Embed Size (px)
Citation preview

T. gondii RP Promoters & Knockdown Reveal MolecularPathways Associated with Proliferation and Cell-CycleArrestSamuel L. Hutson1, Ernest Mui1, Karen Kinsley1, William H. Witola1¤a, Michael S. Behnke2¤b, Kamal El
Bissati1, Stephen P. Muench3, Brittany Rohrman1¤c, Susan R. Liu1, Robert Wollmann4, Yuko Ogata5, Ali
Sarkeshik6, John R. Yates III6, Rima McLeod1,7,8*
1 Department of Surgery (Ophthalmology), The University of Chicago, Chicago, Illinois, United States of America, 2 Department of Veterinary Molecular Biology, Montana
State University, Bozeman, Montana, United States of America, 3 Institute of Membrane and Systems Biology, University of Leeds, Leeds, United Kingdom, 4 Department
of Pathology, The University of Chicago, Chicago, Illinois, United States of America, 5 Seattle Biomedical Research Institute, Seattle, Washington, United States of America,
6 Department of Chemical Physiology, Scripps Research Institute, La Jolla, California, United States of America, 7 Department of Pediatrics (Infectious Disease), The
University of Chicago, Chicago, Illinois, United States of America, 8 Committees on Immunology, Molecular Medicine, and Genetics, Institute of Genomics and Systems
Biology, The College, The University of Chicago, Chicago, Illinois, United States of America
Abstract
Molecular pathways regulating rapid proliferation and persistence are fundamental for pathogens but are not elucidatedfully in Toxoplasma gondii. Promoters of T. gondii ribosomal proteins (RPs) were analyzed by EMSAs and ChIP. One RPpromoter domain, known to bind an Apetela 2, bound to nuclear extract proteins. Promoter domains appeared to associatewith histone acetyl transferases. To study effects of a RP gene’s regulation in T. gondii, mutant parasites (Drps13) wereengineered with integration of tetracycline repressor (TetR) response elements in a critical location in the rps13 promoterand transfection of a yellow fluorescent-tetracycline repressor (YFP-TetR). This permitted conditional knockdown of rps13expression in a tightly regulated manner. Drps13 parasites were studied in the presence (+ATc) or absence ofanhydrotetracycline (-ATc) in culture. -ATc, transcription of the rps13 gene and expression of RPS13 protein were markedlydiminished, with concomitant cessation of parasite replication. Study of Drps13 expressing Myc-tagged RPL22, -ATc, showedRPL22 diminished but at a slower rate. Quantitation of RNA showed diminution of 18S RNA. Depletion of RPS13 causedarrest of parasites in the G1 cell cycle phase, thereby stopping parasite proliferation. Transcriptional differences 6ATcimplicate molecules likely to function in regulation of these processes. In vitro, -ATc, Drps13 persists for months and theproliferation phenotype can be rescued with ATc. In vivo, however, Drps13 could only be rescued when ATc was givensimultaneously and not at any time after 1 week, even when L-NAME and ATc were administered. Immunization with Drps13parasites protects mice completely against subsequent challenge with wildtype clonal Type 1 parasites, and robustlyprotects mice against wildtype clonal Type 2 parasites. Our results demonstrate that G1 arrest by ribosomal proteindepletion is associated with persistence of T. gondii in a model system in vitro and immunization with Drps13 protects miceagainst subsequent challenge with wildtype parasites.
Citation: Hutson SL, Mui E, Kinsley K, Witola WH, Behnke MS, et al. (2010) T. gondii RP Promoters & Knockdown Reveal Molecular Pathways Associated withProliferation and Cell-Cycle Arrest. PLoS ONE 5(11): e14057. doi:10.1371/journal.pone.0014057
Editor: Gordon Langsley, INSERM U1016, Institut Cochin, France
Received June 3, 2010; Accepted October 25, 2010; Published November 22, 2010
Copyright: � 2010 Hutson et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: The authors gratefully acknowledge support of this work by gifts from the Fin Charity Trust, the Rooney Aldens, the Peter Mann and DominiqueCornwell Family Trust, the Morel, Jackson, Orlinsky, Goldberg, Taub, Kapnick, Schilling, and Kiewit families, Intervet/Schering Plough, Toxoplasmosis ResearchInstitute, The Research to Prevent Blindness Foundation, The Howard Hughes Foundation Undergraduate Summer Research Program in Neurosciences (BR), andNational Institutes of Health (NIH) National Institute of Allergy and Infectious Diseases R01AI043228, U01 AI082180-01, U01 AI 77887, R01 AI071319-01 (RM), NIHP41RR0118232 (JY). The funders had no role in study design, data collection and analysis, or decision to publish.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
¤a Current address: Department of Zoonotic Diseases and Veterinary Medicine, Tuskegee University, Tuskegee, Alabama, United States of America¤b Current address: Department of Molecular Microbiology, Washington University School of Medicine, St. Louis, Missouri, United States of America¤c Current address: Department of Bioengineering, Rice University, Houston, Texas, United States of America
Introduction
It was of interest to determine whether transcriptional
regulation of a ribosomal protein (RP) in Toxoplasma gondii could
play a role in control of this parasite’s cell cycle and consequent
phenotype in a model system in vitro. Recent studies of ribosomes
and/or cell cycle [1–18] in a number of organisms provided
precedent as they had demonstrated that RPs are critical for
regulation of cell cycle, differentiation, proliferation, and responses
to stresses [3,10,13–18]: For example, in Saccharomyces cerevisiae the
absence of RPs cause cell cycle arrest in G1 [13–16], slowing
growth, and causing differentiation; RP mutants of Arabidopsis [17]
and Drosophila melanogaster ‘‘minutes’’ [18], have smaller numbers of
ribosomes and/or severe growth impairment phenotypes. Previous
studies suggested stage-associated differences in expression of T.
gondii ribosomal proteins (RP) [19]. Furthermore, promoters of
PLoS ONE | www.plosone.org 1 November 2010 | Volume 5 | Issue 11 | e14057

genes encoding RPs in T. gondii contain highly conserved T. gondii
RP (TRPs) promoter sequences [20,21] suggesting they could
contribute to coordinated transcriptional regulation of RP
synthesis. These consensus sequences of TRPs, TRP1 (TCG-
GCTTATATTCGG) and TRP2 ([T/C]GCATGC[G/A]), are
located close to transcriptional start sites of RPs [20,21] and are
present frequently in the T. gondii genome. For example, a small
RP (rps13’s) promoter has two TRP2s, in addition to a motif found
to regulate bradyzoite gene expression, TGTGTG [22].
T. gondii also has orthologous molecules to those in other
organisms that interact with transcription factors regulating RPs.
Similar to other eukaryotes, activation of gene expression in
Toxoplasma involves acetylation of histone proteins [23,24] and
other components of an epigenetic regulatory complex [23–26]. In
the yeast, S. cerevisiae, MYST family histone acetyltransferase
(HAT), Esa1, is recruited to RP gene promoters to regulate
transcription [27]. Apetela 2 (AP2) transcription factors have been
identified in Plasmodia, T. gondii and plants [28–38], but are not
present in animals. In Plasmodium, a global yeast two hybrid
analysis suggested that ApiAP2 proteins interact with chromatin
remodeling factors and each other [39]. P. falciparum ApiAP2
protein PfSIP2 is key in chromosome end biology involving
heterochromatin formation and epigenetic factors [40]. In the
plant Arabidopsis, especially in stress conditions, a HAT interacts
with an AP2, CBFI [41]. Thus, it seemed feasible that a similar
AP2-HAT complex could play a role in regulation of transcription
of RPs in T. gondii. Consistent with this hypothesis, the TRP2
promoter element had been reported to contain bases that bind a
Plasmodium AP2 [29].
Little was known about which of T. gondii’s ribosomal
components are essential for ribosomes to function properly.
Thus, a new system using reagents described earlier [20,21] was
attractive for use to create a tightly regulated, conditional
knockdown of a RP. To attempt to create this tight, conditional
knockdown of a RP, a putative RP small subunit 13 gene (rps13)
had been selected earlier [20,21]. This RP was selected because it
had one major transcriptional start site [20,21]. It turned out to be
a fortuitous choice because recently, other organism’s RPS13s
have been found to be critical for catalysis and initiation of
translation. These RPS13s also are key in the association of small
and large ribosomal subunits, and the point at which tRNA,
mRNA and amino acids assemble to initiate protein synthesis in
other organisms [4–8]. We considered it possible that expression of
this RP gene selected earlier, if it truly were the homologue of
RPS13 in other organisms, would be essential for ribosome
assembly and function. Its absence would alter T. gondii
profoundly.
The use of a transactivator system had successfully created
conditional knockdowns of T. gondii genes [42]. A tetracycline
repressor (TetR) protein system [43] for T. gondii had been
described earlier although regulation of a functional endogenous
gene product was not accomplished [20,21]. The need for
tetracycline to enter encysted bradyzoites to turn a gene off was
a theoretical limitation of the current transactivator system.
Another challenge for the work presented herein, was to create,
validate, and characterize a tightly regulated, conditional,
‘‘tetracycline on’’ (called herein ‘‘+ATc’’) system in T. gondii that
would stably modify gene expression, independent of promoter
strength. In studies, foundational for the present work, tetracycline
repressor response element (TetO)-modified promoters for T.
gondii rps13 had been created [20,21] demonstrating an optimal
location for insertion of TetO elements by mapping the promoter
to optimize tight regulation. If this putative RPS13 functioned as
RPS13s in other species and was essential, it was hypothesized that
a RPS13 conditional knockdown parasite could be created and
characterized [20,21].
Herein, our studies provide new empirical information
concerning ribosomal protein (RPs) promoters and promoter
associated proteins. A role of a T. gondii ribosomal protein, (and
hence ribosomes) including its regulation and production, was
investigated using an inducible knockdown system [20,21].
Information concerning T. gondii proliferation and persistence in
an in vitro model system was obtained. Conditional knockdown of
rps13 created a G1 arrested parasite that persisted in this tissue
culture system for months. However, this mutant parasite did not
appear to persist in vivo and protected mice against challenge with
wildtype parasites.
Results
Modeling of the structure of the putative T. gondii RPS13demonstrates that it is likely part of the ribozyme
Because RPS13 was selected for conditional knockdown, and
because of its key location in ribosomes of other species (Figure 1A)
we wanted to determine more definitively that the putative RPS13
was likely to be a true RPS13 based on its structure. If it were a
true homologue of other species RPS13s it would be likely to be
essential for ribosome function. Thus, a structural model of T.
gondii RPS13 was produced using the phyre modeling server. This
model used the solved Thermophilus ribosomal protein RPS15
structure as a starting model. Analysis of the sequence alignment
between various RPS13 proteins and the Thermophilus RPS15,
protein from which the T. gondii model is based, shows a significant
amount of sequence similarity between not only the RPS13 family,
but the RPS13 and RPS15 proteins (Figure 1B), particularly at the
C-terminus. Furthermore, the RPS15 structure is in complex with
ribosomal RNA. Analysis of those residues that form the RNA
binding site show strong conservation between the RPS13 and
RPS15 family (blue circles in Figure 1B top panel). Moreover, the
sequence conservation between the RPS13 and RPS15 family is
located primarily around the RNA binding site, suggesting that the
mode of RNA binding in the RPS13 and RPS15 family is similar
(Figure 1B bottom panel; Movie S1). The critical location for the
homologous RPS13 in the catalytic site of ribosomes of other
organisms suggest that this T. gondii RPS13 is likely to be essential.
This finding made the creation and determination of the
phenotype of a conditional knockdown to characterize T. gondii
RPS13 function of interest. There were not obvious unique
features distinguishing the parasite and human RPS13s.
Analyses of proteins bound to RP promoter elementsreveal key regulatory pathways
TRP2 is present in two places in the rps13 gene’s promoter and
TRP1 is present in promoters of other rp genes [20,21]. This
made it of interest to characterize whether TRP1 and TRP2
bound nuclear extract proteins and if so which promoter
elements were critical for this binding. If they bound, further
characterizing nuclear extract proteins that associated with these
promoter elements also would be of interest. EMSAs using 32P
labeled TRP1 or TRP2 promoter elements show that tachyzoite
nuclear extracts cause mobility-shifts in TRP1 and TRP2, which
are competitively inhibited by cold probe (Figures 2A and S2A).
Nucleotide sequences within TRP2 elements of the RPS13
promoter which are critical for binding proteins were more
precisely identified. This was accomplished as follows: Three
base pair linker scan mutants (1 to 8 in Figure 2A) that spanned
TRP2 elements were synthesized individually and radiolabeled.
EMSAs were carried out using T. gondii nuclear extract with these
T. gondii rps13
PLoS ONE | www.plosone.org 2 November 2010 | Volume 5 | Issue 11 | e14057

radiolabeled wildtype or mutant TRP2 elements and binding
pattern observed. With mutant TRP2 probes, alteration of
nucleotide sequences TGCAT, a known AP2 binding motif [29]
shaded yellow in Figure 2A bottom, in mutants 4 to 8 (red font,
bottom panel, Figure 2A) abolishes or markedly reduces protein
binding. The highest, slow-migrating complex is most intense
(Figure 2A), when compared with other bound protein(s),
suggesting that the upper complex contains multiple proteins.
There was absence of binding with mutants 7 and 8, suggesting
that there are associated proteins in addition to those that are in
the complex with TGCAT.
We then studied whether T. gondii HATs MYST A and B
associated with TRP1 and/or TRP2, because Stockinger [41] had
found a HAT associated with an Arabidopsis AP2 and TGCAT is
an AP2 binding motif. Also, LaCount [39] had found that
ApiAP2s interact with chromatin remodeling factors and each
other. In addition, Fluek [40], had found a major role for the
Plasmodium falciparum ApiAP2 protein PfSIP2 in chromosome end
biology. aMYST A and aMYST B antibodies each altered TRP1
or TRP2 element binding of nuclear extract in EMSAs with shift
or modification of intensity of bands (Figure S1B). To confirm by
another method that TgMYST HATs are associated with RP
promoter regions, and more specifically the TRP elements, we also
used a chromatin immunoprecipitation (ChIP) assay. ChIP was
performed using aFlag antibody and parasite lysate derived from
RH strain tachyzoites engineered to express a Flag-tagged form of
TgMYST B. PCR for various regions of RP promoters of RPS13
and L12 was then performed on the immunoprecipitated DNA.
These data indicate that Flag-TgMYST B associates with each RP
promoter, including regions containing a TRP element (Figure 2B).
Additional methodological details and controls are presented in
Text S1. These results indicate that these TRPs in promoters of
Toxoplasma RP genes may associate with HATs.
Mass spectrometry of proteins extracted from EMSAs of TRP1
and TRP2 bound by nuclear extracts of T. gondii were then
performed. TRP1 and TRP2 associated nuclear extract proteins
indentified by mass spectrometry are shown in Table S1. These
included Ap2s, zinc fingers, zinc finger binding protein, a cold
shock DNA binding protein, SWI2/SNF2, CAM kinases, GCN5,
dead box helicases, NMD3, serine threonine phosphase 2C, a
splicing protein, eukaryotic translation initiation factors, SnoRNA,
MA3, transcription initiation factor EIF IIB, a ribosome biogenesis
regulatory protein, nuclear proteins NOPs, chaperones, heat shock
proteins, and RPL5.
Figure 1. Structural model of ribosomes and RPS13 sequence alignment. (A) Structural model of ribosome showing key locations of criticalfeatures of ribosomes and the ribosomal proteins. These are relevant to our model and critical in ribosome function in other organisms. RPS13(59.m03516) is in the catalytic site. RPS13 and RPL5 are contiguous. RPL5 (31.m00914) and RPL11 (583.m00014) occupy key locations and RPL4(42.m0584) is also in a key place at the point of egress of the assembled polypeptide chain from the ribosome. Ribosomal RNA assembly protein(55.m00169) and Elongation Factor (207.m00015_x and 207.m00015) interact with the ribosome at the catalytic site. (B) Sequence alignment of the T.gondii ribosomal protein S13, with similar sequences from mammalian, plant, bacterial and parasitic sources, with the T. thermophilus RPS15 sequenceat the bottom. Those residues that are identical and similar are boxed in red and orange, respectively. Those residues contacting the bound RNA inthe RPS15 structure are highlighted by a blue circle in the sequence alignment, showing that the conservation clusters around the RNA binding site.The right panel shows a structural model of the T. gondii RPS13 structure, based on the T. thermophilus RPS15 structure which is in complex withribosomal RNA, shown as a blue ribbon. For both the cartoon and surface representations of the RPS13 structure those residues conserved are shownin red. (See Movie S1.)doi:10.1371/journal.pone.0014057.g001
T. gondii rps13
PLoS ONE | www.plosone.org 3 November 2010 | Volume 5 | Issue 11 | e14057

Creation and characterization of rps13 conditionalknockdown parasites (Drps13) in vitro
Generation of ATc controlled conditional knockdown of
an endogenous rps13 gene in T. gondii. A targeting vector
construct containing the RPS13 promoter fragment in which four
TetO elements had been integrated in tandem just upstream of the
transcription start site as described previously [21] was used to
make conditional RPS13 knockdown mutant parasites herein
(Figure S2). The targeting vector described above [21] was
transfected into DHXGPRT RH parasites to facilitate recom-
bination of 4 TetO elements into the endogenous RPS13
promoter following the strategy outlined in Figure S2A (top,
middle and bottom). The resolved clone 1 of the mutant parasites
with the RPS13 endogenous promoter bearing 4 TetO elements
was transfected with a plasmid expressing a yellow fluorescent-Tet-
repressor (YFP-TetR) chimeric protein [20] that would block
transcription of RPS13 by binding the TetO elements in the
promoter. Twelve clones were selected for expression of the YPF-
TetR protein using fluorescence microscopy (data shown only for
clone 3 in Figure 3A). Figure S2B shows data comparing growth of
parental RH-HXGPRT and Drps13 parasite clones 3 and 9 in the
presence of anhydrotetracycline (+ATc). The Drps13 parasites
cultured with +ATc grew in fibroblasts at approximately the same
rate as the wildtype RH parasites and the parental HXGPRT
parasites.
Anhydrotetracycline regulates rps13 transcription. To
determine whether there was regulation of rps13 messenger RNA,
quantitative real-time PCR was used to measure rps13
transcription in parasites with and without anhydrotetracycline
(6ATc). RNA samples were taken from clone 3 grown for 4 and
24 hours. Amount of measured rps13 was normalized to total
RNA. Primers at the C terminus outside the region of the
construct for homologous recombinants were used. The effects of
culturing the conditional knockdown parasites -ATc on abrogating
rps13 mRNA were apparent early, at the 4-hour time point.
Reduction of mRNA was robust by 24 hours. We found a 3-fold
diminution of mRNA at 4 hours and a 10-fold diminution at
24 hours (Figure 3A). Abnormal transcription of rps13 within
hours of removing ATc, documents for the first time the utility of
this new knockdown system.
Anhydrotetracycline regulates expression of RPS13 and
proliferation of rps13 conditional knockdown parasites. It
was not known whether T. gondii RPS13 would be essential for
ribosome stability and biogenesis in this parasite, or whether there
would be regulation of or effect on replication of T. gondii with a
four TetO construct. Therefore, clones 3 and 9 were selected for
Figure 2. Ribosome gene promoters and transcription. (A) Nuclear extract proteins that associate with the TRP2 element. In EMSA withradiolabeled TRP2 probes with (+) and without (2) cold competitor TRP2 with tachyzoite nuclear extract, note mobility-shift which indicates thatthere is protein in the nuclear extract that binds to TRP2 which is competed by cold TRP2. Arrows with letters a to g in the TRP2 EMSAs designatesome of the bands that are no longer demonstrated when cold competitor for TRP2 is present. These letters designating bands are the same as inFigure S1B. There is also a similar mobility-shift for TRP1 with cold competitor (Figures S1A and B). Arrow in Figure S1A indicates one level in an EMSAwith TRP2 at which gel shift occurs for which there is no similar shift in mobility for a band in TRP1. Similar mobilities of proteins in TRP1 and TRP2EMSAs do not necessarily indicate identical bound or associated proteins. Mapping of key bases in the TRP2 elements shows that TRP2 contains amotif TGCATG known to interact with an Apicomplexan Apetela 2 transcription factor which we find herein is critical for binding nuclear extractproteins. The loss of different bands with differing mutations in the full TRP2 promoter indicates that additional bases also contribute to bindingmolecules in nuclear extracts. (B) TgMYST B appears to associate with RP gene promoters. Tachyzoites from a stable transgenic parasite cloneexpressing Flag-TgMYST B were processed for ChIP using anti-Flag antibody. PCR analysis for two distinct regions of RP gene promoters RPS13 [i] andL12 are shown [ii]. They are indicated as 1 and 2, respectively. Input DNA was used as a positive control for each primer pair and a ChIP samplewithout antibody was included as a negative control. TRP1 and two TRP2 elements are represented as gray boxes in the promoter region of each RPgene shown.doi:10.1371/journal.pone.0014057.g002
T. gondii rps13
PLoS ONE | www.plosone.org 4 November 2010 | Volume 5 | Issue 11 | e14057
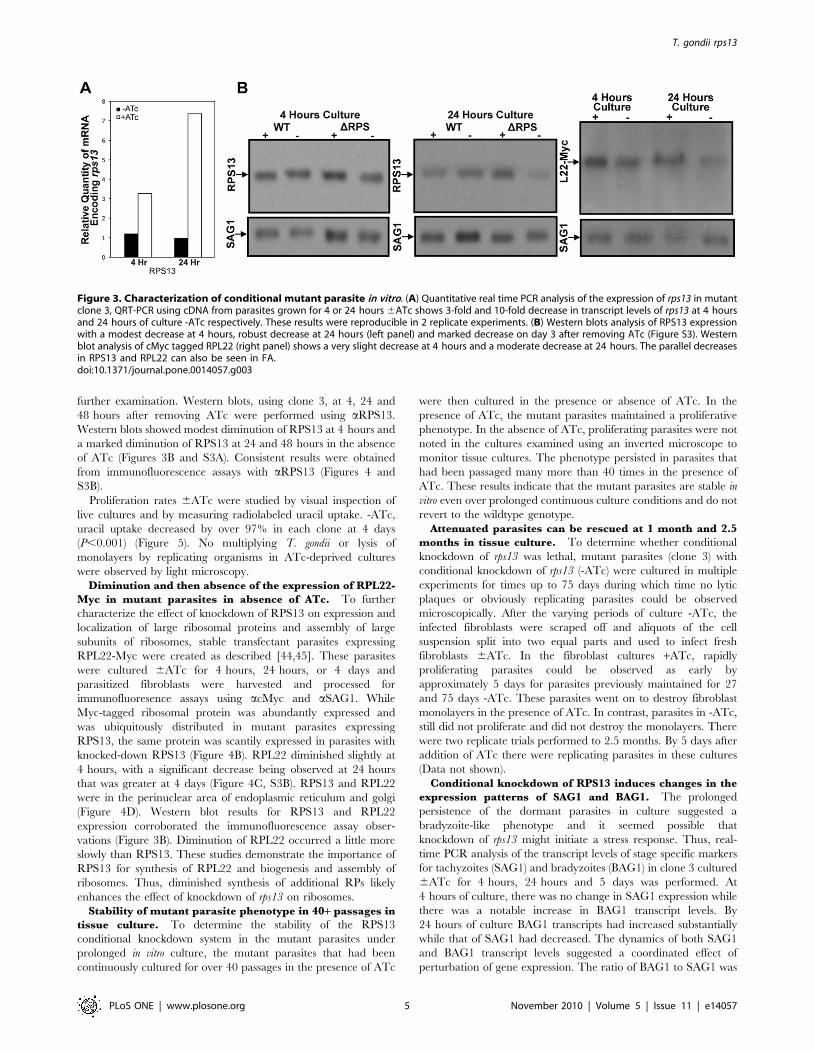
further examination. Western blots, using clone 3, at 4, 24 and
48 hours after removing ATc were performed using aRPS13.
Western blots showed modest diminution of RPS13 at 4 hours and
a marked diminution of RPS13 at 24 and 48 hours in the absence
of ATc (Figures 3B and S3A). Consistent results were obtained
from immunofluorescence assays with aRPS13 (Figures 4 and
S3B).
Proliferation rates 6ATc were studied by visual inspection of
live cultures and by measuring radiolabeled uracil uptake. -ATc,
uracil uptake decreased by over 97% in each clone at 4 days
(P,0.001) (Figure 5). No multiplying T. gondii or lysis of
monolayers by replicating organisms in ATc-deprived cultures
were observed by light microscopy.
Diminution and then absence of the expression of RPL22-
Myc in mutant parasites in absence of ATc. To further
characterize the effect of knockdown of RPS13 on expression and
localization of large ribosomal proteins and assembly of large
subunits of ribosomes, stable transfectant parasites expressing
RPL22-Myc were created as described [44,45]. These parasites
were cultured 6ATc for 4 hours, 24 hours, or 4 days and
parasitized fibroblasts were harvested and processed for
immunofluoresence assays using acMyc and aSAG1. While
Myc-tagged ribosomal protein was abundantly expressed and
was ubiquitously distributed in mutant parasites expressing
RPS13, the same protein was scantily expressed in parasites with
knocked-down RPS13 (Figure 4B). RPL22 diminished slightly at
4 hours, with a significant decrease being observed at 24 hours
that was greater at 4 days (Figure 4C, S3B). RPS13 and RPL22
were in the perinuclear area of endoplasmic reticulum and golgi
(Figure 4D). Western blot results for RPS13 and RPL22
expression corroborated the immunofluorescence assay obser-
vations (Figure 3B). Diminution of RPL22 occurred a little more
slowly than RPS13. These studies demonstrate the importance of
RPS13 for synthesis of RPL22 and biogenesis and assembly of
ribosomes. Thus, diminished synthesis of additional RPs likely
enhances the effect of knockdown of rps13 on ribosomes.
Stability of mutant parasite phenotype in 40+ passages in
tissue culture. To determine the stability of the RPS13
conditional knockdown system in the mutant parasites under
prolonged in vitro culture, the mutant parasites that had been
continuously cultured for over 40 passages in the presence of ATc
were then cultured in the presence or absence of ATc. In the
presence of ATc, the mutant parasites maintained a proliferative
phenotype. In the absence of ATc, proliferating parasites were not
noted in the cultures examined using an inverted microscope to
monitor tissue cultures. The phenotype persisted in parasites that
had been passaged many more than 40 times in the presence of
ATc. These results indicate that the mutant parasites are stable in
vitro even over prolonged continuous culture conditions and do not
revert to the wildtype genotype.
Attenuated parasites can be rescued at 1 month and 2.5
months in tissue culture. To determine whether conditional
knockdown of rps13 was lethal, mutant parasites (clone 3) with
conditional knockdown of rps13 (-ATc) were cultured in multiple
experiments for times up to 75 days during which time no lytic
plaques or obviously replicating parasites could be observed
microscopically. After the varying periods of culture -ATc, the
infected fibroblasts were scraped off and aliquots of the cell
suspension split into two equal parts and used to infect fresh
fibroblasts 6ATc. In the fibroblast cultures +ATc, rapidly
proliferating parasites could be observed as early by
approximately 5 days for parasites previously maintained for 27
and 75 days -ATc. These parasites went on to destroy fibroblast
monolayers in the presence of ATc. In contrast, parasites in -ATc,
still did not proliferate and did not destroy the monolayers. There
were two replicate trials performed to 2.5 months. By 5 days after
addition of ATc there were replicating parasites in these cultures
(Data not shown).
Conditional knockdown of RPS13 induces changes in the
expression patterns of SAG1 and BAG1. The prolonged
persistence of the dormant parasites in culture suggested a
bradyzoite-like phenotype and it seemed possible that
knockdown of rps13 might initiate a stress response. Thus, real-
time PCR analysis of the transcript levels of stage specific markers
for tachyzoites (SAG1) and bradyzoites (BAG1) in clone 3 cultured
6ATc for 4 hours, 24 hours and 5 days was performed. At
4 hours of culture, there was no change in SAG1 expression while
there was a notable increase in BAG1 transcript levels. By
24 hours of culture BAG1 transcripts had increased substantially
while that of SAG1 had decreased. The dynamics of both SAG1
and BAG1 transcript levels suggested a coordinated effect of
perturbation of gene expression. The ratio of BAG1 to SAG1 was
Figure 3. Characterization of conditional mutant parasite in vitro. (A) Quantitative real time PCR analysis of the expression of rps13 in mutantclone 3, QRT-PCR using cDNA from parasites grown for 4 or 24 hours 6ATc shows 3-fold and 10-fold decrease in transcript levels of rps13 at 4 hoursand 24 hours of culture -ATc respectively. These results were reproducible in 2 replicate experiments. (B) Western blots analysis of RPS13 expressionwith a modest decrease at 4 hours, robust decrease at 24 hours (left panel) and marked decrease on day 3 after removing ATc (Figure S3). Westernblot analysis of cMyc tagged RPL22 (right panel) shows a very slight decrease at 4 hours and a moderate decrease at 24 hours. The parallel decreasesin RPS13 and RPL22 can also be seen in FA.doi:10.1371/journal.pone.0014057.g003
T. gondii rps13
PLoS ONE | www.plosone.org 5 November 2010 | Volume 5 | Issue 11 | e14057

elevated in the absence of ATc suggesting an early ‘‘stress’’
response. These findings were reproducible in two replicate
experiments (Figure 6A). Immunostaining for SAG1 and BAG1
confirmed these findings (Figure 6B, C). Staining with aBAG1 and
aSRS9/BRS4 (another bradyzoite protein) were present in some
parasites (Figure 6B, D top), but reactivity with Dolichos was
absent (Figure 6C top, D top). In stressed parasites used as controls
for this staining, BAG1 (Figure 6B) and BRS4/SRS9 (Figure 6C,
D bottom) were also present. Dolichos staining of a cyst wall
component was also present (Figure 6C bottom, D bottom).Parasites are arrested at G1. Abrogation of ribosomal
proteins in yeast results in arrest in G1 [13–15]. Thus, it was of
interest to determine whether abrogating ribosome function might
cause arrest in G1 of the cell cycle. Tachyzoites cultured +ATc
exhibit a normal asynchronous profile but those cultured -ATc are
largely 1N at 24 hours as demonstrated by staining with
propidium iodide and subsequent FACs analysis (Figure 7A).
The 2N peak in -ATc samples is diminished compared to the peak
in +ATc samples. By gating plots, percentage of parasites in
G1 was quantified as 68.5% +ATc, a G1 profile seen in
asynchronously growing parasites [46,47], and as 85% -ATc as
seen in synchronously growing parasites. Therefore, by FACS, -
Figure 4. Immunofluorescence assay analyses of RPS13 and RPL22. (A–C) Immunofluorescence assay analysis of the expression of RPS13 inwildtype (wt,A) and Drps13 (B) parasites at 4 and 24 hours of culture 6ATc. RPS13 protein is shown as green fluorescent protein with the parasite andhost cell nuclei stained blue. In Drps13 parasites, in the absence of ATc, RPS13 expression is shown to decrease modestly at 4 hours and significantlyat 24 hours of culture. Rabbit aRPS13 was used as described in the text. Immunofluorescence assay analysis of the expression of Myc-taggedribosomal L22 protein (L22-Myc) in L22-Myc translateds Drps13 mutant parasites at 4 and 24 hours of culture 6ATc (C). L22-Myc protein is shown asgreen fluorescent protein with the parasite and host cell nuclei stained blue. In the absence of ATc, L22-Myc expression is shown to decreasemodestly at 4 hours and significantly but slightly less than RPS 13, at 24 hours and 4 days (Figure S3) of culture (D) Confocal microscopy shows RPS13 and RPL22-Myc at the perimeter of the nucleus in the area of the endoplasmic reticulum and golgi apparatus in the merged image.doi:10.1371/journal.pone.0014057.g004
Figure 5. Uracil incorporation assay. Parasites were grown for 3days 6ATc at which point radiolabeled uracil was added. Uracilincorporation into parasites was measured 24 hours later. Both clones 3and 9 showed a 97% decrease in incorporation of uracil at 4 days.doi:10.1371/journal.pone.0014057.g005
T. gondii rps13
PLoS ONE | www.plosone.org 6 November 2010 | Volume 5 | Issue 11 | e14057

ATc parasites show an increased G1 distribution. These studies
were replicated twice and the results, 85% -ATc, 68.5% +ATc
were similar.
Global transcriptome analysis indicates that depletion of
rps13 leads to cell cycle arrest in G1. Since FACS analysis
suggested that parasites were arrested in G1, transcriptomes were
analyzed to determine whether RPS13 depletion would have an
effect on global mRNA profile. Electrophoretic analysis of RNA
used for microarray hybridization showed that -ATC, small
subunit RNA (18S) is markedly diminished in RPS13 conditional
mutants, but large subunit RNA is not (28S) (Figure 7B to D).
Ribosomal RNA makes up .80% of total RNA in the cell,
therefore rRNA quality and quantity reflect that of the mRNA
population. Since the majority of the sample RNA consists of 28S
and 18S rRNA species, the 28S:18S rRNA ratio has traditionally
been viewed as the primary indicator of RNA quality, with a ratio
of 1.7 to 2.0 considered to indicate high-quality, intact RNA.
+ATc samples had rRNA ratios of 1.7, demonstrating robust,
intact RNA. -ATc samples had unusually high rRNA ratios (4.5 to
4.8) that were out of the normal 1.7 to 2.0 range, suggesting
abnormal RNA integrity. Decreased area of 18S rRNA peaks in
these samples suggest small subunit (18S) breakdown.
Affymetrix Toxoplasma microarray data was analyzed to
determine whether there was an association with a G1 transcript
profile. Four hundred and seventy-five genes with significant gene
expression differences (Welch T-test, P = 0.01) between 6ATc
samples were identified. These gene expression differences are
represented in the heat map in Figure 7E. Genes were clustered
into those increased (153) or decreased (322) -ATc. There are a
number of genes with diminished expression and pronounced over
expression -ATc including LDH1, 4 zinc finger proteins, leucine
rich protein, ABC transporter-like molecules, a bradyzoite small
heat shock protein, myosin related molecules with many fold
increases and one AP2 protein with a small increase (Table S2).
These gene sets have a large overlap with G1 genes defined in an
ongoing cell cycle expression study in T. gondii (Behnke, White, in
preparation). Over 60% of the genes that have diminished
expression -ATc overlap with genes that are diminished at G1,
and over 40% of genes with increased expression -ATc overlap
with genes that are up-regulated at G1. A hypergeometric
distribution of this association suggests that the probability of this
happening by chance is essentially zero. Thus, by both FACS and
microarray analysis, parasites -ATc display a G1 profile. Figure 8
shows this G1 arrest and its consequences schematically.
Characterization of RPS13 mutant parasites in vivoRPS13 conditional parasites are not lethal for mice -ATc
and are lethal +ATc administered on the day of infection but
Figure 6. Message for SAG1 and BAG1 ±ATc for 4 or 24 hours of culture and immunofluorescence analyses for BAG1, SAG1, SRS9,and Dolichos. (A) Increased message for BAG1 and decreased message for SAG1 -ATc at 24 hours. There was no effect on BAG1 and SAG1 mRNA at4 hours. These results were reproducible in two replicate experiments. WT = wildtype strain. RPS13 = conditional mutant. (B) To visualize the parasitecysts, Dolichos staining is green. Tachyzoite surface protein, SAG1 staining is red. The parasite and fibroblast nuclei stain blue. In either + or – ATcconditions, there is no Dolichos staining. In C, SAG1 staining is green, BAG1 staining is also green, DAPI is stained blue. -ATc BAG1 is present while+ATc BAG1 is absent. Control staining with 5 days of pH stress +ATc also is shown. (D) –Atc for 24 hours BRS4/SRS9 staining (red) is seen in someparasites but Dolichos staining is absent (top). Control staining of +ATc parasites that have been stressed with pH shock (bottom panel) show bothgreen Dolichos staining and red BRS4/SRS9 staining, with dapi marking the parasite nuclei. –ATc parasite BAG1 transcript and protein increase. BRS4/SRS9 protein increases but Dolichos is not present in a cyst wall. These Drps13 parasites when stressed with pH shock are capable of differentiating toparasites that form structures that stain with Dolichos as well as producing BAG1 and BRS4. -ATc, some Drps13 parasites stain for BAG1 and SRS9, butnot for Dolichos.doi:10.1371/journal.pone.0014057.g006
T. gondii rps13
PLoS ONE | www.plosone.org 7 November 2010 | Volume 5 | Issue 11 | e14057
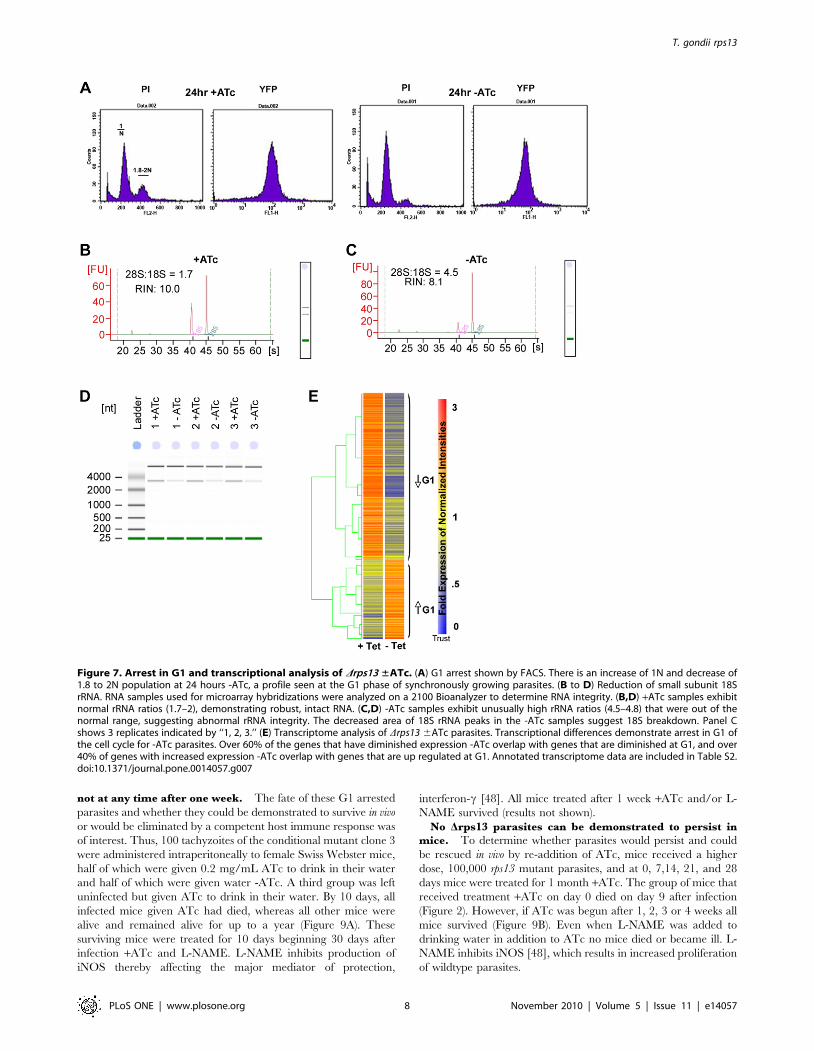
not at any time after one week. The fate of these G1 arrested
parasites and whether they could be demonstrated to survive in vivo
or would be eliminated by a competent host immune response was
of interest. Thus, 100 tachyzoites of the conditional mutant clone 3
were administered intraperitoneally to female Swiss Webster mice,
half of which were given 0.2 mg/mL ATc to drink in their water
and half of which were given water -ATc. A third group was left
uninfected but given ATc to drink in their water. By 10 days, all
infected mice given ATc had died, whereas all other mice were
alive and remained alive for up to a year (Figure 9A). These
surviving mice were treated for 10 days beginning 30 days after
infection +ATc and L-NAME. L-NAME inhibits production of
iNOS thereby affecting the major mediator of protection,
interferon-c [48]. All mice treated after 1 week +ATc and/or L-
NAME survived (results not shown).
No Drps13 parasites can be demonstrated to persist in
mice. To determine whether parasites would persist and could
be rescued in vivo by re-addition of ATc, mice received a higher
dose, 100,000 rps13 mutant parasites, and at 0, 7,14, 21, and 28
days mice were treated for 1 month +ATc. The group of mice that
received treatment +ATc on day 0 died on day 9 after infection
(Figure 2). However, if ATc was begun after 1, 2, 3 or 4 weeks all
mice survived (Figure 9B). Even when L-NAME was added to
drinking water in addition to ATc no mice died or became ill. L-
NAME inhibits iNOS [48], which results in increased proliferation
of wildtype parasites.
Figure 7. Arrest in G1 and transcriptional analysis of Drps13 ±ATc. (A) G1 arrest shown by FACS. There is an increase of 1N and decrease of1.8 to 2N population at 24 hours -ATc, a profile seen at the G1 phase of synchronously growing parasites. (B to D) Reduction of small subunit 18SrRNA. RNA samples used for microarray hybridizations were analyzed on a 2100 Bioanalyzer to determine RNA integrity. (B,D) +ATc samples exhibitnormal rRNA ratios (1.7–2), demonstrating robust, intact RNA. (C,D) -ATc samples exhibit unusually high rRNA ratios (4.5–4.8) that were out of thenormal range, suggesting abnormal rRNA integrity. The decreased area of 18S rRNA peaks in the -ATc samples suggest 18S breakdown. Panel Cshows 3 replicates indicated by ‘‘1, 2, 3.’’ (E) Transcriptome analysis of Drps13 6ATc parasites. Transcriptional differences demonstrate arrest in G1 ofthe cell cycle for -ATc parasites. Over 60% of the genes that have diminished expression -ATc overlap with genes that are diminished at G1, and over40% of genes with increased expression -ATc overlap with genes that are up regulated at G1. Annotated transcriptome data are included in Table S2.doi:10.1371/journal.pone.0014057.g007
T. gondii rps13
PLoS ONE | www.plosone.org 8 November 2010 | Volume 5 | Issue 11 | e14057

Robust protection against challenge with wildtype RH
strain tachyzoites following immunization with Drsp13
demonstrates that DRPS13 can function as a vaccine.
Since Drps13 mutant parasites were markedly attenuated, we
hypothesized that they therefore could be immunogenic,
presenting both tachyzoite and some stress related bradyzoite
antigens (e.g. BAG 1), and persist at least for a short period of time
while they elicited an immune response. Thus, in two replicate
experiments, groups of mice were given 100,000 RPS13
conditional mutant parasites and all mice survived. They then
received booster doses of 100,000 RPS13 conditional mutant
parasites 2 weeks and 4 weeks later without ill effect. Two weeks
later they were challenged with 2,000 RH tachyzoites and all
unimmunized control mice died while all immunized mice
survived (Figure 9C). In separate experiments (data not shown)
immunized mice challenged 1 year later were protected (,80 to
90% survival) against otherwise lethal challenge.
A similar set of two replicate experiments was performed after
immunization with a single dose of 1,000 or 10,000 or 100,000
mutant parasites with challenge with 2,000 RH strain parasites 8
weeks later. A dose of 100,000 parasites resulted in complete
protection. Doses of 10,000 parasites resulted in ,50 to 80%
protection and lower doses were not protective (Figure 9D).
To determine whether any parasite DNA could be found to
persist in brains of these immunized and challenged mice, PCR
with primers to detect the multi-copy B1 gene and quantitative
PCR to detect the 300 repeat copy 529 base pair gene was
performed with brain from mice infected for prolonged periods of
time after immunization with Drps13 -ATc. This did not reveal T.
gondii B1 or 300 copy (529 bp) gene DNA in any of 8 mice (data
not shown). RH tachyzoite DNA (this PCR method was sensitive
in detection of 4 parasites) was the positive control for this
quantitative PCR experiment (r2 = 0.99).
Protection against Type II parasites. In these
experiments, there were 5 unimmunized mice in each trial with
a total of 10 unimmunized mice. There were 5 mice immunized
one time (16) and 5 mice immunized two times (26) mice in each
trial and thus a total of 10 immunized mice for each type of
immunization (16, 26). They were challenged with 50 Me49 cysts
i.p. and at 30 days after challenge, cyst numbers were found to
decrease markedly in immunized mice (Figure 9E; P,0.01). In
these experiments, qPCR of brain using the 300 copy 529 bp gene
was performed following challenge of control and immunized
mice. This demonstrated marked diminution in parasite burden in
brain following immunization, most dramatically for those
immunized two times compared to the unimmunized group
(Figure 9F, P#0.001). Pathology still was present, although less,
and rare cysts were seen following immunization and challenge
when compared with unimmunized controls. Pathology was more
focal in the immunized mice but quantitation of histopathology
did not differ significantly between mice in groups that had been
immunized once or with a booster immunization (26) (Figure 9G).
Discussion
Our finding that abrogation of a ribosomal protein causes a
specific G1 arrest confirms the strength of the refined Tet model
for conditional gene knockdown. This influence of ribosomes on
proliferation and persistence has not been demonstrated previ-
ously in T. gondii, although there is precedent for this fundamental
biological process in other eukaryotes such as yeast. Our findings
provide an in vitro model which suggests that T. gondii’s sensing of
intra-parasite stresses could modulate regulation of transcription of
its rp genes. RPs influence biogenesis of ribosomes, and
subsequently, the ability of the parasite to either proliferate or
arrest in G1. Analysis of promoters of RPs led Schaap and van
Figure 8. Model of interactions of ribosomal protein promoter elements, regulation of RP synthesis and G1 arrest. This model is basedon data herein, which complements and is consonant with data in the literature [24–26,49,52–57,73] and [Kim et al, personal communication, 2010].This image shows a model of interactions of ribosomal protein promoter elements with complexes that regulate their transcription. It also shows theeffects of diminution of transcription of a RP and the additional stress of abrogation of protein synthesis in our in vitro model system. In this model,stress activates signaling pathways. Signaling pathways are indicated by small colored symbols. This signaling decreases transcription of RPs. Thisdecreased RP leads to diminution of ribosomes and arrest of proliferation, and arrest in G1. We hypothesize the following explains our findings: Stressis detected and there is signaling via PKA [24] and TOR [24,26] which leads to diminished transcription of certain ribosomal genes. Transcriptionfactors interacting with RP gene promoter elements such as TRP1 and TRP2 include AP2s (Kim et al, personal communication, 2010), and theresponse elements associate with chromatin remodelers as demonstrated herein. In this model, we propose than when there is incomplete synthesisof ribosomal proteins, then p53 is not degraded and there is cell cycle arrest in G1, as we found herein when rps13 transcription was diminished.doi:10.1371/journal.pone.0014057.g008
T. gondii rps13
PLoS ONE | www.plosone.org 9 November 2010 | Volume 5 | Issue 11 | e14057

Figure 9. In vivo experiments. (A) Mice given RPS13 mutant -ATc survive. Data shown to day 9 but experiment continued to 6 months. Mice givenRPS13 mutant +ATc simultaneously all die. Those given mutant but no ATc all survived. Two hundred RPS13 mutant parasites were administered i.p.(B) Administration of ATc at varying times simultaneously with or after 100,000 RPS13 mutant parasites. Simultaneous administration of ATc andparasites is lethal but not administration of ATc at 1 or more weeks after administration of 100,000 RPS13 mutant parasites. (C) Complete protectionagainst challenge with 2,000 RH tachyzoites by immunization. There was complete protection with a single dose of 100,000 parasites. Challenge was
T. gondii rps13
PLoS ONE | www.plosone.org 10 November 2010 | Volume 5 | Issue 11 | e14057

Poppel to postulate that two conserved rp gene promoter cis
elements, TRP1 and TRP2, regulate expression of rp genes, and
thereby, biogenesis of ribosomes [49]. Schaap and van Poppel had
postulated that TRP2 would be an enhancer and TRP1 a
suppressor of rp transcription [49]. Empirical evidence from our
studies herein, along with the work of others (Kim et al, personal
communication, 2010, [50], [72]) is consistent with a role for AP2
transcription factors that interact with TRPs and regulate RP
synthesis, and thus specify parasite cell and, at least in part, life
cycle stages. Using rolling mutations to study TRP2, we found that
a specific six base TRP2 motif is essential for binding nuclear
extract proteins (Figure 2). Very recently, this same TRP2
promoter element was predicted to bind an AP2 [29], and later
observed to bind a T. gondii AP2, X1-3 (K. Kim, personal
communication, 2010). Movement of AP2s to the nucleus occurs
in waves associated with the cell cycle and transcription of genes
[72]. In G1a of the cell cycle in the tachyzoites, hundreds of genes
including ribosomal genes are expressed. These have the
TGCATG in their promoters [72]. Another group of investigators
found that the TGCATG element in TRP2 in the promoter of rpl9
enhances transcription of a reporter gene [50] in T. gondii, as van
Poppel and Schaap had postulated. These latter studies occurred
while we found that these six bases that are part of the TRP2
promoter element were necessary, but not sufficient, for binding
the nucleoproteins in the complex in our EMSAs. We found that
additional C terminal bases also were critical for binding/
associating with nuclear extract proteins. The loss of different
bands with differing mutations in the full TRP2 promoter element
suggests that additional bases also contribute to a complex or
complexes of associated nuclear extract proteins. For mutants 7
and 8 all associations with all nuclear extract proteins are
abrogated. One explanation for this might be that an association
with some of the other proteins with bases absent in mutants 7 and
8 could be critically dependent for or on protein(s) associated with
TGCATG. Otherwise, it might be expected that only some of the
bands would disappear because the TGCATG core site is intact in
mutants 7 and 8. Recent work of others [51] indicates approaches
whereby transcription factors such as AP2s might be inhibited that
may be pertinent to future work. Future studies of mutations in the
rps13 promoter using a luciferase reporter may be useful in
clarifying the importance of these bases in both of its TRP2
elements in enhancing transcription. Their relative importance
and interaction with the TGTGTG promoter element, previously
shown to be important in bradyzoite gene transcription, in the
rps13 promoter also will be of interest in future studies.
In addition, we found that aMYST A and B caused shifts in
our EMSAs with both TRP1 and TRP2. Myst B (GCN5) was
identified by mass spectrometry of nuclear factors binding TRPs
1 and 2. Also, in ChIPs, antibody to flag tagged MYST B
suggested interactions with TRP1 and 2. Attempts to perform
this ChIP study for TgMYST A were complicated by the fact that
an additional copy of TgMYST A is lethal to Toxoplasma [25].
These findings all support association of MYST A and B HATs
with proteins bound to TRP1 and 2 or direct binding of these
HATs to rp promoters. These results are similar to the association
of HATS with AP2 transcription factors that regulate genes in
Arabidopsis [29] and Plasmodia [39,40]. A protein interaction
network in Plasmodia demonstrated an AP2 interaction with
GCN5 histone acetyl transferases and Swi2/Snf2 ATPases [39].
These MYST family HATs may bind via an AP2 complex as
occurs in Arabidopsis [29] or could bind directly to DNA as occurs
in a mammalian MYST HAT. Another histone interacting
protein, HDAC3, also associated with an AP2 (TgCRC35) [23].
It is likely that other transcription factors also participate in
regulation of production of RPS13 and other RPs. For example,
there is also a TGTGTG in the RPS13 promoter. This motif is
critical in controlling expression of genes in parasites stressed in
vitro [22]. Proteins bound in our EMSAs (Table S1) identified by
mass spectrometry suggest that certain proteins including AP2,
GCN5 (MystB), Swi2/Snf2 ATPases may be in the complex in T.
gondii. Further, future studies with knockdown and tagging these
genes, ChIP, and immunolocalization will be useful in definitely
proving there is truly an association and if so in characterizing the
association.
We note, relevant to our model (Figure 8), as suggested by our
data, that in yeast, TOR and PKA pathways regulate ribosomal
genes in response to conditions of amino acid deprivation and
other cellular stress-associated signals. Sensing of stress is through
long chain fatty acids, ceramide synthases and ceramide
phosphatases. All the molecules in the PKA pathway [53] and
some in the TOR pathway [26] are present in T. gondii and a
TOR-like protein appears to be transported into the nucleus.
Putative homologues of kinases, transcription factors, cyclins and a
ceramide activated protein phosphatase that is involved in sensing
cellular stress and mediates G1 arrest via TORC2 in yeast have
been identified in T. gondii [13–15,54] (K. Kinsley, E. Mui, R.
McLeod, unpublished data; Figure 8). The same families of
proteins that interact with components of the TOR and PKA
pathways in yeast (e.g. HATs) also associate with TRP2 in the
promoter of rps13 and TRP1 in the promoters of other RPs in T.
gondii. This further suggests that TOR and PKA pathways may
participate in regulation of proliferation and persistence in T.
gondii. Roles for long chain fatty acids and ceramide synthases
producing ceramides and signaling through TORC2 [55] are
likely because homologues of these proteins key in stress responses
in yeast also are present in T. gondii. Sensing of cellular stress in
plants is mediated by PYR/PYL/RCAR [56] and abscisic acid
which modulates a PP2 phosphatase via the ligand (ABA)-bound
receptor. These pathways modulate calcineurin and calcium fluxes
and have been found in T. gondii [57].
8 weeks after the immunizing dose (to 3 months). (D) Challenge of Drps13 immunized SW mice that received varying immunizing doses of Drps13.Doses range from 1,000 to 100,000 Drps13 tachyzoites with RH strain tachyzoites. There is one hundred percent survival with 100,000 immunizingdose. (E) Reduction of cyst numbers following challenge of Drps13 immunized mice with Type 2 parasites. Immunization results in reductions in cystnumbers. (F) Reduction of parasite burden following challenge of Drps13 immunized mice with Type 2 parasites. Parasite burden demonstrated byqPCR for the 300 copy 529 base pair gene is diminished by immunization. (G) Reduction of brain pathology following challenge of Drps13 immunizedmice with Type 2 parasites. Histopathology, and brain pathology following challenge of control unimmunized and mice immunized one or two timesand challenged with ME49 (Type 2) parasites was analyzed. There was significantly less parasite burden and pathology following this Type 2challenge of the immunized mice, but infection and pathology were not eliminated completely. There appeared to be somewhat greater protectionfollowing two immunizations but the differences did not reach statistical significance. Cyst number was quantitated on a scale of 0 to 5, as werehippocampal perivascular cuffing, intraparenchymal inflammatory process, inflammation in the leptomeninges, and inflammatory process in thevasculature within the brain parenchyma. Data are expressed as number of mice, mean with standard deviation, median, and range. Differencesbetween immunized and unimmunized mice were statistically significant for all but parenchymal inflammation and perivascular cuffing inparenchyma.doi:10.1371/journal.pone.0014057.g009
T. gondii rps13
PLoS ONE | www.plosone.org 11 November 2010 | Volume 5 | Issue 11 | e14057

Further, growth arrest in yeast and animals can be mediated by
depletion of ribosomal proteins. In yeast, ribosome biogenesis is
linked to G1 of the cell cycle [14]. Recently Ferreiria-Cerca,
Fumagalli, and others [1–3,10] found that eukaryotic ribosomal
proteins and subunits are delicately balanced. In this work with
hepatic cells, Fumagalli found that when RPL11 and MDM2 (a
ubiquitin ligase that degrades p53) interaction is modified, by
diminishing RPL11, p53 degradation is altered. When p53 is not
degraded, it causes cell cycle arrest in G1 [3]. Whether this precise
molecular mechanism also occurs in T. gondii and whether it is
operative in vivo remains to be determined. Plasmodia monitor
nutrient support for translation of proteins before committing to
DNA synthesis. The cell cycle mechanisms sensing ribosomes or
translation factors [73,74] are connected with induction of genes
required for DNA replication in late G1/S through G1 cyclin
regulation, although protein interactions in yeast and metazoans
differ [72].
To study effects of ribosomal protein S13 on proliferation and
persistence in T. gondii in vitro, a new conditional knockdown
system was used with a TetR knockdown system using constructs
discussed earlier [20,21]. Theoretical strengths of this YFP-TetR
conditional knockdown system are that the modified and the
native promoter have the same robustness and also, theoretically
that ATc does not need to access encysted bradyzoites for
knockdown. Similar YFP-TetR conditional knockdown systems
also had demonstrated stringent regulation of gene function with
low concentrations of ATc in other organisms. This system had a
88-fold regulation of transcription of rps13 using a reporter gene in
T. gondii [20]. We subsequently, herein, successfully did find this
system to be effective for conditional knockdown of the T. gondii
endogenous RPS13 gene. Separately we have found this system is
effective in regulating a novel dense granule protein and parasite
enzymes (W. Witola, K. El Bissati, R. McLeod, manuscripts in
preparation, 2010).
The putative RPS13 selected to attempt to interfere with
ribosome function using this new conditional knockdown system
was chosen because it has a single transcription start site [20,21].
Although it was not known at the time the reagents were created
[21] or our studies were initiated, this turned out to be a fortuitous
choice because recently RPS13 was determined to be in the
ribosome catalytic site [5–8,58–60] in other organisms (Figure 1A).
Herein, our deduced structure of the putative RPS13 selected for
conditional knockdown was modeled using the published structure
of Thermophilus RPS15 as a basis because this latter structure had
been solved. This confirmed that this T. gondii molecule has all the
features to be a RPS13. It has residues likely to interact with
ribosomal RNA (blue circles, Figure 1B, on line movie RP link)
and thus to function in catalysis and initiating translation rather
than only as ribosome scaffold. It thereby should be critical for
protein assembly. In S. cerevisiae the RPS13 homologue is in the
rRNA/r-protein neighborhood [4–8] and in E. coli RPS13 is
essential for ratcheting small and large ribosomes contiguous to
each other and is the direct link between the tRNA-binding site
and movements of the head of the small ribosomal subunit
(Figure 1A) [8]. The conditional TetR knockdown method used
herein was robust, resulting in diminished specific rps13 mRNA
and RPS13 protein synthesis in T. gondii and arrest of proliferation.
This validated RPS13, and molecules associated with its
regulation, as molecular targets, and the utility of this conditional
ATc repressor knockdown system for such target validation.
However, since RPS13 is so similar in sequence and deduced
structure to other RPS13s, including human RPS13 (Figure 1B),
our modeling did not offer obvious insights into targeting T. gondii
RPS13 for medicine discovery.
Following reduction in rps13 mRNA in the conditional
knockdown parasite, western blot and immunofluorescence assays
we observed that both RPS13 and RPL22 diminish but RPS13
diminishes earlier (Figure 4A,B). Diminution of RPS13 begins
modestly at 4 hours and is substantial by 24 hours. RPL22 is not
appreciably modulated at 4 hours, is moderately diminished by
24 hours and decreases more over the next days in culture. The
ratio of 18S:28S RNA diminishes (Figure 7). This diminution of
18S rRNA indicates a role for RPS13 in either stabilization or
processing of rRNA.
Conditional knockdown led to increased expression of BAG1
(Figure 6), arrest in the G1 phase of the cell cycle (Figure 7) and
thereby arrest of parasite proliferation and persistent parasites in
this tissue culture model. These observations suggested that
depletion of ribosomes leads to synthesis of BAG1 (Figure 6) and
initiates an early stress response that induces parasite G1 arrest in
vitro (Figure 7, Table S2).
The forced proliferation arrest observed herein shares the G1
arrest feature associated with the in vitro pH or compound 1
stressed parasite stage. This type of G1 arrest also was observed by
Radke in bradyzoites isolated from mouse brain in earlier studies
[61]. However, morphology and other features of this mutant are
not those of typical fully differentiated bradyzoites (Figure 6B-D).
For example, Dolichos staining is absent (Figure 6). Expression
data of 144 ribosomal proteins in type I-II-III tachyzoites and
bradyzoites samples (expression data with the T. gondii Affymetrix
Array available at GSE16037 and ToxoDB.org) suggests that most
ribosomal proteins, including RPS13, do not have altered
expression across strain or developmental stages. There are a
few exceptions, however. For example, RPL4 transcript levels are
increased in type III CTG. RPL4 (42.m05824) was recently found
to be a part of the channel in ribosomes for assembling proteins
[62] as well as being critical in stress responses in yeast [62].
Location of certain relevant RPs in ribosomes is shown in
Figure 1A.
Our work provides novel insights into significance of this protein
for this parasite’s cell cycle, proliferation, and persistence in tissue
culture after removal of ATc. Our Drps13 mutant parasites can
persist in vitro in this proliferation arrested state for prolonged
periods of time (months) and yet be revived by adding ATc. This is
reminiscent of how plants can ‘‘go to sleep for a long, long time’’
through the PYR abscisic acid calcineurin stress response pathway
[56]. T. gondii has this pathway. The recent work of Sibley et al
demonstrates that this ABA system is operative in T. gondii.
Some of the genes that are expressed in fully differentiated
bradyzoites were not transcriptionally upregulated -ATc (Table
S2). This suggests that Drps13 can maintain functions vital for
persistence while neither replicating nor differentiating to encysted
bradyzoites fully. Lack of ribosomes themselves are a model
leading to arrest in G1 in other organisms [3] and may also
function in this manner in T. gondii (Figure 8). The mechanisms
demonstrated in vitro herein suggest that stress may be translated
through diminished transcription of ribosomal proteins to
depletion or markedly diminished numbers of ribosomes and
then to G1 arrest, at least in some cases. With an additional
mechanism(s) that allows escape from the immune response, an
imbalance of ribosomal proteins and diminution of ribosomes
might allow a variety of microorganisms that can stop replicating
and persist in a basal state to persist in vivo for a long time.
Our conditional mutant parasites do not appear able to evade
the immune response in vivo. They appear to be incapable of
persistence in the brain in the presence of a competent immune
response. They cannot be demonstrated to be revived in vivo by
subsequent administration of ATc administered alone or with L-
T. gondii rps13
PLoS ONE | www.plosone.org 12 November 2010 | Volume 5 | Issue 11 | e14057

NAME. Nonetheless, they protect mice against challenge with
wildtype clonal Type 1 parasites completely, or clonal Type 2
parasites robustly. They are proving useful as internal ‘‘gold
standard of protection’’ controls in ongoing studies to develop
biomarkers and to produce a component vaccine to protect
humans against toxoplasmosis (Cong H, McLeod R et al, in
preparation).
Since bradyzoites in vivo, like Drps13 in vitro, also are arrested in
G1 [61], it seems reasonable to speculate that this diminished
replication phenotype is a critical part of a response to stresses in
vivo and could contribute to the slowed replication in the full
encysted, bradyzoite phenotype. However, although it may be a
component part of development of a bradyzoite phenotype and
even necessary, the studies herein demonstrate that it is not
sufficient for the full phenotype. At a minimum, other genes which
allow for the sequestration from the host’s immune response and
for other essential bradyzoite genes to be expressed must also be
necessary for bradyzoites to form and encyst since Drps13 does not
persist in brain in vivo in the presence of a competent immune
response.
It would be of interest to determine whether a similar in vitro
persistent, G1 arrested phenotype in a ‘‘stressed’’ parasite is
common to other parasites that are attenuated and proving to be
immunogenic and protective. Examples of such parasites are those
attenuated by knockout of genes encoding components of nutrient
pathways and other unknown means such as in temperature
sensitive mutant 4. The robustness of protection conferred by
Drps13 combined with its lack of persistence in vivo, and inability to
induce growth of the parasite with ATc or L-NAME after the first
week, may provide insights into establishing protection by other
means. These results suggest that a similar persistent exposure to
immunogenic epitopes for several days might be useful in
development of component vaccines as well. This conditional
knockdown parasite appears to be sufficient to stimulate the
immune system without establishing a chronic infection (Figure 9).
Therefore it may be useful for vaccine development, including
definition of epitopes and immune responses essential for
protection.
Conclusions and SignificanceThe studies herein address regulation of transcription of a RP
and characterize effects of conditional knockdown of this RP.
Promoter analysis included identification of both a transcription
factor binding site in the promoter of RPS13 and associated
molecules. Modulation of the transcription factors that regulate
RPs would be expected to lead to subsequent imbalance of RPs. If
the regulatory mechanisms present in yeast and mammals also are
used by T. gondii, disruption of critical RPs would lead to
abrogation of effect of an MDM2-like molecule, arrest in the cell
cycle in G1, and hence contribute to diminution of proliferation
and persistence. The work described herein, identifies mechanisms
and suggests molecular switches associated with proliferation and
persistence of T. gondii in vitro. Conditional knockdown of rps13
which disrupts ribosomes is associated with arrest in G1 of the cell
cycle. The molecular pathways in this process are molecular
targets for development of antimicrobial agents to eradicate T.
gondii. The method developed herein for conditional knockdown
can be used for validation of other targets. Immunization with
Drps13 parasites protects mice completely against subsequent
challenge with wildtype clonal Type 1 parasites, and robustly
protects mice against wildtype clonal Type 2 parasites. Thus, the
conditional knockdown parasites have potential to be useful in
vaccine development.
Methods
Ethics StatementAll animal studies were approved by the University of Chicago
IACUC (#71734) and conducted according to AAALAC and
USDA guidelines (Animal Welfare Assurance #A3523-01).
Methods are presented in detail in Text S1. They are as
described in the following references: for modeling of RPS13
promoter analysis and EMSAs [63], and mass spectrometry
[23,75–78], ChIP [29,52], creation of RPL22 construct [44,45],
determining expression of RPs in tachyzoite and pH switch
conditions [22], homologous recombination [64], creation of
constructs with four TetOs in an optimal site in the RPS13
promoter and YFP-TetR constructs [20,21], culture of parasites
[65,66], creation of stable transfectants with the YFP-TetR
[20,21], qPCR [67], western blots, IFA [68], tissue culture of T.
gondii and assays that measure uracil uptake [65,66], analysis of G1
arrest [22,46,47,69], transcriptomes [22] use of L-NAME [48],
and immunization and evaluations of immunized mice [70].
Supporting Information
Figure S1 (A) TRP1 and TRP2 EMSA with radio labeled TRP1
and TRP2 probes with (+) and without (2) cold competitor TRP1
or TRP2 with tachyzoite nuclear extract. Note mobility-shift
which indicates that there is protein in the nuclear extract that
binds to TRP2, which is competed by cold TRP2. TRP2 arrow
indicates level in the gel where this occurs. (B) EMSAs with radio
labeled TRP1 [left lanes] and TRP2 [right lanes] with nuclear
extracts from tachyzoites competed by cold probe second lane,
and with antibody to MYST A [third lane] and MYST B [fourth
lane]. Some of the differences are highlighted by inclusion in green
rectangles. Bands are marked by letters a to g so they can be
identified in each of the corresponding lanes. In S1A * indicates a
nonspecific band that is not competed by cold probe. S1A shows
part of the EMSA beginning with band A. In S1A and S1B, bands
of similar mobility for TRP1 and TRP2 are not necessarily
binding the same nuclear extract proteins. Bands are indicated
with letters to indicate corresponding bands between figures 1A,
S1A, and S1B in the individual TRP1 and TRP2 elements. All
EMSA results shown were reproducible in at least two replicate
experiments.
Found at: doi:10.1371/journal.pone.0014057.s001 (2.62 MB
TIF)
Figure S2 Creation of conditional mutant parasite and replica-
tion in vitro +ATc. (A) Creation of parasites. These parasites were
created as described previously for parasites with one TetO in the
rps13 promoter [21]. As shown, to create the Drps13 conditional
knockdown, genomic integration of four TetO elements in the
rps13 locus was accomplished with a hit-and-run mutagenesis
strategy [21,64]. Alternative possible cross-over events could have
occurred between a construct containing in sequence
(TCCCCGACAACACCTTCTAC) and native T. gondii genomic
DNA resulting in different pseudodiploid conformations [20,21].
The constructs were previously described [21]. Pseudodiploid
generation occurred creating a pseudodiploid parasite, as
demonstrated by the colored diagram organization [21] with
primer locations marked. Sequences from the construct in the
diagram are indicated within brackets. The diagram, top, is
adapted from Figure 2A in reference 21 with permission. Other
symbols in this diagram include: orange box with an X represents
four TetOs; blue line represents bluescript vector backbone;
prps13 represents RPS13 promoter; pDHFR represents DHFR
promoter; DHFR is the DHFR coding region; thick arrows
T. gondii rps13
PLoS ONE | www.plosone.org 13 November 2010 | Volume 5 | Issue 11 | e14057

represent continuation of rps13 gene. Crossovers with the
construct and native gene occurred within the rps13 promoter
(prps13) as shown in the schematic diagram top, right. An
alternate pseudodiploid that did not occur could have formed with
a crossover within the rps13 intron. PCR using primers 3
(GTCGAGTCCTGTAGGTTCATC) and 10 (GGAGATCTC-
TATCACTGATAGGGA) on DNA isolated from mycophenolic
acid-xanthine-resistant clones showed that only one clone, B28,
had the replacement construct integrated at the rps13 locus (Figure
S2A top, left). PCR with primers 3 and 10 yielded a product of the
correct size to include the rps13 gene promoter and four TetOs.
Primers 9 (TCCCTATCAGTATAGAGATCTCC) and 4 did not
amplify products (data not shown). PCR with DNA from 6-
thioxanthine-resistant clones using both HXGPRT primers
(Figure S2A middle) as well as primers 3 and 4 (CTCCGAAG-
GAGTCTCTCAGTG) (Figure S2A bottom left panel) show that
pseudodiploidy has been lost for clones 1, 6, 12, 15, and 18. PCR
using primers 3 and 10 (Figure S2A bottom right panel) show that
clones 1, 6, 12, 15, and 18 retained TetO elements in the rps13
promoter. Parasites, e.g., chloramphenicol-resistant clones 3 and
9, express YFP (data not shown). (B) Comparison of uracil uptake
in Drps13 mutant parasites and parental strains +ATc. Uracil
uptake in the mutant parasite compared to wildtype or parental
strain was similar. There was no diminution of uptake in
Drps+ATc. Parental and conditional mutant knockdown parasites
are shown.
Found at: doi:10.1371/journal.pone.0014057.s002 (0.61 MB TIF)
Figure S3 Expression of RPS13 and RPL22. (A) Western blot of
cultures 6ATc at 4 and 48 hours probed with aRPS13 and
aSAG1. (B) IFA at 24 hours and 4 days probed of cultures 6ATc
with aMyc to detect Myc-tagged RPL22.
Found at: doi:10.1371/journal.pone.0014057.s003 (2.22 MB TIF)
Table S1 Mass Spectrometry.
Found at: doi:10.1371/journal.pone.0014057.s004 (0.07 MB
XLS)
Table S2 6ATc Transcriptome.
Found at: doi:10.1371/journal.pone.0014057.s005 (0.29 MB
XLS)
Text S1
Found at: doi:10.1371/journal.pone.0014057.s006 (0.22 MB
DOC)
Movie S1 RPS13.
Found at: doi:10.1371/journal.pone.0014057.s007 (3.65 MB
MOV)
Acknowledgments
We thank Michael White for his contributions to this paper. We thank
Dick Schaap, Nicole van Poppel, Jelle Welagen and Intervet for providing
the constructs pDHFRHXGPRT/rps13subTetO(IV)-23, YFP-TetR con-
struct, aRPS13, and control antibodies, reference [21], and helpful
discussions. We thank Micah Bhatti for assisting with Pfam analysis of
transcriptomes and William J. Sullivan, Nathalie Vonlaufen, and
Arunasalam Naguleswaran for providing aMYST A and B and performing
ChIP. We thank Louis Weiss for aBAG1 antibody and Laura Knoll for
aSRS9/BRS4 antibody. We thank David Morse for the RPL22 construct.
We thank Kristen Wroblewski for assistance with statistical analyses. We
gratefully acknowledge the special encouragement in these studies from R.
Blackfoot, R. Thewind, A. Akfortseven, S. Gemma, S. Jackson, and A.K.
Bump. We thank Joseph McCammon and Matthias Dean-Carpentier for
their assistance with preparation of the manuscript. We thank Louis Weiss,
Albert Einstein University, and Craig Roberts, Strathclyde University, for
reading this manuscript and their helpful suggestions.
Author Contributions
Conceived and designed the experiments: SLH EM KK WHW MSB KEB
SPM RM. Performed the experiments: SLH EM KK WHW MSB KEB
SPM BAR SRL RW YO AS JRY RM. Analyzed the data: SLH EM KK
WHW MSB SPM BAR SRL RW RM. Contributed reagents/materials/
analysis tools: MSB RM. Wrote the paper: SLH WHW MSB KEB YO AS
RM. Approved final manuscript: RM SLH EM KK WHW MSB KEB
SPM BAR SRL RW YO AS JRY.
References
1. Ferreira-Cerca S, Poll G, Kuhn H, Neueder A, Jakob S, et al. (2007) Analysis of
the in vivo assembly pathway of eukaryotic 40S ribosomal proteins. Molecular
Cell 28: 446–457.
2. Ferreira-Cerca S, Poll G, Gleizes PE, Tschochner H, Milkereit P (2005) Roles of
eukaryotic ribosomal proteins in maturation and transport of pre-18S rRNA and
ribosome function. Molecular Cell 20: 263–275.
3. Ferreira-Cerca S, Hurt E (2009) Cell biology: Arrest by ribosome. Nature 459:
46–47.
4. Blaha G, Stanley RE, Steitz TA (2009) Formation of the first peptide bond: the
structure of EF-P bound to the 70S ribosome. Science 325: 966–970.
5. Schmeing TM, Voorhees RM, Kelley AC, Gao YG, Murphy FVt, et al. (2009)
The crystal structure of the ribosome bound to EF-Tu and aminoacyl-tRNA.
Science 326: 688–694.
6. Schmeing TM, Ramakrishnan V (2009) What recent ribosome structures have
revealed about the mechanism of translation. Nature 461: 1234–1242.
7. Yonath A (2006) Molecular biology: triggering positive competition. Nature 444:
435–436.
8. Zhang W, Dunkle JA, Cate JH (2009) Structures of the ribosome in intermediate
states of ratcheting. Science 325: 1014–1017.
9. Opferman JT, Zambetti GP (2006) Translational research? Ribosome integrity
and a new p53 tumor suppressor checkpoint. Cell Death and Differentiation 13:
898–901.
10. Fumagalli S, Di Cara A, Neb-Gulati A, Natt F, Schwemberger S, et al. (2009)
Absence of nucleolar disruption after impairment of 40S ribosome biogenesis
reveals an rpL11-translation-dependent mechanism of p53 induction. Nat Cell
Biol 11: 501–508.
11. Beuvink I, Boulay A, Fumagalli S, Zilbermann F, Ruetz S, et al. (2005) The
mTOR inhibitor RAD001 sensitizes tumor cells to DNA-damaged induced
apoptosis through inhibition of p21 translation. Cell 120: 747–759.
12. Connolly LE, Cox JS (2009) CarD tricks and magic spots: mechanisms of
stringent control in mycobacteria. Cell Host Microbe 6: 1–2.
13. Trotter EW, Berenfeld L, Krause SA, Petsko GA, Gray JV (2001) Protein
misfolding and temperature up-shift cause G1 arrest via a common mechanism
dependent on heat shock factor in Saccharomycescerevisiae. Proc Natl Acad
Sci U S A 98: 7313–7318.
14. Rosado IV, Kressler D, de la Cruz J (2007) Functional analysis of
Saccharomyces cerevisiae ribosomal protein Rpl3p in ribosome synthesis.
Nucleic Acids Res 35: 4203–4213.
15. Nickels JT, Broach JR (1996) A ceramide-activated protein phosphatase mediates
ceramide-induced G1 arrest of Saccharomyces cerevisiae. Genes Dev 10: 382–394.
16. Tschochner H, Hurt E (2003) Pre-ribosomes on the road from the nucleolus to
the cytoplasm. Trends in Cell Biology 13: 255–263.
17. Ito T, Kim GT, Shinozaki K (2000) Disruption of an Arabidopsis cytoplasmic
ribosomal protein S13-homologous gene by transposon-mediated mutagenesis
causes aberrant growth and development. Plant J 22: 257–264.
18. Marygold SJ, Roote J, Reuter G, Lambertsson A, Ashburner M, et al. (2007)
The ribosomal protein genes and Minute loci of Drosophila melanogaster.
Genome Biol 8: R216.
19. Schaap D, Arts G, van Poppel NF, Vermeulen AN (2005) De novo ribosome
biosynthesis is transcriptionally regulated in Eimeria tenella, dependent on its life
cycle stage. Mol Biochem Parasitol 139: 239–248.
20. van Poppel NF, Welagen J, Duisters RF, Vermeulen AN, Schaap D (2006) Tight
control of transcription in Toxoplasma gondii using an alternative tet repressor.
International Journal for Parasitology 36: 443–452.
21. van Poppel NF (2005) Transcriptional regulation of ribosomal protein genes in
Toxoplasma gondii, a way towards conditionally attenuated parasites. Boxmeer
Intervet International. pp 110.
22. Behnke MS, Radke JB, Smith AT, Sullivan WJ, Jr., White MW (2008) The
transcription of bradyzoite genes in Toxoplasma gondii is controlled by
autonomous promoter elements. Mol Microbiol 68: 1502–1518.
23. Saksouk N, Bhatti MM, Kieffer S, Smith AT, Musset K, et al. (2005) Histone-
modifying complexes regulate gene expression pertinent to the differentiation of
the protozoan parasite Toxoplasma gondii. Mol Cell Biol 25: 10301–10314.
24. Sullivan WJ, Jr., Smith AT, Joyce BR (2009) Understanding mechanisms and
the role of differentiation in pathogenesis of Toxoplasma gondii: a review. Mem
Inst Oswaldo Cruz 104: 155–161.
T. gondii rps13
PLoS ONE | www.plosone.org 14 November 2010 | Volume 5 | Issue 11 | e14057

25. Smith AT, Tucker-Samaras SD, Fairlamb AH, Sullivan WJ, Jr. (2005) MYST
family histone acetyltransferases in the protozoan parasite Toxoplasma gondii.Eukaryot Cell 4: 2057–2065.
26. Vonlaufen N, Kanzok SM, Wek RC, Sullivan WJ, Jr. (2008) Stress response
pathways in protozoan parasites. Cell Microbiol 10: 2387–2399.
27. Reid JL, Iyer VR, Brown PO, Struhl K (2000) Coordinate regulation of yeast
ribosomal protein genes is associated with targeted recruitment of Esa1 histoneacetylase. Mol Cell 6: 1297–1307.
28. Lindner SE, De Silva EK, Keck JL, Llinas M (2009) Structural Determinants of
DNA Binding by a P. falciparum ApiAP2 Transcriptional Regulator. Journal ofMolecular Biology.
29. De Silva EK, Gehrke AR, Olszewski K, Leon I, Chahal JS, et al. (2008) Specific
DNA-binding by apicomplexan AP2 transcription factors. Proc Natl AcadSci U S A 105: 8393–8398.
30. Balaji S, Babu MM, Iyer LM, Aravind L (2005) Discovery of the principal
specific transcription factors of Apicomplexa and their implication for the
evolution of the AP2-integrase DNA binding domains. Nucleic Acids Res 33:3994–4006.
31. Koussevitzky S, Nott A, Mockler TC, Hong F, Sachetto-Martins G, et al. (2007)
Signals from chloroplasts converge to regulate nuclear gene expression. Science316: 715–719.
32. Yuda M, Iwanaga S, Shigenobu S, Mair GR, Janse CJ, et al. (2009)
Identification of a transcription factor in the mosquito-invasive stage of malaria
parasites. Molecular Microbiology 71: 1402–1414.
33. Diaz-Martin J, Almoguera C, Prieto-Dapena P, Espinosa JM, Jordano J (2005)
Functional interaction between two transcription factors involved in the
developmental regulation of a small heat stress protein gene promoter. PlantPhysiology 139: 1483–1494.
34. Iyer LM, Anantharaman V, Wolf MY, Aravind L (2008) Comparative genomics
of transcription factors and chromatin proteins in parasitic protists and othereukaryotes. International Journal for Parasitology 38: 1–31.
35. Howitt CA, Wilinski D, Llinas M, Templeton TJ, Dzikowski R, et al. (2009)
Clonally variant gene families in Plasmodium falciparum share a common
activation factor. Molecular Microbiology 73: 1171–1185.
36. Rushton PJ, Bokowiec MT, Han S, Zhang H, Brannock JF, et al. (2008)Tobacco transcription factors: novel insights into transcriptional regulation in
the Solanaceae. Plant Physiology 147: 280–295.
37. Grandbastien MA, Audeon C, Bonnivard E, Casacuberta JM, Chalhoub B,et al. (2005) Stress activation and genomic impact of Tnt1 retrotransposons in
Solanaceae. Cytogenet Genome Res 110: 229–241.
38. Deitsch K, Duraisingh M, Dzikowski R, Gunasekera A, Khan S, et al. (2007)Mechanisms of gene regulation in Plasmodium. American Journal of Tropical
Medicine and Hygiene 77: 201–208.
39. LaCount DJ, Vignali M, Chettier R, Phansalkar A, Bell R, et al. (2005) A
protein interaction network of the malaria parasite Plasmodium falciparum. Nature438: 103–107.
40. Flueck C, Bartfai R, Niederwieser I, Witmer K, Alako BT, et al. (2010) A Major
Role for the Plasmodium falciparum ApiAP2 Protein PfSIP2 in ChromosomeEnd Biology. PLoS Pathog 6: e1000784.
41. Stockinger EJ, Mao Y, Regier MK, Triezenberg SJ, Thomashow MF (2001)
Transcriptional adaptor and histone acetyltransferase proteins in Arabidopsisand their interactions with CBF1, a transcriptional activator involved in cold-
regulated gene expression. Nucleic Acids Res 29: 1524–1533.
42. Meissner M, Schluter D, Soldati D (2002) Role of Toxoplasma gondii myosin A
in powering parasite gliding and host cell invasion. Science 298: 837–840.
43. Gossen M, Bujard H (1993) Anhydrotetracycline, a novel effector fortetracycline controlled gene expression systems in eukaryotic cells. Nucleic
Acids Res 21: 4411–4412.
44. Sanz E, Yang L, Su T, Morris DR, McKnight GS, et al. (2009) Cell-type-specificisolation of ribosome-associated mRNA from complex tissues. Proceedings of the
National Academy of Sciences of the United States of America 106:
13939–13944.
45. Messina M, Niesman I, Mercier C, Sibley LD (1995) Stable DNAtransformation of Toxoplasma gondii using phleomycin selection. Gene 165:
213–217.
46. Radke JR, Striepen B, Guerini MN, Jerome ME, Roos DS, et al. (2001) Definingthe cell cycle for the tachyzoite stage of Toxoplasma gondii. Mol Biochem
Parasitol 115: 165–175.
47. Radke JR, White MW (1998) A cell cycle model for the tachyzoite ofToxoplasma gondii using the Herpes simplex virus thymidine kinase. Mol
Biochem Parasitol 94: 237–247.
48. Roberts F, Roberts CW, Ferguson DJ, McLeod R (2000) Inhibition of nitric
oxide production exacerbates chronic ocular toxoplasmosis. Parasite Immunol22: 1–5.
49. Van Poppel NF, Welagen J, Vermeulen AN, Schaap D (2006) The complete set
of Toxoplasma gondii ribosomal protein genes contains two conserved promoterelements. Parasitology 133: 19–31.
50. Mullapudi N, Joseph SJ, Kissinger JC (2009) Identification and functional
characterization of cis-regulatory elements in the apicomplexan parasiteToxoplasma gondii. Genome Biol 10: R34.
51. Moellering RE, Cornejo M, Davis TN, Del Bianco C, Aster JC, et al. (2009)
Direct inhibition of the NOTCH transcription factor complex. Nature 462:
182–188.
52. Vonlaufen N, Naguleswaran A, Coppens I, Sullivan WJ, Jr. (2010) MYST family
lysine acetyltransferase facilitates ataxia telangiectasia mutated (ATM) kinase-mediated DNA damage response in Toxoplasma gondii. Journal of Biological
Chemistry 285: 11154–11161.
53. Eaton MS, Weiss LM, Kim K (2006) Cyclic nucleotide kinases and tachyzoite-bradyzoite transition in Toxoplasma gondii. Int J Parasitol 36: 107–114.
54. McCourt PC, Morgan JM, Nickels JT, Jr. (2009) Stress-induced ceramide-activated protein phosphatase can compensate for loss of amphiphysin-like
activity in Saccharomyces cerevisiae and functions to reinitiate endocytosis.
Journal of Biological Chemistry 284: 11930–11941.55. Aronova S, Wedaman K, Aronov PA, Fontes K, Ramos K, et al. (2008)
Regulation of ceramide biosynthesis by TOR complex 2. Cell Metab 7:148–158.
56. Nishimura N, Hitomi K, Arvai AS, Rambo RP, Hitomi C, et al. (2009)Structural mechanism of abscisic acid binding and signaling by dimeric PYR1.
Science 326: 1373–1379.
57. Nagamune K, Hicks LM, Fux B, Brossier F, Chini EN, et al. (2008) Abscisic acidcontrols calcium-dependent egress and development in Toxoplasma gondii.
Nature 451: 207–210.58. Cech TR (2000) Structural biology. The ribosome is a ribozyme. Science 289:
878–879.
59. Becker T, Bhushan S, Jarasch A, Armache JP, Funes S, et al. (2009) Structure ofmonomeric yeast and mammalian Sec61 complexes interacting with the
translating ribosome. Science 326: 1369–1373.60. Seidelt B, Innis CA, Wilson DN, Gartmann M, Armache JP, et al. (2009)
Structural insight into nascent polypeptide chain-mediated translational stalling.Science 326: 1412–1415.
61. Radke JR, Guerini MN, Jerome M, White MW (2003) A change in the
premitotic period of the cell cycle is associated with bradyzoite differentiation inToxoplasma gondii. Molecular and Biochemical Parasitology 131: 119–127.
62. Ball HL, Zhang B, Riches JJ, Gandhi R, Li J, et al. (2009) Shwachman-BodianDiamond syndrome is a multi-functional protein implicated in cellular stress
responses. Human Molecular Genetics 18: 3684–3695.
63. Kibe MK, Coppin A, Dendouga N, Oria G, Meurice E, et al. (2005)Transcriptional regulation of two stage-specifically expressed genes in the
protozoan parasite Toxoplasma gondii. Nucleic Acids Res 33: 1722–1736.64. Donald RG, Roos DS (1998) Gene knock-outs and allelic replacements in
Toxoplasma gondii: HXGPRT as a selectable marker for hit-and-runmutagenesis. Mol Biochem Parasitol 91: 295–305.
65. Mack DG, McLeod R (1984) New micromethod to study the effect of antimicrobial
agents on Toxoplasma gondii: comparison of sulfadoxine and sulfadiazineindividually and in combination with pyrimethamine and study of clindamycin,
metronidazole, and cyclosporin A. Antimicrob Agents Chemother 26: 26–30.66. Mui EJ, Schiehser GA, Milhous WK, Hsu H, Roberts CW, et al. (2008) Novel
Triazine JPC-2067-B Inhibits Toxoplasma gondii In Vitro and In Vivo. PLoS
Negl Trop Dis 2: e190.67. Kasper DC, Sadeghi K, Prusa AR, Reischer GH, Kratochwill K, et al. (2009)
Quantitative real-time polymerase chain reaction for the accurate detection ofToxoplasma gondii in amniotic fluid. Diagnostic Microbiology and Infectious
Disease 63: 10–15.68. Ferguson DJ, Henriquez FL, Kirisits MJ, Muench SP, Prigge ST, et al. (2005)
Maternal inheritance and stage-specific variation of the apicoplast in
Toxoplasma gondii during development in the intermediate and definitive host.Eukaryot Cell 4: 814–826.
69. Gubbels MJ, Lehmann M, Muthalagi M, Jerome ME, Brooks CF, et al. (2008)Forward genetic analysis of the apicomplexan cell division cycle in Toxoplasma
gondii. PLoS Pathog 4: e36.
70. McLeod R, Frenkel JK, Estes RG, Mack DG, Eisenhauer PB, et al. (1988)Subcutaneous and intestinal vaccination with tachyzoites of Toxoplasma gondii
and acquisition of immunity to peroral and congenital toxoplasma challenge.J Immunol 140: 1632–1637.
71. Saksouk N, Bhatti NM, Kieffer S, Smith AT, Musset K, et al. (2005) Histone-
modifying complexes regulate gene expression pertinent to the differentiatin ofthe protozoan parasite Toxoplasma gondii. Mol Cell Biol 25: 10301–10314.
72. Behnke MS, Wootton JC, Lehmaann MM, Radke JB, Lucas O, et al. (2010)Coordinated Progression through Two Subscriptomes Underlies the Tachyzoite Cycle
of Toxoplasma gondii. PLoS ONE 5(8): e12354. doi:10.1371/journal.pone.0012354.73. Bernstein KA, Bleichert F, Bean JM, Cross FR, Baserga SJ (2007) Ribosome
biogenesis is sensed at the Start cell cycle checkpoint. Mol Biol Cell 18: 953–964.
74. Jorgensen P, Rupes I, Sharom JR, Schneper L, Broach JR, et al. (2004) Adynamic transcriptional network communicates growth potential to ribosome
synthesis and critical cell size. Genes Dev 18: 2491–2505.75. Keller A, Nesvizhskii AI, Kolker E, Aebersold R (2002) Empirical statistical
model to estimate the accuracy of peptide identifications made by MS/MS and
database search. Anal Chem 74(20): 5383–92.76. Cociorva D, Tabb D, Yates J (2006) Validation of Tandem Mass Spectrometry
Database each Results using DTASelect. Current Protocols in Bioinformatics13.4.1-13.4.14, supplement 16.
77. Diop SB, Bertaux K, Vasanthi D, Sarkeshik A, Goirand B, et al. (2008) Reptinand Pontin function antagonistically with PcG and TrxG complexes to mediate
Hox gene control, EMBO Rep. Mar 9(3): 260–6.
78. Eng J, McCormack A, Yates J (1994) An Approach to Correlate Tandem MassSpectral Data of Peptides with Amino Acid Sequences in a Protein Database.
J Am Soc Mass Spectrom 5: 976–989.
T. gondii rps13
PLoS ONE | www.plosone.org 15 November 2010 | Volume 5 | Issue 11 | e14057