Embed Size (px)
Citation preview

Patronin induces anaphase B • Wang et al. S1
JCB
TH
E J
OU
RN
AL
OF
CE
LL
BIO
LO
GY
Supplemental material
Wang et al., http://www.jcb.org/cgi/content/full/jcb.201306001/DC1
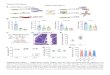
Figure S1. Effect of microinjection of anti-Pa-tronin or excessive cyclin B. (A) Specificity of the anti-KLP10A antibody (left) or anti-Patronin antibody (right) by Western blotting against Drosophila embryo extracts. M, protein mark-ers; HSS, high speed supernatant from Dro-sophila embryonic extract. (B) Spindle dynamics (green, tubulin) and chromosome segregation (red, histone) in a representative embryo injected with anti-Patronin antibody, showing that Patronin inhibition led to a gradi-ent suppression of anaphase B spindle elon-gation. Time is given in seconds from NEB, 14 s after the first recorded image after the micro-injection of the Patronin antibody. The injec-tion site is shown with an arrow in the first frame. Compared with wild-type (WT) spin-dles, Patronin inhibition abolished (spindle 1 [s1] and spindle 2 [s2]) or slowed down (spin-dle 3 [s3]) anaphase B spindle elongation. The spindle s4 shows spindle assembly de-fects after Patronin inhibition. Experiment was repeated in >25 embryos. See also Fig. 2 (A and B). (C) Quantification of phenotypes after Patronin inhibition. (top) For spindle length defects, four or five spindles near the injection site were analyzed for each embryo. (bottom) Chromosome segregation defects were further quantified in spindles that show no evident anaphase B spindle elongation. Note that in most cases, the chromosomes remain linked at the spindle midzone. (D) Spindle (red, tubulin) and kinetochore (green, GFP-cid) dynamics after Patronin inhibition. Time is given in sec-onds from the first frame, 260 s after the first recorded image after microinjection of Patro-nin antibody. (E and F) Microinjection of ex-cessive cyclin B–arrested spindles in metaphase or anaphase A (preanaphase B). It has been reported that the presence of exces-sive or nondegradable cyclin B arrested Dro-sophila embryo mitotic spindles in preanaphase B (Parry and O’Farrell, 2001; Cheerambathur et al., 2007). We injected ex-cessive purified GST–cyclin B into Drosophila embryos. As expected, the cyclin B injection disrupted the synchrony of embryonic mitosis and caused a gradient of phenotypes with spindles arrested between metaphase and anaphase A. By controlling the dose of cyclin B injection, we reproducibly arrested spindles in metaphase or anaphase A without ana-phase B spindle elongation. A high dose of cyclin B injection (intracellular concentration: 0.7 mg/ml) persistently arrested spindles in metaphase for over 20 min (E). A lower dose of cyclin B (intracellular concentration: 0.3 mg/ml) persistently arrested the spindles in anaphase A, that is, at a time point where anaphase A chromosome to pole movement had started but anaphase B spindle elongation had not (F). We should note that the anaphase A chromosome to pole movement was usually not normal in the presence of excessive cyclin B, as previously described (Parry et al., 2003). Green, GFP-tubulin; red, RFP-histone. 0 s shows the first recorded image after microinjection of cyclin B.

JCB S2
Figure S2. KLP10A inhibition led to ana-phase B spindle elongation in preanaphase B–arrested spindles, independent of cortical forces and robust midzone assembly. (A) Spin-dle dynamics (green, tubulin) and chromo-some segregation (red, histone) in a representative embryo sequentially injected (inj) with cyclin B and the anti-KLP10A anti-body. Time is given in seconds from NEB. The first micrograph shows the first recorded image after the injection of cyclin B (12 s after NEB). The left three time-lapse micro-graphs (12–487 s) show that cyclin B injec-tion arrested the spindles in anaphase A. The anti-KLP10A antibody was then injected into the same site between 487 and 552 s, and the right two micrographs (552 and 591 s) show that injection of the anti-KLP10A anti-body led to anaphase B–like spindle elonga-tion, which further segregated the chromosomes. The pole–pole distance as a function of time from NEB (0 s; blue, cyclin B [cyc B] + anti-KLP10A [anti-10A]) is compared with an untreated embryo (black, wild type [WT]) and a representative embryo injected with only cyclin B (cyc B only; gray), showing that KLP10A inhibition initiates anaphase B–like spindle elongation in cyclin B–induced anaphase A–arrested spindles. Cyclin B was injected before NEB, and anti-KLP10A was in-jected at the indicated time. Error bars are SDs. See also Fig. 4. (B). Pole–pole and chro-mosome to pole distances as a function of time for the spindles shown in Fig. 4 D. The chromosome–chromosome (chr-chr) distances were measured between the leading edges of the two sets of chromosomes, and the chromo-some–pole distances were measured between a chromosome leading edge and the nearest pole. Time (0 s) starts from the first frame after anti-KLP10A inhibition. The experiment was repeated in >25 embryos. (C) Spindle elonga-tion rates induced by KLP10A inhibition in cy-clin B–arrested scrambled mutants (16 spindles in 5 embryos) are not significantly different from those in cyclin B–arrested wild-type em-bryos (61 spindles in 26 embryos, P = 0.7969), suggesting that cortical forces play no significant role in the spindle elongation in-duced by KLP10A inhibition. Error bars are SDs. (D) EB1-GFP localization in wild-type and cyclin B/anti-KLP10A–injected embryos. Tubulin (red) and EB1 (green) are shown in
merged images. The bottom shows relative fluorescent intensity line scans of EB1-GFP (blue) and tubulin (gray) along the pole–pole axis. The 0 position shows the spindle equator. In wild-type embryos, EB1-GFP concentrates at the spindle midzone during anaphase B spindle elongation. However, in em-bryos injected with cyclin B and anti-KLP10A, EB1-GFP remains all along MTs during spindle elongation, showing that the spindle elongation induced by KLP10A inhibition is independent of EB1-GFP redistribution. The experiment was repeated in >20 embryos. Bar, 5 µm. (E) Feo-GFP localization in wild-type and injected embryos. In wild-type embryos, Feo, a Drosophila homologue of the MT cross-linker Ase1/PRC1 (Vernì et al., 2004), accumulated at the spin-dle midzone during anaphase B spindle elongation. In our sequential injection experiment, we did not observe accumulation of Feo-GFP at the spindle mid-zone during elongation, showing that the spindle elongation induced by KLP10A inhibition was independent of Feo recruitment to the spindle midzone. Tubulin (red) and Feo (green) are shown in merged images.

Patronin induces anaphase B • Wang et al. S3
Figure S3. Dynamics of Patronin and in silico modeling for the anaphase B switch. (A) FRAP of Patronin-GFP. Example of a Patronin-GFP bleach near the pole. Bar, 5 µm. (B) Corrected and normalized fluorescence for the example shown in A. Fluorescence recovers fully and rapidly. Experiments were re-peated >30 times for each condition. (C) Cartoon of the computational model. The diagram shows three parallel MTs (blue), representing the MTs in an ipMT bundle in the left half-spindle, organized with their plus ends toward the equator (right) and their minus ends away from the equator (left), and a fixed number of KLP10A and Patronin molecules that diffuse freely in the one-dimensional space and bind and dissociate from the MTs at rates kon and koff. The MTs in the model are assumed to slide polewards and polymerize at their plus ends at the sliding rate, effectively giving rise to a fixed position of the MT plus end in the laboratory frame of reference. MT minus ends depolymerize at a rate determined by the number of “active” KLP10A molecules bound, which is calculated as the difference between the number of KLP10A and active inhibiting Patronin bound to the MT tip and is limited by 0 at the lower end. (D) Model result: Pole position as a function of time in wild-type (blue) and Patronin-inhibited (red) embryos. A switch in the percentage of Patronin that can in-hibit the depolymerization activity of KLP10A from 10 to 90% (arrow) leads to movement of the pole away from the equator (at position 0). The experiment was repeated 15 times. (E) Spindle kymograph of Patronin (right) and merged with tubulin (left; red, tubulin; green, Patronin). The distribution of Patronin does not change at anaphase B onset. (F) Model Patronin kymograph: Patronin is uniform throughout preanaphase B and anaphase B as observed in vivo, compare with E. The experiment was repeated 15 times. (G) Example of in silico FRAP. The experiment was repeated 15 times.

JCB S4
Table S1. Model parameters
Parameter Description Range Reference
N_MT Number of MTs 1–15 In a single ipMT bundle; Sharp et al., 1999N_KLP Total number of KLP10A molecules 500–5,000 Assumed in this studyN_Pat Total number of Patronin molecules 500–5,000 Assumed in this study
konK and koff
K KLP10A-MT binding and unbinding rates 0.3 and 0.7 s1 Determined in this study to match in silico FRAP with experimental FRAP
konP and koff
P Patronin-MT binding and unbinding rates 0.15 and 0.9 s1 Determined in this study to match in silico FRAP with experimental FRAP
vdepδ MT minus end depolymerization rate
per number of KLP10A0.06 µm/s Brust-Mascher and Scholey, 2002
Lcr Critical region length at MT minus end for depolymerization rate
10 × 8 nm = 80 nm Assumed in this study
Number of bound KLP10A for vdep
δ 3–5 Assumed in this study
DK and DP Diffusion coefficient for KLP10A and Patronin within the spindle volume
3.2 and 3.4 µm2/s Estimated from measured D of GFP adjusted by size (unpublished data)
Fraction of active Patronin (inhibiting KLP10A) 0–1t Iteration time step 0.1 sNstep Total number of iterations per simulation 2,500x and M Subcompartment size and number 6 nm and 3,000
Video 1. Dynamics of KLP10A-GFP in a representative spindle during Drosophila embryo mitosis. A representative transgenic Drosophila embryo expressing KLP10A-GFP under the polyubiquitin promoter was imaged by time-lapse confocal microscopy. KLP10A-GFP was especially enriched at the centrosomes and spindle poles and also associated with kinetochores from pro-metaphase to anaphase A. In addition, a small but significant localization to cortical structures and spindle MTs was observed. Images were taken every 2.5 s on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer) for 10 min. Display rate is 10 frames/s. See Fig. 1 A for still images.
Video 2. Dynamics of Patronin-GFP in a representative spindle during Drosophila embryo mitosis. A representative transgenic Drosophila embryo expressing Patronin-GFP under the polyubiquitin promoter was imaged by time-lapse confocal microscopy. Patronin-GFP localized along the spindle throughout mitosis and outside the nucleus during interphase. Images were taken every 4.7 s for 7 min on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer). Display rate is 10 frames/s. See Fig. 1 B for still images.
Video 3. Spindle dynamics and chromosome segregation in a control embryo. A representative transgenic Drosophila embryo expressing GFP-tubulin (green) and RFP-histone (red) was imaged by time-lapse confocal microscopy. Image stacks were taken every 17 s for 6 min on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer). Each frame is the projection of nine planes spaced by 0.5 µm. Display rate is 5 frames/s. See Fig. 2 B (top) for still images.

Patronin induces anaphase B • Wang et al. S5
Video 4. Spindle dynamics and chromosome segregation after Patronin inhibition. A representative transgenic Drosophila embryo expressing GFP-tubulin (green) and RFP-histone (red) was injected with an anti-Patronin antibody (intracellular concen-tration of 0.3 mg/ml) at NEB onset and analyzed by time-lapse confocal microscopy. Image stacks were taken every 18 s for 9 min on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer). Each frame is the projection of nine planes spaced by 0.5 µm. Display rate is 5 frames/s. See Fig. 2 B (bottom) for still images.
Video 5. KLP10A inhibition caused anaphase B–like elongation of metaphase-arrested spindles. A representative transgenic Drosophila embryo expressing GFP-tubulin (green) and RFP-histone (red) was imaged by time-lapse confocal microscopy. Before imaging, the embryo was injected with a high dose of cyclin B (intracellular concentration of 0.7 mg/ml). After the high dose cyclin B injection arrested the spindles in metaphase for around 10 min, the embryo was injected with an anti-KLP10A antibody (intracellular concentration of 0.5 mg/ml; KLP10A inhibition) and returned to imaging. This KLP10A inhibition led to anaphase B–like elongation of the metaphase-arrested spindles, with the chromosomes at the spindle equator. Images were taken every 3 s on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer) for a total time of 17 min. Display rate is 10 frames/s. The arrow in the first frame indicates the measured spindle in Fig. 4 A (right).
Video 6. KLP10A inhibition led to anaphase B–like elongation of the anaphase A–arrested spindle. A representative trans-genic Drosophila embryo expressing GFP-tubulin (green) and RFP-histone (red) was imaged by time-lapse confocal microscopy. Before imaging, the embryo was injected with a low dose of cyclin B (intracellular concentration of 0.3 mg/ml), and spindles were arrested in metaphase for a prolonged time and then started to proceed into anaphase A. Then, the embryo was injected with an anti-KLP10A antibody (intracellular concentration of 0.5 mg/ml; KLP10A inhibition) at anaphase A onset and returned to imaging. This KLP10A inhibition led to anaphase B–like elongation of the anaphase A–arrested spindles, which would oth-erwise show no spindle elongation. Images were taken every 3 s on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer) for a total time of 14 min. Display rate is 10 frames/s. The arrows in the first frame indicate the spindles that were measured for Fig. 4 E. The top spindle is also shown in Fig. 4 B (right).
Video 7. KLP10A inhibition led to anaphase B–like spindle elongation, which further segregated chromosomes. A represen-tative transgenic Drosophila embryo expressing GFP-tubulin (green) and RFP-histone (red) was imaged by time-lapse confo-cal microscopy. Before imaging, the embryo was injected with a low dose of cyclin B (intracellular concentration of 0.3 mg/ml), and spindles were arrested in metaphase for a prolonged time and then started to proceed into anaphase A. Then, the embryo was injected with an anti-KLP10A antibody (intracellular concentration of 0.5 mg/ml; KLP10A inhibition) after anaphase A started and returned to imaging. This KLP10A inhibition led to anaphase B–like elongation that further segregated chromosomes, which would otherwise show no spindle elongation. Images were taken every 3 s on a confocal microscope (IX70; Olympus) with spinning-disk confocal head (PerkinElmer) for a total time of 11 min. Display rate is 10 frames/s. See also Fig. S2 A.
ReferencesBrust-Mascher, I., and J.M. Scholey. 2002. Microtubule flux and sliding in mitotic spindles of Drosophila embryos. Mol. Biol. Cell. 13:3967–3975. http://dx.doi.
org/10.1091/mbc.02-05-0069
Cheerambathur, D.K., G. Civelekoglu-Scholey, I. Brust-Mascher, P. Sommi, A. Mogilner, and J.M. Scholey. 2007. Quantitative analysis of an anaphase B switch: pre-dicted role for a microtubule catastrophe gradient. J. Cell Biol. 177:995–1004. http://dx.doi.org/10.1083/jcb.200611113
Parry, D.H., and P.H. O’Farrell. 2001. The schedule of destruction of three mitotic cyclins can dictate the timing of events during exit from mitosis. Curr. Biol. 11:671–683. http://dx.doi.org/10.1016/S0960-9822(01)00204-4
Parry, D.H., G.R. Hickson, and P.H. O’Farrell. 2003. Cyclin B destruction triggers changes in kinetochore behavior essential for successful anaphase. Curr. Biol. 13:647–653. http://dx.doi.org/10.1016/S0960-9822(03)00242-2
Sharp, D.J., K.L. McDonald, H.M. Brown, H.J. Matthies, C. Walczak, R.D. Vale, T.J. Mitchison, and J.M. Scholey. 1999. The bipolar kinesin, KLP61F, cross-links mi-crotubules within interpolar microtubule bundles of Drosophila embryonic mitotic spindles. J. Cell Biol. 144:125–138. http://dx.doi.org/10.1083/jcb.144.1.125
Vernì, F., M.P. Somma, K.C. Gunsalus, S. Bonaccorsi, G. Belloni, M.L. Goldberg, and M. Gatti. 2004. Feo, the Drosophila homolog of PRC1, is required for central-spindle formation and cytokinesis. Curr. Biol. 14:1569–1575. http://dx.doi.org/10.1016/j.cub.2004.08.054