Embed Size (px)
Citation preview

ANRV343-BB37-16 ARI 9 April 2008 11:38
Structure of EukaryoticRNA PolymerasesP. Cramer, K.-J. Armache, S. Baumli, S. Benkert,F. Brueckner, C. Buchen, G.E. Damsma, S. Dengl,S.R. Geiger, A.J. Jasiak, A. Jawhari, S. Jennebach,T. Kamenski, H. Kettenberger, C.-D. Kuhn,E. Lehmann, K. Leike, J.F. Sydow, and A. VanniniGene Center Munich and Center for Integrated Protein Science CIPSM, Departmentof Chemistry and Biochemistry, Ludwig-Maximilians-Universitat Munchen,81377 Munich, Germany; email: [email protected]
Annu. Rev. Biophys. 2008. 37:337–52
First published online as a Review in Advance onFebruary 7, 2008
The Annual Review of Biophysics is online atbiophys.annualreviews.org
This article’s doi:10.1146/annurev.biophys.37.032807.130008
Copyright c© 2008 by Annual Reviews.All rights reserved
1936-122X/08/0609-0337$20.00
Key Words
gene transcription, RNA synthesis, nucleic acid polymerase, X-raycrystallography, structural biology, multiprotein complex
AbstractThe eukaryotic RNA polymerases Pol I, Pol II, and Pol III are thecentral multiprotein machines that synthesize ribosomal, messenger,and transfer RNA, respectively. Here we provide a catalog of avail-able structural information for these three enzymes. Most structuraldata have been accumulated for Pol II and its functional complexes.These studies have provided insights into many aspects of the tran-scription mechanism, including initiation at promoter DNA, elon-gation of the mRNA chain, tunability of the polymerase active site,which supports RNA synthesis and cleavage, and the response ofPol II to DNA lesions. Detailed structural studies of Pol I and PolIII were reported recently and showed that the active center regionand core enzymes are similar to Pol II and that strong structuraldifferences on the surfaces account for gene class-specific functions.
337
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
Gene transcription:the process of RNAsynthesis based on aDNA template
DNA-dependentRNA polymerase:an enzyme that usesa DNA template tosynthesize acomplementary copyof RNA
Pol I, Pol II, andPol III: RNApolymerase I, II, III
Rpb: RNApolymerase B (II)subunit
Contents
INTRODUCTION. . . . . . . . . . . . . . . . . 338POL II STRUCTURE . . . . . . . . . . . . . . 338THE ELONGATION COMPLEX
AND NUCLEOTIDEINCORPORATION . . . . . . . . . . . . . 341
OVERCOMING OBSTACLESDURING ELONGATION . . . . . . 344
THE POL II INITIATIONCOMPLEX. . . . . . . . . . . . . . . . . . . . . . 345
STRUCTURAL STUDIESOF POL I . . . . . . . . . . . . . . . . . . . . . . . 345
STRUCTURAL STUDIESOF POL III . . . . . . . . . . . . . . . . . . . . . . 346
CONCLUSIONS ANDOUTLOOK . . . . . . . . . . . . . . . . . . . . . 346
INTRODUCTION
Gene transcription in eukaryotic cells iscarried out by the three different DNA-dependent RNA polymerases Pol I, Pol II,and Pol III. Pol I produces ribosomal RNA,Pol II synthesizes messenger RNAs and smallnuclear RNAs, and Pol III produces transferRNAs and other small RNAs. A fourth RNApolymerase, Pol IV, which was recently dis-covered in plants, is not included here, as itscomposition and structure are currently un-known. The RNA polymerases are multisub-unit enzymes. Pol I, II, and III comprise 14,12, and 17 subunits, respectively, and havea total molecular weight of 589, 514, and693 kDa, respectively. Ten subunits form astructurally conserved core, and additionalsubunits are located on the periphery. Com-prehensive reviews that summarized struc-tural studies of Pol II were published fouryears ago (21, 35). Here we provide an up-to-date catalog of structural studies of multisub-unit RNA polymerases to facilitate access tothe structural information. We emphasize re-cent structures and, in particular, recent struc-tural studies of Pol I and Pol III, which allowfor a first structural comparison of the threeRNA polymerases.
POL II STRUCTURE
Pol II consists of a 10-subunit core en-zyme and a peripheral heterodimer of sub-units Rpb4 and Rpb7 (Rpb4/7 subcomplex,Table 1). The core enzyme comprises sub-units Rpb1, Rpb2, Rpb3, and Rpb11, whichcontain regions of sequence and structuralsimilarity in Pol I, Pol III, bacterial RNApolymerases (72, 82), and the archaeal RNApolymerase (49). The Pol II core also com-prises subunits Rpb5, Rpb6, Rpb8, Rpb10,and Rpb12, which are shared between PolI, II, and III (common subunits, Table 1).Counterparts of these common subunits ex-cept Rpb8 exist in the archaeal polymerase,but only a counterpart of Rpb6 exists in thebacterial enzyme (60). Finally, homologuesof the core subunit Rpb9 exist in Pol I andPol III, but not in the archaeal or bacterialenzyme.
Initial electron microscopic studies of PolII revealed the overall shape of the enzyme(26) (Table 2). The core Pol II could subse-quently be crystallized, leading to an electrondensity map at 6 A resolution (32). Crystal im-provement by controlled shrinkage and phas-ing at 3 A resolution resulted in a backbonemodel of the Pol II core (23). This revealedthat Rpb1 and Rpb2 form opposite sides of apositively charged active center cleft, whereasthe smaller subunits are arrayed around theperiphery. Refined atomic structures of thecore Pol II were obtained in two differentconformations and revealed domain-like re-gions within the subunits, as well as surface el-ements predicted to have functional roles (24)(Figure 1 and Figure 2). The active site andthe bridge helix, which spans the cleft, linea pore in the floor of the cleft. The Rpb1side of the cleft forms a mobile clamp, whichwas trapped in two different open states inthe free core structures (24) but was closed inthe structure of a core complex that includedDNA and RNA (34). The mobile clamp isconnected to the body of the polymerase byfive switch regions that show conformationalvariability. The Rpb2 side of the cleft consists
338 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
Table 1 RNA polymerase subunits
RNA polymerase Pol I Pol II Pol IIITen-subunit core A190 Rpb1 C160
A135 Rpb2 C128
AC40 Rpb3 AC40
AC19 Rpb11 AC19
A12.2 Rpb9 C11
Rpb5 (ABC27) Rpb5 Rpb5
Rpb6 (ABC23) Rpb6 Rpb6
Rpb8 (ABC14.5) Rpb8 Rpb8
Rpb10 (ABC10α) Rpb10 Rpb10
Rpb12 (ABC10β) Rpb12 Rpb12Rpb4/7 subcomplex A14 Rpb4 C17
A43 Rpb7 C25TFIIF-like subcomplexa A49 (Tfg1/Rap74) C37
A34.5 (Tfg2/Rap30) C53Pol III-specific subcomplex – – C82
– – C34
– – C31Number of subunits 14 12 17
aThe two subunits in Pol I and Pol III are predicted to form heterodimers that resemble part of the Pol II initiation/elongation factor TFIIF, which is composed of subunits Tfg1, Tfg2, and Tfg3 in Saccharomyces cerevisiae, and of subunitsRap74 and Rap30 in human.
of the lobe and protrusion domains. Rpb2 alsoforms a protein wall that blocks the end of thecleft.
The Pol II core structures lacked sub-units Rpb4 and Rpb7, which can dissoci-ate from the yeast enzyme (28). A structureof the archaeal homologue of the Rpb4/7heterodimer showed that Rpb7 contains anN-terminal domain, later called the tip do-main, and a C-terminal domain that includesan oligosaccharide-binding (OB) fold (70).The approximate location of Rpb4/7 on thecore polymerase was first determined by elec-tron microscopy (EM) of two-dimensionalcrystals (40). Later, EM analysis of single par-ticles revealed a closed clamp and showed thatthe Rpb4/7 subcomplex protrudes from out-side the core enzyme below the clamp (20).A different open-closed transition that in-volved the polymerase jaws was observed byEM of two-dimensional crystals (5). Crystal-
EM: electronmicroscopy
lographic backbone models of the completePol II then revealed the exact position and ori-entation of Rpb4/7 and showed that it formeda wedge between the clamp and the linkerto the unique tail-like C-terminal repeat do-main (CTD) of the polymerase (2, 11). Thecrystal structure of free Rpb4/7 (Figure 1d ),together with an improved resolution of thecomplete Pol II crystals, finally enabled re-finement of a complete atomic model of PolII (3) (Figure 1b).
The CTD of Pol II is flexibly linkedto the core enzyme and consists of hep-tapeptide repeats of the consensus sequenceYSPTSPS. The CTD integrates nuclearevents by binding proteins involved in mRNAbiogenesis (for reviews see References 9, 37,and 56). CTD-binding proteins recognize aspecific CTD phosphorylation pattern, whichchanges during the transcription cycle. Struc-tural and functional studies of CTD-binding
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 339
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
Table 2 Structural information on eukaryotic RNA polymerases
MethodaResolutionc
(A) PDB code(s) Reference(s)Pol II subunitsRpb9 N-terminal domain NMR – 1qyp (76)Rpb10 NMR – 1ef4 (54)Rpb5 X-ray 1.9 1dzf (71)Rpb4/7 yeast X-ray 2.3 1y14 (3)Rpb4/7 human X-ray 2.7 2c35 (58)Core Pol IIb
Free EM (2D) 16.0 – (26)+TFIIB or TFIIE EM (2D) 15.7 – (52)Core ECb EM (2D) 20 – (62)Free X-ray 6.0 – (32)Free (form 1) X-ray 3.1 1i3q (23, 24)Free (form 2) X-ray 2.8 1i50 (24)EC (tailed-template) X-ray 3.3 1i6h (34)+α-amanitin X-ray 2.8 1k83 (10)Core EC X-ray 3.6 1sfo (81)Core EC +NTP (insertion site) X-ray 4.25 1r9s (80)Core EC +NTP (entry site) 3.5 1r9tFree +ATP X-ray 3.2 1twa (80)+GTP 3.0 1twc+UTP 2.3 1twf+CTP 3.3 1twg+2′dATP 3.4 1twhFree core, 150 mM Mn2+ X-ray 3.4 2nvy (77)Core EC +GTP, 5 mM Mg2+ 3.95 2e2h+2′dGTP, 5 mM Mg2+ 3.4 2e2i+GMPCPP, 5 mM Mg2+ 3.5 2e2j+2′dUTP, 150 mM Mg2+ 3.0 2nvq+GMPCPP, 150 mM Mg2+ 3.36 2nvt+2′dUTP, 5 mM Mg2+ 3.6 2nvx+UTP (updated) 4.3 2nvz+UTP, 150 mM Mg2+ 3.4 2yu9Core +TFIIB X-ray 4.5 1r5u (12)Complete Pol IIb
Free EM (2D) 24 – (40)Free EM (cryo)
18– (20)
Free (backbone) X-ray 4.2 1nt9 (2)Free (backbone) X-ray 4.1 1nik (11)Free (atomic) X-ray 3.8 1wcm (3)+TFIIF EM (cryo) 18 – (19)+TFIIS X-ray 3.8 1pqv (41)EC X-ray 4.0 1y1w (42)EC +TFIIS X-ray 3.8 1y1v (42)
(Continued )
340 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
Table 2 (Continued )
MethodaResolutionc
(A) PDB code(s) Reference(s)EC +NTP (inactive preinsertion-like state) X-ray 4.5 1y77 (42)CPD-damaged EC complex A X-ray 3.8 2ja5 (8)
complex B 4.0 2ja6complex C 3.8 2ja7complex D 3.8 2ja8
Cisplatin-damaged EC X-ray 3.8 2r7z (25)RNA template-product complex X-ray 3.8 2r92 (51)HDV RNA hairpin X-ray 4.0 2r93 (51)Free human Pol II EM 20 EMD-1283/1284 (46)Pol II CTD peptidesb
Free heptapeptide repeat NMR – – (36)Peptide with eight repeats NMR – – (13)Cyclic model peptides NMR – – (48)Complex with WW domain X-ray 1.84 1f8a (75)Complex with capping enzyme X-ray 2.7 1p16 (29)Complex with Pcf11 CIDb X-ray 2.2 1sza (55)
2.1 1sz9Complex with Scp1 X-ray 2.05 2ghq (83)
1.8 2ghtPol IFree EM (2D) 40 – (65)Free EM (cryo) 34 – (7)Free EM (cryo negative stain) 22 – (27)Free EM (cryo) 11.9 EMD-1435 (47)�A49/34.5 EM (cryo) 25 – (47)Core Homology model – – (47)A14/43 X-ray 3.10 2rf4 (47)Pol IIIFree EM (cryo) 17 EMD-1322 (30)Core Homology model – – (39)C17/25 X-ray 3.2 2ckz (39)
aX-ray, X-ray crystallography; NMR, nuclear magnetic resonance; EM, electron microscopy (2D, two-dimensional electron crystallography; stain,negative staining; cryo, single particle cryo-EM).bCore, 10-subunit Pol II core enzyme; complete, complete 12-subunit Pol II; EC, elongation complex; CTD, C-terminal repeat domain of Pol II;CID, C-terminal repeat domain interacting domain.cResolutions are not always comparable since different data cut-off criteria were used in different studies both in electron microscopy and X-raycrystallography.
and CTD-modifying proteins and theircomplexes with CTD peptides elucidatedCTD structure and revealed some ofthe mechanisms underlying CTD function(Table 2).
THE ELONGATION COMPLEXAND NUCLEOTIDEINCORPORATION
During the elongation phase of transcription,the polymerases move along a DNA template
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 341
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
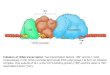
RNA polymerase I RNA polymerase II RNA polymerase III
A14/43 Rpb4/7
EM density
core Pol I model
C17/25
core Pol IIImodelcore Pol II
C53/37
C82/34/31
A49/34.5
EM density
C17/25 (Pol III)
HRDCdomain
Tip-associateddomain
OBdomain
Tipdomain
N
N C
A14/43 (Pol I)
C
N
Tip-associateddomain
Tipdomain
OBdomain
C
Rpb4/7 (Pol II)
H1H*
Tipdomain
N
N
C
C
Tip-associateddomain
HRDCdomain
OBdomain
C
a b c
d
Figure 1RNA polymerase structures. (a) Pol I hybrid structure (47). (b) Ribbon model of the refined completePol II crystal structure (3). (c) Pol III EM structure (30) with the Pol II-based homology model and thecrystal structure of C17/25 (39) fitted as published in Reference 30. (d ) Rpb4/7 subcomplex structures:A14/43 (left), Rpb4/7 (center) (3), C17/25 (right).
and synthesize a complementary chain of ri-bonucleotides. RNA extension begins withbinding of a nucleoside triphosphate (NTP)substrate to the transcription elongation com-plex (EC) that is formed by the polymerase,DNA, and RNA. Catalytic addition of the nu-cleotide to the growing RNA 3′ end then re-leases a pyrophosphate ion. Finally, transloca-tion of DNA and RNA frees the substrate sitefor binding of the next NTP. The EC is char-acterized by an unwound DNA region, thetranscription bubble. The bubble contains ashort hybrid duplex formed between the DNAtemplate strand and the RNA product emerg-ing from the active site.
The mechanism of RNA elongation waselucidated by structural studies of Pol II–
nucleic acid complexes (Figure 2). EM firstrevealed the point of DNA entry to the PolII cleft (62). The first crystal structure of aPol II–nucleic acid complex was that of thecore Pol II transcribing a tailed template DNA(34), which allows for promoter-independenttranscription initiation. This structure re-vealed downstream DNA entering the cleftand a 8 to 9 base pair DNA-RNA hybridin the active center. Comparison with thehigh-resolution core Pol II structure (24) re-vealed protein surface elements predicted toplay functional roles. Later, polymerase ECstructures utilized synthetic DNA-RNA scaf-folds (42, 81) and revealed the exact location ofthe downstream DNA and several nucleotidesupstream of the hybrid. Mechanisms were
342 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
B
A
Bridge helix
Wall Zipper
Switch 2
Switch 3 Fork loop 1
Fork loop 2
βDloopII
180°
Switch 1
a b
c
Transcription
Active sitemetal A DNA template
DNA non-templateRNA
NTP (insertion site)NTP (inactive pre-insertion state)Mismatched NTP (entry site)
Functional Pol II elements
Rudder
Trigger loopBridge helix
Figure 2Structure of the Pol II elongation complex (EC). (a) Overview of the EC structure (42, 77, 80).(b) Superposition of NTP-binding sites [red, insertion site; violet, entry site (42, 77, 80); pink, inactivepreinsertion-like state (42, 77, 80)]. (c) Functional Pol II surface elements in the EC.
suggested for how Pol II unwinds downstreamDNA and how it separates the RNA prod-uct from the DNA template at the end of thehybrid. Although Pol II generally uses DNAas a template, there is also evidence that PolII can use RNA templates. Recent structuresshowed that an RNA template-product du-plex can bind to the site normally occupiedby the DNA-RNA hybrid and provided thestructural basis for the phenomenon of RNA-dependent RNA synthesis by Pol II (51).
Additional structures of Pol II ECs in-cluded the NTP substrate (42, 77, 80). Thesestudies suggested how the polymerase se-lects the correct NTP and how it incorpo-
rates a nucleotide into RNA. The NTP wascrystallographically trapped in the insertionsite (77, 80), which is apparently occupiedduring catalysis, but also in an overlapping,slightly different location, suggesting an in-active NTP-bound preinsertion state of theenzyme (42). Both NTPs form Watson-Crickinteractions with a base in the DNA tem-plate strand. Binding of the NTP to theinsertion site involves folding of the trig-ger loop (77), a mobile part of the activecenter first observed in free bacterial RNApolymerase (72), and in the Pol II-TFIIScomplex (41). Folding of the trigger loopcloses the active site and may be involved in
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 343
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
selection of the correct NTP. The NTP com-plex structures revealed contacts of the nu-cleotide with the polymerase, which explaindiscrimination of ribonucleotides against de-oxyribonucleotides, and provided insightsinto the selection of the nucleotide comple-mentary to the templating DNA base.
Catalytic nucleotide incorporation appar-ently follows a two-metal ion mechanism sug-gested for all polymerases (69). The Pol II ac-tive site contains a persistently bound metalion (metal A) and a second, mobile metal ion(metal B) (24). Metal A is held by three invari-ant aspartate side chains and binds the RNA3′ end (24), whereas metal B binds the NTPtriphosphate moiety (80). Recent studies offunctional complexes of the bacterial RNApolymerase revealed the close conservationof the EC structure (73) and provided addi-tional insights into nucleotide incorporation(74). As for Pol II, NTP binding to the inser-tion site can induce folding of the trigger loop.In the presence of the antibiotic streptolydi-gin, however, the NTP binds in the inactive,preinsertion state, in which the triphosphateand metal B are too far from metal A to per-mit catalysis. This finding supported a two-step mechanism of nucleotide incorporation(42, 74). The NTP would first bind in theinactive state to an open active center confor-mation. Complete folding of the trigger loopthen leads to closure of the active center, de-livery of the NTP to the insertion site, andcatalysis. An alternative model for nucleotideaddition involves binding of the NTP to a pu-tative entry site in the pore, in which the nu-cleotide base is oriented away from the DNAtemplate, and rotation of the NTP aroundmetal ion B directly into the insertion site(80).
OVERCOMING OBSTACLESDURING ELONGATION
During active transcription, Pol II must over-come intrinsic DNA arrest sites, which aregenerally rich in A-T base pairs and posea natural obstacle to transcription. At such
sites, Pol II moves backward along DNA andRNA, resulting in extrusion of the RNA 3′
end through the polymerase pore beneaththe active site and transcriptional arrest. TheRNA cleavage stimulatory factor TFIIS canrescue an arrested polymerase by creating anew RNA 3′ end at the active site from whichtranscription can resume. The mechanism ofTFIIS function was elucidated with the struc-tures of Pol II and a Pol II EC in complexwith TFIIS (41, 42). TFIIS inserts a hairpininto the polymerase pore and complementsthe active site with acidic residues, changesthe enzyme conformation, and repositions theRNA transcript (41, 42). These studies sup-ported the idea that the Pol II active site istunable, as it can catalyze different reactions,including RNA synthesis and RNA cleavage(41, 68).
Other obstacles to transcription are bulkylesions in the DNA template strand. Re-cent structural studies of Pol II ECs thatcontain lesions in the template strandunraveled the mechanisms of polymerasestalling at two different bulky lesions, a UVlight-induced thymine-thymine cyclobutanepyrimidine dimer (CPD) (8) and a guanine-guanine intrastrand cross-link induced by theanticancer drug cisplatin (25). Cells efficientlyeliminate CPDs and cisplatin cross-links bytranscription-coupled DNA repair (TCR),which begins with Pol II stalling at the le-sion. TCR continues with assembly of the nu-cleotide excision repair machinery, removal ofa lesion-containing DNA fragment, and re-pair of the resulting DNA gap. The structuralstudies revealed that the structure of the EC isgenerally not influenced by the presence of thelesions, except for some changes in the down-stream DNA, arguing against allosteric mod-els for the assembly of the repair machineryduring TCR. The studies also demonstrateda translocation barrier for bulky dinucleotidelesions that impairs their delivery to the activesite.
The important conclusion from thesestudies was that the detailed mechanismsof transcriptional stalling at two different
344 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
dinucleotide lesions differ. The cisplatin le-sion cannot overcome the translocation bar-rier, leading to polymerase stalling. However,the CPD can overcome the barrier and stablybinds in the active site. Inefficient AMP incor-poration occurs at both lesions by an appar-ently template-independent mechanism, butonly for the CPD does this enable bindingof the lesion at the active site via formationof a stable thymine-adenine base pair. Op-posite the CPD, UMP misincorporation oc-curs, which finally stalls Pol II because theresulting mismatch impairs further translo-cation of nucleic acids. These studies alsoshowed that Pol II can, under certain artificialconditions, bypass a distorting dinucleotidelesion, and suggested that it is currently im-possible to predict the mechanisms of tran-scriptional stalling or mutagenesis at othertypes of lesions.
THE POL II INITIATIONCOMPLEX
For transcription initiation, Pol II assembleswith the initiation factors TFIIB, -D, -E,-F, and -H into an initiation complex at pro-moter DNA. Our current understanding ofthe architecture of the initiation complex wasreviewed recently (4, 22, 35). Briefly, cur-rent models of initiation assume that the pro-moter DNA is first bound outside the Pol IIcleft. Upon DNA melting, the template sin-gle strand is predicted to slip inside the Pol IIcleft and bind near the active site for the de-termination of the transcription start site andthe initiation of RNA synthesis. The mech-anisms underlying these events, however, arepoorly understood.
The location of initiation factors alongpromoter DNA was revealed by site-specificDNA-protein cross-linking (6, 15, 44, 59).With the use of photo-cross-linking and radi-cal probing, the N-terminal domain of TFIIBwas located to the polymerase dock domain(16), and the C-terminal domain of TFIIBwas placed over the polymerase cleft and wall(17). Crystallographic analysis of the Pol II
core-TFIIB complex confirmed that the N-terminal TFIIB zinc ribbon domain bindsthe dock domain and showed that a regionbetween the N- and C-terminal domains ofTFIIB, the B-finger, extended into the activecenter (12). EM of the Pol II-TFIIF complexsuggested that the second largest TFIIF sub-unit extends along the cleft, whereas densityat the Rpb4/7 subcomplex was attributed tothe TFIIF largest subunit (19). A recent cross-linking study revealed an interaction of TFIIFwith the opposite side of the cleft (18). It islikely that TFIIF, similar to TFIIB, extendswith a loop into the active center (17, 31).Early EM placed the initiation factor TFIIEat the downstream jaws of Pol II (52), whereascross-linking studies recently found an inter-action of TFIIE with the clamp domain (18).TFIIH is located near downstream DNA (44,59). Finally, studies of a complete Pol II com-plex with an RNA aptamer showed how tran-scription initiation can be inhibited by a smallRNA (43). The RNA inhibitor binds the ac-tive center cleft and apparently prevents load-ing of the DNA into the cleft during initiation(43).
STRUCTURAL STUDIESOF POL I
Compared with Pol II, little structural infor-mation is available for Pol I and Pol III. Thefold of the 10-subunit core enzymes is con-served to a large extent, and the active centerregions are highly similar in all three enzymes.Pol I and Pol III also contain counterparts ofthe Rpb4/7 subcomplex, A14/43 and C17/25,respectively, but there is hardly any sequenceconservation between the corresponding sub-units in these heterodimers (38, 57, 61, 63, 66,67). Pol I further contains the specific sub-units A49 and A34.5, and Pol III additionallycontains the specific subunits C82, C53, C37,C34, and C31.
For Pol I, the overall shape and dimen-sions were first revealed by EM analysisof two-dimensional crystals (65). Subsequentcryo-EM at 34 A resolution [Fourier shell
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 345
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
correlation (FSC) = 0.5] visualized a stalkcontaining A14/43 and densities for A49 andA34.5 over the central cleft (7, 61). Later EManalysis with cryo-negative staining at 22 A(FSC = 0.5) confirmed the stalk but not thelocation of A49 and A34.5 (27). Recently, a12 A (FSC = 0.5) cryo-EM structure wasreported for yeast Pol I, as well as a ho-mology model for the core enzyme, and thecrystal structure of the subcomplex A14/43(47). In the resulting hybrid structure of Pol I(Figure 1a), A14/43, the clamp, and thedock domain contribute to a unique sur-face interacting with promoter-specific ini-tiation factors, in particular Rrn3. A14/43lacks the HRDC domain present in Rpb4/7and C17/25 (Figure 1d ). Subunits A49 andA34.5 form a heterodimer near the enzymefunnel that is important for normal elonga-tion activity and is predicted to be partly re-lated to the Pol II initiation/elongation factorTFIIF.
STRUCTURAL STUDIESOF POL III
An interaction map between Pol III subunitswas derived by yeast two-hybrid analysis andcopurification assays (reviewed in References14, 33, 64). The five Pol III–specific subunitsapparently form two subcomplexes. SubunitsC82, C34, and C31 form a trimeric subcom-plex involved in initiation (78, 79), whereasC37 and C53 form another subcomplex in-volved in termination and reinitiation (30, 50,53).
The first structural information for Pol IIIhas recently become available and includes ahomology model for the 10-subunit core en-zyme (39), the crystal structure of the C17/25subcomplex (39), and a 17 A EM structure(30) (Figure 1c,d ). Compared with Pol I andPol II, Pol III shows a structurally differentupstream face for specific initiation complexassembly during promoter selection. The PolIII upstream face includes the C82/34/31 sub-complex (30), which is involved in initiation,
and a HRDC domain in subunit C17 thatadopts a position different from its Pol IIcounterpart, Rpb4/7 (39). The C37/53 het-erodimer may adopt a position on the out-side of the core enzyme near the lobe andfunnel (30) and may be the counterpart ofthe A49/34.5 heterodimer in Pol I (47). Massspectrometry is consistent with a subcomplexarchitecture of Pol III that includes a 10-subunit Pol II-like core; the peripheral het-erodimers C17/25, C37/53, and C82/34; andsubunit C31, which bridges between C82/34,C17/25, and the core (53).
CONCLUSIONS AND OUTLOOK
The complexity and large size of multisub-unit RNA polymerases have prevented elu-cidation of their structure for a long time.The first detailed insights into the architec-ture and possible mechanism of multisub-unit RNA polymerases were obtained aroundthe millennium, when the core Pol II crystalstructures (23, 24, 34) and structural studies ofthe related bacterial enzyme (45, 82) becameavailable. In the following years, the discoveryof many additional structures of Pol II com-plexes, and structures of the bacterial enzymethat are not reviewed here, elucidated differ-ent aspects of transcription. Only over the pasttwo years, more detailed structural informa-tion for Pol I and Pol III became available andrevealed similarities and differences betweenthe three eukaryotic RNA polymerases thatemerged to match different functional prop-erties and requirements. To explain the de-tailed differences in structure and function ofthese enzymes, it is necessary to also inves-tigate Pol I and Pol III crystallographically,which is a tremendous experimental chal-lenge. In addition, many aspects of the Pol IItranscription mechanism remain unresolved,and more complex structures are required toclarify them. Among these open issues are themechanisms of initial promoter binding andmelting, initial transcription, translocation ofnucleic acids after nucleotide addition, and
346 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
termination, to name a few. A long-term goalof structural biology on RNA polymerasesremains to obtain clues for how these en-
zymes are regulated by coregulatory assem-blies, which transfer signals from transcrip-tional activators and repressors.
SUMMARY POINTS
1. Crystallographic studies of Pol II have elucidated the structure and conformationalflexibility of a central biological multiprotein machine.
2. Structural studies of Pol II complexes with nucleic acids revealed the location of theDNA template, the RNA product, and the NTP substrate, as well as elucidated manyaspects of the transcription mechanism.
3. Structural studies of Pol II complexes with protein factors provided insights into tran-scription initiation and characterized the enzyme’s tunable active site, which catalyzesRNA synthesis and cleavage.
4. Recent structural studies of Pol I and Pol III revealed structural differences betweenthe three eukaryotic RNA polymerases that account for functional differences, andenabled a detailed comparative structure-function analysis.
FUTURE ISSUES
1. What is the mechanism of nucleic acid translocation over the polymerase surface?How can structural and functional data on the nucleotide addition cycle be combinedinto a movie of the transcribing polymerase?
2. How can the crystal structures of Pol I and Pol III be determined to elucidate thedetailed structural differences between eukaryotic RNA polymerases and to explainenzyme-specific properties and regulation?
3. What is the three-dimensional structure of the Pol II transcription initiation complex,and what does this tell us about promoter recognition, loading, and melting?
4. What are the mechanisms of transcriptional regulation? How do transcription factorschange the amount of RNAs produced by the polymerases?
DISCLOSURE STATEMENT
The authors are not aware of any biases that might be perceived as affecting the objectivity ofthis review.
ACKNOWLEDGMENTS
We apologize to the many colleagues whose work was not mentioned because of the specialfocus of this review on structural studies of eukaryotic RNA polymerases and because of spacerestraints. This work was supported by the Deutsche Forschungsgemeinschaft, the Nanosys-tems Initiative Munich (NIM), the EU Network 3D Repertoire, the Elitenetzwerk Bayern,
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 347
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
the Boehringer-Ingelheim Fonds, the EU Marie-Curie training program, and the Fonds derChemischen Industrie.
NOTE ADDED IN PROOF
While this review was in production the first crystal structure of an archaeal RNA polymerasebecame available (Hirata A, Klein BJ, Murakami KS. 2008. The X-ray crystal structure of RNApolymerase from archaea. Nature 451:851–54). This work confirmed the conclusions from acryo-EM structure of another archaeal RNA polymerase (49) and provided many additionalstructural details.
LITERATURE CITED
1. Alic N, Ayoub N, Landrieux E, Favry E, Baudouin-Cornu P, et al. 2007. Selectivity andproofreading both contribute significantly to the fidelity of RNA polymerase III transcrip-tion. Proc. Natl. Acad. Sci. USA 104:10400–5
2. Armache K-J, Kettenberger H, Cramer P. 2003. Architecture of the initiation-competent12-subunit RNA polymerase II. Proc. Natl. Acad. Sci. USA 100:6964–68
3. Refined crystalstructure of thecomplete12-subunit Pol II.
3. Armache K-J, Mitterweger S, Meinhart A, Cramer P. 2005. Structures of completeRNA polymerase II and its subcomplex, Rpb4/7. J. Biol. Chem. 280:7131–34
4. Asturias FJ. 2004. Another piece in the transcription initiation puzzle. Nat. Struct. Mol. Biol.11:1031–33
5. Asturias FJ, Meredith GD, Poglitsch CL, Kornberg RD. 1997. Two conformations of RNApolymerase II revealed by electron crystallography. J. Mol. Biol. 272:536–40
6. Bartlett MS, Thomm M, Geiduschek EP. 2000. The orientation of DNA in an archaealtranscription initiation complex. Nat. Struct. Biol. 7:782–85
7. Bischler N, Brino L, Carles C, Riva M, Tschochner H, et al. 2002. Localization of the yeastRNA polymerase I-specific subunits. EMBO J. 21:4136–44
8. Brueckner F, Hennecke U, Carell T, Cramer P. 2007. CPD damage recognition by tran-scribing RNA polymerase II. Science 315:859–62
9. Buratowski S. 2003. The CTD code. Nat. Struct. Biol. 10:679–8010. Bushnell DA, Cramer P, Kornberg RD. 2002. Structural basis of transcription: alpha-
amanitin-RNA polymerase II cocrystal at 2.8 A resolution. Proc. Natl. Acad. Sci. USA99:1218–22
11. Bushnell DA, Kornberg RD. 2003. Complete RNA polymerase II at 4.1 A resolution:implications for the initiation of transcription. Proc. Natl. Acad. Sci. USA 100:6969–73
12. Bushnell DA, Westover KD, Davis RE, Kornberg RD. 2004. Structural basis of transcrip-tion: an RNA polymerase II-TFIIB cocrystal at 4.5 Angstroms. Science 303:983–88
13. Cagas PM, Corden JL. 1995. Structural studies of a synthetic peptide derived from thecarboxyl-terminal domain of RNA polymerase II. Proteins 21:149–60
14. Chedin S, Ferri ML, Peyroche G, Andrau JC, Jourdain S, et al. 1998. The yeast RNApolymerase III transcription machinery: a paradigm for eukaryotic gene activation. ColdSpring Harb. Symp. Quant. Biol. 63:381–89
15. Chen BS, Mandal SS, Hampsey M. 2004. High-resolution protein-DNA contacts for theyeast RNA polymerase II general transcription machinery. Biochemistry 43:12741–49
16. Chen HT, Hahn S. 2003. Binding of TFIIB to RNA polymerase II: mapping the bindingsite for the TFIIB zinc ribbon domain within the preinitiation complex. Mol. Cell 12:437–47
348 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
17. Chen HT, Hahn S. 2004. Mapping the location of TFIIB within the RNA polymerase IItranscription preinitiation complex: a model for the structure of the PIC. Cell 119:169–80
18. Chen HT, Warfield L, Hahn S. 2007. The positions of TFIIF and TFIIE in the RNApolymerase II transcription preinitiation complex. Nat. Struct. Mol. Biol. 14:696–703
19. Chung WH, Craighead JL, Chang WH, Ezeokonkwo C, Bareket-Samish A, et al. 2003.RNA polymerase II/TFIIF structure and conserved organization of the initiation complex.Mol. Cell 12:1003–13
20. Craighead JL, Chang WH, Asturias FJ. 2002. Structure of yeast RNA polymerase II insolution: implications for enzyme regulation and interaction with promoter DNA. Struct.Fold. Des. 10:1117–25
21. Cramer P. 2004. Structure and function of RNA polymerase II. Adv. Protein Chem. 67:1–4222. Cramer P. 2007. Finding the right spot to start transcription. Nat. Struct. Mol. Biol. 14:686–
8723. Cramer P, Bushnell DA, Fu J, Gnatt AL, Maier-Davis B, et al. 2000. Architecture of
RNA polymerase II and implications for the transcription mechanism. Science 288:640–53
24. Refined crystalstructure of the10-subunit Pol IIcore enzyme.
24. Cramer P, Bushnell DA, Kornberg RD. 2001. Structural basis of transcription: RNApolymerase II at 2.8 angstrom resolution. Science 292:1863–76
25. Damsma GE, Alt A, Brueckner F, Carell T, Cramer P. 2007. Mechanism of transcriptionalstalling at cisplatin-damaged DNA. Nat. Struct. Mol. Biol. 14:1127–33
26. Darst SA, Edwards AM, Kubalek EW, Kornberg RD. 1991. Three-dimensional structureof yeast RNA polymerase II at 16 A resolution. Cell 66:121–28
27. De Carlo S, Carles C, Riva M, Schultz P. 2003. Cryo-negative staining reveals conforma-tional flexibility within yeast RNA polymerase I. J. Mol. Biol. 329:891–902
28. Edwards AM, Kane CM, Young RA, Kornberg RD. 1991. Two dissociable subunits of yeastRNA polymerase II stimulate the initiation of transcription at a promoter in vitro. J. Biol.Chem. 266:71–75
29. Fabrega C, Shen V, Shuman S, Lima CD. 2003. Structure of an mRNA capping enzymebound to the phosphorylated carboxy-terminal domain of RNA polymerase II. Mol. Cell11:1549–61
30. Pol III EMstructure.
30. Fernandez-Tornero C, Bottcher B, Riva M, Carles C, Steuerwald U, et al. 2007.Insights into transcription initiation and termination from the electron microscopystructure of yeast RNA polymerase III. Mol. Cell 25:813–23
31. Freire-Picos MA, Krishnamurthy S, Sun ZW, Hampsey M. 2005. Evidence that theTfg1/Tfg2 dimer interface of TFIIF lies near the active center of the RNA polymeraseII initiation complex. Nucleic Acids Res. 33:5045–52
32. Fu J, Gnatt AL, Bushnell DA, Jensen GJ, Thompson NE, et al. 1999. Yeast RNA polymeraseII at 5 A resolution. Cell 98:799–810
33. Geiduschek EP, Kassavetis GA. 2001. The RNA polymerase III transcription apparatus.J. Mol. Biol. 310:1–26
34. First structureof a core Pol IIelongationcomplex.
34. Gnatt AL, Cramer P, Fu J, Bushnell DA, Kornberg RD. 2001. Structural basis oftranscription: an RNA polymerase II elongation complex at 3.3 A resolution. Science
292:1876–8235. Hahn S. 2004. Structure and mechanism of the RNA polymerase II transcription machinery.
Nat. Struct. Mol. Biol. 11:394–40336. Harding MM. 1992. NMR studies on YSPTSPSY: implications for the design of DNA
bisintercalators. J. Med. Chem. 35:4658–6437. Hirose Y, Manley JL. 2000. RNA polymerase II and the integration of nuclear events. Genes
Dev. 14:1415–29
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 349
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
38. Hu P, Wu S, Sun Y, Yuan CC, Kobayashi R, et al. 2002. Characterization of humanRNA polymerase III identifies orthologues for Saccharomyces cerevisiae RNA polymerase IIIsubunits. Mol. Cell Biol. 22:8044–55
39. Pol III corehomology modeland crystalstructure ofC17/25.
39. Jasiak AJ, Armache KJ, Martens B, Jansen RP, Cramer P. 2006. Structural biologyof RNA polymerase III: subcomplex C17/25 X-ray structure and 11 subunit enzymemodel. Mol. Cell 23:71–81
40. Jensen GJ, Meredith G, Bushnell DA, Kornberg RD. 1998. Structure of wild-type yeastRNA polymerase II and location of Rpb4 and Rpb7. EMBO J. 17:2353–58
41. Kettenberger H, Armache K-J, Cramer P. 2003. Architecture of the RNA polymeraseII-TFIIS complex and implications for mRNA cleavage. Cell 114:347–57
42. Structure of thecomplete Pol IIelongationcomplex.
42. Kettenberger H, Armache K-J, Cramer P. 2004. Complete RNA polymerase II elon-gation complex structure and its interactions with NTP and TFIIS. Mol. Cell 16:955–65
43. Kettenberger H, Eisenfuhr A, Brueckner F, Theis M, Famulok M, Cramer P. 2006. Struc-ture of an RNA polymerase II-RNA inhibitor complex elucidates transcription regulationby noncoding RNAs. Nat. Struct. Mol. Biol. 13:44–48
44. Kim TK, Ebright RH, Reinberg D. 2000. Mechanism of ATP-dependent promoter meltingby transcription factor IIH. Science 288:1418–22
45. Korzheva N, Mustaev A, Kozlov M, Malhotra A, Nikiforov V, et al. 2000. A structuralmodel of transcription elongation. Science 289:619–25
46. Kostek SA, Grob P, De Carlo S, Lipscomb JS, Garczarek F, Nogales E. 2006. Molec-ular architecture and conformational flexibility of human RNA polymerase II. Structure14:1691–700
47. Pol I hybridstructure,including the 12 AEM structure, ahomology modelfor the coreenzyme, and thecrystal structure ofA14/43.
47. Kuhn C-D, Geiger SR, Baumli S, Gartmann M, Jennebach S, et al. 2007. Functionalarchitecture of RNA polymerase I. Cell 131:1260–72
48. Kumaki Y, Matsushima N, Yoshida H, Nitta K, Hikichi K. 2001. Structure of the YSPTSPSrepeat containing two SPXX motifs in the CTD of RNA polymerase II: NMR studies ofcyclic model peptides reveal that the SPTS turn is more stable than SPSY in water. Biochim.Biophys. Acta 1548:81–93
49. Kusser A, Bertero M, Naji S, Becker T, Thomm M, et al. 2007. Structure of an archaealRNA polymerase. J. Mol. Biol. doi:10.1016/j.jmb.2007.08.066
50. Landrieux E, Alic N, Ducrot C, Acker J, Riva M, Carles C. 2006. A subcomplex of RNApolymerase III subunits involved in transcription termination and reinitiation. EMBO J.25:118–28
51. Lehmann E, Brueckner F, Cramer P. 2007. Molecular basis of RNA-dependent RNApolymerase II activity. Nature 450:445–49
52. Leuther KK, Bushnell DA, Kornberg RD. 1996. Two-dimensional crystallography of tran-scription factor IIB- and IIE-RNA polymerase II complexes: implications for start siteselection and initiation complex formation. Cell 85:773–79
53. Lorenzen K, Vannini A, Cramer P, Heck AJ. 2007. Structural biology of RNA polymeraseIII: Mass spectrometry elucidates subcomplex architecture. Struct. Fold. Des. 15:1237–45
54. Mackereth CD, Arrowsmith CH, Edwards AM, McIntosh LP. 2000. Zinc-bundle structureof the essential RNA polymerase subunit RPB10 from Methanobacterium thermoautotroph-icum. Proc. Natl. Acad. Sci. USA 97:6316–21
55. Meinhart A, Cramer P. 2004. Recognition of RNA polymerase II carboxy-terminal domainby 3′-RNA-processing factors. Nature 430:223–26
56. Meinhart A, Kamenski T, Hoeppner S, Baumli S, Cramer P. 2005. A structural perspectiveof CTD function. Genes Dev. 19:1401–15
350 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
57. Meka H, Daoust G, Arnvig KB, Werner F, Brick P, Onesti S. 2003. Structural and functionalhomology between the RNAP(I) subunits A14/A43 and the archaeal RNAP subunits E/F.Nucleic Acids Res. 31:4391–400
58. Meka H, Werner F, Cordell SC, Onesti S, Brick P. 2005. Crystal structure and RNA bindingof the Rpb4/Rpb7 subunits of human RNA polymerase II. Nucleic Acids Res. 33:6435–44
59. Miller G, Hahn S. 2006. A DNA-tethered cleavage probe reveals the path for promoterDNA in the yeast preinitiation complex. Nat. Struct. Mol. Biol. 13:603–10
60. Minakhin L, Bhagat S, Brunning A, Campbell EA, Darst SA, et al. 2001. Bacterial RNApolymerase subunit omega and eukaryotic RNA polymerase subunit RPB6 are sequence,structural, and functional homologs and promote RNA polymerase assembly. Proc. Natl.Acad. Sci. USA 98:892–97
61. Peyroche G, Levillain E, Siaut M, Callebaut I, Schultz P, et al. 2002. The A14-A43 het-erodimer subunit in yeast RNA pol I and their relationship to Rpb4-Rpb7 pol II subunits.Proc. Natl. Acad. Sci. USA 99:14670–75
62. Poglitsch CL, Meredith GD, Gnatt AL, Jensen GJ, Chang WH, et al. 1999. Electron crystalstructure of an RNA polymerase II transcription elongation complex. Cell 98:791–98
63. Sadhale PP, Woychik NA. 1994. C25, an essential RNA polymerase III subunit related tothe RNA polymerase II subunit RPB7. Mol. Cell Biol. 14:6164–70
64. Schramm L, Hernandez N. 2002. Recruitment of RNA polymerase III to its target pro-moters. Genes Dev. 16:2593–620
65. Schultz P, Celia H, Riva M, Sentenac A, Oudet P. 1993. Three-dimensional model of yeastRNA polymerase I determined by electron microscopy of two-dimensional crystals. EMBOJ. 12:2601–7
66. Shpakovski GV, Shematorova EK. 1999. Characterization of the rpa43+ cDNA ofSchizosaccharomyces pombe: structural similarity of subunit Rpa43 of RNA polymerase I andsubunit Rpc25 of RNA polymerase III. Russ. J. Bioorgan. Chem. 25:700–5
67. Siaut M, Zaros C, Levivier E, Ferri ML, Court M, et al. 2003. An Rpb4/Rpb7-like complexin yeast RNA polymerase III contains the orthologue of mammalian CGRP-RCP. Mol. CellBiol. 23:195–205
68. Sosunov V, Sosunova E, Mustaev A, Bass I, Nikiforov V, Goldfarb A. 2003. Unifiedtwo-metal mechanism of RNA synthesis and degradation by RNA polymerase. EMBOJ. 22:2234–44
69. Steitz TA. 1998. A mechanism for all polymerases. Nature 391:231–3270. Todone F, Brick P, Werner F, Weinzierl RO, Onesti S. 2001. Structure of an archaeal
homolog of the eukaryotic RNA polymerase II RPB4/RPB7 complex. Mol. Cell 8:1137–4371. Todone F, Weinzierl RO, Brick P, Onesti S. 2000. Crystal structure of RPB5, a universal
eukaryotic RNA polymerase subunit and transcription factor interaction target. Proc. Natl.Acad. Sci. USA 97:6306–10
72. Vassylyev DG, Sekine S, Laptenko O, Lee J, Vassylyeva MN, et al. 2002. Crystal structureof a bacterial RNA polymerase holoenzyme at 2.6 A resolution. Nature 417:712–19
73. Vassylyev DG, Vassylyeva MN, Perederina A, Tahirov TH, Artsimovitch I. 2007. Structuralbasis for transcription elongation by bacterial RNA polymerase. Nature 448:157–62
74. Vassylyev DG, Vassylyeva MN, Zhang J, Palangat M, Artsimovitch I, Landick R. 2007.Structural basis for substrate loading in bacterial RNA polymerase. Nature 448:163–68
75. Verdecia MA, Bowman ME, Lu KP, Hunter T, Noel JP. 2000. Structural basis forphosphoserine-proline recognition by group IV WW domains. Nat. Struct. Biol. 7:639–43
www.annualreviews.org • Structure of Eukaryotic RNA Polymerases 351
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-16 ARI 9 April 2008 11:38
76. Wang B, Jones DN, Kaine BP, Weiss MA. 1998. High-resolution structure of an archaealzinc ribbon defines a general architectural motif in eukaryotic RNA polymerases. Structure6:555–69
77. Wang D, Bushnell DA, Westover KD, Kaplan CD, Kornberg RD. 2006. Structural basis oftranscription: role of the trigger loop in substrate specificity and catalysis. Cell 127:941–54
78. Wang Z, Roeder RG. 1997. Three human RNA polymerase III-specific subunits form asubcomplex with a selective function in specific transcription initiation. Genes Dev. 11:1315–26
79. Werner M, Hermann-Le Denmat S, Treich I, Sentenac A, Thuriaux P. 1992. Effect ofmutations in a zinc-binding domain of yeast RNA polymerase C (III) on enzyme functionand subunit association. Mol. Cell Biol. 12:1087–95
80. Extendedstructure of thecore Pol IIelongation complexwith NTPsubstrate bound.
80. Westover KD, Bushnell DA, Kornberg RD. 2004. Structural basis of transcrip-tion: nucleotide selection by rotation in the RNA polymerase II active center. Cell
119:481–8981. Westover KD, Bushnell DA, Kornberg RD. 2004. Structural basis of transcription: sepa-
ration of RNA from DNA by RNA polymerase II. Science 303:1014–1682. Zhang G, Campbell EA, Minakhin L, Richter C, Severinov K, Darst SA. 1999. Crystal
structure of Thermus aquaticus core RNA polymerase at 3.3 A resolution. Cell 98:811–2483. Zhang Y, Kim Y, Genoud N, Gao J, Kelly JW, et al. 2006. Determinants for dephospho-
rylation of the RNA polymerase II C-terminal domain by Scp1. Mol. Cell 24:759–70
352 Cramer et al.
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

AR343-FM ARI 10 April 2008 7:2
Annual Review ofBiophysics
Volume 37, 2008
Contents
FrontispieceRobert L. Baldwin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �xiv
The Search for Folding Intermediates and the Mechanismof Protein FoldingRobert L. Baldwin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1
How Translocons Select Transmembrane HelicesStephen H. White and Gunnar von Heijne � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 23
Unique Rotary ATP Synthase and Its Biological DiversityChristoph von Ballmoos, Gregory M. Cook, and Peter Dimroth � � � � � � � � � � � � � � � � � � � � � � � � 43
Mediation, Modulation, and Consequencesof Membrane-Cytoskeleton InteractionsGary J. Doherty and Harvey T. McMahon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 65
Metal Binding Affinity and Selectivity in Metalloproteins:Insights from Computational StudiesTodor Dudev and Carmay Lim � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 97
Riboswitches: Emerging Themes in RNA Structure and FunctionRebecca K. Montange and Robert T. Batey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �117
Calorimetry and Thermodynamics in Drug DesignJonathan B. Chaires � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �135
Protein Design by Directed EvolutionChristian Jäckel, Peter Kast, and Donald Hilvert � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �153
PIP2 Is A Necessary Cofactor for Ion Channel Function:How and Why?Byung-Chang Suh and Bertil Hille � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �175
RNA Folding: Conformational Statistics, Folding Kinetics,and Ion ElectrostaticsShi-Jie Chen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �197
Intrinsically Disordered Proteins in Human Diseases: Introducingthe D2 ConceptVladimir N. Uversky, Christopher J. Oldfield, and A. Keith Dunker � � � � � � � � � � � � � � � �215
Crowding Effects on Diffusion in Solutions and CellsJames A. Dix and A.S. Verkman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �247
vii
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.

AR343-FM ARI 10 April 2008 7:2
Nanobiotechnology and Cell Biology: Micro- and NanofabricatedSurfaces to Investigate Receptor-Mediated SignalingAlexis J. Torres, Min Wu, David Holowka, and Barbara Baird � � � � � � � � � � � � � � � � � � � � � �265
The Protein Folding ProblemKen A. Dill, S. Banu Ozkan, M. Scott Shell, and Thomas R. Weikl � � � � � � � � � � � � � � � � � �289
Translocation and Unwinding Mechanisms of RNAand DNA HelicasesAnna Marie Pyle � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �317
Structure of Eukaryotic RNA PolymerasesP. Cramer, K.-J. Armache, S. Baumli, S. Benkert, F. Brueckner, C. Buchen,G.E. Damsma, S. Dengl, S.R. Geiger, A.J. Jasiak, A. Jawhari, S. Jennebach,T. Kamenski, H. Kettenberger, C.-D. Kuhn, E. Lehmann, K. Leike, J.F. Sydow,and A. Vannini � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �337
Structure-Based View of Epidermal Growth Factor ReceptorRegulationKathryn M. Ferguson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �353
Macromolecular Crowding and Confinement: Biochemical,Biophysical, and Potential Physiological ConsequencesHuan-Xiang Zhou, Germán Rivas, and Allen P. Minton � � � � � � � � � � � � � � � � � � � � � � � � � � � � �375
Biophysics of Catch BondsWendy E. Thomas, Viola Vogel, and Evgeni Sokurenko � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �399
Single-Molecule Approach to Molecular Biology in Living BacterialCellsX. Sunney Xie, Paul J. Choi, Gene-Wei Li, Nam Ki Lee, and Giuseppe Lia � � � � � � � � �417
Structural Principles from Large RNAsStephen R. Holbrook � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �445
Bimolecular Fluorescence Complementation (BiFC) Analysisas a Probe of Protein Interactions in Living CellsTom K. Kerppola � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �465
Multiple Routes and Structural Heterogeneity in Protein FoldingJayant B. Udgaonkar � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �489
Index
Cumulative Index of Contributing Authors, Volumes 33–37 � � � � � � � � � � � � � � � � � � � � � � � �511
Errata
An online log of corrections to Annual Review of Biophysics articles may be found athttp://biophys.annualreviews.org/errata.shtml
viii Contents
Ann
u. R
ev. B
ioph
ys. 2
008.
37:3
37-3
52. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by M
eteo
rolo
gisc
hes
Inst
itut -
Uni
vers
ity M
uenc
hen
on 0
5/08
/08.
For
per
sona
l use
onl
y.