag-ments of dCBP expressed as GST fusion proteins and immo-bilized on GST-Sepharose beads. Our results show that apolypeptide corresponding to a nearly full-length ASH1 (aa 47to 2011) interacts strongly with GST-dCBP-2278-2678 contain-ing the C-terminal C/H3 domain (Fig. 7A). No binding ofASH1 to GST fusion proteins containing the CBP CBD (GST-dCBP-825-1043) or the bromo-zinc finger domain (aa 1699 to1997) was observed. Different domains of ASH1 were used toshow that the N-terminal regions of ASH1 (domains contain-ing aa 47 to 456 and aa 458 to 853) and the SET domain (aa1245 to 1525) are responsible for the interaction with GST-dCBP-2278-2678 (Fig. 7B and C). Unlike the nearly full-lengthASH1, the ASH1-458-853 polypeptide and the SET domainbind with the GST fusion protein containing the dCBP bromo-zinc finger domain (Fig. 7A to C). Thus, these results indi-cate that ASH1 interacts predominantly with the C/H3 do-main of dCBP through both its N-terminal region and itsSET domain.
The two N-terminal regions of ASH1 that mediate the in-teraction with dCBP were also expressed as GST fusion pro-teins and immobilized on GST-Sepharose beads. Using nu-clear extracts from Drosophila Kc cells, we showed that dCBPis retained on GST-ASH1(47-456) and GST-ASH1(458-853)proteins but not on the GST protein alone (Fig. 7D). Theseinteractions are specific, because an E1A polypeptide thatbinds to the ASH1 binding region in dCBP blocks the GST-ASH1(47-456)-dCBP interaction more efficiently than an E1A-RG2 polypeptide that carries a mutation that reduces E1Abinding to dCBP (Fig. 7E).
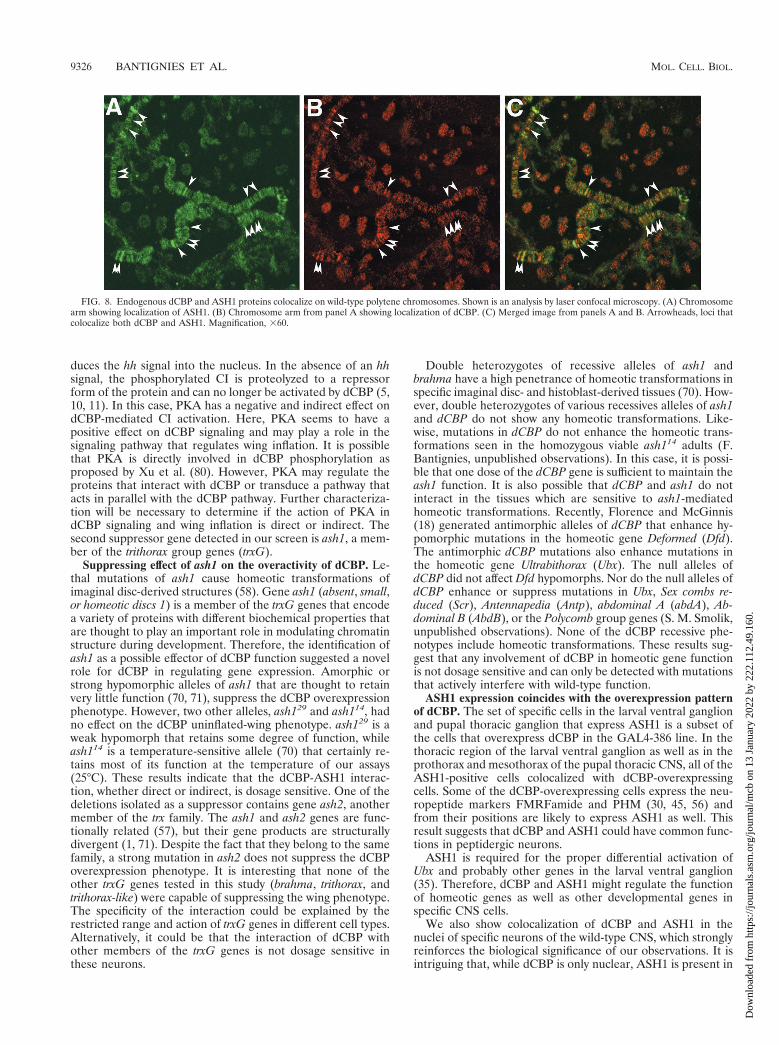
An antibody against a fragment of the ASH1 protein iden-tifies approximately 100 sites on polytene chromosomes (71).A chicken polyclonal antibody directed against dCBP also de-tects specific sites on polytene chromosomes (Fig. 8). Severalsites were found to stain both ASH1 and dCBP, suggesting thatdCBP and ASH1 may cooperate in the transcription of specificgenes.
Taken together, the binding assays and the colocalization atspecific sites on the polytene chromosome strongly support theidea that dCBP and ASH1 can be part of the same transcrip-tional regulatory complex in vivo.
FIG. 5. CNS expression of ASH1 in dCBP-overexpressing or wild-type third-instar larvae, as analyzed by laser confocal microscopy. (A to C) Expression ofASH1 and dCBP in the larval ventral ganglion by dCBP-overexpressing cells. (A)ASH1 immunostaining with fluorescein isothiocyanate (FITC); (B) dCBP im-munostaining with rhodamine; (C) merged image from panels A and B. (D to F)Expression of ASH1 and dCBP in the larva ventral ganglia of wild-type flies. (D)ASH1-FITC immunostaining; (E) dCBP-rhodamine immunostaining; (F)merged image from panels D and E. (G to I) Colocalization of ASH1 and dCBPin the nuclei of wild-type larval ganglion neurons. (G) ASH1-FITC immuno-staining; (H) dCBP-rhodamine immunostaining; (I) merged image from panelsG and H (arrowheads, fine haze of yellow colocalization). Magnifications, 340(A to F) and 360 (G to I).
FIG. 6. dCBP overexpression in a wild-type or ash122 heterozygous genetic background. Shown is an analysis by laser confocal microscopy of dCBP normalexpression and overexpression in the ventral ganglia of third-instar larvae. (A) dCBP ubiquitous expression in wild-type larvae; (B) dCBP overexpression inGAL4-386/1; Tr21/1 larvae; (C) dCBP overexpression in GAL4-386/ash122; Tr21/1 larvae.
9324 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

DISCUSSION
Previous studies have shown that CBP affects transcriptionthrough interactions with components of the basal transcrip-tional machinery and through its intrinsic and associatedacetyltransferase activities. In this report, we used a geneticapproach in Drosophila to further examine the in vivo functionof dCBP. Overactivity of dCBP in particular cell types causesseveral distinct adult phenotypes. By screening for deletionsthat could suppress one dCBP overexpression phenotype, weidentified ASH1, a member of the trithorax group of chroma-tin modifiers, as a potential interacting partner of dCBP. ASH1and dCBP colocalize to a subset of CNS neurons and to spe-cific bands in polytene chromosomes. Furthermore, dCBP andASH1 interact specifically at the molecular level. Our geneticand biochemical analyses link dCBP to a second class of pro-teins involved in epigenetic gene regulation.
Screen for suppressor genes of the uninflated-wing pheno-type. Despite the fact that the dCBP-overexpressing flies with
an uninflated-wing phenotype were weak and difficult to cul-ture, they were fertile and could be used for our screen. Theuse of the deficiency kit allowed us to rapidly define regions ofthe genome that contain genes capable of influencing the effectof dCBP overexpression. In five deletions, the genetic elementsthat cause the suppressions remain unknown and must becharacterized. We anticipate that the sequencing of the entireDrosophila genome and the generation of more P-elementmutations in these regions will help us to identify novel dCBPinteractors.
In this report, we identified two genes in two deletions that,when hemizygous, suppress the uninflated-wing phenotype.The first suppressor gene is the PKA-C1/DCO gene encodingthe catalytic subunit of the PKA. Interestingly, PKA has beeninvolved in different pathways that require dCBP activity, bothin mammalian cells and in Drosophila. In Drosophila, PKAnegatively regulates the hedgehog (hh) signal transduction cas-cade by phosphorylating CI, the transcription factor that trans-
FIG. 7. dCBP interacts with ASH1. (A) Equimolar amounts of immobilized GST and GST-dCBP fusion proteins were incubated with in vitro-translated 35S-labeledASH1 protein (nearly full-length ASH1 protein; aa 49 to 2011). (B) Equimolar amounts of immobilized GST fusion proteins were incubated with various invitro-translated 35S-labeled ASH1 fragments. The most C-terminal ASH1 fragment (aa 1639 to 2011) contains the PHD domain, but none of these fragments containthe SET domain. (C) Equimolar amounts of immobilized GST fusion proteins were incubated with the in vitro-translated 35S-labeled SET domain. (D) Equimolaramounts of immobilized GST and GST-ASH1 fusion proteins were incubated with Kc cell nuclear extracts. dCBP was detected by Western blotting using the dCBPchicken polyclonal antibody. (E) GST-ASH1-47-456 fusion protein was incubated with Kc cell nuclear extracts in the presence of 1, 4, or 12 mg of E1A or E1A-RG2.Similar results were obtained for GST-ASH1-458-853 (data not shown).
VOL. 20, 2000 The dCBP-ASH1 INTERACTION 9325
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.
duces the hh signal into the nucleus. In the absence of an hhsignal, the phosphorylated CI is proteolyzed to a repressorform of the protein and can no longer be activated by dCBP (5,10, 11). In this case, PKA has a negative and indirect effect ondCBP-mediated CI activation. Here, PKA seems to have apositive effect on dCBP signaling and may play a role in thesignaling pathway that regulates wing inflation. It is possiblethat PKA is directly involved in dCBP phosphorylation asproposed by Xu et al. (80). However, PKA may regulate theproteins that interact with dCBP or transduce a pathway thatacts in parallel with the dCBP pathway. Further characteriza-tion will be necessary to determine if the action of PKA indCBP signaling and wing inflation is direct or indirect. Thesecond suppressor gene detected in our screen is ash1, a mem-ber of the trithorax group genes (trxG).
Suppressing effect of ash1 on the overactivity of dCBP. Le-thal mutations of ash1 cause homeotic transformations ofimaginal disc-derived structures (58). Gene ash1 (absent, small,or homeotic discs 1) is a member of the trxG genes that encodea variety of proteins with different biochemical properties thatare thought to play an important role in modulating chromatinstructure during development. Therefore, the identification ofash1 as a possible effector of dCBP function suggested a novelrole for dCBP in regulating gene expression. Amorphic orstrong hypomorphic alleles of ash1 that are thought to retainvery little function (70, 71), suppress the dCBP overexpressionphenotype. However, two other alleles, ash129 and ash114, hadno effect on the dCBP uninflated-wing phenotype. ash129 is aweak hypomorph that retains some degree of function, whileash114 is a temperature-sensitive allele (70) that certainly re-tains most of its function at the temperature of our assays(25°C). These results indicate that the dCBP-ASH1 interac-tion, whether direct or indirect, is dosage sensitive. One of thedeletions isolated as a suppressor contains gene ash2, anothermember of the trx family. The ash1 and ash2 genes are func-tionally related (57), but their gene products are structurallydivergent (1, 71). Despite the fact that they belong to the samefamily, a strong mutation in ash2 does not suppress the dCBPoverexpression phenotype. It is interesting that none of theother trxG genes tested in this study (brahma, trithorax, andtrithorax-like) were capable of suppressing the wing phenotype.The specificity of the interaction could be explained by therestricted range and action of trxG genes in different cell types.Alternatively, it could be that the interaction of dCBP withother members of the trxG genes is not dosage sensitive inthese neurons.
Double heterozygotes of recessive alleles of ash1 andbrahma have a high penetrance of homeotic transformations inspecific imaginal disc- and histoblast-derived tissues (70). How-ever, double heterozygotes of various recessives alleles of ash1and dCBP do not show any homeotic transformations. Like-wise, mutations in dCBP do not enhance the homeotic trans-formations seen in the homozygous viable ash114 adults (F.Bantignies, unpublished observations). In this case, it is possi-ble that one dose of the dCBP gene is sufficient to maintain theash1 function. It is also possible that dCBP and ash1 do notinteract in the tissues which are sensitive to ash1-mediatedhomeotic transformations. Recently, Florence and McGinnis(18) generated antimorphic alleles of dCBP that enhance hy-pomorphic mutations in the homeotic gene Deformed (Dfd).The antimorphic dCBP mutations also enhance mutations inthe homeotic gene Ultrabithorax (Ubx). The null alleles ofdCBP did not affect Dfd hypomorphs. Nor do the null alleles ofdCBP enhance or suppress mutations in Ubx, Sex combs re-duced (Scr), Antennapedia (Antp), abdominal A (abdA), Ab-dominal B (AbdB), or the Polycomb group genes (S. M. Smolik,unpublished observations). None of the dCBP recessive phe-notypes include homeotic transformations. These results sug-gest that any involvement of dCBP in homeotic gene functionis not dosage sensitive and can only be detected with mutationsthat actively interfere with wild-type function.
ASH1 expression coincides with the overexpression patternof dCBP. The set of specific cells in the larval ventral ganglionand pupal thoracic ganglion that express ASH1 is a subset ofthe cells that overexpress dCBP in the GAL4-386 line. In thethoracic region of the larval ventral ganglion as well as in theprothorax and mesothorax of the pupal thoracic CNS, all of theASH1-positive cells colocalized with dCBP-overexpressingcells. Some of the dCBP-overexpressing cells express the neu-ropeptide markers FMRFamide and PHM (30, 45, 56) andfrom their positions are likely to express ASH1 as well. Thisresult suggests that dCBP and ASH1 could have common func-tions in peptidergic neurons.
ASH1 is required for the proper differential activation ofUbx and probably other genes in the larval ventral ganglion(35). Therefore, dCBP and ASH1 might regulate the functionof homeotic genes as well as other developmental genes inspecific CNS cells.
We also show colocalization of dCBP and ASH1 in thenuclei of specific neurons of the wild-type CNS, which stronglyreinforces the biological significance of our observations. It isintriguing that, while dCBP is only nuclear, ASH1 is present in
FIG. 8. Endogenous dCBP and ASH1 proteins colocalize on wild-type polytene chromosomes. Shown is an analysis by laser confocal microscopy. (A) Chromosomearm showing localization of ASH1. (B) Chromosome arm from panel A showing localization of dCBP. (C) Merged image from panels A and B. Arrowheads, loci thatcolocalize both dCBP and ASH1. Magnification, 360.
9326 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

both the nucleus and the cytoplasm surrounding the nuclearperiphery. Preparation of nuclear and cytoplasmic Kc cell ex-tracts revealed the presence of the ASH1 protein in bothcompartments (data not shown). At this time, we do not knowthe significance of this pattern of localization, but it is possiblethat ASH1 nuclear localization might be regulated throughposttranscriptional modifications.
Screens for enhancers and suppressors of overexpressionphenotypes have been useful in identifying components of reg-ulatory pathways. Nevertheless, overexpression systems havedrawbacks and can potentially identify secondary effectors of anonspecific phenotype. However, we believe that this screenhas identified genes that affect dCBP function for several rea-sons. First, the number of deficiencies that suppress the unin-flated-wing phenotype is small. A large number of suppressorsmight suggest that the overexpression of dCBP was not elicit-ing a specific cell phenotype. Second, two of the deletionssuppressed both the wing and the eye overexpression pheno-types, suggesting that the overexpression of dCBP in the twotissues has some common effects. One of the deletions dem-onstrated that the dosage of PKA could affect the dCBP over-expression phenotype. CBP and dCBP are known to play a rolein PKA signaling, so the fact that PKA was identified in thisscreen is consistent with the idea that dCBP overexpressionreflects an overactivation of the PKA pathway. We have ruledout trivial explanations for the suppression of dCBP overex-pression by ASH1; dCBP overexpression does not cause thedeath of ASH1-expressing cells, nor do ash1 mutations affectthe overexpression of dCBP. A characterization of dCBP lossof function in these cells both in wild-type and ash1 mutantbackgrounds is necessary to complete this analysis. A clonalanalysis of dCBP mutant cells is not feasible because dCBP isrequired for cell viability and only small clones can be gener-ated. This analysis will have to await reagents that allow us toknock out dCBP function in the GAL4-386 cells in the ash1mutant background. In addition, it will be important to identifythe targets of dCBP and ASH1 in these cells as well as thepathways that activate them. Although the genetic analysis isnot complete, it is likely that the genetic suppression of dCBPoverexpression by ash1 mutations reflects a functional associ-ation between ASH1 and dCBP because these two proteinshave specific interactions in vitro.
Overexpression of dCBP in specific CNS cells causes winginflation defects. In many tissues, overexpression of dCBPcauses lethality, suggesting that the dose of this effector isimportant for its function. The overproduction of dCBP inspecific cells of the CNS with two different GAL4 lines pro-duced defects in wing inflation with various degrees of pen-etrance. However, overexpression of dCBP in wing tissuesthroughout development does not interfere with wing inflation.
Previous studies have implicated specific CNS cells in theregulation of wing inflation. In Drosophila, the death of specificcells is triggered after eclosion and is strongly correlated withwing inflation behavior (29). In addition, two specific neuronsin the fly brain are responsible for the production of the neu-ropeptide eclosion hormone (EH). The specific knockout ofEH-producing cells (EH cells) during early development re-sults in eclosion delays and a disruption of eclosion behaviors,such as wing inflation (41). In the moth Manduca sexta, EHtriggers a neuroendocrine cascade that regulates both ecdysisand postecdysis processes such as wing inflation. It was sug-gested that the frequent failure of EH cell knockout flies toinflate their wings successfully is due to a lack of excitability ofneuroendocrine-responsive EH cells that release importantsignals for proper eclosion behaviors (41). In Manduca, differ-ent neuropeptides, such as bursicon and the cardioacceleratory
peptides, are usually released after eclosion to aid in wingexpansion (72, 75, 76). It may be that the neurons which over-express dCBP are the neurosecretory cells that are targeted bythe EH cascade and that produce the peptides that signal thewing inflation process. In this case, the overexpression of dCBPinterferes with normal cell function. Of course the wing infla-tion defect could be due to the death of the neurons caused bythe overexpression of dCBP. However, the pattern of cells thatoverexpress LacZ and dCBP in the GAL4-386 backgroundremains the same throughout development, and cells that over-express dCBP and express ASH1 are viable at least 24 hposteclosion, so the overexpression of dCBP does not appearto affect the viability of these cells. Two additional GAL4 lines,GAL4-c929 and GAL4-c191, also drive specific expression inthe CNS, specifically in most of the peptidergic neurons of thebrain and ventral ganglion (R. Hewes and P. Taghert, personalcommunication). At 25°C, escapers were obtained only withthe GAL4-c191 line. Approximately 30% of these flies haveuninflated or partially inflated wings.
We propose that the overexpression of dCBP in specificCNS cells affects the regulation of signaling pathways thatinvolve dCBP and that are important for proper eclosion be-haviors. Our preliminary data suggest that at least some of thecells that overexpress dCBP are neuropeptidergic neurons andcolocalize with the neuropeptides FMRFamide and PHM.However, antibody incompatibility does not allow us to deter-mine whether these cells also express ASH1. Clearly, morecharacterization will be required to determine the exact path-ways affected by dCBP. The dominant wing phenotype ob-tained by overexpressing dCBP with GAL4-386 is a goodmodel to elucidate some of the cells and signaling pathwaysinvolved in wing inflation.
Specific interaction between ASH1 and dCBP. Our bio-chemical experiments show that coactivator dCBP bindsstrongly to trxG protein ASH1. This observation supports theidea that ASH1 and dCBP interact in vivo and implicates anovel class of chromatin binding proteins in mediating dCBPfunction.
The ASH1 protein contains three motifs that are character-istic of some proteins that regulate transcription and/or arebound to chromosomes: there are two AT hook motifs in theN-terminal region, a SET domain, and a PHD finger in theC-terminal domain. The AT hook motif is important for thebinding of some proteins to DNA (52). PHD fingers are Cys-rich Zn finger-like motifs implicated in protein-protein inter-actions and are found in other trxG proteins (1, 40, 70). TheSET domain is an approximately 130-aa region found in anumber of other chromatin-associated proteins, including theTRX factor (40), PcG protein Enhancer of Zeste [E(Z)] (26),and the modifier of position effect variegation Su(Var)3-9 (73).The TRX SET domains have been proposed to mediate asso-ciation with components of chromatin-remodeling complexes(54), and ASH1 and TRX interact directly through their SETdomains (55). Our binding assays indicate that two N-terminalregions and the SET domain of ASH1 interact strongly withdCBP. However, no interaction with the PHD domain wasobserved. Thus, the SET and the PHD domains of ASH1might function for the recruitment of other chromatin-associ-ated proteins, such as TRX, and the N-terminal region couldserve to interact with the DNA, possibly through the AT mo-tifs, to direct the targeting of HATs to the promoter. Furtherbiochemical characterization will be necessary to confirm thismodel, but the interaction between dCBP and ASH1 providesnew insights on the possible function of ASH1 in gene regu-lation.
The binding of ASH1 to dCBP requires the C-terminal C/H3
VOL. 20, 2000 The dCBP-ASH1 INTERACTION 9327
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

domain. In mammalian CBP and p300, this region mediatesinteractions with numerous sequence-specific transcriptionfactors, the adenovirus E1A protein, TFIIB, RNA helicase A,and P/CAF, a GCN5-like histone acetylase (19, 25, 60). IndCBP, the C/H3 domain mediates the interaction with tran-scription factor dTCF and Mad (78, 79), demonstrating animportant role for this domain in dCBP function. Our findingsreveal that this domain contributes to the interaction withchromatin-associated protein ASH1, suggesting that dCBPmay function in epigenetic regulatory complexes. The C/H3domain is adjacent to HAT and might contribute to the regu-lation of the histone acetylation activity of CBP and p300 (32)or might recruit targets of acetylation close to the enzymaticdomain. Thus, it will be interesting to determine whetherASH1 has any effect on dCBP HAT functions or if it is a targetof dCBP acetyltransferase activity.
The recent work of Dhalluin et al. (15) has shown that thebromodomain of P/CAF binds histone peptides in an acetyla-tion-dependent manner. The bromodomain of GCN5, a mem-ber of the SAGA complex, is required for SWI/SNF remodel-ing of the nucleosome and stabilizing the SWI/SNF complex onthe promoter (66). Thus, it appears that the bromodomaininteracts with acetylated proteins and may form a link betweendifferent regulatory complexes. Although the full-length ASH1does not interact with the bromodomain of dCBP, both theASH1-458-853) polypeptide and the SET domain do interactwith this domain. It may be that full-length ASH1 undergoes amodification, upon binding with the dCBP C/H3 domain, thatallows other regions of ASH1 to interact with the dCBP bro-modomain. In this case, it would appear that the interaction isnot dependent on acetylation.
Our results also show that dCBP and ASH1 colocalized to anumber of specific sites on polytene chromosomes, suggestingthat they might serve as coregulators of a specific set of genesincluding the homeotic selector genes. The mapping of thespecific sites where dCBP and ASH1 colocalize will help us toidentify target genes that are regulated by ASH1 and dCBP.An analysis of these genes, their promoters, and their regula-tion by dCBP and ASH1 will further define the functional roleof the dCBP-ASH1 interaction.
It has been recently shown that ASH1 is a component of alarge-molecular-weight complex (47). The components of thisASH1 complex have not been identified, and it will be inter-esting to test whether it includes dCBP. BRM, another trxGmember, is also contained in a large-molecular-mass complexof 2 MDa. Four of the subunits of the BRM complex arerelated to subunits of the yeast chromatin-remodeling com-plexes SWI/SNF and RSC (remodels the structure of chroma-tin) (47), suggesting that trxG proteins are important regula-tors of chromatin structure. However, no other trxG membersare present in this complex, suggesting that each trxG memberis involved with different complexes and probably has divergentfunctions. Thus, it might be anticipated that ASH1 could bethe only trithorax member that affected the overexpression ofdCBP function.
Our genetic approach allowed us to characterize a specificcellular and biochemical interaction between dCBP and ASH1,a member of the trithorax group of chromatin modifiers. Thisfinding may provide important new insights into the functionsof both proteins. trxG proteins are thought to be importantcomponents of chromatin-remodeling complexes, and ourstudy provides evidence that they might also be involved in therecruitment of transcriptional activators. While the molecularmechanism of the interaction between dCBP and ASH1 is notknown, it probably involves the modification of chromatinstructure, and this suggests that CBP may not only affect the
nucleosome but may also be involved in the regulation ofhigher-order chromatin structure as well.
ACKNOWLEDGMENTS
We thank A. Shearn for the ash1 and ash2 mutants, ash1 cDNA, andASH1 antibody; we are grateful to J. Lundblad for the E1A andRG2.E1A proteins. We also thank D. Kalderon for the Pka mutantand transgenic flies; C. O’Kane and A. Brand for GAL4 lines; G.Rubin for the GMR-GAL4 line; J. Roote for the pk-sple33 pr cn/T(2;3)SM6.TM6B,Tb balancer chromosome strain; and R. Hewes and P.Taghert for GAL4 lines, the FMRFamide, and PHM antibodies andfor sharing unpublished information. We also thank the Bloomingtonand the Umea Drosophila Stock Centers for providing numerousstocks. We are very grateful to A. Snyder (MMI department, OHSU,and the Oregon Hearing Research Center) for confocal analysis.
This work was partly supported by grants from the Association pourla Recherche contre le Cancer and the National Institutes of Health(DK4Y239).
REFERENCES
1. Adamson, A., and A. Shearn. 1996. Molecular genetic analysis of Drosophilaash2, a member of the trithorax group required for imaginal disc patternformation. Genetics 144:621–633.
2. Akimaru, H., Y. Chen, P. Dai, D.-X. Hou, M. Nonaka, S. M. Smolik, S.Armstrong, R. H. Goodman, and S. Ishii. 1997. Drosophila CBP is a co-activator of cubitus interruptus in hedgehog signalling. Nature 386:735–738.
3. Akimaru, H., D.-X. Hou, and S. Ishii. 1998. Drosophila CBP is required fordorsal-dependent twist gene expression. Nat. Genet. 17:211–214.
4. Arany, Z., D. Newsome, E. Oldread, D. M. Livingston, and R. Eckner. 1995.A family of transcriptional adaptor proteins targeted by the E1A oncopro-tein. Nature 374:81–84.
5. Aza-Blanc, P., F.-A. Ramirez-Weber, M.-P. Laget, C. Schwartz, and T. Korn-berg. 1997. Proteolysis that is inhibited by Hedgehog targets Cubitus inter-ruptus protein to the nucleus and converts it to a repressor. Cell 89:1043–1053.
6. Bannister, A. J., and T. Kouzarides. 1996. The CBP co-activator is a histoneacetyltransferase. Nature 384:641–643.
7. Borrow, J., V. P. Stanton, Jr., J. M. Andresen, R. Becher, F. G. Behm, R. S.Chaganti, C. I. Civin, C. Disteche, I. Dube, A. M. Frischauf, D. Horsman, F.Mitelman, S. Volinia, A. E. Watmore, and D. E. Housman. 1996. The trans-location t(8;16)(p11;p13) of acute myeloid leukaemia fuses a putative acetyl-transferase to the CREB-binding protein. Nat. Genet. 14:33–41.
8. Brand, A., and N. Perrimon. 1993. Targeted gene expression as a means ofaltering cell fates and generating dominant phenotypes. Development 118:401–415.
9. Chen, H., R. J. Lin, R. L. Schiltz, D. Chakravarti, A. Nash, L. Nagy, M. L.Privalsky, Y. Nakatani, and R. M. Evans. 1997. Nuclear receptor coactivatorACTR is a novel histone acetyltransferase and forms a multimeric activationcomplex with P/CAF and CBP/p300. Cell 90:569–580.
10. Chen, Y., J.-R. Cardinaux, R. H. Goodman, and S. M. Smolik. 1999. Mutantsof cubitus interruptus that are independent of PKA regulation are indepen-dent of hedgehog signaling. Development 126:3607–3616.
11. Chen, Y., N. Gallaher, R. H. Goodman, and S. M. Smolik. 1998. Proteinkinase A directly regulates the activity and proteolysis of cubitus interruptus.Proc. Natl. Acad. Sci. USA 95:2349–2354.
12. Chen, Y., R. H. Goodman, and S. M. Smolik. 2000. Cubitus interruptusrequires Drosophila CREB-binding protein to activate wingless expression inthe Drosophila embryo. Mol. Cell. Biol. 20:1616–1625.
13. Chinwalla, V., E. P. Jane, and P. J. Harte. 1995. The Drosophila trithoraxprotein binds to specific chromosomal sites and is co-localized with Poly-comb at many sites. EMBO J. 14:2056–2065.
14. Chrivia, J. C., R. P. Kwok, N. Lamb, M. Hagiwara, M. R. Montminy, andR. H. Goodman. 1993. Phosphorylated CREB binds specifically to the nu-clear protein CBP. Nature 365:855–859.
15. Dhalluin, C., J. E. Carlson, L. Zeng, C. He, A. K. Aggarwal, and M.-M. Zhou.1999. Structure and ligand of a histone acetyltransferase bromodomain.Nature 399:491–496.
16. Dingwall, A. K., S. J. Beek, C. M. McCallum, J. W. Tamkun, G. V. Kalpana,S. P. Goff, and M. P. Scott. 1995. The Drosophila snr1 and brm proteins arerelated to yeast SWI/SNF proteins and are components of a large proteincomplex. Mol. Biol. Cell. 6:777–791.
17. Eckner, R., M. E. Ewen, D. Newsome, M. Gerdes, J. A. DeCaprio, J. B.Lawrence, and D. M. Livingston. 1994. Molecular cloning and functionalanalysis of the adenovirus E1A-associated 300-kD protein (p300) reveals aprotein with properties of a transcriptional adaptor. Genes Dev. 8:869–884.
18. Florence, B., and W. McGinnis. 1998. A genetic screen of the drosophila Xchromosome for mutations that modify Deformed function. Genetics 150:1497–1511.
9328 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

19. Giles, R. H., D. J. Peters, and M. H. Breuning. 1998. Conjunction dysfunc-tion: CBP/p300 in human disease. Trends Genet. 14:178–183.
20. Goldman, P. S., V. K. Tran, and R. H. Goodman. 1997. The multifunctionalrole of the co-activator CBP in transcriptional regulation. Recent Prog.Horm. Res. 52:103–119.
21. Goodman, R. H., and S. Smolik. 2000. CBP/p300 in cell growth, transfor-mation, and development. Genes Dev. 14:1553–1577.
22. Gu, W., and R. G. Roeder. 1997. Activation of p53 sequence-specific DNAbinding by acetylation of the p53 C-terminal domain. Cell 90:595–606.
23. Henninghausen, L., and H. Lubon. 1987. Interaction of protein with DNA invitro. Methods Enzymol. 152:721–735.
24. Imhof, A., X. J. Yang, V. V. Ogryzko, Y. Nakatani, A. P. Wolffe, and H. Ge.1997. Acetylation of general transcription factors by histone acetyltrans-ferases. Curr. Biol. 7:689–692.
25. Janknecht, R., and T. Hunter. 1996. A growing coactivator network. Nature383:22–23.
26. Jones, R. S., and W. M. Gelbart. 1993. The Drosophila Polycomb-group geneEnhancer of zeste contains a region with sequence similarity to trithorax. Mol.Cell. Biol. 13:6357–6366.
27. Kawasaki, H., R. Eckner, T. P. Yao, K. Taira, R. Chiu, D. M. Livingston, andK. K. Yokoyama. 1998. Distinct roles of the co-activators p300 and CBP inretinoic-acid-induced F9-cell differentiation. Nature 393:284–289.
28. Kee, B. L., J. Arias, and M. R. Montminy. 1996. Adaptor-mediated recruit-ment of RNA polymerase II to a signal-dependent activator. J. Biol. Chem.271:2373–2375.
29. Kimura, K. I., and J. Truman. 1990. Postmetamorphic cell death in thenervous and muscular systems of Drosophila melanogaster. J. Neurosci. 10:403–411.
30. Kolhekar, A. S., M. S. Roberts, N. Jiang, R. C. Johnson, R. E. Mains, B. A.Eipper, and P. H. Taghert. 1997. Neuropeptide amidation in Drosophila:separate genes encode the two enzymes catalyzing amidation. J. Neurosci.17:1363–1376.
31. Kraus, W. L., and J. T. Kadonaga. 1998. p300 and estrogen receptor coop-eratively activate transcription via differential enhancement of initiation andreinitiation. Genes Dev. 12:331–342.
32. Kraus, W. L., E. T. Manning, and J. T. Kadonaga. 1999. Biochemical anal-ysis of distinct activation functions in p300 that enhance transcription initi-ation with chromatin templates. Mol. Cell. Biol. 19:8123–8135.
33. Kuzin, B., S. Tillib, Y. Sedkov, L. Mizrokhi, and A. Mazo. 1994. The Dro-sophila trithorax gene encodes a chromosomal protein and directly regulatesthe region specific homeotic gene fork head. Genes Dev. 8:2478–2490.
34. Kwok, R. P., J. R. Lundblad, J. C. Chrivia, J. P. Richards, H. P. Bachinger,R. G. Brennan, S. G. Roberts, M. R. Green, and R. H. Goodman. 1994.Nuclear protein CBP is a coactivator for the transcription factor CREB.Nature 370:223–226.
35. LaJeunesse, D., and A. Shearn. 1995. Trans-regulation of thoracic homeoticselector genes of the Antennapedia and bithorax complexes by trithoraxgroup genes: absent, small, and homeotic discs 1 and 2. Mech. Dev. 53:123–139.
36. Lane, M. E., and D. Kalderon. 1993. Genetic investigation of cAMP-depen-dent protein kinase function in Drosophila development. Genes Dev.7:1229–1243.
37. Li, W., J. T. Ohlmeyer, M. E. Lane, and D. Kalderon. 1995. Function ofprotein kinase A in hedgehog signal transduction and Drosophila imaginaldisc development. Cell 80:553–562.
37a.Lindsley, D. L., and G. C. Zimm. 1992. The genome of Drosophila melano-gaster. Academic Press, San Diego, Calif.
38. Lundblad, J. R., R. P. Kwok, M. E. Laurance, M. L. Harter, and R. H.Goodman. 1995. Adenoviral E1A-associated protein p300 as a functionalhomologue of the transcriptional co-activator CBP. Nature 374:85–88.
39. Mannervik, M., Y. Nibu, H. Zhang, and M. Levine. 1999. Transcriptionalcoregulators in development. Science 284:606–609.
40. Mazo, A. M., D.-H. Huang, B. A. Mozer, and I. B. Dawid. 1990. The trithoraxgene, a trans-acting regulator of the bithorax complex in Drosophila, en-codes a protein with zinc-binding domains. Proc. Natl. Acad. Sci. USA87:2112–2116.
41. McNabb, S. L., J. D. Baker, J. Agapite, H. Steller, L. Riddiford, and J. W.Truman. 1997. Disruption of a behavioral sequence by targeted death ofpeptidergic neurons in Drosophila. Neuron 19:813–823.
42. Miller, R. W., and J. H. Rubinstein. 1995. Tumors in Rubinstein-Taybisyndrome. Am. J. Med. Genet. 56:112–115.
43. Muraoka, M., M. Konoshi, R. Kituchi-Yanoshita, K. Tanaka, N. Shitara,J.-M. Chong, T. Iwama, and M. Miyaki. 1996. p300 gene alterations incolorectal and gastric carcinomas. Oncogene 12:1565–1569.
44. Nakajima, T., C. Uchida, S. F. Anderson, C. G. Lee, J. Hurwitz, J. D. Parvin,and M. Montminy. 1997. RNA helicase A mediates association of CBP withRNA polymerase II. Cell 90:1107–1112.
45. Nassel, D. R. 1996. Neuropeptides, amines, and amino acids in an elemen-tary insect ganglion. Prog. Neurobiol. 48:325–420.
46. Ogryzko, V. V., R. L. Schiltz, V. Russanova, B. H. Howard, and Y. Nakatani.1996. The transcriptional coactivators p300 and CBP are histone acetyltrans-ferases. Cell 87:953–959.
47. Papoulas, O., S. J. Beek, S. L. Moseley, C. M. McCallum, M. Sarte, A.Shearn, and J. W. Tamkun. 1998. The Drosophila trithorax group proteinsBRM, ASH1 and ASH2 are subunits of distinct protein complexes. Devel-opment 125:3955–3966.
48. Petrij, F., R. H. Giles, H. G. Dauwerse, J. J. Saris, R. C. M. Hennekam, M.Masuno, N. Tommerup, G.-J. B. V. Ommen, R. H. Goodman, D. J. M. Peters,and M. H. Breuning. 1995. Rubinstein-Taybi syndrome caused by mutationsin the transcriptional coactivator CBP. Nature 376:348–351.
49. Pirrotta, V. 1995. Chromatin complexes regulating gene expression in Dro-sophila. Curr. Opin. Genet. Dev. 5:466–472.
50. Pirrotta, V. 1998. Polycombing the genome: PcG, trxG, and chromatin si-lencing. Cell 93:333–336.
51. Rastelli, L., C. S. Chan, and V. Pirotta. 1993. Related chromosome bindingsites for zeste, suppressors of zeste and Polycomb group proteins in Dro-sophila and their dependence on enhancers of zeste function. EMBO J.12:1513–1522.
52. Reeves, R., and M. S. Nissen. 1990. The AT DNA-binding domain of mam-malian high mobility group I proteins. A novel peptide motif for recognizingDNA structure. J. Biol. Chem. 265:8573–8582.
53. Rose, R. E., N. M. Gallaher, D. J. Andrew, R. H. Goodman, and S. M.Smolik. 1997. The CRE binding protein dCREB-A is required for Drosoph-ila embryonic development. Genetics 146:595–606.
54. Rozenblatt-Rosen, O., T. Rozovskaia, D. Buracov, Y. Sedkov, S. Tillib, J.Blechman, C. Croce, A. Mazo, and E. Canaani. 1998. The C-terminal SETdomains of ALL-1 and Trithorax interact with the INI1 and SNR1 proteins,components of the SWI/SNF complex. Proc. Natl. Acad. Sci. USA 95:4152–4157.
55. Rozovskaia, T., S. Tillib, S. Smith, Y. Sedkov, O. Rozenblatt-Rosen, S.Petruk, T. Yano, T. Nakamura, L. Ben-Simchon, J. Gildea, C. M. Croce, A.Shearn, E. Canaani, and A. Mazo. 1999. Trithorax and ASH1 interact di-rectly and associate with the trithorax group-responsive bxd region of theUltrabithorax promoter. Mol. Cell. Biol. 19:6441–6447.
56. Schneider, L. E., E. T. Sun, D. J. Garland, and P. H. Taghert. 1993. Animmunocytochemical study of the FMRFamide neuropeptide gene productsin Drosophila. J. Comp. Neurol. 337:446–460.
57. Shearn, A. 1989. The ash-1, ash-2 and trithorax genes of Drosophila melano-gaster are functionally related. Genetics 121:517–525.
58. Shearn, A., E. Hesperger, and G. Hesperger. 1987. Genetic studies of mu-tation at two loci of Drosophila melanogaster which cause a wide variety ofhomeotic transformations. Roux’s Arch. Dev. Biol. 196:231–242.
59. Shi, Y., and C. Mello. 1998. A CBP/p300 homolog specifies multiple differ-entiation pathways in Caenorhabditis elegans. Genes Dev. 12:943–955.
60. Shikama, M., J. Lyon, and N. B. Thangue. 1997. The p300/CBP family:integrating signals with transcription factors and chromatin. Trends CellBiol. 7:230–236.
61. Simon, J. 1995. Locking in stable states of gene expression: transcriptionalcontrol during Drosophila development. Curr. Opin. Cell Biol. 7:376–385.
62. Sobulo, O. M., J. Borrow, R. Tomek, S. Reshmi, A. Harden, B. Schlegel-berger, D. Housman, N. A. Doggett, J. D. Rowley, and N. J. Zeleznik-Le.1997. MLL is fused to CBP, a histone acetyltransferase, in therapy-relatedacute myeloid leukemia with a t(11;16)(q23;p13.3). Proc. Natl. Acad. Sci.USA 94:8732–8737.
63. Spencer, T. E., G. Jenster, M. M. Burcin, C. D. Allis, J. Zhou, C. A. Mizzen,N. J. McKenna, S. A. Onate, S. Y. Tsai, M. J. Tsai, and B. W. O’Malley. 1997.Steroid receptor coactivator-1 is a histone acetyltransferase. Nature 389:194–198.
64. Spradling, A. C. 1986. P element-mediated transformation, p. 175–198. InD. B. Roberts (ed.), Drosophila: a practical approach. IRL Press Limited,Oxford, United Kingdom. DC.
65. Swope, D. L., C. L. Mueller, and J. C. Chrivia. 1996. CREB-binding proteinactivates transcription through multiple domains. J. Biol. Chem. 271:28138–28145.
66. Syntichaki, P., I. Topalidou, and G. Thireos. 2000. The Gcn5 bromodomaincoordinates nucleosome remodeling. Nature 404:414–417.
67. Taghert, P. H., and L. E. Schneider. 1990. Interspecific comparison of aDrosophila gene encoding FMRFamide-related neuropeptides. J. Neurosci.10:1929–1942.
68. Tamkun, J. W., R. Deuring, M. P. Scott, M. Kissinger, A. M. Pattatucci, T. C.Kaufman, and J. A. Kennison. 1992. brahma: a regulator of Drosophilahomeotic genes structurally related to the yeast transcriptional activatorSNF2/SWI2. Cell 68:561–572.
69. Tanaka, Y., I. Naruse, T. Maekawa, H. Masuya, T. Shiroishi, and S. Ishii.1997. Abnormal skeletal patterning in embryos lacking a single Cbp allele: apartial similarity with Rubinstein-Taybi syndrome. Proc. Natl. Acad. Sci.USA 94:10215–10220.
70. Tripoulas, N. A., E. Hersperger, D. LaJeunesse, and A. Shearn. 1994. Mo-lecular genetic analysis of the Drosophila melanogaster gene absent, small orhomeotic discs (ash1). Genetics 137:1027–1038.
71. Tripoulas, N. A., D. LaJeunesse, J. Gildea, and A. Shearn. 1996. The Dro-sophila ash1 gene product, which is localized at specific sites on polytenechromosomes, contains a SET domain and a PHD finger. Genetics 143:913–928.
VOL. 20, 2000 The dCBP-ASH1 INTERACTION 9329
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.

72. Truman, J. W., and P. T. Endo. 1974. Physiology of insect ecdysis. Neuronaland hormonal factors involved in wing spreading behavior of moths. J. Exp.Biol. 61:47–55.
73. Tschiersch, B., A. Hofmann, V. Krauss, R. Dorn, G. Korge, and G. Reuter.1994. The protein encoded by the Drosophila position-effect variegationsuppressor gene Su(var)3-9 combines domains of antagonistic regulators ofhomeotic gene complexes. EMBO J. 13:3822–3831.
74. Tsukiyama, T., and C. Wu. 1997. Chromatin remodeling and transcription.Curr. Opin. Genet. Dev. 7:182–191.
75. Tublitz, N. J., and J. W. Truman. 1985. Insect cardioactive peptides. I.Distribution and molecular characterization of two cardioacceleratory pep-tides (CAP) in the tobacco hawkmoth, Manduca sexta. J. Exp. Biol. 114:365–379.
76. Tublitz, N. J., and J. W. Truman. 1985. Insect cardioactive peptides. II.Neurohormonal control of heart activity by two cardioacceleratory peptidesin the tobacco hawkmoth, Manduca sexta. J. Exp. Biol. 114:381–395.
77. Wade, P. A., and A. P. Wolffe. 1997. Histone acetyltransferases in control.Curr. Biol. 7:R82–R84.
78. Waltzer, L., and M. Bienz. 1998. Drosophila CBP represses the transcriptionfactor TCF to antagonize Wingless signaling. Nature 395:521–525.
79. Waltzer, L., and M. Bienz. 1999. A function of CBP as a transcriptionalco-activator during Dpp signalling. EMBO J. 18:1630–1641.
80. Xu, L., R. M. Lavinsky, J. S. Dasen, S. E. Flynn, E. M. McInerney, T.-M.Mullen, T. Heinzel, D. Szeto, E. Korzus, R. Kurokawa, A. K. Aggarwal, D. W.Rose, C. K. Glass, and M. G. Rosenfeld. 1998. Signal-specific co-activatordomain requirements for Pit-1 activation. Nature 395:301–306.
81. Yang, X. J., V. V. Ogryzko, J. Nishikawa, B. H. Howard, and Y. Nakatani.1996. A p300/CBP-associated factor that competes with the adenoviral on-coprotein E1A. Nature 382:319–324.
82. Yao, T. P., S. P. Oh, M. Fuchs, N.-D. Zhou, L.-E. Ch’ng, D. Newsome, R. T.Bronson, E. Li, D. M. Livingston, and R. Eckner. 1998. Gene dosage-dependent embryonic development and proliferation defects in mice lackingthe transcriptional integrator p300. Cell 93:361–372.
83. Zink, B., and R. Paro. 1989. In vivo binding of a trans-regulator of homeoticgenes in Drosophila melanogaster. Nature 337:468–471.
9330 BANTIGNIES ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
222
.112
.49.
160.





























