Embed Size (px)
Citation preview

doi:10.1152/ajpgi.00295.2006 292:98-112, 2007. First published Aug 24, 2006;Am J Physiol Gastrointest Liver Physiol
Sungwon Choi, Mike Lee, Amy L. Shiu, Sek Jin Yo and Gregory W. Aponte
You might find this additional information useful...
65 articles, 35 of which you can access free at: This article cites http://ajpgi.physiology.org/cgi/content/full/292/1/G98#BIBL
2 other HighWire hosted articles: This article has been cited by
[PDF] [Full Text] [Abstract]
, November 10, 2006; 314 (5801): 982-985. ScienceG. Kirillov, Z. Guo, S. Lyle, E. K. Ginter and E. I. Rogaev A. Kazantseva, A. Goltsov, R. Zinchenko, A. P. Grigorenko, A. V. Abrukova, Y. K. Moliaka, A.
LIPH.Human Hair Growth Deficiency Is Linked to a Genetic Defect in the Phospholipase Gene
[PDF] [Full Text] [Abstract], May 1, 2007; 292 (5): G1366-G1375. Am J Physiol Gastrointest Liver Physiol
S. Choi, M. Lee, A. L. Shiu, S. J. Yo, G. Hallden and G. W. Aponte STC-1 cells
GPR93 activation by protein hydrolysate induces CCK transcription and secretion in
including high-resolution figures, can be found at: Updated information and services http://ajpgi.physiology.org/cgi/content/full/292/1/G98
can be found at: AJP - Gastrointestinal and Liver Physiologyabout Additional material and information http://www.the-aps.org/publications/ajpgi
This information is current as of February 5, 2008 .
http://www.the-aps.org/.Society. ISSN: 0193-1857, ESSN: 1522-1547. Visit our website at American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the American Physiologicalabnormal function of the gastrointestinal tract, hepatobiliary system, and pancreas. It is published 12 times a year (monthly) by the
publishes original articles pertaining to all aspects of research involving normal orAJP - Gastrointestinal and Liver Physiology
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

Identification of a protein hydrolysate responsive G protein-coupled receptorin enterocytes
Sungwon Choi, Mike Lee, Amy L. Shiu, Sek Jin Yo, and Gregory W. AponteDepartment of Nutritional Sciences and Toxicology, University of California at Berkeley, Berkeley, California
Submitted 5 July 2006; accepted in final form 21 August 2006
Choi S, Lee M, Shiu AL, Yo SJ, Aponte GW. Identification of aprotein hydrolysate responsive G protein-coupled receptor in enterocytes.Am J Physiol Gastrointest Liver Physiol 292: G98–G112, 2007. Firstpublished August 24, 2006; doi:10.1152/ajpgi.00295.2006.—G protein-coupled receptors (GPCRs) have the potential to play a role as molecularsensors responsive to luminal dietary contents. Although such a role forGPCRs has been implicated in the intestinal response to protein hydro-lysate, no GPCR directly involved in this process has been previouslyidentified. In the present study, for the first time, we identified GPR93expression in enterocytes and demonstrated its activation in these cells byprotein hydrolysate with EC50 of 10.6 mg/ml as determined by theinduction of intracellular free Ca2�. In enterocytes, GPR93 was syner-gistically activated by protein hydrolysate in combination with an agonist,oleoyl-L-�-lysophosphatidic acid (LPA), which activated the receptor inthese enterocytes with EC50 of 7.9 nM. The increased intracellular Ca2�
by GPR93 activation was observed without the addition of a promiscuousG� protein and was pertussis toxin sensitive, which suggests G�q- andG�i-mediated pathways. Activated GPR93 also induced pertussis toxin-sensitive ERK1/2 phosphorylation. Both nuclear factor of activated Tcells and 12-O-tetradecanoylphorbol 13-acetate responsive elements re-porter activities were induced by protein hydrolysate in cells exogenouslyexpressing GPR93. The peptidomimetic cefaclor by itself did not activateGPR93 but potentiated the protein hydrolysate response and furtheramplified the synergistic enhancement of GPR93 activation by proteinhydrolysate and LPA. These data suggest that, physiologically, thecomposition of stimuli might determine GPR93 activity or its sensitivitytoward a given activator and suggest a new mechanism of the regulationof mucosal cell proliferation and differentiation and hormonal secretionby dietary products in the lumen.
extracellular signal-regulated protein kinase 1/2; intestine; GPR92;GPR93; lysophosphatidic acid
INTESTINAL MUCOSAL HOMEOSTASIS necessitates the coordinationof physiological and chemical factors that regulate functionsfrom cellular renewal and differentiation along the crypt-to-villus axis to the regulation of hormone secretion, immuneresponse, and nutrient assimilation along the proximal-to-distalintestine. This coordination is partly facilitated by cellularfactors that initiate intracellular signals in response to theluminal content. The importance of the presence of luminaldietary nutrients is well exemplified in patients undergoingtotal parenteral nutrition (TPN). Chronic TPN resulting in adramatic mucosal remodeling can lead to a compromisedabsorptive capacity and intestinal immune function and thedevelopment of intestinal and liver diseases.
Studies investigating nutrient-induced signaling and generegulation have mostly centered on the effects of fatty acidsand glucose on the expression of proteins involved in transportand metabolism in adipocytes and pancreatic �-cells, respec-
tively (15, 19, 33, 52, 65). Signaling events initiated by luminalprotein hydrolysate have been a focus of studies for definingthe mechanisms leading to the release of CCK (48, 49). In theenteroendocrine STC-1 cells, protein hydrolysate activatesERK1/2, CaMK pathways, as well as the PKA pathways (19).Several studies have demonstrated that the uptake of proteinhydrolysate in the intestine is through the proton-coupledoligopeptide transporter PepT1 (1, 13, 14). Oligopeptides up-take through this transporter is linked to events that subse-quently lead to an induced transcription of CCK (19) and therelease of CCK (14). There are numerous examples of nutrient-sensing events in the lumen that may be mediated by trans-porters or receptors. The indirect involvement of G protein-coupled receptors (GPCRs) in these sensing events in whichthe release of their ligands is directly induced by luminalnutrients is well characterized. For example, free fatty acidsdirectly induce the release of peptide YY (3), which in turnactivates neuropeptide Y (NPY) receptors, or protein hydroly-sate induces the release of CCK (12, 42, 48), which in turnactivates the receptor CCK1R (63). The role of GPCRs assensors that can directly respond to changes in luminal contentsis not well defined. The characteristics of their seven trans-membrane configuration not only allow for a given GPCR torecognize a wide range of molecular structures but also activatemultiple pathways depending on the conditions of the stimuli.
Some GPCRs have been reported to be directly activated bybasic L-amino acids such as the Ca2�-sensing receptor CaR (9);the GPRC6A, which has wide tissue distribution outside theintestine (61); and the heterodimeric taste receptor T1R1/T1R3
(47). The identification of G� gustducin and transducin, whichassociate with taste receptors, in the gastrointestinal (GI) mu-cosa as well as the presence of T2R in the GI tract suggests thisfamily of GPCRs can act as sensors that may respond toluminal contents such amino acids and toxins (27, 66).
Using the enterocyte-like hybrid Berkeley Rat IntestineEpithelial 380 (hBRIE 380i) cells, which do not expressPepT1, we explored the possibility that signaling cascades inthe enterocytes initiated by protein hydrolysate could be di-rectly mediated by the activation of a GPCR. In the presentstudy, we identified for the first time a GPCR, GPR93, in therat enterocytes that is directly responsive to protein hydroly-sate. GPR93 activation by protein hydrolysate mobilized intra-cellular Ca2� concentration ([Ca2�]i) and activated ERK1/2through both G�q- and G�i-mediated pathways. Protein hydro-lysate and oleoyl-L-�-lysophosphatidic acid (LPA) synergisti-cally activated GPR93. Our data suggest that GPR93 could bepartly responsible for luminal protein hydrolysate-inducedERK1/2 activation and for the subsequent effects of this
Address for reprint requests and other correspondence: G. W. Aponte, Univ.of California, Dept. of Nutritional Sciences and Toxicology, 119 Morgan Hall,Berkeley, CA 94720-3104 (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Gastrointest Liver Physiol 292: G98–G112, 2007.First published August 24, 2006; doi:10.1152/ajpgi.00295.2006.
0193-1857/07 $8.00 Copyright © 2007 the American Physiological Society http://www.ajpgi.orgG98
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

activation, such as alterations in cell proliferation and differ-entiation.
MATERIALS AND METHODS
Materials. All reagents, including meat protein hydrolysate(peptone) type I and LPA, which were used for GPR93 char-acterization in the entire study, were purchased from Sigma-Aldrich, unless indicated differently.
Cell culture condition and transfection. The hBRIE 380icells used in this study were from a well-characterized sub-clone expressing enterocyte phenotypes and protein markers(2, 24, 25). Experiments were performed using hBRIE 380icells of the same passage number. The hBRIE 380i andChinese hamster ovary (CHO) cells were maintained inIscove’s modified Dulbecco’s medium (IMDM; Invitrogen)with 10% bovine calf serum (BCS; Hyclone), 100 U/ml pen-icillin, and 100 �g/ml streptomycin as additional supplementsat 37°C in 5% CO2-95% air. For experiments, cells were plated24 h before experiments and were at �80% confluency on theday of use. The hBRIE 380i cells used for Western blotanalysis were laid down in 35-mm tissue culture dishes (Corn-ing), coated with rat-tail collagen type I (2). For transfection,cells were trypsinized, resuspended in IMDM (7.5 � 106
cells/ml), and incubated with the plasmid DNA in a volumebetween 0.2 and 0.7 ml at room temperature for 5 min. Thetotal amount of DNA in each transfection was 8 �g/106 ofcells, unless otherwise noted. Electroporation was carried outin a 0.4-mm cuvette at 0.25 kV and 960 �F, using a GenePulser (Bio-Rad). One milliliter of IMDM-10% BCS wasadded to the cuvette immediately after electroporation. Thecells were then plated and allowed to recover for 20 h. ThehBRIE 380i cells, stably expressing mitochondria-targeted ae-quorin (mtAEQ), were prepared as follows: 36 h after electro-poration with 2 �g mtAEQ construct/106 hBRIE 380i cells,resistant clones were selected in the presence of 800 �g/mlG418 (Invitrogen). Expression of functional mtAEQ was ver-ified using [Ca2�]i mobilization assay.
Plasmid construction. The open reading frames of GPR93,G�15, G�q, NPY receptor subtype 1 (NPY1R), �2-adrenergicreceptor (�2AR), GPR103, P2Y5, P2Y9, and P2Y10 were PCRamplified using Pfu DNA polymerase (Stratagene) from ratintestine or brain cDNA. The sequences of oligonucleotideprimers (Integrated DNA Technology) and GenBank accessionnumbers are listed in Table 1. Each receptor cDNA productwas ligated into the multiple cloning site of pCI-neo expressionvector (Promega), and each G protein cDNA was ligated intothe multiple cloning site of pcDNA3.0 (Invitrogen). G�q
cDNA was used as a PCR template for the construction ofG��6qi5myr (10, 35, 36). GPR93 tagged at the COOH-terminuswith the enhanced green fluorescent protein (GPR93-EGFPfusion), was constructed by ligating the GPR93 open readingframe (its stop codon removed using PCR) at the BamHI site ofthe enhanced green fluorescent protein (EGFP) expressionvector (Clontech). The mtAEQ (53, 58), i.e., Ca2�-sensitiveAEQ cDNA (AY604000) with the first 3 bases replaced with a99-base fragment encoding the NH2-terminal 33 amino acidsof human cytochrome c oxidase subunit VIII (including the 25amino acid mitochondria signal sequence) (J04823), was con-structed using overlapping synthetic oligonucleotide primersand ligated into pcDNA3.0. TPA response element (TRE) and T
able
1.O
ligo
nucl
eoti
depr
imer
sus
edfo
rR
T-P
CR
anal
yses
and
cDN
Acl
onin
g
Gen
eA
cces
sion
Num
ber
Forw
ard
Prim
er(5
�to
3�)
Rev
erse
Prim
er(5
�to
3�)
RT
-PC
Ran
alys
isG
PR93
XM
_575
667
GC
TC
TG
CC
TG
GG
CG
TG
TG
GG
CT
CT
CA
TC
CT
GC
GC
GT
CG
GG
CC
TC
GC
CA
GT
GT
CC
AG
AA
GA
CC
PepT
1N
M_0
5712
1A
AG
AG
GA
AC
CA
CT
GG
CT
GG
AC
TG
GG
CT
AA
AG
GG
GA
GG
TG
AA
GT
TG
AA
AC
CA
CA
TT
TT
GC
cDN
Acl
onin
gP2
Y5
BC
0987
03T
GT
GA
CC
AC
AT
CT
GA
AT
AC
CA
AG
GC
GA
CC
GA
CC
GT
CC
GT
AG
GT
TA
AT
TT
TC
TG
TT
GG
AG
CP2
Y9
XM
_228
501
GT
CC
CT
AG
TC
TC
AA
GA
GT
GG
TG
AA
CC
CC
TG
CA
CT
GT
AG
CT
GA
AC
CT
GA
AA
GA
CA
AT
TC
CT
GP2
Y10
XM
_228
500
CT
GA
GA
TA
TC
CA
CC
AT
GG
GC
AG
CA
AT
AG
CA
CC
AC
AT
TA
TG
CT
TC
CT
TC
AT
CT
GT
GG
AG
AC
C�
GPR
93X
M_5
7566
7A
TG
TT
TG
CC
AA
TT
CT
TC
AG
CC
AA
CA
CC
TC
TT
CT
CA
GA
GG
GC
TG
AA
TC
TT
GG
GG
GC
CG
TT
GT
CC
GPR
103
NM
_198
199
AT
GC
AG
GC
GC
TC
AA
CA
TC
AC
CG
CG
GA
GC
AG
TT
CG
CT
TA
CA
GT
TC
AT
GG
CC
AC
TA
CC
AA
AA
GT
AG
AG
NPY
IRZ
I150
4G
CG
GA
TC
CA
CC
AT
GA
AC
TC
AA
CT
CT
GT
TC
TC
CA
GC
TT
CA
GA
TT
TT
TT
CA
TT
GT
CA
TT
CA
TA
CT
�2A
RB
C08
6538
AC
CC
GC
TG
AG
AG
CG
TC
TG
GG
CA
CT
GA
AA
GC
CC
GT
CT
TA
AA
GA
GT
AG
AA
AG
CC
TA
CA
CT
AC
AG
TG
GG
�q
NM
_031
036
TG
GC
CA
CC
AT
GA
CT
CT
GG
AG
TC
CA
TC
AT
GG
CG
TG
CT
GC
CT
GA
GC
CT
AG
CT
AG
CT
TA
GA
CC
AG
AT
TG
TA
CT
CC
TT
CA
GG
TT
CA
GC
TG
CA
GG
�15
NM
_053
542
CG
CC
CA
CC
AT
GG
CC
CG
GT
CC
CT
GA
CT
TG
GG
GC
TG
CT
GT
CC
CG
GT
TC
CC
CG
TG
TC
AC
AA
CA
GG
TT
GA
TC
TC
GT
CC
AG
GT
AC
CG
G�
�6qi5
my
AG
TC
CA
CC
AT
GG
GG
TG
CT
GC
CT
GA
GC
GA
GG
AG
GC
CA
AG
GC
TA
GC
TA
GC
TC
AG
AA
GA
GG
CC
AC
AG
TC
CT
TC
AG
GT
TC
AG
CT
GC
AG
Rat
geno
mic
sequ
ence
(NW
_048
043)
was
also
used
inde
sign
ing
the
prim
ers
for
P2Y
10.
NPY
1R,
neur
opep
tide
Yre
cept
orsu
btyp
e1;
�2A
R,
�2-a
dren
ergi
cre
cept
or.
G99PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

nuclear factor of activated T cells (NFAT) luciferase reporterconstructs are pBV-luc (a generous gift from Dr. Bert Vo-gelstein, The Johns Hopkins Kimmel Cancer Center) with 9�TPA response elements (TGACTAA) or 8� NFAT responseelements (GGAGGAAAAACTGTTTCATACAGAAGGCGT),respectively, inserted in its multiple cloning site. All constructswere verified by DNA sequencing (DNA Sequencing Facility,University of California at Berkeley).
Tissue preparation and RNA isolation for GPR93 expressionprofile. Overnight fasted male Sprague-Dawley rats (14 wkold) were used as tissues sources (n 4). The protocol foranimal use was reviewed and approved by the Animal Care andUse Committee of the University of California at Berkeley. GItissue samples were prepared as follows: intestines were ex-tracted and cut into segments (each �5 cm long). Residualluminal contents were removed by running ice-cold PBSthrough the intestinal segments. The mucosal layer of theintestine was obtained by gentle scraping of the exposedluminal surface. The muscle layer was obtained after furtherscraping to remove the residual mucosal layer. The stomachmucosa was prepared in a similar fashion. The purity of themucosa was verified by the relative expression of villin andintestinal fatty acid binding proteins (I-FABP), differentiationmarkers of intestinal enterocytes, as determined by RT-PCR.
Non-GI tissue samples were obtained by removing theorgans, followed by rinsing with ice-cold PBS twice andmincing with surgical scissors. Immediately, the tissue sampleswere immersed in ice-cold TRIzol (Invitrogen), homogenized,and frozen in liquid nitrogen. All tissue preparation steps weredone on ice. RNA was isolated from the TRIzol tissue homog-enate according to the manufacturer’s protocol.
Semiquantitative RT-PCR. Reverse transcription was as de-scribed previously (40). PCR was carried out using Taq DNApolymerase (New England Biolab). GPR93 specific primersamplified a cDNA fragment of 249 bp. PCR parameters forGPR93 were as follows: 20 s at 94°C, 15 s at 55°C, and 30 sat 72°C for 32 cycles. The primers and PCR condition forvillin, I-FABP, and 18S RNA were as previously described(40). Amplified cDNA fragments were analyzed by agarose gelelectrophoresis followed by densitometry. The specificity ofthe PCR products was confirmed by DNA sequencing.
Laser microscopy dissection. Rat duodenum tissue sectionswere prepared for cryostat as previously described (24).Briefly, the duodenum section was removed from overnightfasted male Sprague-Dawley rats (12 wk old), and the residualluminal contents were washed out by running ice-cold PBSthrough the duodenum segment. The duodenum was further cutinto 2-mm sections in the horizontal direction after shortfixation in 70% ethanol in PBS (pH 7.4). The tissue sectionswere briefly rinsed with ice-cold PBS and immersed in ice-cold30% (wt/vol) sucrose in PBS overnight at 4°C. The sucrose-equilibrated sections were embedded into optimum cuttingtemperature compound (TissueTeK), frozen on dry ice, cryo-sectioned at 10 �m thickness, and then stored at 80°C.
For the laser microscopy dissection (LMD) procedure, 10�m cryosections were mounted on slides, fixed with 70%ethanol for 30 s, and stained with eosin, followed by a 5-sdehydration step in each of 70%, 95%, and 100% ethanol.After brief air drying, the sections were laser microdissectedusing a Leica AS LMD system with the following setting:aperture, 6 –10; intensity, 45; and speed, 2–5. Total RNA
from laser-captured villus and crypt regions (15 patchesfrom the crypt and villus area, separately, were pooled) wasisolated using RNeasy Micro Kit (Qiagen). SemiquantitativeRT-PCR was then performed. The purity of harvested LMDsamples was confirmed by comparing the expression level ofvillin and I-FABP, differentiation markers of intestinal ep-ithelial cells.
Localization of GPR93 in CHO cells. CHO cells weretransfected with the GPR93-EGFP fusion construct by electro-poration (4 �g plasmid DNA/106 cells). After 24 h of recoveryincubation in IMDM-10% BCS under normal culture condi-tions, cells were trypsinized, resuspended in phenol red-freeIMDM-10% BCS, plated on six-well slides coated with colla-gen type I at a density of 104/well, and incubated for 1 h inhumidified petri dishes. Slides were then transferred into freshmedia and further incubated for 16 h under normal cultureconditions. The localization of EGFP-tagged GPR93 was vi-sualized by using Laser Scanning Confocal Microscopy (Zeiss510 UV/Vis Meta system).
AEQ-based [Ca2�]i mobilization assay. [Ca2�]i mobiliza-tion assay was performed as previously described (54) withslight modifications. Briefly, mtAEQ expression vector wascoelectroporated (2 �g/106 cells) with other plasmid constructsas indicated in the figures, and the cells were allowed torecover for 20 h in IMDM-10% BCS. For CHO cells, cellswere dislodged with 5 mM EDTA-PBS and loaded with 5 �Mcoelenterazine-h (Promega)-300 �M glutathione in IMDM(2 � 106 cells/ml) at 37°C for 2 h with gentle rolling. ForhBRIE 380i cells, cells were trypsinized and gently rolled for1 h in IMDM-10% BCS, followed by loading in HBSS (In-vitrogen) for 1 h under the same condition as used for CHOcells. A 100-�l aliquot (5 � 104 cells in HBSS) was thenassayed in a luminometer equipped with an injector (TurnerBioSystem). The stimulus was injected into the cell suspensionin a 100-�l aliquot at a 2� final concentration in PBS. A100-�l aliquot of lysis buffer (300 mM CaCl2 and 300 �Mdigitonin) was injected 40 s later to react with the remainingAEQ. Luminescence [as relative light units (RLU)] was re-corded continuously. Fractional RLU is increased RLU due toa stimulus normalized to the total RLU, i.e., the integratedRLU value for 30 s after injection of the stimulus plus that for20 s after the addition of the lysis buffer. All reagents testedwere dissolved in PBS (pH 7.4).
To determine whether protons could activate GPR93, stimuliin the form of the buffer at varying pH were used. Phosphatebuffer (PB; 10 mM Na2HPO4 and 2 mM KH2PO4) was pHadjusted with HCl or NaOH to 6.5 (acidic), 7.4 (neutral), or 8.5(basic), and NaCl was added to bring the osmolality to 300osmol/kgH2O, as determined by microosmometer (PrecisionSystems). To test whether osmotic pressure could stimulateGPR93, modified PB (PB at 100 osmol/kgH2O, adjusted withNaCl, pH. 7.4), 2% glycerol in modified PB (211 osmol/kgH2O), and 4% glycerol in modified PB (553 osmol/kgH2O)were used as stimuli.
BSA digestion. Fatty acid-free BSA (Roche) was dissolvedin PBS (pH 7.4) at a concentration of 10 mg/ml (wt/vol).Digested BSA was prepared by incubating a BSA solution (10mg/ml) with proteinase K (5 �g enzyme/mg BSA, Invitrogen)at 37°C for 20 h with gentle agitation. The solution was heatedat 80°C for 15 min to inactivate proteinase K and then centri-fuged at 16,000 g at 4°C for 5 min.
G100 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

Luciferase reporter assay. Two micrograms of the reporterconstruct/106 of either CHO or hBRIE 380i cells were elec-troporated with other constructs as indicated in the figures, andcells were seeded into 12-well plates at 5 � 105 cells/well inIMDM-10% BCS. For the NFAT reporter study, cells wereallowed to recover in IMDM-10% BCS for 20 h after trans-fection. On the day of each experiment, cells were first washedthree times with PBS, serum starved for 2 h, and then treatedwith either 10 �M LPA or 50 mg/ml peptone in serum-freeIMDM for 6 h. For the TRE reporter study, cells were allowedto recover in IMDM-10% BCS for 36 h. On the day of eachexperiment, cells were washed three times with PBS andtreated with either 10 �M LPA or 50 mg/ml peptone inserum-free IMDM for 12 h. Fatty acid-free BSA, at a concen-tration of 0.1% (wt/vol), was added as a carrier in the treat-ments. Forty microliters of passive lysis buffer (Promega) wereadded to each well after the treatments. The luciferase activi-ties of the samples were determined according to the manu-facturer’s protocol using a luminometer and normalized to thetotal protein concentration, determined by the Bio-Rad proteinassay (Bio-Rad).
Inhibitors treatment. For [Ca2�]i mobilization assay, cellswere incubated with 80 ng/ml pertussis toxin (PTX) for 24 hbefore the assay. U-73122, or its inactive analog U-73343, at10 or 20 �M, was mixed with the stimulus without preincu-bation. Nifedipine (10 �M) and thapsigargin (20 nM) wereadded to the cells 5 min and 30 min, respectively, before astimulus was added. For Western blot analysis, cells werepreincubated with 100 ng/ml PTX for 20 h. The preincubationwith 100 nM wortmannin, 5 �M U-73122 or U-73343, or 50�M PD-98059, was for 30 min.
Western immunoblotting analysis. To determine the effect ofprotein hydrolysate on ERK1/2 activation, hBRIE 380i cellswere transfected with 6 �g GPR93 cDNA/106 cells and laiddown in 60-mm dishes at a density of 2 � 105 per dish. Aftera 24-h incubation in IMDM-10% BCS, cells were serumstarved in IMDM-0.1% BCS for 12 h, followed by a 1-hincubation in HBSS. The cells were treated with 20 mg/ml ofpeptone or 100 nM LPA for 4 min, unless otherwise noted inthe figures. After the treatment, cells were immediately placedon ice, rinsed twice with ice-cold PBS, and scraped with 40 �lof 2� Laemmli sample buffer per 60-mm dish. Western blotanalysis on polyvinylidene difluoride membrane was carriedout as previously described (24). Protein concentration wasdetermined by the Bio-Rad protein assay. The amount ofprotein per lane was 0.2 �g for ERK1 detection and 10 �g forphosphorylated ERK1/2 (pERK1/2). The primary antibodyagainst ERK1 (Santa Cruz Biotechnology) was used at 1:3,000and against pERK1/2 (Cell Signaling) at 1:2,000. The horse-radish peroxidase-conjugated secondary antibody was used at1:10,000.
Sequence analysis. Identity score between different GPCRswas determined by the ClustalW alignment method.
Statistical analysis. Data are expressed as means � SD.Statistical difference between multiple groups was determinedby one-way ANOVA with Tukey’s post hoc test performedusing SPSS version 11. Significance was accepted at P � 0.05.Dose-response curves were generated using the curve-fittingsoftware GraphPad Prism version 4.
RESULTS
Our objective was to study the GPCRs that transduce exter-nal dietary signals into the intestinal epithelial cells. We choseprotein hydrolysate (peptone), a mixture of enzymatically de-rived peptide fragments (mostly between 120 and 1,200 Da)and free amino acids, as a representative dietary componentsince peptone mimics dietary proteins digest in the luminalchyme (12). Protein hydrolysate stimulates mucosal brushborder ERK1/2 (6), but a GPCR that mediates this event hasyet to be identified. The significance of ERK1/2 activity in theintestinal mucosa has been illustrated by the prominent sub-cellular localization of ERK1/2 in the brush border and by theresponsiveness of ERK1/2 signaling to changes in the luminalcontent in response to feeding (6).
We determined that peptone also stimulated the phosphor-ylation of ERK1/2 in enterocyte-derived hBRIE 380i cells(Fig. 1). The oligopeptide transporter PepT1, a reported medi-ator of peptone responses in intestinal cells (14, 44), is notexpressed in hBRIE 380i cells as determined by RT-PCR (datanot shown). We took advantage of this fact to search for othermediators of peptone response using hBRIE 380i cells.
We focused on orphan GPCRs because the activation ofmany GPCRs often leads to increased ERK1/2 (28, 57), andGPCRs have a wide variety of activators: from large glyco-protein hormones, neurotransmitters, and metabolites to exter-nal stimuli, such as light and odorant molecules. Besides beingactivated by their high affinity ligands, GPCRs can also bestimulated by activators with different molecular characteris-tics; for example, the Ca2�-sensing GPCR can also be acti-vated by L-amino acids (9), and the proton-sensing GPCR canalso be activated by lysolipids (59).
The GenBank and Ensembl genomic DNA databases weresearched for putative GPCRs with non-olfactory-like se-quences. RNAs from small intestinal epithelial cells isolated bydispersion (2) and from hBRIE 380i cells were used to screencandidate GPCRs by RT-PCR (unpublished data). The selectedcandidate GPCRs were tested for their responsiveness to pep-tone in a [Ca2�]i mobilization assay by cotransfecting thepromiscuous G�15 (unpublished data).
Fig. 1. Induction of ERK1/2 phosphorylation by peptone in hybrid BerkeleyRat Intestine Epithelial 380 (hBRIE 380i) cells. The hBRIE 380i cells werelaid down in 60-mm tissue culture dishes at the density of 2 � 105/dish. Aftera 24-h incubation at 37°C in Iscove’s modified Dulbecco’s medium (IMDM)-10% bovine calf serum (BCS), cells were serum starved in IMDM-0.1% BCSfor 12 h, followed by a 1-h incubation in HBSS. The cells were treated with 10or 20 mg/ml of peptone for 4 min. The levels of ERK1/2 phosphorylation weredetermined by Western blot analysis as described in MATERIALS AND METHODS.A representative image was shown. The intensity of the phosphorylatedERK1/2 (pERK1/2) bands was normalized to total ERK1, and the result wasexpressed as fold change with the value without peptone arbitrarily set to 1.
G101PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

We determined GPR93 was a candidate GPCR that wasresponsive to peptone. O’Dowd and coworkers (39) reportedits expression in several mouse tissues. GPR93 is a family AGPCR that belongs to a group of purinoreceptor-like GPCRs(29), i.e., P2Y5, P2Y9, and P2Y10. The aim of this study was tocharacterize GPR93 activation by peptone in hBRIE 380i cells,focusing on the collective net [Ca2�]i flux and ERK1/2 phos-phorylation.
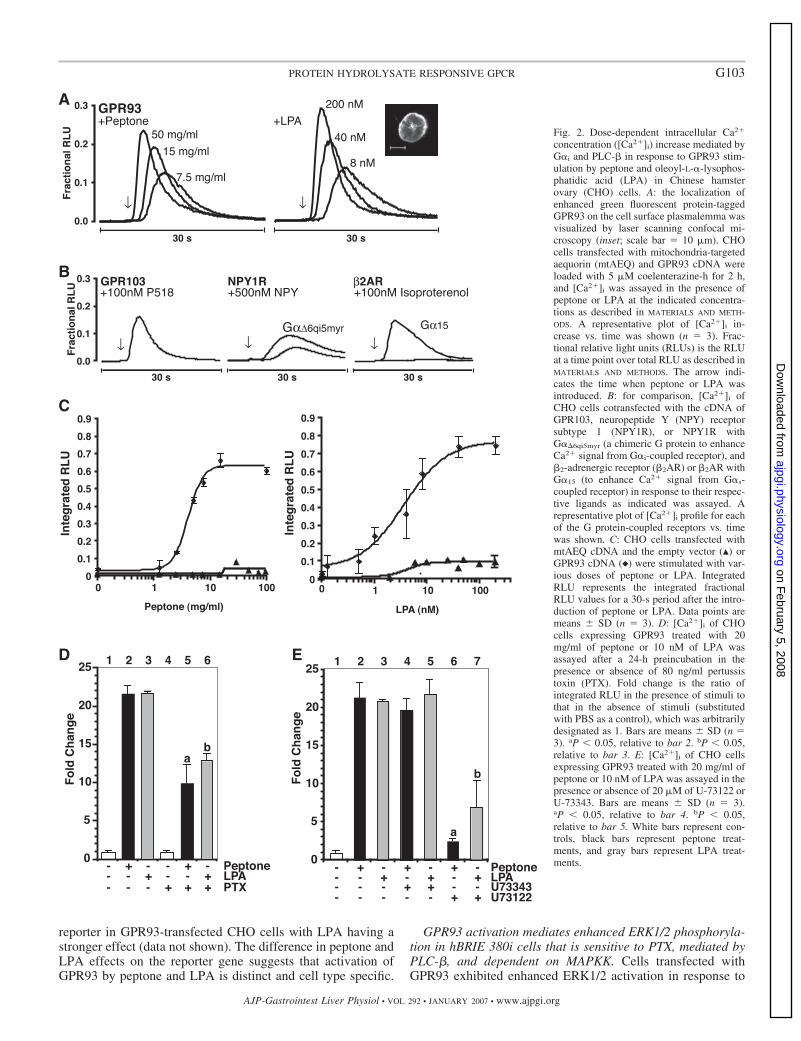
Peptone and LPA induce [Ca2�]i in CHO cells overexpress-ing GPR93. The first step of the characterization of GPR93activation was performed in CHO cells using the mtAEQ-based [Ca2�]i mobilization assay. The proper localization ofthe transfected GPR93 to the cell surface plasmalemma wasconfirmed by fluorescence confocal microscopy using GPR93-EGFP fusion (Fig. 2A, inset). Although in the initial screeningprocess G�15 was cotransfected with GPR93, cells transientlytransfected only with GPR93 also showed a significant, tran-sient [Ca2�]i flux in response to peptone. There was no [Ca2�]i
in the empty vector transfectant in response to peptone, with orwithout a promiscuous G� (data not shown). Thus all experi-ments were carried out with GPR93-transfected cells withoutcotransfecting a promiscuous G� (G�6qi5myr or G�15).
We tested the possibility that peptone might activate otherGPCRs nonspecifically. Peptone-treated CHO cells overex-pressing GPR103, NPY1R (cotransfected with G�6qi5myr), or�2AR (cotransfected with G�15) did not show an increase in[Ca2�]i. P2Y5, P2Y9, and P2Y10, with or without cotransfec-tion with G�15, were also not stimulated by peptone (data notshown). The possibility of GPR93 activation by compoundsother than peptone, most of which could be in the diet, was alsoexplored. Soy protein hydrolysate stimulated GPR93 in asimilar extent to peptone. The compounds that did not induce[Ca2�]i are listed in Table 2. The possibility of the activationof GPR93 by proton/pH or hypo/hypertonicity was also ex-plored. pH from 6.5 to 8.5, as well as osmolality from 100 to553 osmol/kgH2O, did not induce [Ca2�]i in GPR93-overex-pressing CHO cells.
Another compound that was tested was LPA. External LPAcan be present in the diet as it is or as a product of PLA2
digestion in the lumen of the intestine. P2Y9, which has a 30%amino acid identity with GPR93, was recently reported to beactivated by LPA (37). We treated GPR93-transfected cellswith 5 �M LPA and observed that [Ca2�]i was significantlyinduced, but there was no [Ca2�]i release observed in theempty vector transfectant (data not shown). Therefore, LPAwas included as a stimulus to further characterize GPR93activation.
Peptone and LPA activation of GPR93 in CHO and hBRIE380i cells is PTX sensitive and mediated by PLC-�. GPR93-mediated [Ca2�]i flux in response to peptone or LPA was doseresponsive (Fig. 2, A and C). The EC50 was 4 mg/ml forpeptone and 3.6 nM for LPA (Fig. 2C). For comparison, knownGPCRs were activated by their agonists, i.e., NPY1R (a G�i-GPCR) with/without chimeric G�6qi5myr (a chimeric G proteinto enhance signals from G�i-coupled receptor) by NPY, �2AR(a G�s-GPCR) with/without G�15 (to enhance signals fromG�s-coupled receptor) by isoproterenol, and GPR103 [a G�q-GPCR (31)] by RFamide P518 (Fig. 2B). CotransfectingG�6qi5myr was needed to observe a significant increase in[Ca2�]i flux elicited through NPY1R. This was similar to�2AR, for which a cotransfection with G�15 was required to
yield an observable [Ca2�]i peak. In contrast, upon its activa-tion, GPR103 yielded an increased [Ca2�]i similar to that ofGPR93 without the addition of a promiscuous G� protein. Thissuggests that a G�q-mediated pathway is downstream ofGPR93 activation. A [Ca2�]i flux due to a GPCR activationcan be caused by G�q or G� dissociated from G�i. PTX (80ng/ml), a specific inhibitor of G�i, reduced peptone stimulationby 50% and LPA stimulation by 60% (Fig. 2D), suggesting thatGPR93 activation partially leads to a pathway involving G�i.GPCRs often exert intracellular signaling events by couplingwith more than one kind of G proteins; as an example, LPAreceptor 1, 2, and 3 can couple with both G�i and G�q (8). Thepossible involvement of other G� proteins in GPR93-mediated[Ca2�]i increase in our system remains to be determined. Apossible downstream effector of G�i- or G�q-coupled recep-tors mediating the increase of [Ca2�]i is PLC-�. A treatmentwith PLC-� inhibitor U-73122, but not its inactive analogU-73343, almost abolished peptone induction and inhibitedLPA stimulation by more than 50% (Fig. 2E). PLC-� liberatesinositol triphosphate, which activates the Ca2� channel on theendoplasmic reticulum membrane resulting in a [Ca2�]i flux.Nifedipine (10 �M), an inhibitor of plasma membrane L-typevoltage-gated Ca2� channel, did not affect the [Ca2�]i fluxinduced by peptone or LPA stimulation of GPR93 (data notshown). Thapsigargin (20 nM), an inhibitor of sarco(endo)plas-mic reticulum Ca2� ATPase, completely eliminated the in-crease in [Ca2�]i, suggesting that GPR93 activation inducesCa2� release from intracellular stores (data not shown).
We repeated the characterization of GPR93 activation inhBRIE 380i cells. In hBRIE 380i cells transiently transfectedwith GPR93, peptone or LPA induced a [Ca2�]i flux withoutcotransfecting a promiscuous G� protein. The increase in[Ca2�]i levels was concentration dependent (Fig. 3A). TheEC50 for peptone was 10.6 mg/ ml and 7.9 nM for LPA (Fig.3A). PTX reduced the peptone stimulation by 68% and LPAstimulation by 50% (Fig. 3B). U-73122 decreased peptonestimulation by 83%, whereas U-73343 had no effects (Fig. 3C).The effect of U-73122 was dose responsive (data not shown).Similarly, U-73122, but not U-73343, inhibited LPA stimula-tion by 52% (Fig. 3C). These data are consistent with what wasobserved in CHO cells.
GPR93 activation leads to induced NFAT- and TRE-lucif-erase reporter activities. We tested whether the increase in[Ca2�]i levels due to GPR93 activation could lead to down-stream gene responses by the luciferase reporter assay. Weexamined the luciferase activity of reporter constructs contain-ing response elements to NFAT and TPA (TRE) as indicatorsfor increased [Ca2�]i. GPR93 activation by peptone or LPA inCHO cells induced luciferase activity from a construct con-taining NFAT responsive elements (Fig. 4A). The NFAT lu-ciferase reporter could not be used in hBRIE 380i cells becauseeven the positive control (TPA plus ionomycin) did not inducethe luciferase activity, which might be due to the lack ofexpression of the transcription factor (unpublished data).Therefore, we used a luciferase reporter construct containingTRE. The activation of GPR93 by peptone or LPA inducedluciferase expression (Fig. 4B). Interestingly, the effect ofpeptone on NFAT-driven reporter was weaker than that of LPAin CHO cells, but it was LPA that had a weaker effect on theTRE luciferase reporter in hBRIE 380i cells. Peptone and LPAalso induced luciferase expression from the TRE-luciferase
G102 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from
reporter in GPR93-transfected CHO cells with LPA having astronger effect (data not shown). The difference in peptone andLPA effects on the reporter gene suggests that activation ofGPR93 by peptone and LPA is distinct and cell type specific.
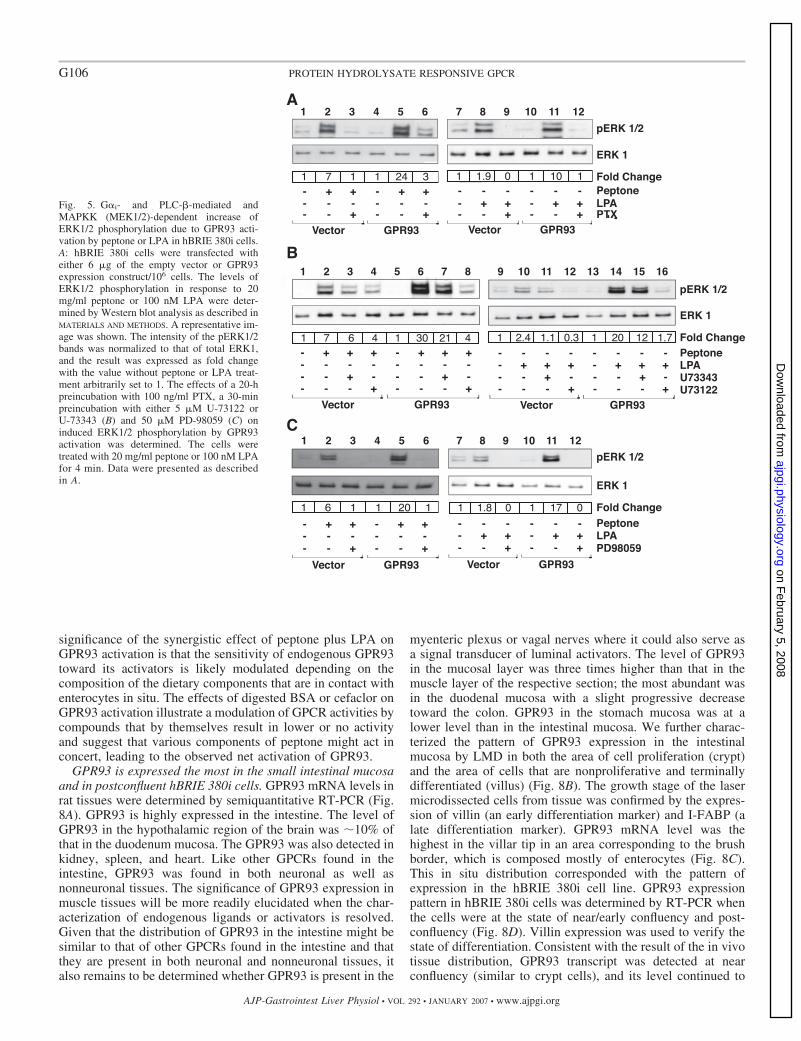
GPR93 activation mediates enhanced ERK1/2 phosphoryla-tion in hBRIE 380i cells that is sensitive to PTX, mediated byPLC-�, and dependent on MAPKK. Cells transfected withGPR93 exhibited enhanced ERK1/2 activation in response to
Fig. 2. Dose-dependent intracellular Ca2�
concentration ([Ca2�]i) increase mediated byG�i and PLC-� in response to GPR93 stim-ulation by peptone and oleoyl-L-�-lysophos-phatidic acid (LPA) in Chinese hamsterovary (CHO) cells. A: the localization ofenhanced green fluorescent protein-taggedGPR93 on the cell surface plasmalemma wasvisualized by laser scanning confocal mi-croscopy (inset; scale bar 10 �m). CHOcells transfected with mitochondria-targetedaequorin (mtAEQ) and GPR93 cDNA wereloaded with 5 �M coelenterazine-h for 2 h,and [Ca2�]i was assayed in the presence ofpeptone or LPA at the indicated concentra-tions as described in MATERIALS AND METH-ODS. A representative plot of [Ca2�]i in-crease vs. time was shown (n 3). Frac-tional relative light units (RLUs) is the RLUat a time point over total RLU as described inMATERIALS AND METHODS. The arrow indi-cates the time when peptone or LPA wasintroduced. B: for comparison, [Ca2�]i ofCHO cells cotransfected with the cDNA ofGPR103, neuropeptide Y (NPY) receptorsubtype 1 (NPY1R), or NPY1R withG��6qi5myr (a chimeric G protein to enhanceCa2� signal from G�i-coupled receptor), and�2-adrenergic receptor (�2AR) or �2AR withG�15 (to enhance Ca2� signal from G�s-coupled receptor) in response to their respec-tive ligands as indicated was assayed. Arepresentative plot of [Ca2�]i profile for eachof the G protein-coupled receptors vs. timewas shown. C: CHO cells transfected withmtAEQ cDNA and the empty vector (Œ) orGPR93 cDNA (}) were stimulated with var-ious doses of peptone or LPA. IntegratedRLU represents the integrated fractionalRLU values for a 30-s period after the intro-duction of peptone or LPA. Data points aremeans � SD (n 3). D: [Ca2�]i of CHOcells expressing GPR93 treated with 20mg/ml of peptone or 10 nM of LPA wasassayed after a 24-h preincubation in thepresence or absence of 80 ng/ml pertussistoxin (PTX). Fold change is the ratio ofintegrated RLU in the presence of stimuli tothat in the absence of stimuli (substitutedwith PBS as a control), which was arbitrarilydesignated as 1. Bars are means � SD (n 3). aP � 0.05, relative to bar 2. bP � 0.05,relative to bar 3. E: [Ca2�]i of CHO cellsexpressing GPR93 treated with 20 mg/ml ofpeptone or 10 nM of LPA was assayed in thepresence or absence of 20 �M of U-73122 orU-73343. Bars are means � SD (n 3).aP � 0.05, relative to bar 4. bP � 0.05,relative to bar 5. White bars represent con-trols, black bars represent peptone treat-ments, and gray bars represent LPA treat-ments.
G103PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

peptone or LPA compared with cells transfected with theempty vector. The level of pERK1/2 was induced rapidly inresponse to peptone or LPA, reaching a maximum at 4 min andthen gradually decreasing (data not shown). Peptone at a dosebetween 20 and 50 mg/ml maximally induced ERK1/2 phos-phorylation in GPR93-overexpressing cells, whereas specificLPA stimulation was only observed at 100 nM (data notshown).
To determine the correlation between GPR93 activation andthe increase in pERK, PTX and U-73122 were used. PTX (100ng/ml) almost abolished peptone (Fig. 5A) as well as LPA (Fig.5A) induction. Likewise, U-73122 decreased pERK inductionby peptone (Fig. 5B) and LPA (Fig. 5B) to the basal levels. Thetreatment with phosphatidylinositol 3-kinase inhibitor wort-mannin (100 nM) did not change the effect of peptone onpERK (data not shown). These results suggest an involvementof G� subunits from G�i and PLC-� activation in the induc-tion of pERK. This does not exclude the possibility of otherpathways, which were not explored in this study, contributingto ERK1/2 activation (including those that are G�q mediated).The observed induction of pERK due to GPR93 activationrequires MAPKK (MEK1/2), since PD-98059 (MEK1/2 inhib-itor) at 50 �M abolished induced pERK by peptone (Fig. 5C)and LPA (Fig. 5C).
Peptone and LPA synergistically activate GPR93. Whetherpeptone and LPA could act in concert to activate GPR93 wastested by treating GPR93-transfected hBRIE 380i cells withvarying doses of peptone and LPA simultaneously in a [Ca2�]i
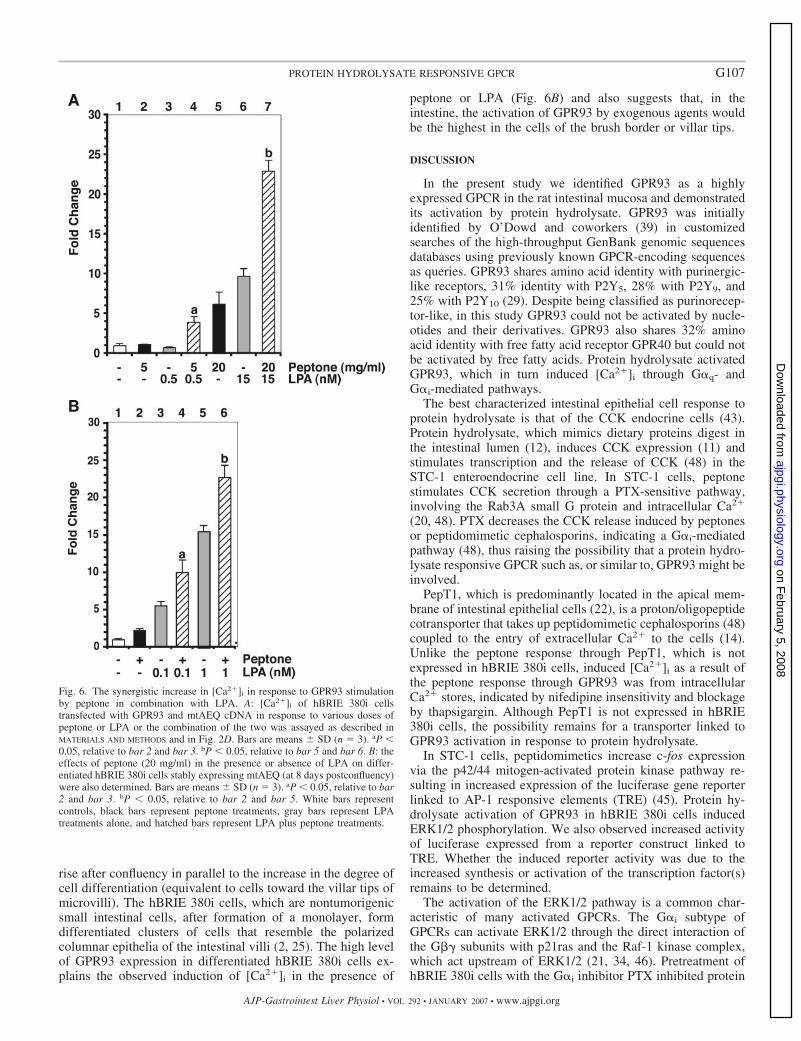
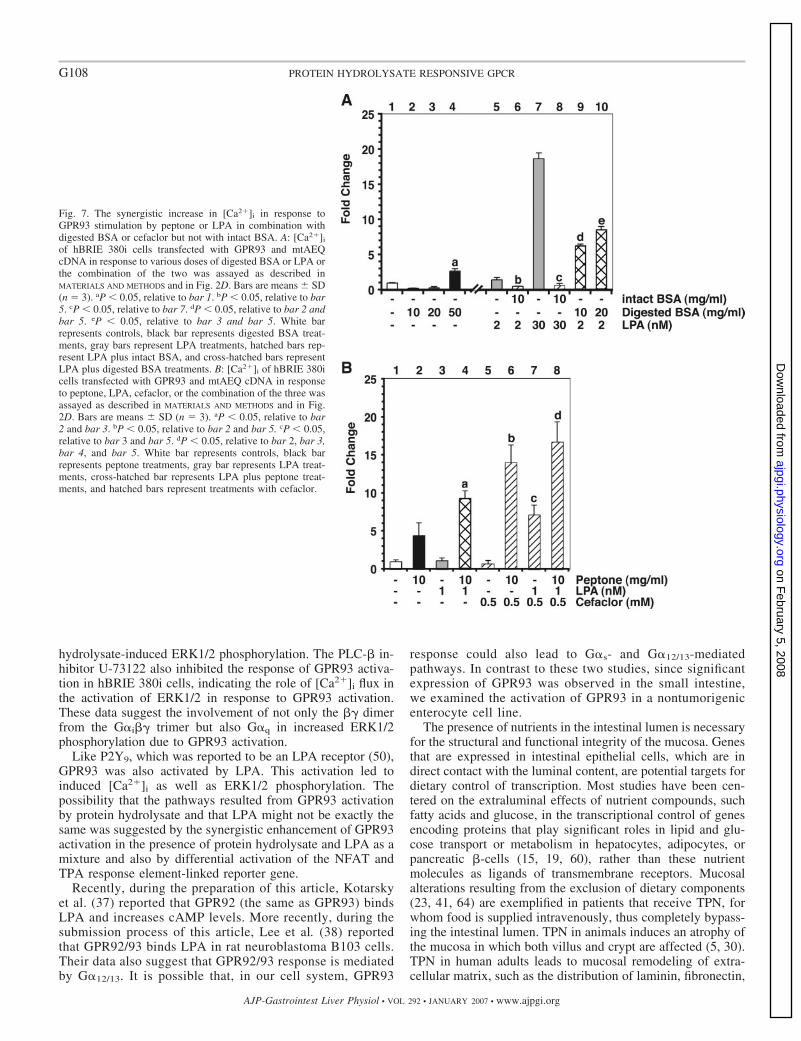
mobilization assay (Fig. 6A). Peptone or LPA individuallyactivated GPR93. However, [Ca2�]i increase was more thanadditive when both were combined and used as a stimulus (Fig.6A, bar 4 vs. 2 and 3; and bar 7 vs. 5 and 6). Fatty acid-freeBSA at 0.1% and peptone were added together in this partic-ular experiment because LPA solutions used in all our exper-iments contained 0.1% fatty acid-free BSA as a carrier. Weobserved that GPR93 did not significantly respond to BSAstimulation up to 100 mg/ml (data not shown); however, BSAreduced [Ca2�]i flux due to peptone (data not shown) or LPAstimulation (Fig. 7A, bar 6 vs. 5; and bar 8 vs. 7). Therefore,[Ca2�]i induction by peptone in Fig. 6 was to a less extent thanwhat was shown in Fig. 3C.
GPR93 could be activated by peptone and LPA as shown inFig. 3. We tested whether endogenous GPR93 activation inhBRIE 380i cells could also be determined by [Ca2�]i mobi-lization assay using hBRIE 380i cells stably expressingmtAEQ. At the cell stage before confluency, peptone or LPAdid not increase [Ca2�]i (data not shown). However, when thecells were allowed to become differentiated at 8-d postconflu-ency, peptone and LPA induced [Ca2�]i (Fig. 6B). Inductionby peptone was much weaker than LPA compared with whatwas observed in proliferating cells in the transient transfectionsystem. This might be due to increased expression of otherendogenous LPA receptors in our cell culture system that,unlike GPR93, are not peptone responsive. The induction of[Ca2�]i was synergistically enhanced when peptone andLPA were combined (Fig. 6B, bar 4 vs. 2 and 3; and bar 6vs. 2 and 5).
We used two approaches to verify that the peptide compo-nents in peptone were indeed responsible for GPR93 activationand for synergistic effects of peptone plus LPA on GPR93activation. We tested the effect of proteinase K-digested fattyacid-free BSA on the [Ca2�]i flux in the GPR93-overexpress-ing hBRIE 380i cells. In contrast to undigested BSA, which didnot induce a change in [Ca2�]i, digested BSA (50 mg/ml)showed a significant increase over basal (Fig. 7A, bar 4 vs. 1).The change in [Ca2�]i was less than that observed with peptoneat a similar dose. This is likely due to the fact that the peptidecomponents in peptone are derived from more than one kind ofprotein. Digested BSA synergistically induced [Ca2�]i releasewhen it was combined with LPA (Fig. 7A, bar 9 vs. 2 and 5;and bar 10 vs. 3 and 5). We also utilized cefaclor, a peptido-mimetic substrate of the oligopeptide transporter PepT1 (7),which did not induce [Ca2�]i release in hBRIE 380i cells (Fig.7B, bar 5; and tested up to 5 mM; data not shown). Cefaclorsynergistically enhanced GPR93 activation by peptone or LPA(Fig. 7B, bar 6 vs. 2 and 5; and bar 7 vs. 3 and 5). Enhancedactivation of GPR93 by a combination of peptone and LPAwas also further amplified in the presence of cefaclor (Fig. 7B,bar 8 vs. 4).
The implication of this result is that LPA and peptone mightact on different sites of the GPR93 molecule. Although at thedoses of peptone and LPA that were used, the effects weresynergistic whether the GPR93 activation by saturated doses ofpeptone could be amplified by LPA and vice versa could not bedetermined due to the nature of the assay. The physiological
Table 2. The list of compounds that did not activate GPR93
Category Compounds Tested
Amino acids Casamino acidsEach of the 20 amino acids
Di_tripeptides Glycyl-glycine -Glutamyl - cysteinyl - glycine
Monosaccharides ArabinoseGlucoseXyloseFructoseGalactose
Disaccharides �-Lactose�-LactoseMaltoseMelibioseSucroseTrehalose
Sugar alcohol SorbitolShort chain fatty acids Acetic acid
Butyric acidPropionic acid
Long-chain saturated fatty acids Lauric acidMyristic acidPalmitic acid
Long-chain unsaturated fatty acid Oleic acidBile acids Cholic acid
Deoxycholic acidTaurocholic acid
Nucleotides CMPADPCDPATPUTP
Sugar nucleotides UDP-glucoseUDP-galactoseUDP-glucuronic acidUDP-N-acetylglucosamine
G104 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from
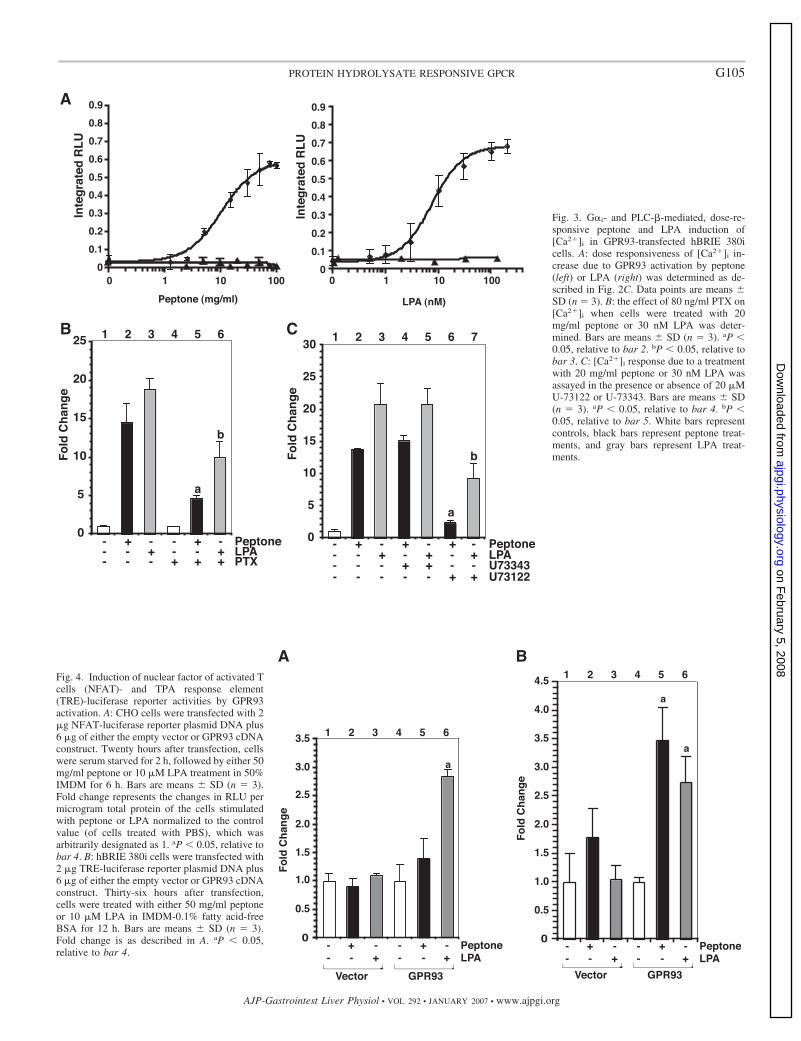
Fig. 3. G�i- and PLC-�-mediated, dose-re-sponsive peptone and LPA induction of[Ca2�]i in GPR93-transfected hBRIE 380icells. A: dose responsiveness of [Ca2�]i in-crease due to GPR93 activation by peptone(left) or LPA (right) was determined as de-scribed in Fig. 2C. Data points are means �SD (n 3). B: the effect of 80 ng/ml PTX on[Ca2�]i when cells were treated with 20mg/ml peptone or 30 nM LPA was deter-mined. Bars are means � SD (n 3). aP �0.05, relative to bar 2. bP � 0.05, relative tobar 3. C: [Ca2�]i response due to a treatmentwith 20 mg/ml peptone or 30 nM LPA wasassayed in the presence or absence of 20 �MU-73122 or U-73343. Bars are means � SD(n 3). aP � 0.05, relative to bar 4. bP �0.05, relative to bar 5. White bars representcontrols, black bars represent peptone treat-ments, and gray bars represent LPA treat-ments.
Fig. 4. Induction of nuclear factor of activated Tcells (NFAT)- and TPA response element(TRE)-luciferase reporter activities by GPR93activation. A: CHO cells were transfected with 2�g NFAT-luciferase reporter plasmid DNA plus6 �g of either the empty vector or GPR93 cDNAconstruct. Twenty hours after transfection, cellswere serum starved for 2 h, followed by either 50mg/ml peptone or 10 �M LPA treatment in 50%IMDM for 6 h. Bars are means � SD (n 3).Fold change represents the changes in RLU permicrogram total protein of the cells stimulatedwith peptone or LPA normalized to the controlvalue (of cells treated with PBS), which wasarbitrarily designated as 1. aP � 0.05, relative tobar 4. B: hBRIE 380i cells were transfected with2 �g TRE-luciferase reporter plasmid DNA plus6 �g of either the empty vector or GPR93 cDNAconstruct. Thirty-six hours after transfection,cells were treated with either 50 mg/ml peptoneor 10 �M LPA in IMDM-0.1% fatty acid-freeBSA for 12 h. Bars are means � SD (n 3).Fold change is as described in A. aP � 0.05,relative to bar 4.
G105PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from
significance of the synergistic effect of peptone plus LPA onGPR93 activation is that the sensitivity of endogenous GPR93toward its activators is likely modulated depending on thecomposition of the dietary components that are in contact withenterocytes in situ. The effects of digested BSA or cefaclor onGPR93 activation illustrate a modulation of GPCR activities bycompounds that by themselves result in lower or no activityand suggest that various components of peptone might act inconcert, leading to the observed net activation of GPR93.
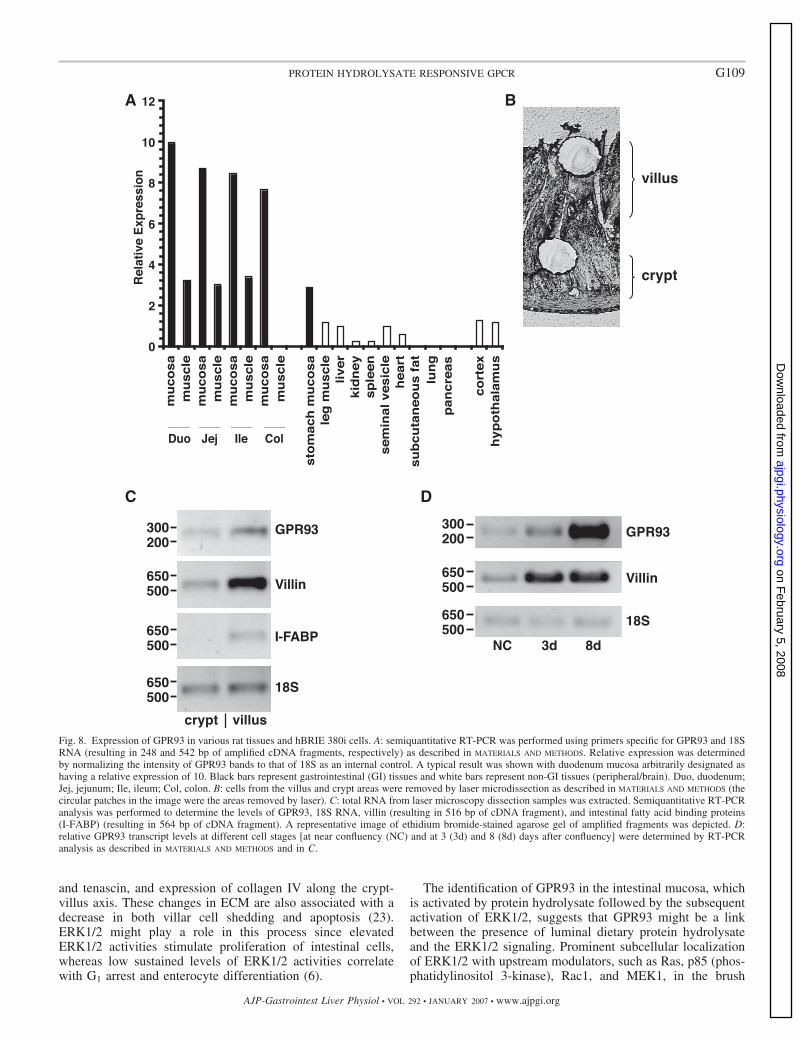
GPR93 is expressed the most in the small intestinal mucosaand in postconfluent hBRIE 380i cells. GPR93 mRNA levels inrat tissues were determined by semiquantitative RT-PCR (Fig.8A). GPR93 is highly expressed in the intestine. The level ofGPR93 in the hypothalamic region of the brain was �10% ofthat in the duodenum mucosa. The GPR93 was also detected inkidney, spleen, and heart. Like other GPCRs found in theintestine, GPR93 was found in both neuronal as well asnonneuronal tissues. The significance of GPR93 expression inmuscle tissues will be more readily elucidated when the char-acterization of endogenous ligands or activators is resolved.Given that the distribution of GPR93 in the intestine might besimilar to that of other GPCRs found in the intestine and thatthey are present in both neuronal and nonneuronal tissues, italso remains to be determined whether GPR93 is present in the
myenteric plexus or vagal nerves where it could also serve asa signal transducer of luminal activators. The level of GPR93in the mucosal layer was three times higher than that in themuscle layer of the respective section; the most abundant wasin the duodenal mucosa with a slight progressive decreasetoward the colon. GPR93 in the stomach mucosa was at alower level than in the intestinal mucosa. We further charac-terized the pattern of GPR93 expression in the intestinalmucosa by LMD in both the area of cell proliferation (crypt)and the area of cells that are nonproliferative and terminallydifferentiated (villus) (Fig. 8B). The growth stage of the lasermicrodissected cells from tissue was confirmed by the expres-sion of villin (an early differentiation marker) and I-FABP (alate differentiation marker). GPR93 mRNA level was thehighest in the villar tip in an area corresponding to the brushborder, which is composed mostly of enterocytes (Fig. 8C).This in situ distribution corresponded with the pattern ofexpression in the hBRIE 380i cell line. GPR93 expressionpattern in hBRIE 380i cells was determined by RT-PCR whenthe cells were at the state of near/early confluency and post-confluency (Fig. 8D). Villin expression was used to verify thestate of differentiation. Consistent with the result of the in vivotissue distribution, GPR93 transcript was detected at nearconfluency (similar to crypt cells), and its level continued to
Fig. 5. G�i- and PLC-�-mediated andMAPKK (MEK1/2)-dependent increase ofERK1/2 phosphorylation due to GPR93 acti-vation by peptone or LPA in hBRIE 380i cells.A: hBRIE 380i cells were transfected witheither 6 �g of the empty vector or GPR93expression construct/106 cells. The levels ofERK1/2 phosphorylation in response to 20mg/ml peptone or 100 nM LPA were deter-mined by Western blot analysis as described inMATERIALS AND METHODS. A representative im-age was shown. The intensity of the pERK1/2bands was normalized to that of total ERK1,and the result was expressed as fold changewith the value without peptone or LPA treat-ment arbitrarily set to 1. The effects of a 20-hpreincubation with 100 ng/ml PTX, a 30-minpreincubation with either 5 �M U-73122 orU-73343 (B) and 50 �M PD-98059 (C) oninduced ERK1/2 phosphorylation by GPR93activation was determined. The cells weretreated with 20 mg/ml peptone or 100 nM LPAfor 4 min. Data were presented as describedin A.
G106 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from
rise after confluency in parallel to the increase in the degree ofcell differentiation (equivalent to cells toward the villar tips ofmicrovilli). The hBRIE 380i cells, which are nontumorigenicsmall intestinal cells, after formation of a monolayer, formdifferentiated clusters of cells that resemble the polarizedcolumnar epithelia of the intestinal villi (2, 25). The high levelof GPR93 expression in differentiated hBRIE 380i cells ex-plains the observed induction of [Ca2�]i in the presence of
peptone or LPA (Fig. 6B) and also suggests that, in theintestine, the activation of GPR93 by exogenous agents wouldbe the highest in the cells of the brush border or villar tips.
DISCUSSION
In the present study we identified GPR93 as a highlyexpressed GPCR in the rat intestinal mucosa and demonstratedits activation by protein hydrolysate. GPR93 was initiallyidentified by O’Dowd and coworkers (39) in customizedsearches of the high-throughput GenBank genomic sequencesdatabases using previously known GPCR-encoding sequencesas queries. GPR93 shares amino acid identity with purinergic-like receptors, 31% identity with P2Y5, 28% with P2Y9, and25% with P2Y10 (29). Despite being classified as purinorecep-tor-like, in this study GPR93 could not be activated by nucle-otides and their derivatives. GPR93 also shares 32% aminoacid identity with free fatty acid receptor GPR40 but could notbe activated by free fatty acids. Protein hydrolysate activatedGPR93, which in turn induced [Ca2�]i through G�q- andG�i-mediated pathways.
The best characterized intestinal epithelial cell response toprotein hydrolysate is that of the CCK endocrine cells (43).Protein hydrolysate, which mimics dietary proteins digest inthe intestinal lumen (12), induces CCK expression (11) andstimulates transcription and the release of CCK (48) in theSTC-1 enteroendocrine cell line. In STC-1 cells, peptonestimulates CCK secretion through a PTX-sensitive pathway,involving the Rab3A small G protein and intracellular Ca2�
(20, 48). PTX decreases the CCK release induced by peptonesor peptidomimetic cephalosporins, indicating a G�i-mediatedpathway (48), thus raising the possibility that a protein hydro-lysate responsive GPCR such as, or similar to, GPR93 might beinvolved.
PepT1, which is predominantly located in the apical mem-brane of intestinal epithelial cells (22), is a proton/oligopeptidecotransporter that takes up peptidomimetic cephalosporins (48)coupled to the entry of extracellular Ca2� to the cells (14).Unlike the peptone response through PepT1, which is notexpressed in hBRIE 380i cells, induced [Ca2�]i as a result ofthe peptone response through GPR93 was from intracellularCa2� stores, indicated by nifedipine insensitivity and blockageby thapsigargin. Although PepT1 is not expressed in hBRIE380i cells, the possibility remains for a transporter linked toGPR93 activation in response to protein hydrolysate.
In STC-1 cells, peptidomimetics increase c-fos expressionvia the p42/44 mitogen-activated protein kinase pathway re-sulting in increased expression of the luciferase gene reporterlinked to AP-1 responsive elements (TRE) (45). Protein hy-drolysate activation of GPR93 in hBRIE 380i cells inducedERK1/2 phosphorylation. We also observed increased activityof luciferase expressed from a reporter construct linked toTRE. Whether the induced reporter activity was due to theincreased synthesis or activation of the transcription factor(s)remains to be determined.
The activation of the ERK1/2 pathway is a common char-acteristic of many activated GPCRs. The G�i subtype ofGPCRs can activate ERK1/2 through the direct interaction ofthe G� subunits with p21ras and the Raf-1 kinase complex,which act upstream of ERK1/2 (21, 34, 46). Pretreatment ofhBRIE 380i cells with the G�i inhibitor PTX inhibited protein
Fig. 6. The synergistic increase in [Ca2�]i in response to GPR93 stimulationby peptone in combination with LPA. A: [Ca2�]i of hBRIE 380i cellstransfected with GPR93 and mtAEQ cDNA in response to various doses ofpeptone or LPA or the combination of the two was assayed as described inMATERIALS AND METHODS and in Fig. 2D. Bars are means � SD (n 3). aP �0.05, relative to bar 2 and bar 3. bP � 0.05, relative to bar 5 and bar 6. B: theeffects of peptone (20 mg/ml) in the presence or absence of LPA on differ-entiated hBRIE 380i cells stably expressing mtAEQ (at 8 days postconfluency)were also determined. Bars are means � SD (n 3). aP � 0.05, relative to bar2 and bar 3. bP � 0.05, relative to bar 2 and bar 5. White bars representcontrols, black bars represent peptone treatments, gray bars represent LPAtreatments alone, and hatched bars represent LPA plus peptone treatments.
G107PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from
hydrolysate-induced ERK1/2 phosphorylation. The PLC-� in-hibitor U-73122 also inhibited the response of GPR93 activa-tion in hBRIE 380i cells, indicating the role of [Ca2�]i flux inthe activation of ERK1/2 in response to GPR93 activation.These data suggest the involvement of not only the � dimerfrom the G�i� trimer but also G�q in increased ERK1/2phosphorylation due to GPR93 activation.
Like P2Y9, which was reported to be an LPA receptor (50),GPR93 was also activated by LPA. This activation led toinduced [Ca2�]i as well as ERK1/2 phosphorylation. Thepossibility that the pathways resulted from GPR93 activationby protein hydrolysate and that LPA might not be exactly thesame was suggested by the synergistic enhancement of GPR93activation in the presence of protein hydrolysate and LPA as amixture and also by differential activation of the NFAT andTPA response element-linked reporter gene.
Recently, during the preparation of this article, Kotarskyet al. (37) reported that GPR92 (the same as GPR93) bindsLPA and increases cAMP levels. More recently, during thesubmission process of this article, Lee et al. (38) reportedthat GPR92/93 binds LPA in rat neuroblastoma B103 cells.Their data also suggest that GPR92/93 response is mediatedby G�12/13. It is possible that, in our cell system, GPR93
response could also lead to G�s- and G�12/13-mediatedpathways. In contrast to these two studies, since significantexpression of GPR93 was observed in the small intestine,we examined the activation of GPR93 in a nontumorigenicenterocyte cell line.
The presence of nutrients in the intestinal lumen is necessaryfor the structural and functional integrity of the mucosa. Genesthat are expressed in intestinal epithelial cells, which are indirect contact with the luminal content, are potential targets fordietary control of transcription. Most studies have been cen-tered on the extraluminal effects of nutrient compounds, suchfatty acids and glucose, in the transcriptional control of genesencoding proteins that play significant roles in lipid and glu-cose transport or metabolism in hepatocytes, adipocytes, orpancreatic �-cells (15, 19, 60), rather than these nutrientmolecules as ligands of transmembrane receptors. Mucosalalterations resulting from the exclusion of dietary components(23, 41, 64) are exemplified in patients that receive TPN, forwhom food is supplied intravenously, thus completely bypass-ing the intestinal lumen. TPN in animals induces an atrophy ofthe mucosa in which both villus and crypt are affected (5, 30).TPN in human adults leads to mucosal remodeling of extra-cellular matrix, such as the distribution of laminin, fibronectin,
Fig. 7. The synergistic increase in [Ca2�]i in response toGPR93 stimulation by peptone or LPA in combination withdigested BSA or cefaclor but not with intact BSA. A: [Ca2�]i
of hBRIE 380i cells transfected with GPR93 and mtAEQcDNA in response to various doses of digested BSA or LPA orthe combination of the two was assayed as described inMATERIALS AND METHODS and in Fig. 2D. Bars are means � SD(n 3). aP � 0.05, relative to bar 1. bP � 0.05, relative to bar5. cP � 0.05, relative to bar 7. dP � 0.05, relative to bar 2 andbar 5. eP � 0.05, relative to bar 3 and bar 5. White barrepresents controls, black bar represents digested BSA treat-ments, gray bars represent LPA treatments, hatched bars rep-resent LPA plus intact BSA, and cross-hatched bars representLPA plus digested BSA treatments. B: [Ca2�]i of hBRIE 380icells transfected with GPR93 and mtAEQ cDNA in responseto peptone, LPA, cefaclor, or the combination of the three wasassayed as described in MATERIALS AND METHODS and in Fig.2D. Bars are means � SD (n 3). aP � 0.05, relative to bar2 and bar 3. bP � 0.05, relative to bar 2 and bar 5. cP � 0.05,relative to bar 3 and bar 5. dP � 0.05, relative to bar 2, bar 3,bar 4, and bar 5. White bar represents controls, black barrepresents peptone treatments, gray bar represents LPA treat-ments, cross-hatched bar represents LPA plus peptone treat-ments, and hatched bars represent treatments with cefaclor.
G108 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from
and tenascin, and expression of collagen IV along the crypt-villus axis. These changes in ECM are also associated with adecrease in both villar cell shedding and apoptosis (23).ERK1/2 might play a role in this process since elevatedERK1/2 activities stimulate proliferation of intestinal cells,whereas low sustained levels of ERK1/2 activities correlatewith G1 arrest and enterocyte differentiation (6).
The identification of GPR93 in the intestinal mucosa, whichis activated by protein hydrolysate followed by the subsequentactivation of ERK1/2, suggests that GPR93 might be a linkbetween the presence of luminal dietary protein hydrolysateand the ERK1/2 signaling. Prominent subcellular localizationof ERK1/2 with upstream modulators, such as Ras, p85 (phos-phatidylinositol 3-kinase), Rac1, and MEK1, in the brush
Fig. 8. Expression of GPR93 in various rat tissues and hBRIE 380i cells. A: semiquantitative RT-PCR was performed using primers specific for GPR93 and 18SRNA (resulting in 248 and 542 bp of amplified cDNA fragments, respectively) as described in MATERIALS AND METHODS. Relative expression was determinedby normalizing the intensity of GPR93 bands to that of 18S as an internal control. A typical result was shown with duodenum mucosa arbitrarily designated ashaving a relative expression of 10. Black bars represent gastrointestinal (GI) tissues and white bars represent non-GI tissues (peripheral/brain). Duo, duodenum;Jej, jejunum; Ile, ileum; Col, colon. B: cells from the villus and crypt areas were removed by laser microdissection as described in MATERIALS AND METHODS (thecircular patches in the image were the areas removed by laser). C: total RNA from laser microscopy dissection samples was extracted. Semiquantitative RT-PCRanalysis was performed to determine the levels of GPR93, 18S RNA, villin (resulting in 516 bp of cDNA fragment), and intestinal fatty acid binding proteins(I-FABP) (resulting in 564 bp of cDNA fragment). A representative image of ethidium bromide-stained agarose gel of amplified fragments was depicted. D:relative GPR93 transcript levels at different cell stages [at near confluency (NC) and at 3 (3d) and 8 (8d) days after confluency] were determined by RT-PCRanalysis as described in MATERIALS AND METHODS and in C.
G109PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

border of differentiated human enterocytes suggests that one ofthe sites of action of ERK1/2 signaling is at the apex of theseepithelial cells in the brush border where ERK1/2 activitycould also be responsive to changes in luminal contents inresponse to feeding. Although it remains to be determinedwhether GPR93 colocalizes with microvillar ERK1/2, the iden-tification of this receptor in the brush border of the intestinalmucosal epithelial cells raises the possibility that GPR93 mightbe partly responsible for the activation of ERK1/2 observed inresponse to luminal factors, such as dietary protein hydroly-sate, which results in the ERK1/2-dependent changes alongcrypt-to-villar length and in the cellular absorptive capac-ity (6).
The physiological significance of the synergistic effect ofpeptone plus LPA on GPR93 activation is that the sensitivityof endogenous GPR93 toward its activators is likely mod-ulated by the composition of dietary components that are incontact with enterocytes in situ. This is further exemplifiedby the action of the peptide mimic cefaclor, which by itselfdid not activate GPR93 as determined by the lack of Ca2�
response but significantly enhanced GPR93 activation byprotein hydrolysate and LPA. Unlike the receptors ex-pressed in enterocytes under cell culture conditions, recep-tors expressed in enterocytes in the mucosa are not likelyexposed to a signal activator in exclusion of other extracel-lular molecules that could alter its activity. Our observationthat the activation of GPR93 by LPA was significantlyattenuated in the presence of intact BSA is consistent withwhat was reported for the LPA endothelial cell differentia-tion gene 7 receptor, the activation of which by LPA iscompletely blocked with 1% BSA (26). The presence offactors that could contribute to the negative or positiveregulation of GPR93 in response to activators, such as LPAor protein hydrolysate, likely varies depending on the phys-iological state of the organism or the receptor expressingtissue. The effects of cefaclor also illustrate that, physiolog-ically, the activation of a GPCR may not be determined bythe presence of an isolated agonist since the cell in whichthat GPCR is expressed encounters various compounds,both agonists and nonagonists. This is particularly importantfor enterocytes that are exposed to external luminal mole-cules.
Although protein hydrolysate stimulates GPR93, proteinhydrolysate may also act as a receptor modulator in responseto a specific agonist. The mechanism for the activation ofGPR93 by the components of protein hydrolysate remains tobe determined. GPCR activation involves the relaxation ofconstraining intramolecular interactions and the formationof new interactions involving specific movements of thetransmembrane helixes (4, 17, 32). In response to differentinteractions with agonists, antagonists, and inverse agonists,GPCRs have the potential to adopt multiple activated con-formational states (18, 51). Each of these states may havedifferent affinities for ligands, G proteins, and the proteinsthat control receptor internalization (16, 62). Therefore, thecomponents of protein hydrolysate provide many possiblereceptor-protein hydrolysate interactions that might lead tothe observed net activation of GPR93. The ability of GPCRsto act as a toggle switch when activated (55), as well as theirability to integrate its response to a multitude of extracel-lular and intracellular effectors, and to deliver a graded
signaling response make these seven transmembrane recep-tors suitable candidates for molecular sensors that respondto the luminal content as a cocktail of activators.
The present study is the first to identify a GPCR in theenterocytes that could be partly responsible for the reportedmucosal responses to luminal protein hydrolysate, such asERK1/2 activation, endocrine cell secretions, and cell prolif-eration and differentiation. Future studies may reveal the pos-sibility that luminal molecules derived from the digestion ofdietary proteins act to stabilize an active receptor conformationby binding to segments of the extracellular loops or act anal-ogous to ago-allosteric modulators, i.e., exogenous compoundsacting both as agonists themselves and as enhancers for en-dogenous agonist(s), thereby influencing the potency of thatendogenous agonist activity (56).
ACKNOWLEDGMENTS
We thank Drs. Steven E. Ruzin and Denise E. Schichnes (BiologicalImaging Facility, University of California at Berkeley) for generously provid-ing expert technical help with LMD and confocal microscopy. We acknowl-edge the use of the GPCRDB information system (http://www.gpcr.org/7tm/).
GRANTS
This work was supported by National Institute of Diabetes and Digestiveand Kidney Diseases Grant DK-58592 (to G. W. Aponte).
REFERENCES
1. Adibi SA. Regulation of expression of the intestinal oligopeptide trans-porter (Pept-1) in health and disease. Am J Physiol Gastrointest LiverPhysiol 285: G779–G788, 2003.
2. Aponte GW, Keddie A, Hallden G, Hess R, Link P. Polarized intestinalhybrid cell lines derived from primary culture: establishment and charac-terization. Proc Natl Acad Sci USA 88: 5282–5286, 1991.
3. Aponte GW, Taylor IL, Soll AH. Primary culture of PYY cells fromcanine colon. Am J Physiol Gastrointest Liver Physiol 254: G829–G836,1988.
4. Ballesteros JA, Jensen AD, Liapakis G, Rasmussen SG, Shi L, GetherU, Javitch JA. Activation of the beta 2-adrenergic receptor involvesdisruption of an ionic lock between the cytoplasmic ends of transmem-brane segments 3 and 6. J Biol Chem 276: 29171–29177, 2001.
5. Biasco G, Callegari C, Lami F, Minarini A, Miglioli M, Barbara L.Intestinal morphological changes during oral refeeding in a patient previ-ously treated with total parenteral nutrition for small bowel resection. Am JGastroenterol 79: 585–588, 1984.
6. Boucher MJ, Rivard N. Regulation and role of brush border-associatedERK1/2 in intestinal epithelial cells. Biochem Biophys Res Commun 311:121–128, 2003.
7. Bretschneider B, Brandsch M, Neubert R. Intestinal transport of beta-lactam antibiotics: analysis of the affinity at the H�/peptide symporter(PEPT1), the uptake into Caco-2 cell monolayers and the transepithelialflux. Pharm Res 16: 55–61, 1999.
8. Chun J, Goetzl EJ, Hla T, Igarashi Y, Lynch KR, Moolenaar W, PyneS, Tigyi G. International Union of Pharmacology. XXXIV. Lysophospho-lipid receptor nomenclature. Pharmacol Rev 54: 265–269, 2002.
9. Conigrave AD, Quinn SJ, Brown EM. L-Amino acid sensing by theextracellular Ca2�-sensing receptor. Proc Natl Acad Sci USA 97: 4814–4819, 2000.
10. Conklin BR, Farfel Z, Lustig KD, Julius D, Bourne HR. Substitution ofthree amino acids switches receptor specificity of Gq alpha to that of Gi
alpha. Nature 363: 274–276, 1993.11. Cordier-Bussat M, Bernard C, Haouche S, Roche C, Abello J,
Chayvialle JA, Cuber JC. Peptones stimulate cholecystokinin secretionand gene transcription in the intestinal cell line STC-1. Endocrinology138: 1137–1144, 1997.
12. Cuber JC, Bernard G, Fushiki T, Bernard C, Yamanishi R, SugimotoE, Chayvialle JA. Luminal CCK-releasing factors in the isolated vascu-larly perfused rat duodenojejunum. Am J Physiol Gastrointest LiverPhysiol 259: G191–G197, 1990.
G110 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

13. Daniel H. Molecular and integrative physiology of intestinal peptidetransport. Annu Rev Physiol 66: 361–384, 2004.
14. Darcel NP, Liou AP, Tome D, Raybould HE. Activation of vagalafferents in the rat duodenum by protein requires PepT1. J Nutr 135:1491–1495, 2005.
15. Duplus E, Glorian M, Forest C. Fatty acid regulation of gene transcrip-tion. J Biol Chem 275: 30749–30752, 2000.
16. Eilers M, Hornak V, Smith SO, Konopka JB. Comparison of class Aand D G protein-coupled receptors: common features in structure andactivation. Biochemistry (Mosc) 44: 8959–8975, 2005.
17. Farrens DL, Altenbach C, Yang K, Hubbell WL, Khorana HG.Requirement of rigid-body motion of transmembrane helices for lightactivation of rhodopsin. Science 274: 768–770, 1996.
18. Gether U. Uncovering molecular mechanisms involved in activation of Gprotein-coupled receptors. Endocr Rev 21: 90–113, 2000.
19. Gevrey JC, Cordier-Bussat M, Nemoz-Gaillard E, Chayvialle JA,Abello J. Co-requirement of cyclic AMP- and calcium-dependent proteinkinases for transcriptional activation of cholecystokinin gene by proteinhydrolysates. J Biol Chem 277: 22407–22413, 2002.
20. Gevrey JC, Laurent S, Saurin JC, Nemoz-Gaillard E, Regazzi R,Chevrier AM, Chayvialle JA, Abello J. Rab3a controls exocytosis incholecystokinin-secreting cells. FEBS Lett 503: 19–24, 2001.
21. Grammer TC, Blenis J. Evidence for MEK-independent pathways reg-ulating the prolonged activation of the ERK-MAP kinases. Oncogene 14:1635–1642, 1997.
22. Groneberg DA, Doring F, Eynott PR, Fischer A, Daniel H. Intestinalpeptide transport: ex vivo uptake studies and localization of peptide carrierPEPT1. Am J Physiol Gastrointest Liver Physiol 281: G697–G704, 2001.
23. Groos S, Reale E, Hunefeld G, Luciano L. Changes in epithelial cellturnover and extracellular matrix in human small intestine after TPN.J Surg Res 109: 74–85, 2003.
24. Hallden G, Aponte GW. Evidence for a role of the gut hormone PYY inthe regulation of intestinal fatty acid-binding protein transcripts in differ-entiated subpopulations of intestinal epithelial cell hybrids. J Biol Chem272: 12591–12600, 1997.
25. Hallden G, Holehouse EL, Dong X, Aponte GW. Expression ofintestinal fatty acid binding protein in intestinal epithelial cell lines,hBRIE 380 cells. Am J Physiol Gastrointest Liver Physiol 267: G730 –G743, 1994.
26. Hama K, Bandoh K, Kakehi Y, Aoki J, Arai H. Lysophosphatidic acid(LPA) receptors are activated differentially by biological fluids: possiblerole of LPA-binding proteins in activation of LPA receptors. FEBS Lett523: 187–192, 2002.
27. Hofer D, Puschel B, Drenckhahn D. Taste receptor-like cells in the ratgut identified by expression of alpha-gustducin. Proc Natl Acad Sci USA93: 6631–6634, 1996.
28. Holst B, Holliday ND, Bach A, Elling CE, Cox HM, Schwartz TW.Common structural basis for constitutive activity of the ghrelin receptorfamily. J Biol Chem 279: 53806–53817, 2004.
29. Horn F, Bettler E, Oliveira L, Campagne F, Cohen FE, Vriend G.GPCRDB information system for G protein-coupled receptors. NucleicAcids Res 31: 294–297, 2003.
30. Hughes CA, Dowling RH. Speed of onset of adaptive mucosal hypoplasiaand hypofunction in the intestine of parenterally fed rats. Clin Sci (Lond)59: 317–327, 1980.
31. Jiang Y, Luo L, Gustafson EL, Yadav D, Laverty M, Murgolo N,Vassileva G, Zeng M, Laz TM, Behan J, Qiu P, Wang L, Wang S,Bayne M, Greene J, Monsma F Jr, Zhang FL. Identification andcharacterization of a novel RF-amide peptide ligand for orphan G-protein-coupled receptor SP9155. J Biol Chem 278: 27652–27657,2003.
32. Karnik SS, Gogonea C, Patil S, Saad Y, Takezako T. Activation ofG-protein-coupled receptors: a common molecular mechanism. TrendsEndocrinol Metab 14: 431–437, 2003.
33. Khoo S, Griffen SC, Xia Y, Baer RJ, German MS, Cobb MH.Regulation of insulin gene transcription by ERK1 and ERK2 in pancreaticbeta cells. J Biol Chem 278: 32969–32977, 2003.
34. Kim MS, Yoon CY, Jang PG, Park YJ, Shin CS, Park HS, Ryu JW,Pak YK, Park JY, Lee KU, Kim SY, Lee HK, Kim YB, Park KS. Themitogenic and antiapoptotic actions of ghrelin in 3T3-L1 adipocytes. MolEndocrinol 18: 2291–2301, 2004.
35. Kostenis E. Is Galpha16 the optimal tool for fishing ligands of orphanG-protein-coupled receptors? Trends Pharmacol Sci 22: 560–564, 2001.
36. Kostenis E, Zeng FY, Wess J. Functional characterization of a series ofmutant G protein alphaq subunits displaying promiscuous receptor cou-pling properties. J Biol Chem 273: 17886–17892, 1998.
37. Kotarsky K, Boketoft A, Bristulf J, Nilsson NE, Norberg A, HanssonS, Sillard R, Owman C, Leeb-Lundberg FL, Olde B. Lysophosphatidicacid binds to and activates GPR92, a G protein-coupled receptor highlyexpressed in gastrointestinal lymphocytes. J Pharmacol Exp Ther 318:619–628, 2006.
38. Lee CW, Rivera R, Gardell S, Dubin AE, Chun J. GPR92 as a newG12/13- and Gq-coupled lysophosphatidic increases cAMP, LPA5. J BiolChem 281: 23589–23597, 2006.
39. Lee DK, Nguyen T, Lynch KR, Cheng R, Vanti WB, Arkhitko O,Lewis T, Evans JF, George SR, O’Dowd BF. Discovery and mapping often novel G protein-coupled receptor genes. Gene 275: 83–91, 2001.
40. Lee M, Hadi M, Hallden G, Aponte GW. Peptide YY and neuropeptideY induce villin expression, reduce adhesion, and enhance migration insmall intestinal cells through the regulation of CD63, matrix metallopro-teinase-3, and Cdc42 activity. J Biol Chem 280: 125–136, 2005.
41. Levine GM, Deren JJ, Steiger E, Zinno R. Role of oral intake inmaintenance of gut mass and disaccharide activity. Gastroenterology 67:975–982, 1974.
42. Li Y, Owyang C. Peptone stimulates CCK-releasing peptide secretion byactivating intestinal submucosal cholinergic neurons. J Clin Invest 97:1463–1470, 1996.
43. Liddle RA. Cholecystokinin cells. Annu Rev Physiol 59: 221–242, 1997.44. Matsumura K, Miki T, Jhomori T, Gonoi T, Seino S. Possible role of
PEPT1 in gastrointestinal hormone secretion. Biochem Biophys Res Com-mun 336: 1028–1032, 2005.
45. Murai A, Noble PM, Deavall DG, Dockray GJ. Control of c-fosexpression in STC-1 cells by peptidomimetic stimuli. Eur J Pharmacol394: 27–34, 2000.
46. Neer EJ, Clapham DE. Roles of G protein subunits in transmembranesignalling. Nature 333: 129–134, 1988.
47. Nelson G, Chandrashekar J, Hoon MA, Feng L, Zhao G, Ryba NJ,Zuker CS. An amino-acid taste receptor. Nature 416: 199–202, 2002.
48. Nemoz-Gaillard E, Bernard C, Abello J, Cordier-Bussat M,Chayvialle JA, Cuber JC. Regulation of cholecystokinin secretion bypeptones and peptidomimetic antibiotics in STC-1 cells. Endocrinology139: 932–938, 1998.
49. Nishi T, Hara H, Hira T, Tomita F. Dietary protein peptic hydrolysatesstimulate cholecystokinin release via direct sensing by rat intestinalmucosal cells. Exp Biol Med (Maywood) 226: 1031–1036, 2001.
50. Noguchi K, Ishii S, Shimizu T. Identification of p2y9/GPR23 as a novelG protein-coupled receptor for lysophosphatidic acid, structurally distantfrom the Edg family. J Biol Chem 278: 25600–25606, 2003.
51. Parnot C, Miserey-Lenkei S, Bardin S, Corvol P, Clauser E. Lessonsfrom constitutively active mutants of G protein-coupled receptors. TrendsEndocrinol Metab 13: 336–343, 2002.
52. Poitout V, Hagman D, Stein R, Artner I, Robertson RP, Harmon JS.Regulation of the insulin gene by glucose and fatty acids. J Nutr 136:873–876, 2006.
53. Rizzuto R, Simpson AW, Brini M, Pozzan T. Rapid changes of mito-chondrial Ca2� revealed by specifically targeted recombinant aequorin.Nature 358: 325–327, 1992.
54. Schaeffer MT, Cully D, Chou M, Liu J, Van der Ploeg LH, FongTM. Use of bioluminescent aequorin for the pharmacological charac-terization of 5HT receptors. J Recept Signal Transduct Res 19: 927–938, 1999.
55. Schwartz TW, Frimurer TM, Holst B, Rosenkilde MM, Elling CE.Molecular mechanism of 7TM receptor activation—a global toggle switchmodel. Annu Rev Pharmacol Toxicol 46: 481–519, 2006.
56. Schwartz TW, Holst B. Ago-allosteric modulation and other types ofallostery in dimeric 7TM receptors. J Recept Signal Transduct Res 26:107–128, 2006.
57. Snabaitis AK, Muntendorf A, Wieland T, Avkiran M. Regulation ofthe extracellular signal-regulated kinase pathway in adult myocardium:differential roles of G(q/11), Gi and G(12/13) proteins in signalling byalpha1-adrenergic, endothelin-1 and thrombin-sensitive protease-activatedreceptors. Cell Signal 17: 655–664, 2005.
58. Stables J, Green A, Marshall F, Fraser N, Knight E, Sautel M,Milligan G, Lee M, Rees S. A bioluminescent assay for agonist activityat potentially any G-protein-coupled receptor. Anal Biochem 252: 115–126, 1997.
G111PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from

59. Tomura H, Mogi C, Sato K, Okajima F. Proton-sensing and lysolipid-sensitive G-protein-coupled receptors: a novel type of multi-functionalreceptors. Cell Signal 17: 1466–1476, 2005.
60. Vaulont S, Vasseur-Cognet M, Kahn A. Glucose regulation of genetranscription. J Biol Chem 275: 31555–31558, 2000.
61. Wellendorph P, Hansen KB, Balsgaard A, Greenwood JR, Egebjerg J,and Brauner-Osborne H. Deorphanization of GPRC6A: a promiscuousL-alpha-amino acid receptor with preference for basic amino acids. MolPharmacol 67: 589–597, 2005.
62. Whistler JL, Gerber BO, Meng EC, Baranski TJ, von Zastrow M,Bourne HR. Constitutive activation and endocytosis of the complementfactor 5a receptor: evidence for multiple activated conformations of a Gprotein-coupled receptor. Traffic 3: 866–877, 2002.
63. Whited KL, Thao D, Lloyd KC, Kopin AS, Raybould HE. Targeteddisruption of the murine CCK1 receptor gene reduces intestinal lipid-induced feedback inhibition of gastric function. Am J Physiol GastrointestLiver Physiol 291: G156–G162, 2006.
64. Williamson RC. Intestinal adaptation (first of two parts). Structural,functional and cytokinetic changes. N Engl J Med 298: 1393–1402, 1978.
65. Wu Q, Ortegon AM, Tsang B, Doege H, Feingold KR, Stahl A. FATP1is an insulin-sensitive fatty acid transporter involved in diet-inducedobesity. Mol Cell Biol 26: 3455–3467, 2006.
66. Wu SV, Rozengurt N, Yang M, Young SH, Sinnett-Smith J, Rozen-gurt E. Expression of bitter taste receptors of the T2R family in thegastrointestinal tract and enteroendocrine STC-1 cells. Proc Natl Acad SciUSA 99: 2392–2397, 2002.
G112 PROTEIN HYDROLYSATE RESPONSIVE GPCR
AJP-Gastrointest Liver Physiol • VOL 292 • JANUARY 2007 • www.ajpgi.org
on February 5, 2008
ajpgi.physiology.orgD
ownloaded from





























