Embed Size (px)
Citation preview

THE JOURNAL OP BIOLOGICAL CHEMISTRY Vol. 249, No. 14,Issue of July 26, pp. 4487-4494, 1974
Printed in U.S.A.
The Activation of Bovine Pepsinogen
SEQUENCE OF THE PEPTIDES RELEASED, IDENTIFICATION OF A PEPSIN INHIBITOR*
(Received for publication, January 18, 1974)
MARIANNE HARBOE, PER MAACK ANDERSEN, AND BENT FOLTMANN
From the Institute of Biochemical Genetics, University of Copenhagen, Copenhagen K, Denmark
JOHN KAY$ AND BEATRICE KASSELL
From the Department of Biochemistry, The Medical College of Wisconsin, Milwaukee, Wisconsin 53.233
SUMMARY
Bovine pepsinogen is converted into pepsin by removal of 45 amino acid residues from the NH, terminus of the single polypeptide chain. The complete sequence of the activation peptides has been deduced from two overlapping sets of pep- tides, one set obtained by autoactivation of the zymogen at pH 2 and the second set from a tryptic digest of maleylated pepsinogen. A peptide derived from the 17 residues at the NH2 terminus of bovine pepsinogen has been identified as an inhibitor of the milk-clotting action of pepsin.
Extensive sequence homologies exist among the activation peptides of bovine and porcine pepsinogens and bovine prochymosin. In the peptides from the two pepsinogens, 30 of the 44 or 45 residues are identical, and most of the substi- tutions in the other residues are conservative. The preser- vation of positive charges in the NH,-terminal portion of the zymogen molecules, lost during activation, agrees with current ideas on the involvement of this segment in stabilizing the physical structure of the zymogens.
Vertebrate enzymes involved in the digestion of diet)ary pro- teins are synthesized in precursor form and are secreted as such into the gastrointestinal tract. Recent studies (reviewed in (1)) have shown that these precursor molecules, or zymogens, have inherent activity by which those that undergo autoactiva- tion can convert themselves into the more active form of the respective enzymes. In the case of the zymogens of the gastric proteases, this activity is developed by a reversible change into the active conformation at low pH (2-4).
The irreversible transformation into active enzymes takes place by limited proteolysis during which a peptide segment is
* The part of the work carried out in Denmark was supported by a grant from the Danish Natural Science Research Council. The part of the work conducted in the United States was supported by National Institutes of Health Grant AM-09826 from the Na- tional Institute of Arthritis, Metabolism, and Digestive Diseases and by National Science Foundation Grant GB-12630.
$ Present address, Department of Biochemistry, University College, Cardiff, Great Britain.
cleaved from the NH2 terminus in each case (cf. Ref. 5). The primary structures of the NH*-terminal portions of the zymogens are of considerable interest in order to understand, first, how the zymogens are able to maintain their less active conformation, and second, how the irreversible activation occurs under the proper conditions.
In the conversion of porcine pepsinogen to pepsin,’ 44 amino acid residues are cleaved from the NH2 terminus of the molecule. Ong and Perlmann (6) determined the sequence of the first 41 residues, while Pedersen and Foltmann (7) and Stepanov el al. (8) found an additional sequence of 3 residues which connects the first 41 residues with the pepsin moiety of the zymogen.
Preliminary studies on the sequence of the activation peptides of bovine pepsinogen have been reported by Kassell et al. (9), Kay (lo), and Foltmann et al. (11). The major portion of the sequence, determined independently in both laboratories, was in agreement, although there were some errors in both sequences. The discrepancies have now been resolved by re-examination of the earlier results and by further experiments. The total se- quence is presented here.
A pepsin-inhibiting substance formed during activation of porcine pepsinogen has been known for many years (12). This inhibitor was partially characterized by Van Vunakis and Her- riott (13). A recent analysis by Anderson and Harthill (14) showed that it corresponds to the NHz-terminal 16 amino acids in the sequence of One; and Perlmann (6), except for 1 extra lysine residue. We report here the identification of an inhibitory pep- tide containing the first 17 amino acids of bovine pepsinogen.
MATERIALS AND METHODS
Bovine Pepsinogen-The pepsinogen was extracted from the mucosa of fourth-stomachs (abomasa), which had been stored frozen. Stomachs from both Danish and American cattle were used, so that two different preparations of pepsinogen were ob- tained, designated [D]% and [A]. For the [D] preparation, the
r Where nothing else is stated, the terms pepsinogen and pepsin refer to the bovine proteins.
2 The letters [D] and [A] are used to designate Danish and Amer- ican preparations, procedures, etc. Other abbreviations used are : TPCK-, n-1-tosvlamido-2-phenvlethyl chloromethgl ketone deriv- ative of; DNSI or dansyl-, 5Ldimethylaminonaphthalene-l-sul- fonyl-; PTH-, phenylthiohydantoin of; BAWP, 1-butanol-acetic acid-water-pyridine, 15:3:12:10; TLC, thin layer chromatography; -, Edman degradation with identification as the PTH derivative
4487
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4488
procedure of Chow and Kassell (15) was modified as previously described (16). For the [A] preparation, purified pepsinogen (15) was subjected to chromatography on polylysine-Sepharose 4B at pH 6.5 (17) as the final step of purification (10). Pepsinogen [A] was resolved into three potentially active peaks differing only in their content of covalently bound phosphate (0.4, 1.5, and 2.8 moles per mole of protein). During activation of bovine pepsino- gen, all the phosphate groups remain attached to the pepsin moiety (18), indicating that no phosphate is present in the activa- tion peptides. The three components were therefore pooled and treated as one. Pepsinogens [D] and [A] were identical in com- position within the limits of analysis.
Other Enzymes and Inhibitors-TPCK-trypsin was obtained from Worthington Biochemicals, Inc., Freehold, N. J. Carboxy- peptidase A and B were either Worthington or Sigma preparations (Sigma Chemical Co.. St. Louis. MO.). Portions of the carboxv- peptidase A suspension were prepared for use as described by Ambler (19). Pancreatic trypsin inhibitor (Trasylol@) was pur- chased from Bayer, Germany.
Reagents-Reagents for manual sequencing were redistilled ac- cording to Edman and Begg (20) and were stored under Ns at -15”. The reagents for the Sequencer, phenylisothiocyanate, Quadrol buffer, heptafluorobutyric acid, heptane, benzene, ethyl acetate, and 1-chlorobutane were all Sequencer Grade from Beck- man Instruments. 1,4-Butanedithiol (obtained from Fluka) was added (0.005%;) to the chlorobutane in order to obtain higher yields of serink: Other reagents were the best grade available.
Chromatography and Ele&ophoresis-Sephadex and Sepharose were obtained from Pharmacia. DEAE-cellulose was either from Whatman Biochemicals, Ltd., Maidstone, U. K., or from Schlei- cher and Schuell, Keene, N. H. The anion exchanger AG l-X2 was nurchased from Bio-Rad. Richmond. Calif. Polvlvsine (mol wt 5?),000 to 100,000) was purchased from Pierce Chemical eo.,
Rockford, 111. Polylysine-Sepharose 4B was prepared in two batches of 75 g each for a column (2.5 X 45 cm) by the method of Nevaldine and Kassell (17). Whatman chromatography papers were used for electrophoresis and chromatography. Polyamide sheets were obtained from Cheng Chin Trading Co., Taipei, Tai- wan.
High voltage paper electrophoresis at about 50 volts per cm was conducted [I>] in liquid-cooled tanks (coolants and buffers as described by Ambler (21)) or [A] with the use of a flat-plate ap- paratus (Savant Instruments, Inc., Hicksville, N. Y.). Either a visible marker containing methyl green and crystal violet was used (IA1 (22)). or l>NS-OH and DNS-arainine were used as internal markkrs’iD]. In the latter case, mtbilities at pH 6.5 and pH 2 were calculated relative to DNS-OH (-1.0) and DNS-arginine (+l.O), respectively. Peptides were partially purified and de- salted by passage through Sephadex.
Pepsin Assays-Hemoglobin and milk-clotting assays were carried out as described previously ([D] (23), [A] (15)).
Amino Acid Analysis-Protein samples were hydrolyzed in re- distilled, constant boiling HCl in sealed, evacuated tubes at 110” for 24, 48, or. 72 hours. -Peptides were’hydrolyzed for 16 to 24 hours. Analvses were run on a Beckman 120B IAl. a BioCal 201 . -, [D], or a Durrum 500 [D] amino acid analyzer.
Sequencing-The sequence of the first 20 residues of intact pep- sinogen [D] was established by Edman degradation (20) in a Beck- man Sequencer (model 890B). The liberated thiazolinones were converted to PTH derivatives, which were identified (both silylated and unsilylated) by gas-liquid chromatography and also on the amino acid analyzer after hydrolysis to free amino acids with HI (24), taking into account the formation of known artifacts during HI hydrolysis (24). Direct identification by gas chroma- tography gave only very faint indications of PTH-glutamic and aspartic acids and their amides, but after conversion to the free amino acids, the corresponding dicarboxylic acids were easily identified. Arginine was identified only as the free amino acid.
Peptides were sequenced by manual Edman degradation ([Al (25), [D] and [A] (26)). The DNS-amino acids were identified on standard size polyamide sheets ([D] and [A] (27, 28)) or by a micromethod ([D] (29)) using polyamide sheets (5 X 5 cm), which permitted the identification of 0.2 nmole of DNS-amino acid.
or as the free amino acid after conversion; -, Edman degradation with identification as the dansyl amino acid; -, identification by carboxypeptidase A or B.
Usually for the tryptic peptides and sometimes for the activation peptides, the thiazolinones or phenylthiohydantoinswere converted to free amino acids ([D] (24), [A] (30)) for identification on the analyzer. The repetitive yield during manual Edman degradation is about 90% [D] except for the acidic and basic amino acids, which give somewhat less. The recovery of free amino acids after con- version of the PTH derivatives with HI is 20 to 60% for histidine and arginine, but 60 to 90% for the other amino acids, including the artifacts produced during the conversion. In addition, some residues were identified by thin layer chromatography of the PTH derivatives, with the use of the solvent systems (31) cited below. The last residue of a sequence was identified either as the dansyl derivative by TLC or as the free amino acid without hydrolysis.
Maleylation and Tryptic Digestion of Pepsinogen [D]-The con- ditions of the experiment have been described previously (16). Briefly, denatured pepsinogen was maleylated in 5 M guanidine HCl. After dialysis and lyophilization, 140 mg of the modified zymogen in 10 ml of water was digested with 2 mg of trypsin for 10 min at lo”, the pH being kept at 7.8. The reaction was termi- nated by the addition of 0.5 ml of Trasylol solution.
Maleylation of Peptides [Al-Modification of lysine residues was carried out at pH 9 (32), the maleic anhydride being added in dioxane. The excess of reagent was removed by passage through a Sephadex G-15 column, equilibrated with 0.1 N NHIOH, followed by lyophilization of the peptide fractions. Deprotection was achieved by dissolving the material in 5% acetic acid-170 pyridine and heating at 60” for 1 hour.
Designation of Peplides and Numbering of Amino Acid Residues -Peptides are numbered according to their finally determined positions in the peptide chain. TM means that the peptide is from a tryptic digest of a maleylated preparation. T stands for “true” tryptic peptides. Activation peptides are marked Act.
RESULTS
Automatic Edman Degradation of Intact Pepsinogen In Beckman
Sequencer (Residues l-20) Sir-Val-Val-L~s-Ile-Pro-Leu-V~l- ~-~4----L-~--L~~
Lys-Lys-Lys-Ser-Leu-Arg-Gln-Asn-Leu-Ile-Gln-Asn
The average repetitive yield during the operation was approxi- mately 90%. As far as was possible, all the residues were iden- tified both as the PTH derivatives and as the free amino acids after conversion with HI. However, arginine-14, glutamine-15, asparagine-16, glutamic acid-19, and asparagine-20 were iden- tified with certainty only after conversion to the free amino acids. Amide assignment was made by comparison to the cor- responding activation peptides, although the amides were seen in low yields as the PTH derivatives also. After 20 cycles, the background due to unspecific cleavage reached a level prohibitive of further identification.
Activation of Bovine Pepsinogen
To avoid degradation of activation peptides by the newly formed pepsin, optimum conditions of activation were first de- termined. Fig. 1 shows the course of activation at 0” between pH 1.5 and 3.0; the optimum pH for activation is 2.0.
Activation experiments were carried out in both laboratories under the same conditions except for the time of activation and the method of separation of the peptides. In the first experi-
ment, 370 mg of pepsinogen [A] was activated at pH 2 and 0” for 4 min at a protein concentration of 15 mg per ml. The pH was raised to 3.5, and the solution was immediately applied to the column of polylysine-Sepharosea at pH 3.5 (Fig. 2). The pep-
3 Separation on polylysine-Sepharose depends on both affinity chromatography and ion exchange (17). With pepsinogen at pH 6.5, ion exchange appears to predominate, resulting in the separa- tion of 3 peaks differing in bound phosphate. At pH 3.5, the pep- sin appeared in a single peak, indicating the predominance of the affinity effect.
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

I Wg7
pH1.5
%I
1ooF F--
~~2.5 I / pH 3.0
5Okb'
'* p 20 40 60
/ / Mlnutel; , -..
1 2 3 4 5
Exposure To Low pH, Min
FIG. 1. The rate of activation of bovine pepsinogen at 0” under varying conditions of pH. A solution of 2 mg of pepsinogen in 3 ml of water, cooled to 0”, was rapidly adjusted to the desired pH with a predetermined amount of 2.5 N HCl. At the designated times, O.l-ml samples were added to 0.4 ml of 0.1 M Tris-acetate buffer, pH 6.5, also at 0”. The final pH was 5.5. The amount of active pepsin was determined by the milk-clotting method.
4489
TABLE I Amino acid composition of mixture of peptides liberated by activation
of bovine pepsinogen
Amino acid
Lysine. Histidine Arginine. Aspartic acid. Asparagine. Threonine. Serine. Glutamic acid. Glutamine.. Proline Glycine Alanine.... Cystine. . . Valine.. Methionine Isoleucine Leucine . Tyrosine. Phenylalanine Tryptophand.. . .
,
--
-
8 7.4 6.8 1 0.8 0.9 3 2.3 3.0 4 4.0 3.6
2 2.2 2.2 6 3.1 3.0 4 4.6 4.5
0 1.4 1.3 2 2.4 2.6 2 2.0 2.6 0 0 0 2 2.3 3.0 1 0.9 1.5 4 3.0 3.2 6 5.2 5.9 2 1.7 1.8 1 1.3 1.2 0 ND8 0.04’
P -h4-b~-%31r_r4_r4_.Al/-“ti- n Pepsinogen minus pepsin (15,33).
I=“” b 24-hour hydrolysate.
8 1 3
3 2 3 1 3 1 2 2 0 3 1 3 6 2 1 0
c Calculated values from 24-, 48-, and 72-hour hydrolysates. d Analyses following hydrolysis with p-toluenesulfonic acid con-
taining 3(2-aminoethyl) indole (34) indicated that there are 5
‘a 1 ~z%,,‘++,~-,~ residues of tryptophan in both the pepsinogen and the pepsin.’ I. This corrects the previous value of 6 tryptophan residues in
pepsinogen, determined spectrophotometrically (15). e N D = not determined.
1 Determined spectrophotometrically (35).
FIG. 2 (left). Chromatography of pepsinogen activation mixture [A] on polylysine-Sepharose 4B (2.5 X 45 cm) equilibrated with 0.05 M ammonium formate buffer, pH 3.5. A linear gradient of 900 ml of formate buffer and 900 ml of 1 M NaCl in the buffer was ap- plied after 380 ml. Fractions (10 ml) were collected at a flow rate of 150 ml per hour. Where the enzyme was expected, the collec- tion tubes contained 0.6 ml of 4 M sodium acetate buffer, pH 5.6, to prevent autolysis. O---O, ASO; O- - -0, ASTO (ninhydrin).
FIG. 3 (right). The sequences of the activation peptides and of
The amino acid compositions of the peptide mixtures from the two activations were nearly identical (Table I) and were in reasonable agreement with the composition estimated from the difference in composition between pepsinogen (15) and pepsin (33). The composition determined by the sequence studies below is included in Table I.
the peptides from the tryptic digest of maleylated pepsinogen. -, sequential Edman degradation-dansylation; -, sequential Edman degradation with identification as PTH derivative or after conversion to free amino acid; -, carboxypeptidase A or B (or both) ; --, present from amino acid analysis.
tides were eluted with 0.05 M ammonium formate buffer, pH 3.5 (Peak I), and the pepsin was eluted by a salt gradient (Peak II). In the second experiment, 225 mg of pepsinogen [D] was acti- vated for 8 min. The pH was raised to 5.3, and the peptides were separated from pepsin by chromatography on DEAE-cel- lulose in sodium phosphate buffer, pH 5.3, with a salt gradient (16). The elution pattern (not shown) was quite similar to Fig. 2, the activation peptides being in Peak I and the pepsin in Peak II.
Composition and Sequence of Activation Peptides
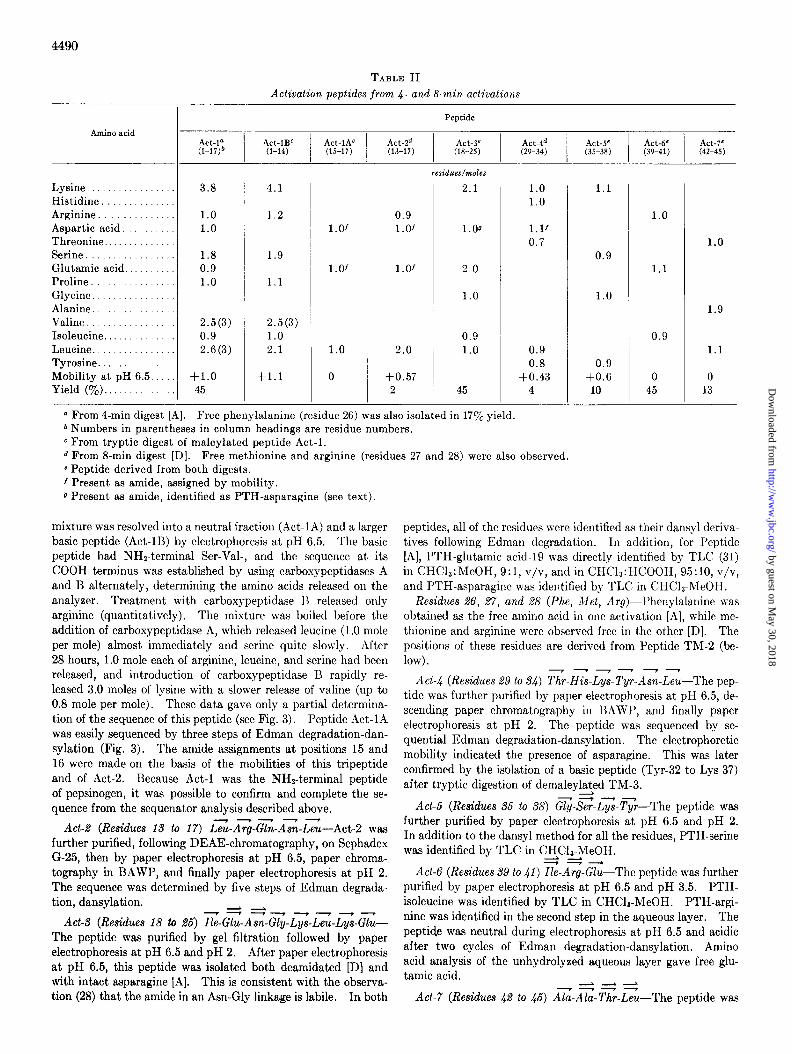
The two peptide mixtures were fractionated by chromatog- raphy on Bio-Gel P-2 (2 x 90 cm [A]) or on Sephadex G-25 (1.5 x 90 cm [D]). Each fraction was further purified by high voltage paper electrophoresis and occasionally by chromatog- raphy in BAWP (36). Details of the composition, electropho- retie mobility, and yield of each activation peptide are given in Table II. The sequence, determined as described in this sec- tion, is summarized in Fig. 3.
Act-l (Residues 1 to 17) ~-Vai-(Vccl,Lys,Ile,Pro,Leu)-Pal-
LTs-LG-LG-&-LL-A$-kn-A;-L%-Act-1 was purified fur- ther by paper electrophoresis at pH 6.5. Edman degradation gave Ser-Val-, corresponding to the NH2 terminus of pepsinogen. A portion of the peptide was maleylated (32) and digested with TPCK-trypsin (peptide concentration 0.75 rnrd, 4 hours, 37”, sub- strate to enzyme ratio 85: 1). After deblocking, the peptide
4 B. Kassell, unpublished.
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4490
TABLE II Activation peptides from 4- and 8-min activations
Peptide
Act-la (l-17)b
Act-lBC Act-lAC
I I
Act-2d Act-3e (l-14) (15-17) (13-17) (18-25)
residues/moles
Amino acid
Lysine................. Histidine Arginine Aspartic acid. Threonine. Serine Glutamic acid Proline. Glycine Alanine Valine Isoleucine. Leucine. ............... Tyrosine. ............ Mobility at pH 6.5. .... Yield (‘$&). ..........
3.8
1.0 1.0
4.1
1.2
1.8 0.9 1.0
1.9
1.1
2.5(3) 2.5(3)
0.9 1.0 2.6 (3) 2.1
+1.0 45
+1.1
l.Of
l.Of
1.0
0
0.9 l.Of
1.01
2.0
+0.57 2
2.1
1.00
2.0
1.0
0.9 1.0
45
0.9 0.8
+0.43 4
-
0.9
1.0
0.9
+0.6 10
1.0
1.1
0.9
0 45
1.0
1.9
1.1
0 13
a From 4-min digest [A]. Free phenylalanine (residue 26) was also isolated in 17yo yield * Numbers in parentheses in column headings are residue numbers. c From tryptic digest of maleylated peptide Act-l. d From 8-min digest [D]. Free methionine and arginine (residues 27 and 28) were also observed. c Peptide derived from both digests. 1 Present as amide, assigned by mobility. g Present as amide, identified as PTH-asparagine (see text).
mixture was resolved into a neutral fraction (Act-1A) and a larger basic peptide (Act-1B) by electrophoresis at pH 6.5. The basic peptide had NHz-terminal Ser-Val-, and the sequence at its COOH terminus was established by using carboxypeptidases A and B alternately, determining the amino acids released on the analyzer. Treatment with carboxypeptidase 1~ released only arginine (quantitatively). The mixture was boiled before the addition of carboxypeptidase A, which released leucine (1 .O mole per mole) almost immediately and serine quite slowly. After 28 hours, 1.0 mole each of arginine, leucine, and serine had been released, and introduction of carboxypeptidase B rapidly re- leased 3.0 moles of lysine with a slower release of valine (up to 0.8 mole per mole). These data gave only a partial determina- tion of the sequence of this peptide (see Fig. 3). Peptide Act-1A was easily sequenced by three steps of Edman degradation-dan- sylation (Fig. 3). The amide assignments at positions 15 and 16 were made on the basis of the mobilities of this tripeptide and of Act-2. Because Act-l was the NHz-terminal peptide of pepsinogen, it was possible to confirm and complete the se- quence from the sequenator analysis described above.
Act-2 (Residues 13 to 17) Leu-ATg-Gin-AT%-LG---Act-2 was further purified, following DEAE-chromatography, on Sephadex G-25, then by paper electrophoresis at pH 6.5, paper chroma- tography in BAWP, and finally paper electrophoresis at pH 2. The sequence was determined by five steps of Edman degrada- tion, dansylation.
-7i--L-?-.-?-- Act-3 (Residues 18 lo 26) Ile-Glu-Asn-Gly-Lys-Leu-Lys-Glu-
The peptide was purified by gel filtration followed by paper electrophoresis at pH 6.5 and pH 2. After paper electrophoresis at pH 6.5, this peptide was isolated both deamidated [D] and with intact asparagine [A]. This is consistent with the observa- tion (28) that the amide in an Asn-Gly linkage is labile. In both
peptides, all of the residues were identified as their dansyl deriva- tives following Edman degradation. 111 addition, for Peptide [A], PTH-glutamic acid-19 was directly identified by TLC (31) in CHC13:MeOH, 9:1, v/v, and in CHC13:HCOOH, 95:10, v/v, and PTH-asparagine was identified by TLC in CHCls-MeOH.
Residues 26, 27, and 28 (Phe, Jlet, Arg)-Phenylalanine was obtained as the free amino acid in one activation [A], while me- thionine and arginine were observed free in the other [D]. The positions of these residues are derived from Peptide TM-2 (be- low).
7-y--- Act-4 (Residues 29 to 34) Thr-His-Lys-Tyr-Am-Leu-The pep-
tide was further purified by paper electrophoresis at pH 6.5, de- scending paper chromatography in BAWP, and finally paper electrophoresis at pH 2. The peptide was sequenced by se- quential Edman degradation-dansylation. The electrophoretic mobility indicated the presence of asparagine. This was later confirmed by the isolation of a basic peptide (Tyr-32 to Lys-37) after tryptic digestion of demaleylated TM-3.
Act-6 (Residues 36 to 38) CT-S?-Lyd-T$--The peptide was further purified by paper electrophoresis at pH 6.5 and pH 2. In addition to the dansyl method for all the residues, PTH-serine was identified by TLC in CX+C$-I+MeO,H.
Act-6 (Residues 39 to 41) Ile-Arg-Glu-The peptide was further purified by paper electrophoresis at pH 6.5 and pH 3.5. PTH- isoleucine was identified by TLC in CHCls-MeOH. PTH-argi- nine was identified in the second step in the aqueous layer. The peptide was neutral during electrophoresis at pH 6.5 and acidic after two cycles of Edman degradation-dansylation. Amino acid analysis of the unhydrolyzed aqueous layer gave free glu- tamic acid. --L----L --7-.-P
Act-7 (Residues 42 to 46) Ala-Ala-Thr-Leu-The peptide was
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4491
EFFLUENT, ml
FIG. 4. Chromatography of the tryptic digest of maleylated pepsinogen [D] on Sephadex G-100 (2.5 X 100 cm) equilibrated in 0.05 M ammonium bicarbonate buffer, pH 8.0. Fractions (5 ml) were collected at a flow rate of 25 ml per hour. --, Azao; - - -, A255.
further purified by paper electrophoresis at pH 6.5 and 3.5. In addition to identification of each residue as the dansyl derivative during Edman degradation, at the second cycle alanine was directly identified as the PTH derivative by TLC in CHCla- MeOH. At this stage, a portion of the residue was hydrolyzed and yielded only threonine to leucine, 1: 1 on the analyzer. After the third cycle, the analysis of the unhydrolyzed residue gave only leucine.
Tryptic Peptides from Maleyl Pepsinogen
Pepsinogen contains 6 arginine residues, three in the peptides released upon activation and three in the pepsin moiety of the molecule. Of the latter, two are located within 20 residues of the COOH terminus (37), and the third is 55 residues from the NH2 terminus of the enzyme.s Thus, tryptic digestion of maleyl- ated pepsinogen should produce one large fragment, one medium- sized fragment, and five small pieces (three from the NH2 termi- nus and two from the COOH terminus).
The digest (see under “Materials and Methods”) was frac- tionated on Sephadex G-100 (Fig. 4, taken from (16)). Peaks I, ZZ, and ZZZ were described previously (16). Peak Z was ah aggregation product; Peak ZZ contained the expected large frag- ment from the pepsin moiety. Peak ZZZ was almost pure Pep- tide TX-IV, the overlapping sequence between the activation peptides and pepsin (16) ; part of its sequence is included in Fig. 3.
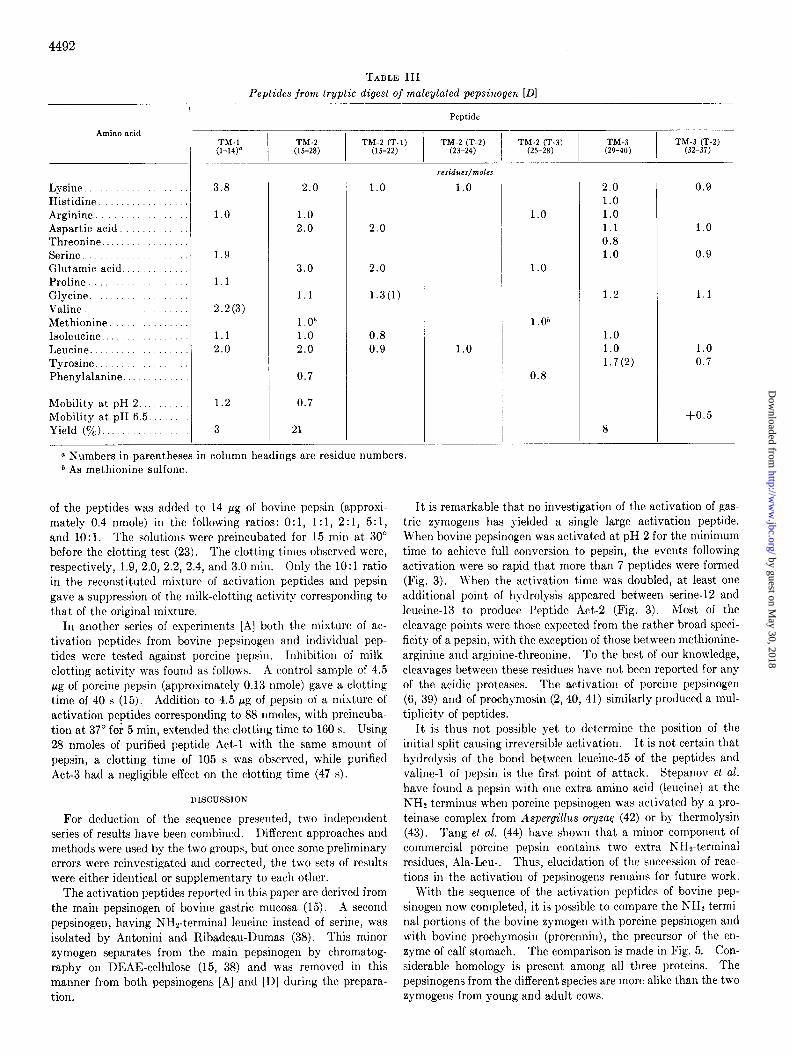
Peaks IV and V are of present interest. The detailed descrip- tions of the peptided isolated from these two components are given below and are summarized in Table III and Fig. 3. In
addition to the peptides to be described, 2 peptides from the COOH-terminal portion of the pepsinogen were also isolated. The sequences of these have been published (37).
--d--L--- TX1 (Residues 1 to 14) Ser-Val-Val-Lys-Zle-Pro-Leu-Val-Lys-
Lys-Lys-Ser.Leu-Arg-After gel filtration on Sephadex G-100 (Fig. 4), the peptides in Peak IV were demaleylated; TM-l was purified by paper electrophoresis at pH 2 (mobility 1.2 relative to DNS-arginine) and was sequenced unambiguously by Edman degradation and identification of the free amino acids after con- version with HI.
The low yield (Table III) resulted from unsuccessful attempts to separate TM1 and TM-2 on DEAE-cellulose and paper elec- trophoresis without demaleylation before it was found that the deblocked peptides separated well at pH 2.
TJZ-2 (Residues 15 to 28) (Gln,Asn,Leu,Zle,Glu,Asn,Gly,
6 M. Harboe and B. Foltmann, unpublished observations.
Lys, Leu, Lys, Glu)-%-&t-Ayg-This peptide was isolated from Peak IV in the same operations as TM-1 (electrophoretic mobil- ity 0.7 at pH 2 relative to DNS-arginine). After elution and drying, the NHP-terminal glutamine was apparent,ly blocked by cyclization to pyrrolidone carboxylic acid, since it was not SUS-
ceptible to Edman degradation. By digestion with a mixture of carboxypeptidase A and B, phenylalanine, methionine, and argi- nine were liberated in the ratios 0.5:0.85: 1.0. After performic acid oxidation (to protect methionine by conversion to the sul-
fone), the peptide was digested with trypsin at pH 8 and room temperature overnight at an enzyme to substrate ratio of 1:75. Electrophoresis at pH 3.5 resolved 3 peptides.
TM-2 (T-l) (Residues 15 to 22) (Gln,Asn, Leu,Zle,Glu,Asn, Gly,Lys)-This peptide had a blocked NH2 terminus, but its amino acid composition (Table III) corresponded well to the NHz-terminal part of TM-2.
TM-2 (T-2) (Residues 23 to 2.4) Lz-L<--Leucine was the NH2 terminus. After one Edman degradation, only lysine was left.
Td4-2 (T-S) (Residues 25 to 28) &-Pz-G-A;-The se- quence was determined by Edman degradation and conversion with HI to free amino acids. Methionine was determined as the sulfone. This peptide represents the COOH-terminal part of TM-2.
----L--L -TM13 @esizues 29 to 40) T<r-Hz-LTs-TTr-Asn-Leu-Gly-Ser-
Lys-Tyr-Zle-Arg-This peptide was purified from Peak V of Fig. 4 by gel filtration on Sephadex G-25, followed by paper elec- trophoresis at pH 3.5. The sequence was obtained by Edman degradation and identification of each amino acid after conver- sion with HI. Digestion with trypsin was conducted as de- scribed for TM-2. The digest yielded 3 basic peptides, resolved by paper electrophoresis at pH 6.5.
TM-3 (T-2) (Residues 32 to 37) TT-A%-L%-Gz-S?-L<-- TM-3 (T-2), the peptide of lowest mobility (+0.5), was further purified by paper electrophoresis at pH 2. Edman degradation- dansylation established the sequence. The presence of aspara- gine was determined by the basicity of the peptide.
TJI-C-This was a large fragment that included the NHz- terminal part of pepsin. The sequence of the first 12 residues has been published (16).
Deduction of Structures and Order of Activation Peptides
The results, summarized in Fig. 3, show that every residue has been sequenced, although not in every peptide. The activation peptides plus the tryptic peptides allow an unequivocal sequence to be deduced. Overlaps are complete except for the bond be- tween residues 28 and 29, but this missing overlap does not cause any problem. The confirmation of the NHz-terminal portion of the sequence by the sequenator and of the COOH-terminal por- tion by the continuation of peptide TM-4 into pepsin (16) makes this the only possible arrangement.
Pepsin-inhibiting Activity of Activation Peptides
When pepsinogen was activated, the milk-clotting activity of the reaction product almost doubled after the activation pep- tides were separated by gel filtration [D] or by chromatography on DEAE-cellulose [D] or polylysine-Sepharose [A].
Preliminary experiments have been carried out to investigate the inhibiting properties of the activation peptides. In one series of experiments [D] the activation peptides were first sepa- rated from pepsin by chromatography. A concentrated mixture
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4492
Amino acid
Lysine ....... ..... Histidine ............. Arginine ............ Aspartic acid. ........ Threonine. ....... ...... Serine ........ Glutamic acid ... Proline. ... Glycine. .. Valine ....... Methionine ...... Isoleucine. ......... Leucine. ............. Tyrosine. .............. Phenylalanine. .....
Mobility at pH 2 ...... Mobility at pH 6.5 ... Yield (%). ........
-
-
-
TABLE III Peptides from tryptic digest of maleylated pepsixogen [Dl
Peptide
TM-l I
TM-2 (l-14)” (1.5-28)
TW;- 1) ‘“;‘;3’_2’$2’
3.8
1.0
1.9
1.1
2.2(3)
1.1 2.0
1.2
3
2.0
1.0 2.0
3.0
1.1
l.O* 1.0 2.0
0.7
0.7
21
1.0
2.0
2.0
1.3(l)
0.8 0.9
residues/moles
1.0
1.0
1.0
1.0
1.0*
0.8
2.0 1.0 1.0 1.1 0.8 1.0
1.2
1.0 1.0 1.7(2)
8
0.9
1.0
0.9
1.1
1.0 0.7
+0.5
a Numbers in parentheses in column headings are residue numbers. * As methionine sulfone.
of the peptides was added to 14 pg of bovine pepsin (approxi- mately 0.4 nmole) in the following ratios: O:l, 1 :I, 2:1, 5:1, and 10: 1. The solutions were preincubated for 15 min at 30” before the clotting test (23). The clotting times observed were, respectively, 1.9, 2.0, 2.2, 2.4, and 3.0 min. Only the 1O:l ratio in the reconstituted mixture of activation peptides and pepsin gave a suppression of the milk-clotting activity corresponding to that of the original mixture.
In another series of experiments [A] both the mixture of ac- tivation peptides from bovine pepsinogen and individual pep- tides were tested against porcine pepsin. Inhibition of milk- clotting activity was found as follows. A control sample of 4.5 pg of porcine pepsin (approximately 0.13 nmole) gave a clotting time of 40 s (15). Addition to 4.5 pg of pepsin of a mixture of activation peptides corresponding to 88 nmoles, with preincuba- tion at 37” for 5 min, extended the clotting time to 160 s. Using 28 nmoles of purified peptide Act-l with the same amount of pepsin, a clotting time of 105 s was observed, while purified Act-3 had a negligible effect on the clotting time (47 s).
It is remarkable that no investigation of the activation of gas- tric zymogens has yielded a single large activation peptide. When bovine pepsinogen was activat,ed at pH 2 for the minimum time to achieve full conversion to pepsin, the events following activation were so rapid that more than 7 peptides were formed (Fig. 3). When the activation time was doubled, at least one additional point of hydrolysis appeared between serine-12 and leucine-13 to produce l’eptide Act-2 (Fig. 3). Most of the cleavage points were those expected from the rather broad speci- ficity of a pepsin, with the exception of those between methionine- arginine and arginine-threonine. To the best of our knowledge, cleavages between these residues have not been reported for any of the acidic proteases. The activation of porcine pepsinogen (6, 39) and of prochymosin (2,40, 41) similarly produced a mul- tiplicity of peptides.
DISCUSSION
For deduction of the sequence presented, two independent series of results have been combined. Different approaches and methods were used by the two groups, but once some preliminary errors were reinvestigated and corrected, the two sets of results were either identical or supplementary to each other.
It is thus not possible yet to determine the position of the initial split causing irreversible activation. It is not certain that hydrolysis of the bond between leucine-45 of the peptides and vaiine-1 of pepsin is the first point of attack. Stepanov et al.
have found a pepsin with one extra amino acid (leucine) at the NH2 terminus when porcine pepsinogen was activated by a pro- teinase complex from Aspergillus oryzae (42) or by thermolysin (43). Tang et al. (44) have shown that a minor component of commercial porcine pepsin contains two extra NHt-terminal residues, Ala-Leu-. Thus, elucidation of the succession of reac- tions in the activation of pepsinogens remains for future work.
The activation peptides reported in this paper are derived from With the sequence of the activation peptides of bovine pep- the main pepsinogen of bovine gastric mucosa (15). A second sinogen now completed, it is possible to compare the NHs-termi- pepsinogen, having NHz-terminal leucine instead of serine, was nal portions of the bovine zymogen with porcine pepsinogen and isolated by Antonini and Ribadeau-Dumas (38). This minor with bovine prochymosin (prorennin), the precursor of the et)- zymogen separates from the main pepsinogen by chromatog- zyme of calf stomach. The comparison is made in Fig. 5. Con- raphy on DEAE-cellulose (15, 38) and xas removed in this siderable homology is present among all three proteins. The manner from both pepsinogens [A] and [D] during the prepara- pepsinogens from the different species are more alike than the two tion. zymogens from young and adult cows.
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Pch: Ala-Glu-lle-Thr-Arg-ILE-PRO-LEU-Tyr-LYS-Gly-LYS-SER-LEU-ARG-Lys-Ala-LEU-LYS-GLU-His-GLY-Leu-LEU-Glu-
Pch: ASP-PHE-LEU-LYS-
FIQ 5. Comparison of the NH?-terminal portions of bovine pepsinogen (Bpg), porcine pepsinogen (Ppg), and prochymosin (proren- nin, Pch). Amino acids in homologous positions in any two of the peptide chains are written in capitals. Homologous sequences of the two pepsinogens are in boxes.
For the two pepsinogens, 30 of the 44 or 45 residues are identi- cal. Most of the substitutions are conservative, e.g. one basic amino acid for another or one hydrophobic residue for another. Other substitutions, glutamic acid for lysine in position 19 and proline for either leucine or arginine in positions 34 and 40, re- spectively, may result from single base changes. In all other cases the positive charges are conserved. This substantiates the idea previously proposed (2, 45-47) that the positive charges in the NHz-terminal portions of the zymogens are necessary to maintain a symogen conformation different from that of t,he active enzyme.
The striking similarity in structure in positions 1 to 17, cor- responding to pepsin-inhibiting peptides identified for both species, may indicate an important physiological role for these inhibitors. The inhibiting activity of the bovine activation pep- tides against porcine pepsin complements the action of porcine inhibitor against bovine pepsin reported by Herriott (12). These cross-reactions are not surprising in view of the homology in primary structure.
Several interesting aspects of inhibition by the activation pep- tides require additional investigation. At the present time, we are not able to explain why the inhibiting effect is greater in the original activation mixture than in a reconstituted mixture; a tentative suggestion is that a minor conformational change oc- curs in bovine pepsin once the activation peptides are removed. We have riot had material available to test other peptides in the bovine sequence; for kxample, the portion that corresponds to porcine sequence 27 to 38, a peptide reported by Wang and Edel- man (48) to have inhibiting activity. Further work is in prog- ress to determine the minimal requirements for an inhibiting peptide and the relationship of the peptides to the conformation of the pepsins.
Acknourledgmenls-One of us (M. H.) is grateful to the Royal Danish School of Pharmacy for a Fellowship. We thank Dr. J. Thomsen of the Danish Institute of Protein Chemistry, Copen- hagen, for the analysis on the Beckman Sequencer, Dr. Patricia A. Meitner, formerly of The Medical College of Wisconsin, for the experiments on the rate of activation of pepsinogen, and Mrs. Meredith Marciniszyn for preparing pepsinogen [A].
REFERENCES
1. KASSELL, B., AND Kay, J. (1973) Science 180, 1022-1027 2. FOLTMANN. B. (1966) C. R. Trav. Lab. Carlsbera 36. 143-231 3. FUNATSU, h., HARA~A, Y., HAYASHI, K., AND ~IR&NSONS, B.
(1971) Agr. Biol. Chem. 26, 566-572
4. 5. 6.
7.
8.
9.
10. 11.
12. 13.
14. 16.
16.
17.
18.
19. 20. 21. 22. 23. 24.
25.
26. 27.
28. 29.
30.
31.
32.
33.
34.
MCPHIE, P. (1972) J. Biol. Chem. 247.4277-4281 OTTESEN, M. (1967) Annu. Rev. Biochem. 36, 55-76 ONG, E. B., AND PERLMANN, G. E. (1968) J. Biol. Chem. 243,
6104-6109 PEDERSEN, V. B., AND FOLTMANN, B. (1973) Fed. Eur. Biochem.
Sot. Lett. 36, 256-256 STEPANOV, V. M., BARATOVA, L. A., PUGACHEV~~, I. B.,
BELYANOVA, L. P., REVINA, L. P., AND TIMOKHINA, E. A. (1973) Biochem. Biophys. Res. Commun. 64, 1164-1170
KASSELL, B., KAY, J.; AND MARCINISZYN, J. P., JR. (1972) in Third American Pevtide Sumvosium. Boston (MEIENHOFER. J., ed) pp. 713-716,‘Ann Aibdr Press; Ann A&or
KAY, J. (1972) Fed. Eur. Biochem. Sot. Proc. Meet., Abstr. 458 FOLTMANN, B., KAUFFMAN, D., PARL, M., AND ANDERSEN,
P. M. (1973) Neth. Milk Dairy J. 27, 288-297 HERRIOTT, R. M. (1941) J. Gen. Physiol. 24, 325-338 VAN VUNAKIS, H., AND HERRIOTT, R. M. (1956) Biochim.
Biophys. Acta 22, 637-543 ANDERSON, W., AND HARTHILL, J. E. (1973) Nature 243,417-419 CHOW, R. B., >\ND K.\SSELL, B. (1968) J. Biol. Chem. 243,
1718-1724 HARBOE, M., AND FOLTM~~NN, B. (1973) Fed. Eur. Biochem. Sot.
Lett. 94, 311-314 NEVALDINF:, B., .\ND KASSELL, B. (1971) Biochim. Biophys.
Acta 260, 207-209 MEITNER, P. A., AND KUSELL, B. (1971) Biochem. J. 121,
249-256 AMBLER, R. P. (1967) Methods Enzymol. 11, 436-445 EDMAN, P., AND BEGG, G. (1967) Eur. J. Biochem. 1, 80-91 AMBLER, R. P. (1963) Biochem. J. 89, 349-378 STEVENSON, K. J. (1971) Anal. Biochem. 40, 29-34 FOLTMANN, B. (1970) Methods Enzymol. 19, 421-436 SMITHIES, O., GIBSON, D., FANNING, E. M., GOODFLIESH,
R. M., GILMAN, J. G., AND BALLANTYNE, D. L. (1971) Bio- chemistry 10, 4912-4921
BLOMBBCK, B., BLOMB~CK, M., EDMAN, P., AND HESSEL, B. (1966) Biochim. Biophys. Acta 116, 371-396
GRAY, W. R. (1967) Methods Enzymol. 11, 469-475 WOODS, K. R., AND W.~NG, K.-T. (1967) Biochim. Biophys.
Acta 133, 369-370 HARTLEY, B. S. (1970) Biochem. J. 119, 805-822 BRUTON, C. J., AND HARTLEY, B. S. (1970) Biochem. J. 117,
18P-19P AFRICA, B., AND CARPENTER, F. H. (1966) Biochem. Biophys.
Res. Commun. 24, 113-119 BRENNER, M., NIEDERWIESER, A., AND PATAKI, G. (1969) in
Thin Layer Chromatography (STAHL, E., ed) pp. 73&786, Springer-Verlag, New York
BUTLER, P. J. G., HARRIS, J. I., HARTLEY, B. S., ,\ND LEBER- MAN, R. (1969) Biochem. J. 112, 679-689
LANG, H. M., AND KASSELL, B. (1971) Biochemistry 10, 2296- 2301
Lru, T.-Y., .~ND CHANG, Y.-H. (1971) J. Biol. Chem. 246, 2842-2848
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4494
35. BEAVEN, G. H., AND HOLIDAY, E. R. (1952) Advan. Protein Chem. 7, 319-387
36. WALEY, S. G., AND WATSON, J. (1953) Biochem. J. 66. 328-337 37. RASMUSSEN, K. T., AND FOLTMANN, B. (1971) Acta Chem.
&and. 26, 3873-3874 38. ANTONINI, J., AND RIBADEAU-DUMAS, B. (1971) Biochimie
(Paris) 63, 321-329 39. VAN VUNAKIS, H., AND HERRIOTT, R. M. (1957) Biochim. Bio-
phys. Acta 23, 6W608 40. FOLTMANN, B., AND ANDERSEN, B. (1971) Fed. Eur. Biochem.
Sot. Lett. 17, 87-89 41. FOLTMANN. B. (1971) Fed. Eur. Biochem. Sot. Proc. Meet.,
Abstr. 137. ’ 42. STEPANOV, V. M., TIMOKHINA, E. A., AND ZYAKUN, A. M.
(1969) Biochem. Biophys. Res. Commun. 37, 470-476
43. STEPANOV, V. M., TIMOKHINA, E. A., BARATOVA, L. A., BELYANOVA, L. P., KORZHENKO, V. P., AND ZHUKOVA, I. G. (1971) Biochem. Biophys. Res. Commun. 46, 1482-1487
44. TANG. J.. HUANG. W.-Y.. SEPULVEDA. P.. CHEN. K. C. S., MA&I&SZYN, J: P., JR:, TAO, N., AAD Jhu, D. ‘(1973) Fed: Proc. 32, Part I, 577
45. FOLTMANN, B. (1971) in Milk Proteins, Chemistry and Molecu- lar Biology (MCKENZIE, H. A., ed) pp. 217-254, Academic Press, New York --
46. WONG. R. C.. NAKAGAWA. Y.. AND PERLMANN. G. E. (1972) J. &ol. Chkm. 247, 1625-1631
. ,
47. FINLAY, T. H., DHARMGRONGARTAMA, E. D., AND PERLMANN, G. E. (1973) J. Biol. Chem. 248,48274833
48. WANG, J. L., AND EDELMAN, G. M. (1971) J. Biol. Chem. 246, 11861191
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Marianne Harboe, Per Maack Andersen, Bent Foltmann, John Kay and Beatrice KassellRELEASED, IDENTIFICATION OF A PEPSIN INHIBITOR
The Activation of Bovine Pepsinogen: SEQUENCE OF THE PEPTIDES
1974, 249:4487-4494.J. Biol. Chem.
http://www.jbc.org/content/249/14/4487Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/249/14/4487.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 30, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























