Embed Size (px)
Citation preview

Transcription
RNA Polymerases and General Transcription Factors

Subjects, covered in this lecture
• A short overview • Structure and mechanism of RNA
polymerases• Eukaryotic promoters• General transcription factors• Elongation factors• Shortly about eukaryotic RNA polymerases,
other than RNA Pol II
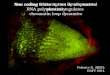
Nucleic acid polymerases: classification
Nucleic acid polymerases
RNA polymerases DNA polymerases
DNA dependent
DNA polymerases
DNA dependent
RNA polymerases
RNA dependent
DNA polymerases
RNA dependent
RNA polymerases
Template specificity
HIV reversetranscriptase
HCV replicase E.coli DNA polymerase III
RNA polymerase II
Sugar specificityAdds dNTPsAdds rNTPs

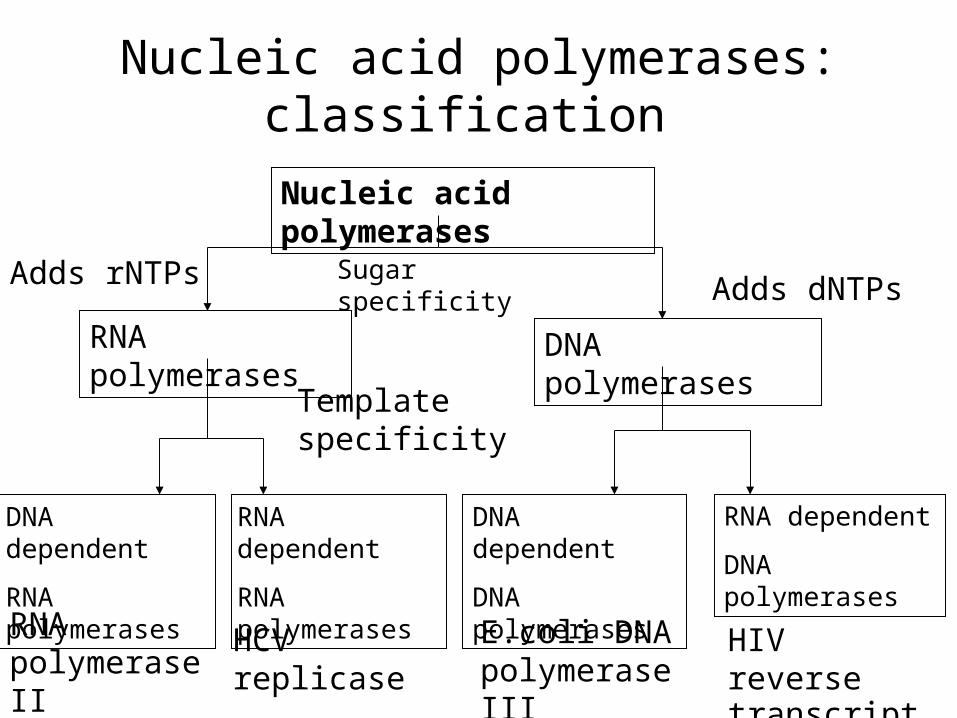
Sugar specificity – how?
• dNTP- yes! ; rNTP – no! that’s easy!
• some residue blocks entry of 2’OH
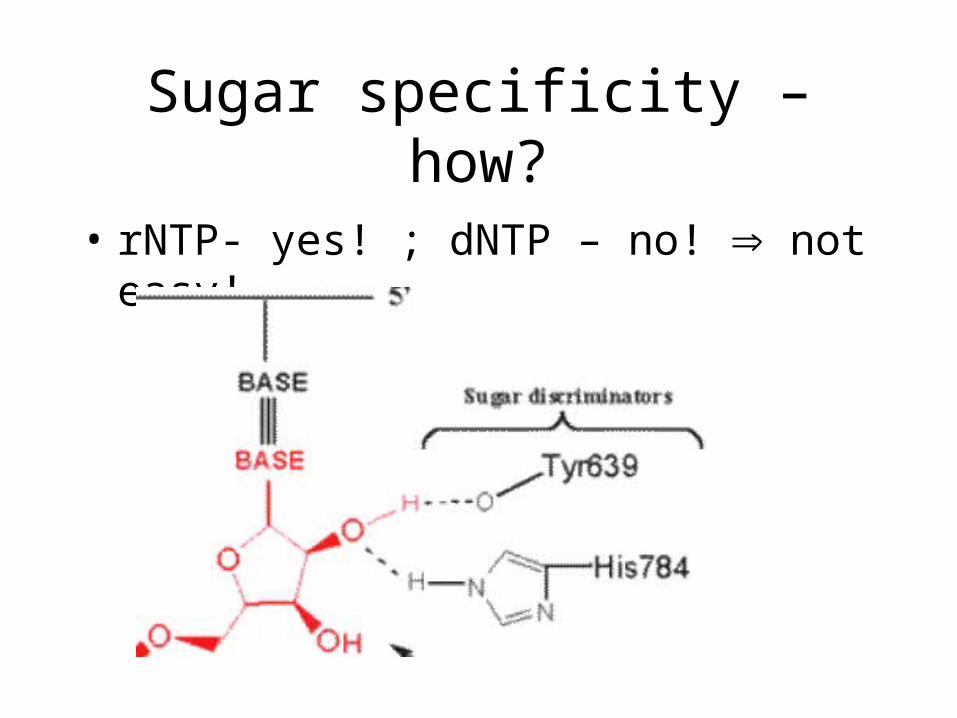
Sugar specificity – how?
• rNTP- yes! ; dNTP – no! not easy!

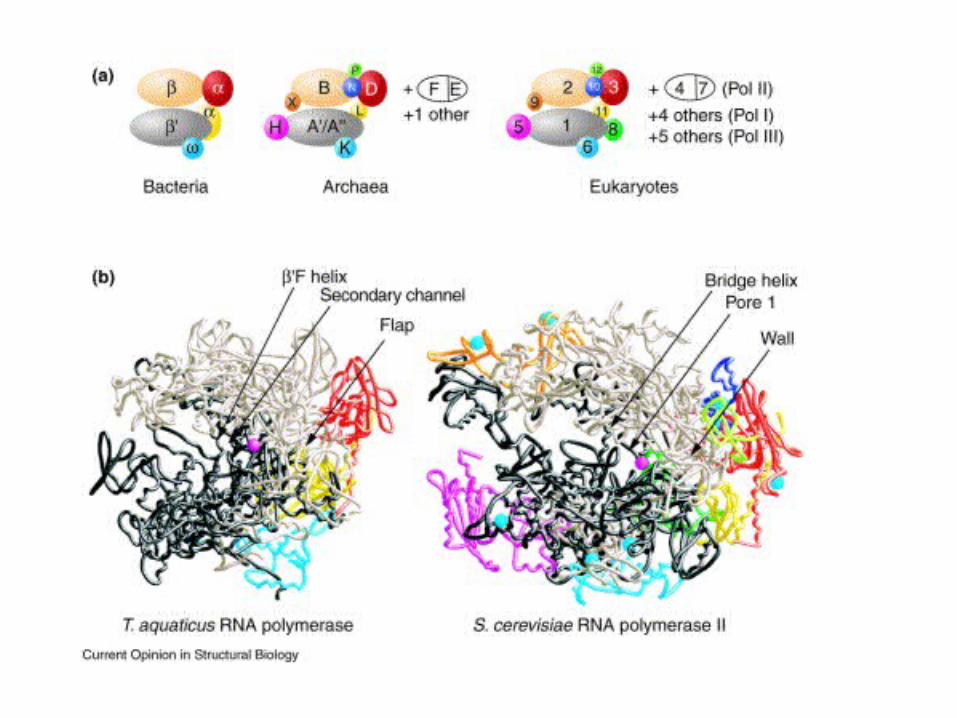
Simple and complex DNA – dependent RNA polymerases
• Viral RNA polymerases (1 subunit)
• Prokaryotic RNA polymerases (4 different subunits)
• Eukaryotic and archeal RNA polymerases (at least 12 subunits)
Viruses

The structural features of RNA polymerases
Based on 3D sructures of bacteriophage T7 RNA pol and yeast
RNA Pol II
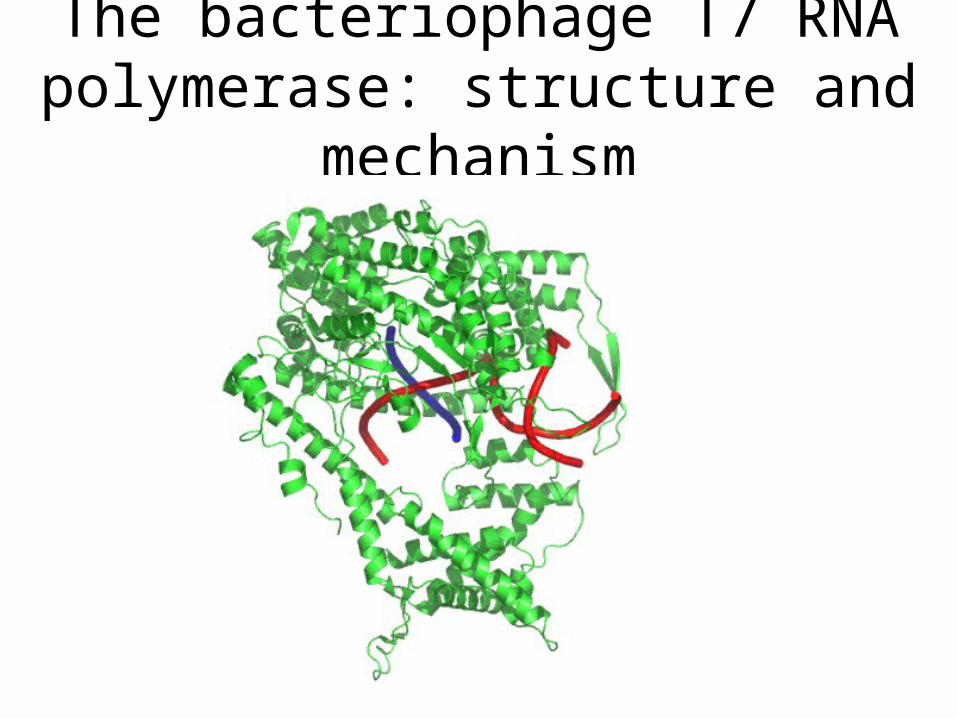
The bacteriophage T7 RNA polymerase: structure and mechanism
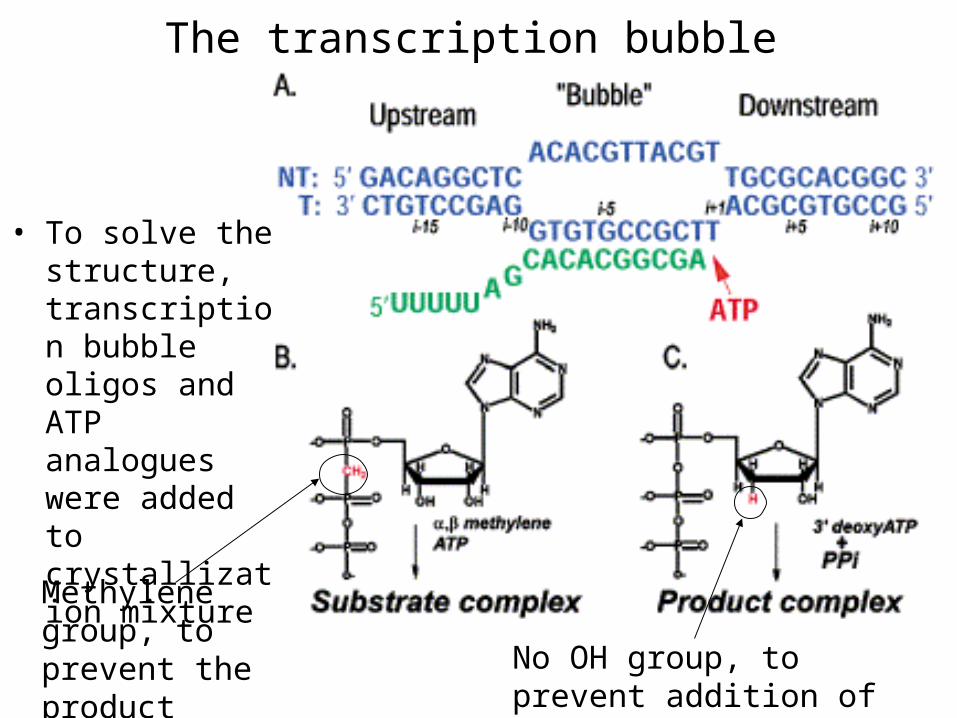
The transcription bubble
• To solve the structure, transcription bubble oligos and ATP analogues were added to crystallization mixture
Methylene group, to prevent the product formation No OH group, to prevent addition
of the next nucleotide
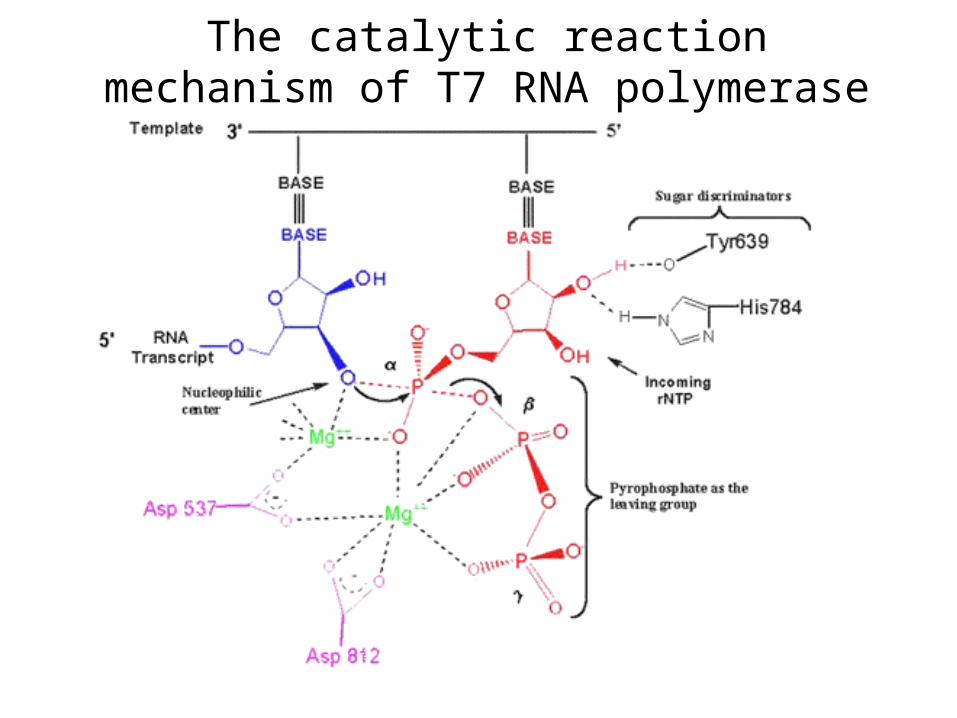
The catalytic reaction mechanism of T7 RNA polymerase
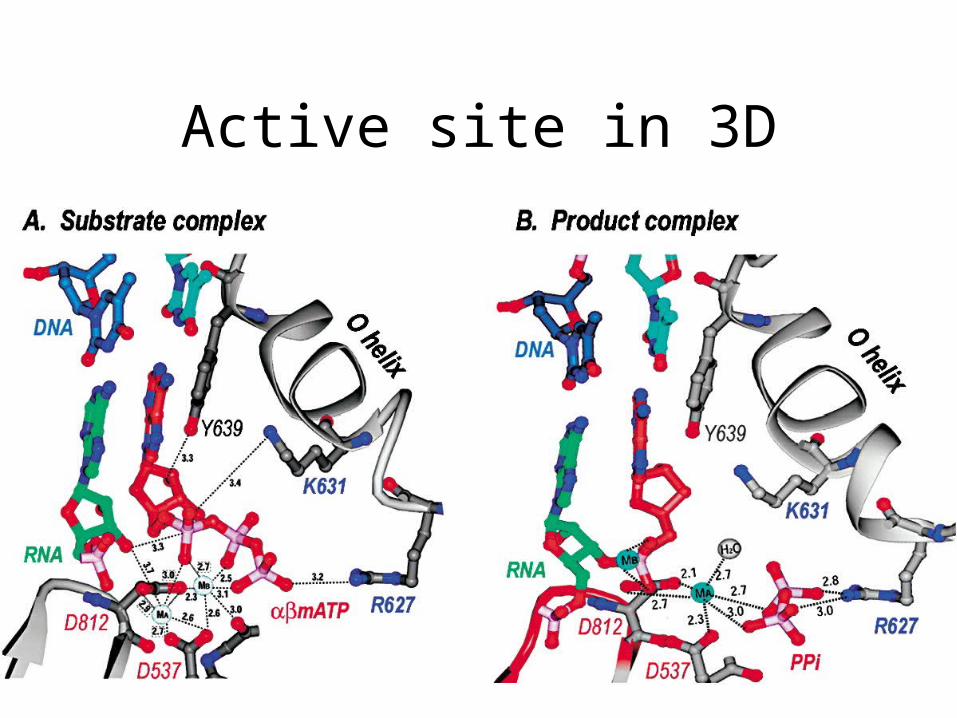
Active site in 3D
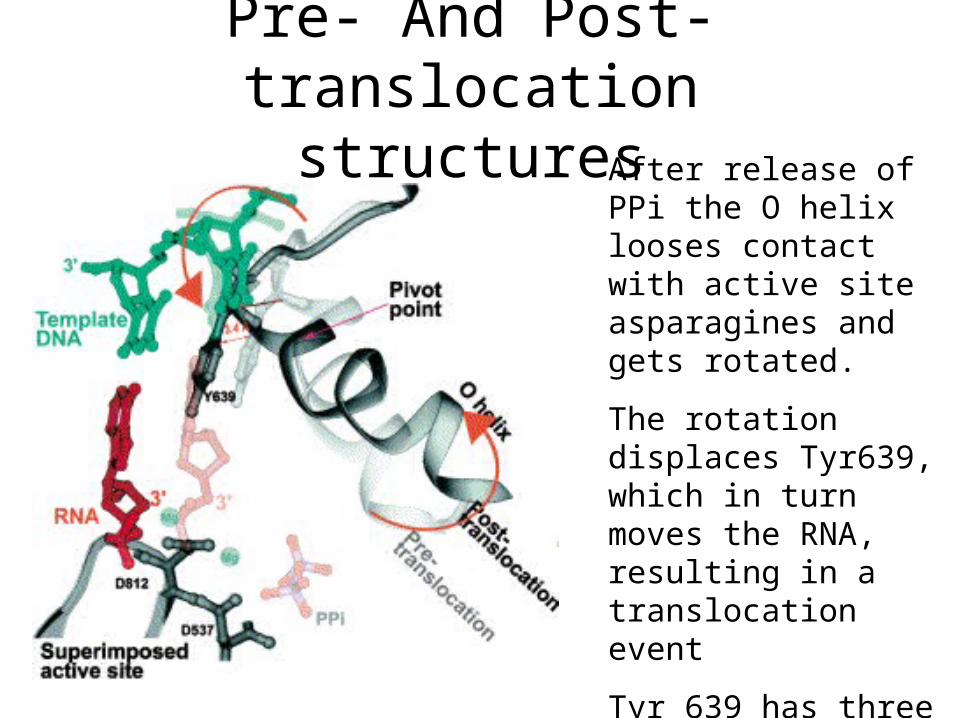
Pre- And Post-translocation structures
After release of PPi the O helix looses contact with active site asparagines and gets rotated.
The rotation displaces Tyr639, which in turn moves the RNA, resulting in a translocation event
Tyr 639 has three functions –(1) sugar discrimination, (2) stacking against incoming base and (3) translocation
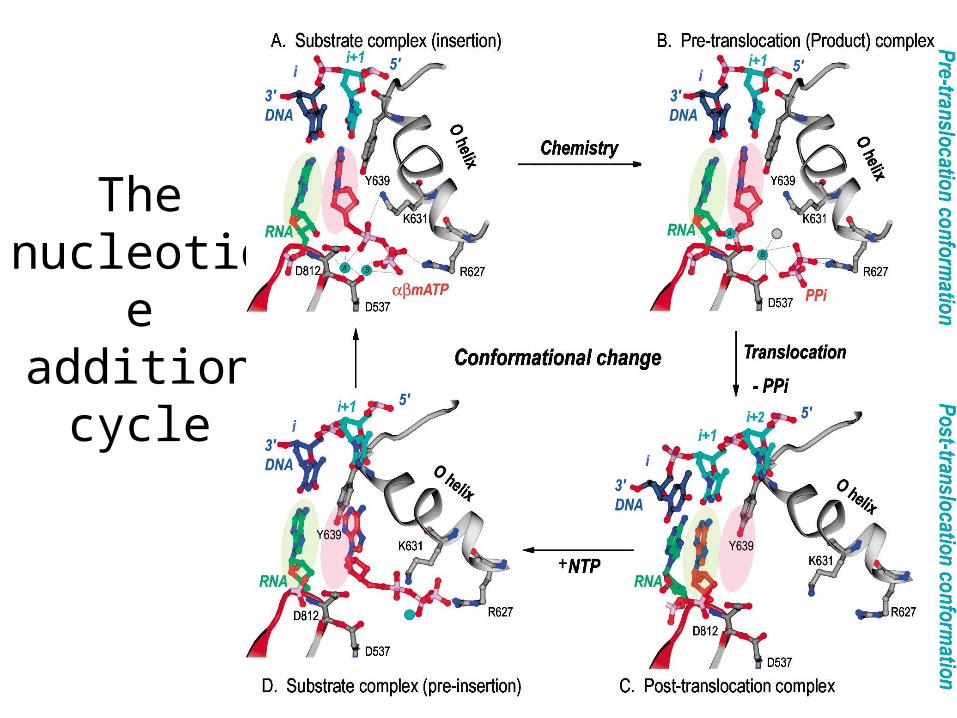
The nucleotide addition
cycle

Differences between non-transcribing and transcribing structures of RNA Pol II
• The non-transcribing complex is more “open”, which faciliates DNA binding
• Upon the binding to promoter, the structure, called “clamp” closes the complex like a lid
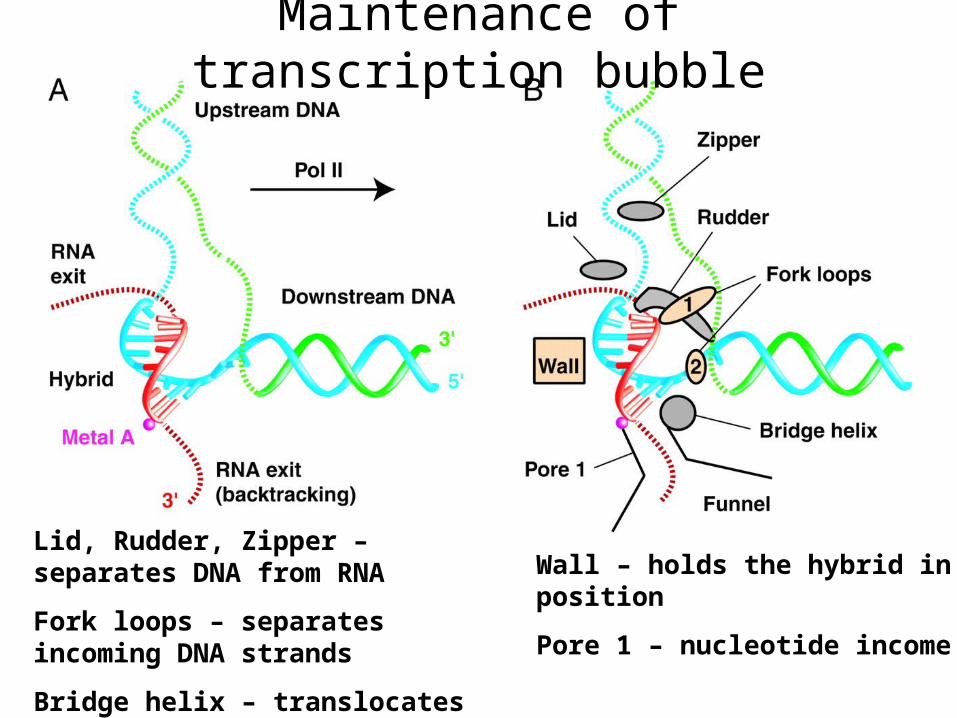
Maintenance of transcription bubble
Lid, Rudder, Zipper – separates DNA from RNA
Fork loops – separates incoming DNA strands
Bridge helix – translocates the bubble
Wall – holds the hybrid in position
Pore 1 – nucleotide income
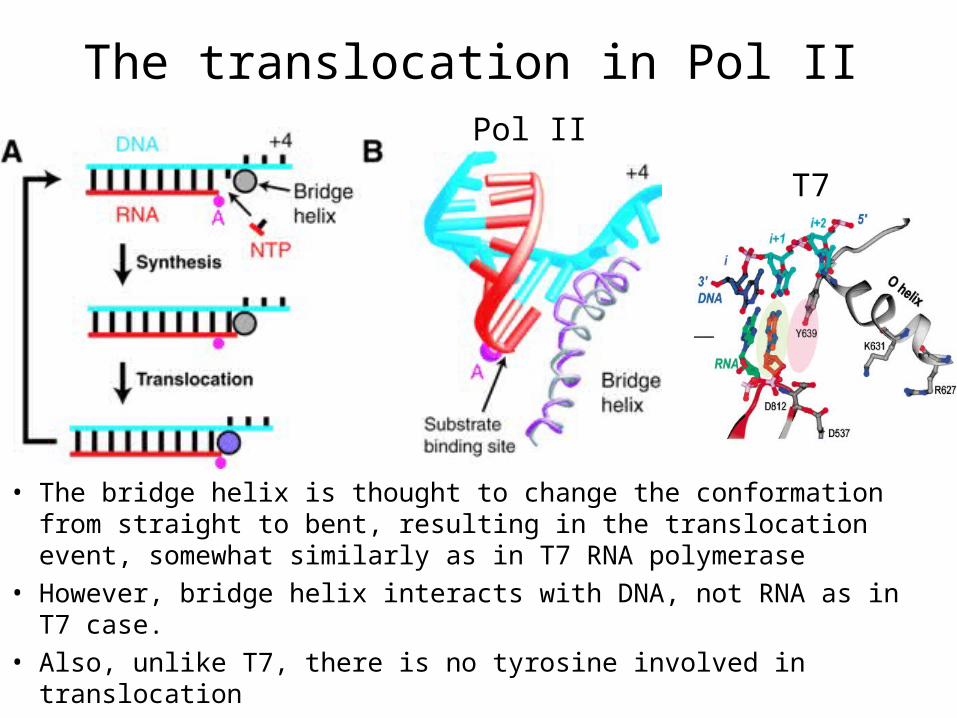
The translocation in Pol II
• The bridge helix is thought to change the conformation from straight to bent, resulting in the translocation event, somewhat similarly as in T7 RNA polymerase
• However, bridge helix interacts with DNA, not RNA as in T7 case.
• Also, unlike T7, there is no tyrosine involved in translocation
T7
Pol II

Are multi-subunit (RNA pol II) and single subunit (T7) RNA polymerases evolutionary related?
• Probably not• One similar feature is involvement of two magnesium
ions in catalytic activity• Another similar feature is involvment of Bridge helix
(Pol II) and O helix (T7) in translocation, although there is no involvement of tyrosine in Pol II
• However, there are no sequence or structure similarities among T7 and Pol II
• Differences exist also in ribose recognition – no obvious sugar discriminators in PolII
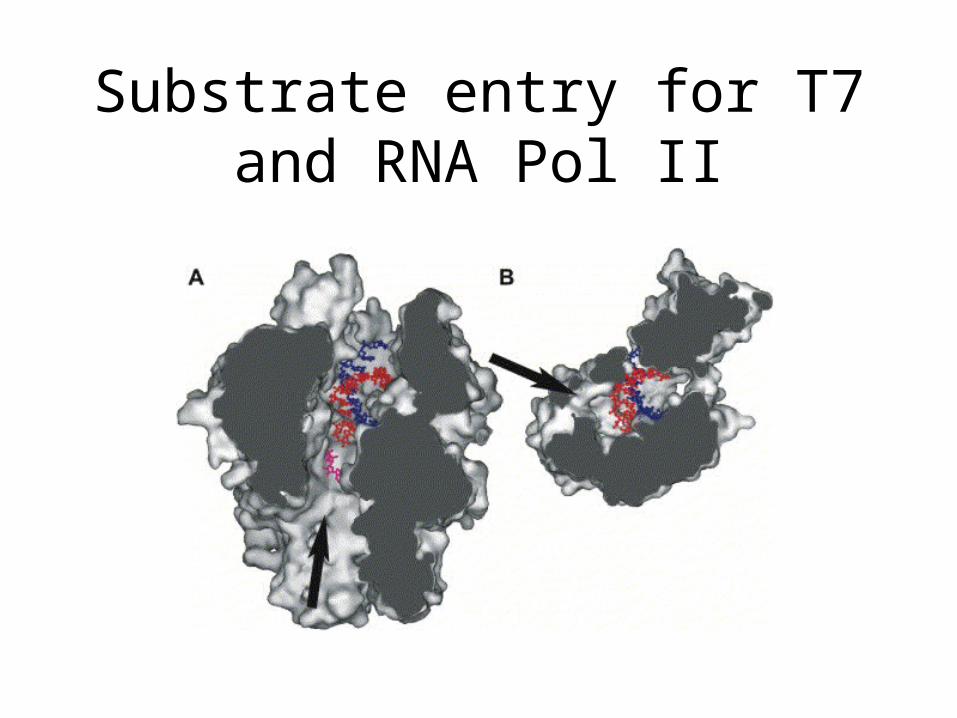
Substrate entry for T7 and RNA Pol II

Carboxyl terminal domain (CTD) of Pol II
• A stretch of 7aa repeats in c-terminus of Pol II subunit 1.• Consensus sequence of repeats YSPTSPS• In yeast 26 repeats, in humans - 50 • CTDs get phosphorylated at several Ser and Tyr residues
after transcription initiation• Phosphorylation of CTDs is important for promoter
escape, elongation and post-transcritption events• CTDs have shown to act as a “landing pad” for different
proteins, involved in elongation and post-transcription events
• The base of CTDs is located near the exit groove of RNA

The eukaryotic promoters
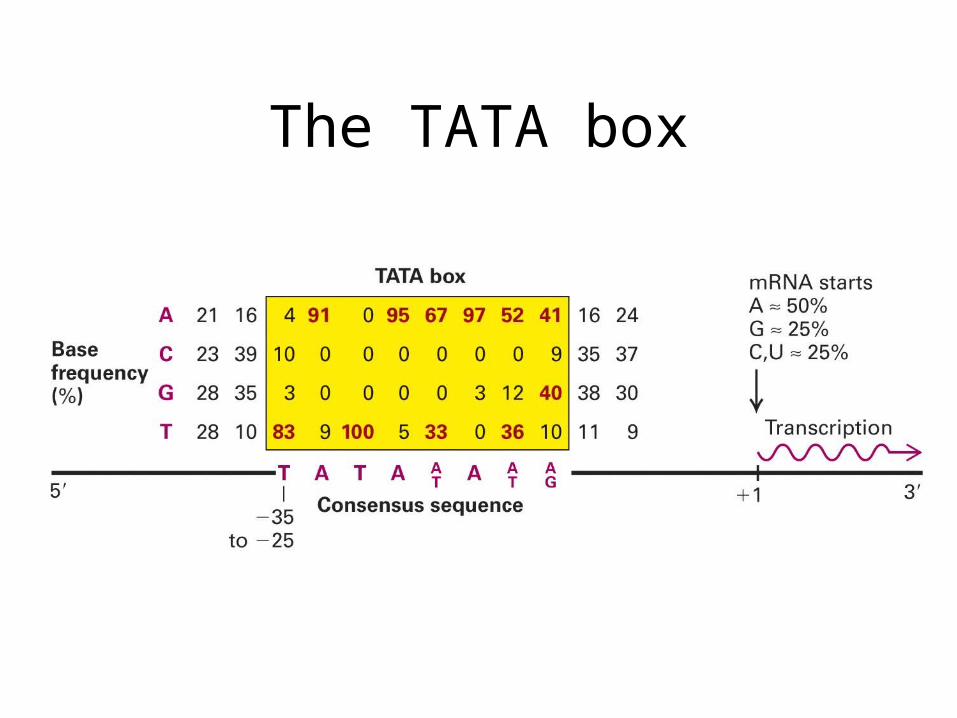
The TATA box

Other promoter sequences
• Initiator (Inr): Y-Y-A+1-N-T/A-Y-Y-Y. Some promoters contain both initiator and TATA box, whereas others only one of them
• CpG islands: CG rich stretch of 20-50 nucleotides within ~100 base pairs upstream the start site.
• BRE (TFIIB recognition element) sequence, which has the consensus G/C-G/C-G/A-C-G-C-C, is located immediately upstream of the TATA element of some promoters and
increases the affinity of TFIIB for the promoter • Downstream promoter element (DPE) is present in some
genes with initiator promoter. It is located about 30 nucleotides downstream of the transcription start site and includes a common G-A/T-C-G sequence
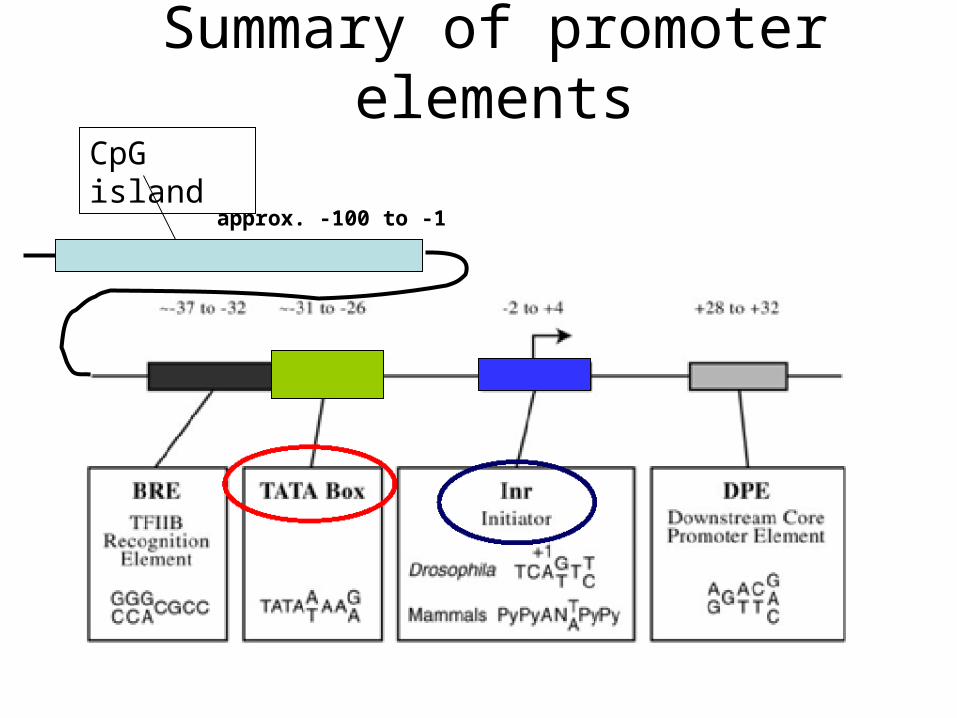
Summary of promoter elements
CpG island
approx. -100 to -1

General transcription factors• General transcription factors are required for
transcription in eukaryotes from all genes • GTFs assist RNA Pol II in transcription initiation • GTFs are designated TFIIA, TFIIB,... and most of
them are multimeric proteins• Equivalent GTFs are highly conserved among the
eukaryotes• In prokaryotes, only one general transcription
factor, known as factor is required
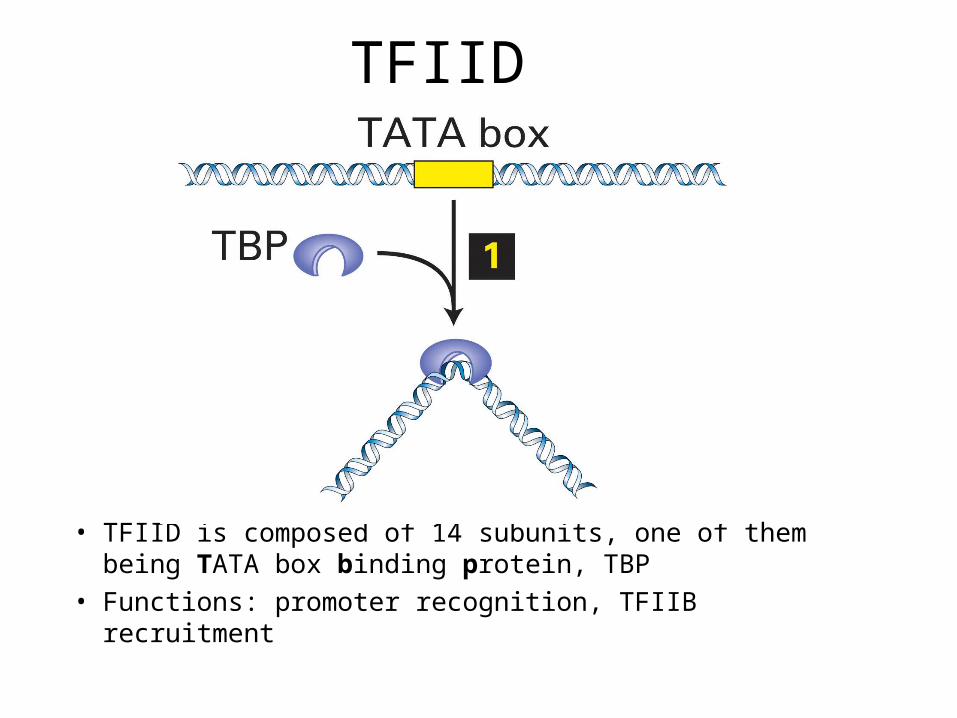
TFIID
• TFIID is composed of 14 subunits, one of them being TATA box binding protein, TBP
• Functions: promoter recognition, TFIIB recruitment
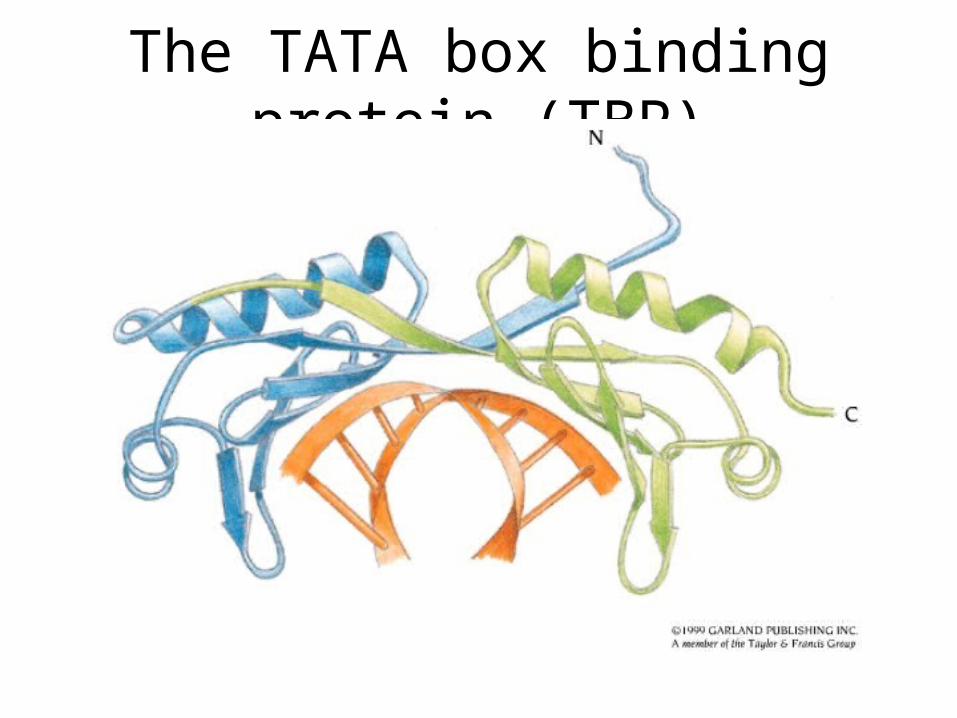
The TATA box binding protein (TBP)
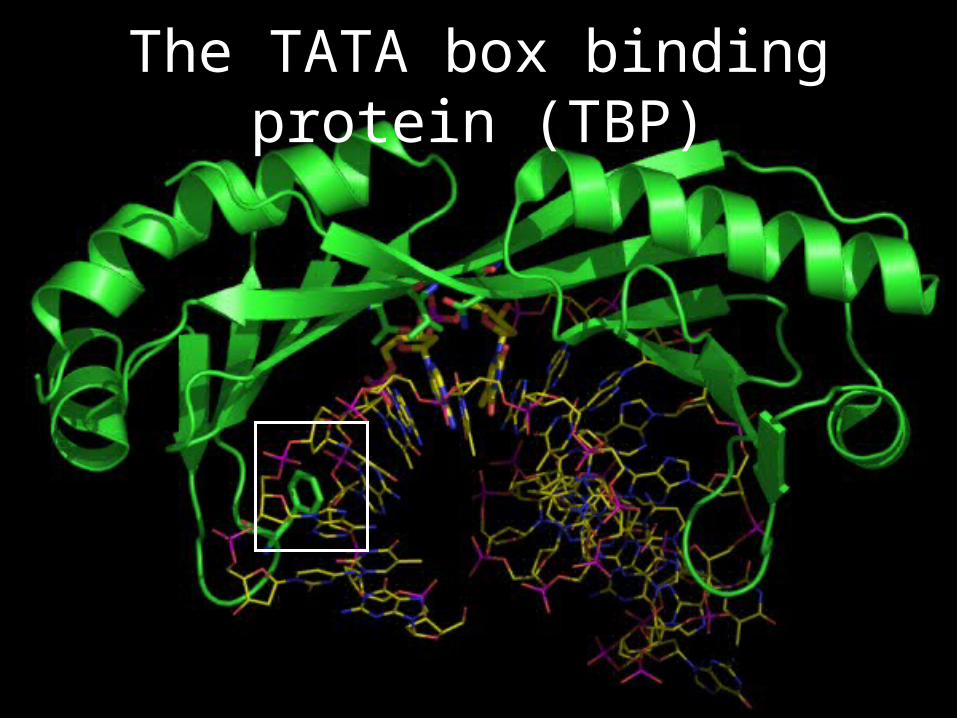
The TATA box binding protein (TBP)
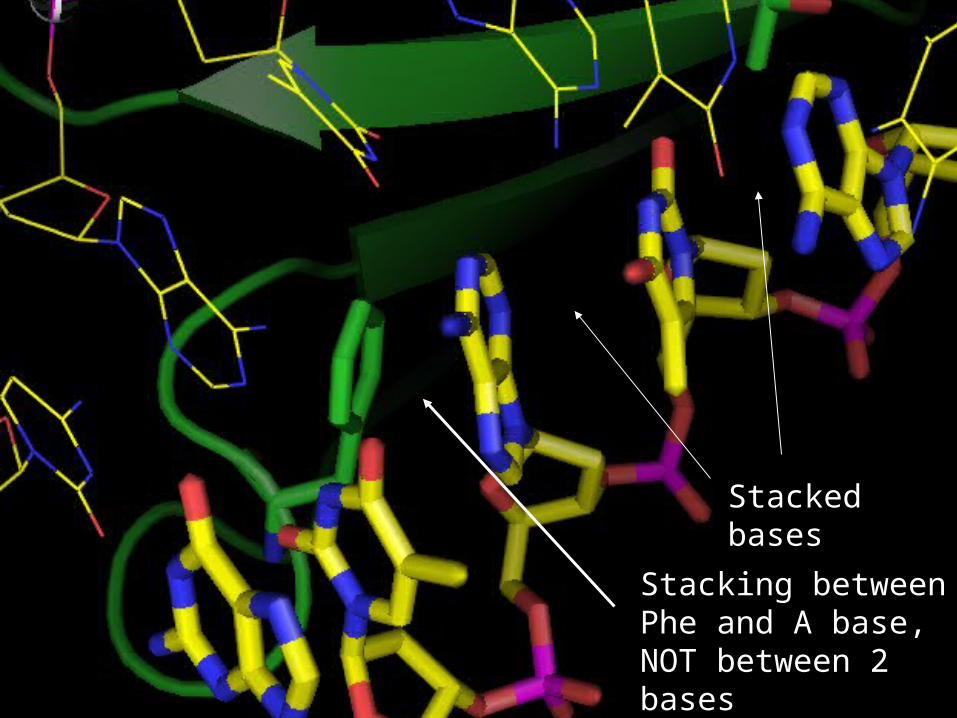
Stacked bases
Stacking between Phe and A base, NOT between 2 bases
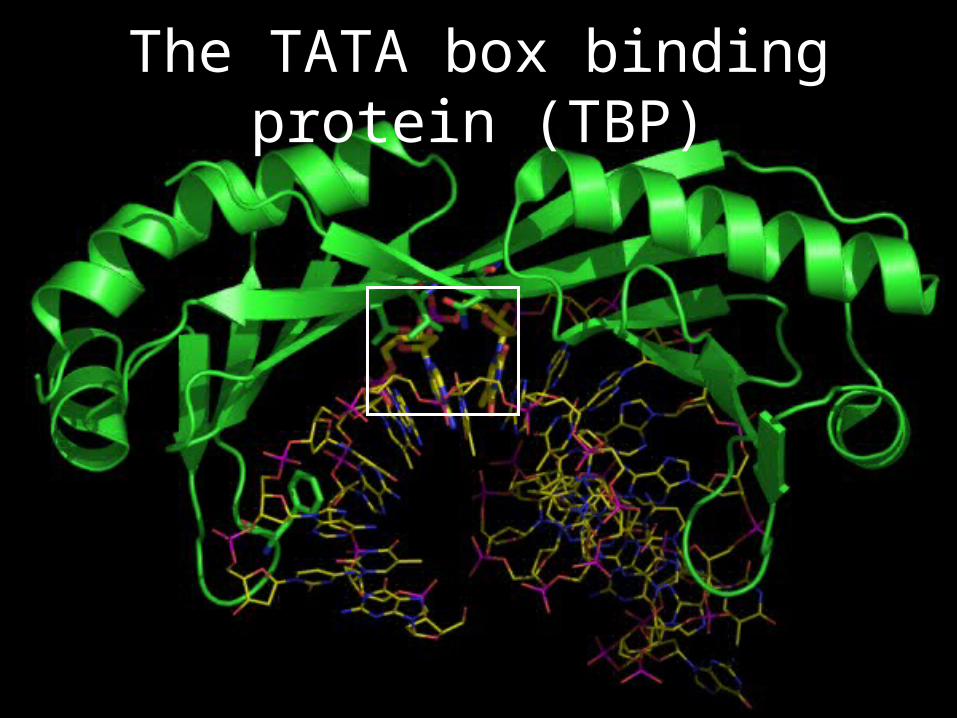
The TATA box binding protein (TBP)
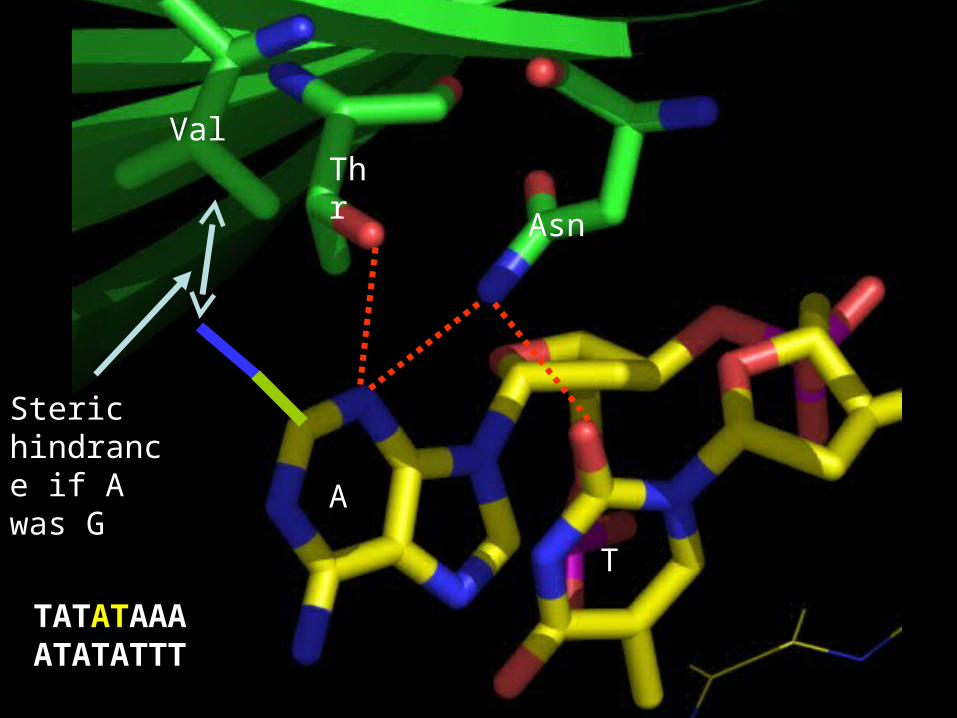
Steric hindrance if A was G
ValThr
Asn
A
T
TATATAAAATATATTT
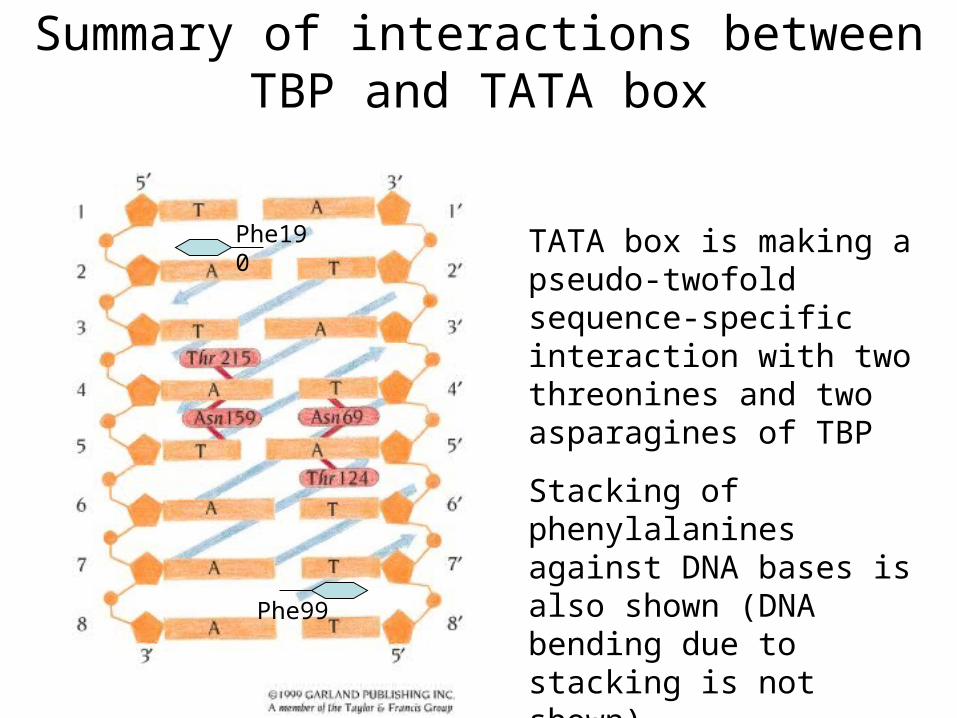
Summary of interactions between TBP and TATA box
TATA box is making a pseudo-twofold sequence-specific interaction with two threonines and two asparagines of TBP
Stacking of phenylalanines against DNA bases is also shown (DNA bending due to stacking is not shown)
Phe190
Phe99
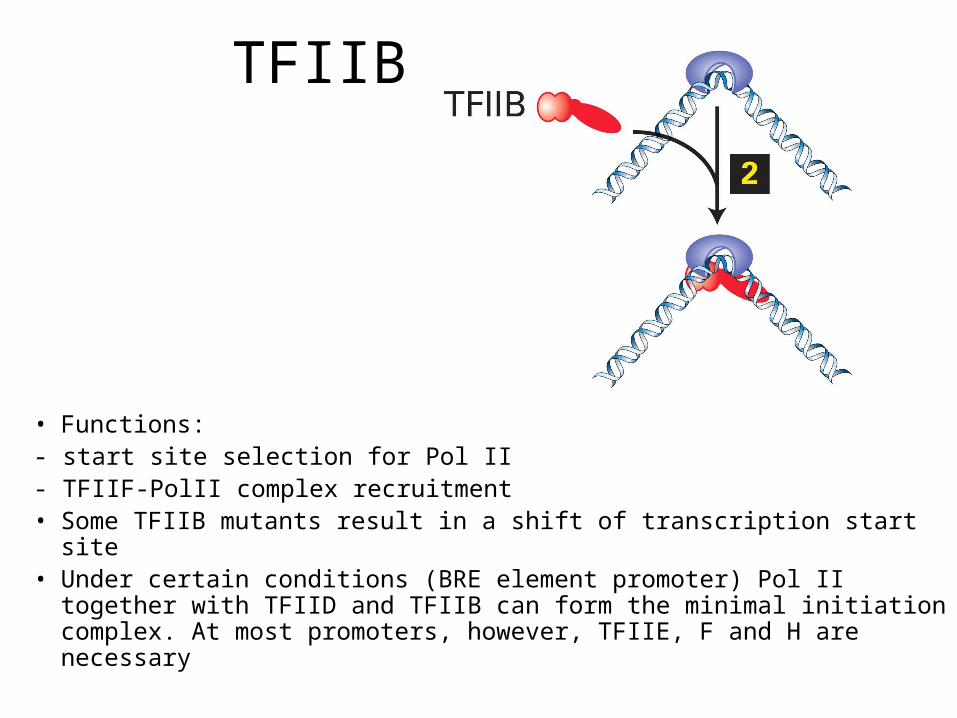
TFIIB
• Functions: - start site selection for Pol II - TFIIF-PolII complex recruitment• Some TFIIB mutants result in a shift of transcription start site• Under certain conditions (BRE element promoter) Pol II together
with TFIID and TFIIB can form the minimal initiation complex. At most promoters, however, TFIIE, F and H are necessary
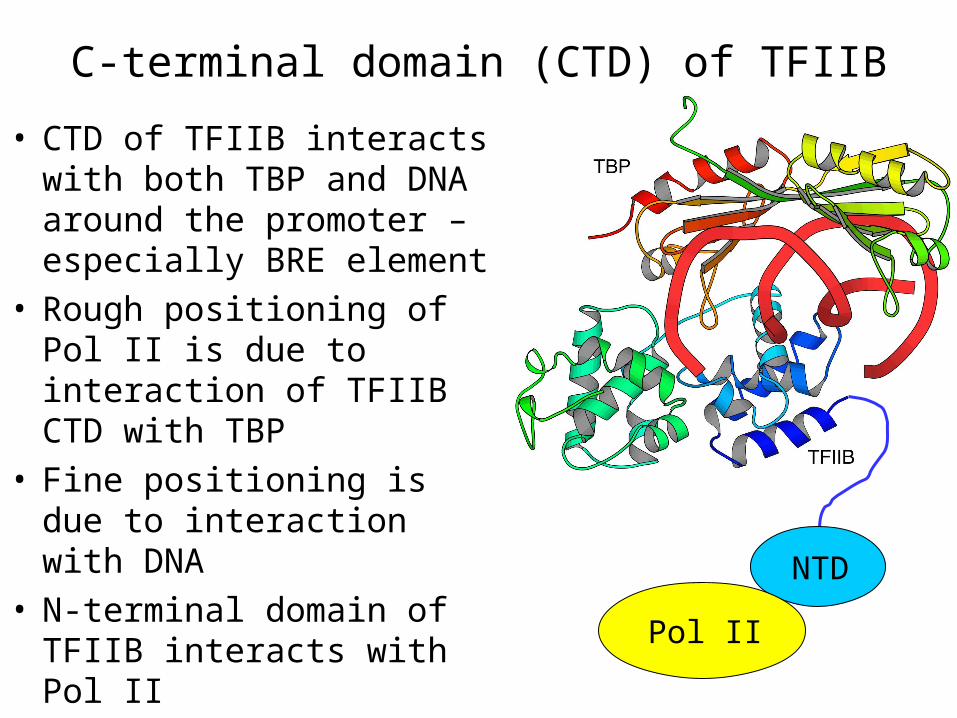
C-terminal domain (CTD) of TFIIB
• CTD of TFIIB interacts with both TBP and DNA around the promoter – especially BRE element
• Rough positioning of Pol II is due to interaction of TFIIB CTD with TBP
• Fine positioning is due to interaction with DNA
• N-terminal domain of TFIIB interacts with Pol II Pol II
NTD
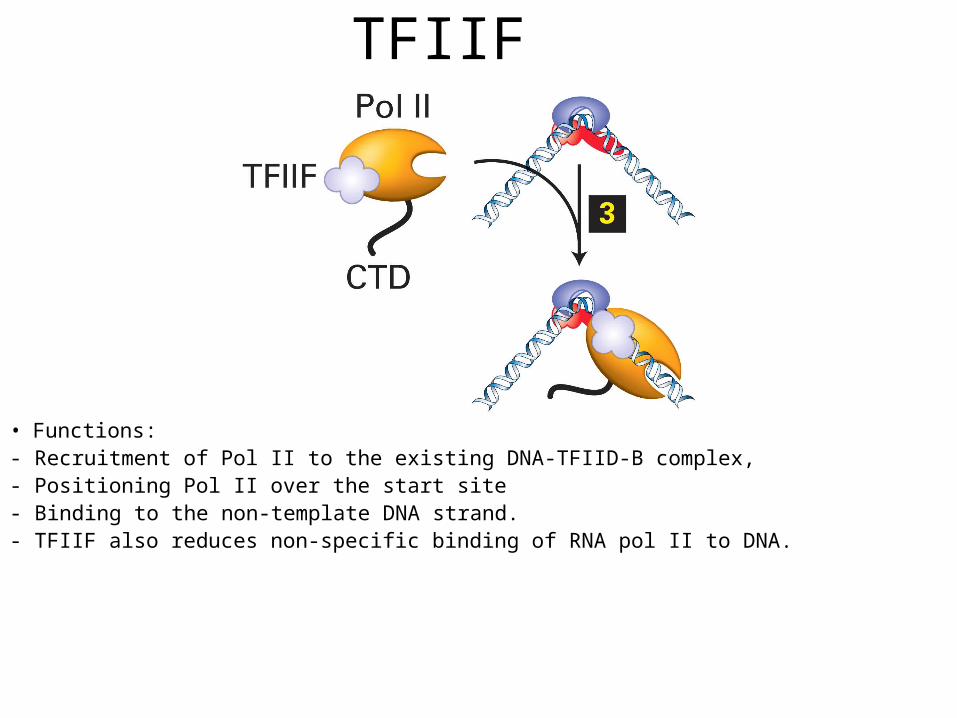
TFIIF
• Functions: - Recruitment of Pol II to the existing DNA-TFIID-B complex, - Positioning Pol II over the start site- Binding to the non-template DNA strand. - TFIIF also reduces non-specific binding of RNA pol II to DNA.
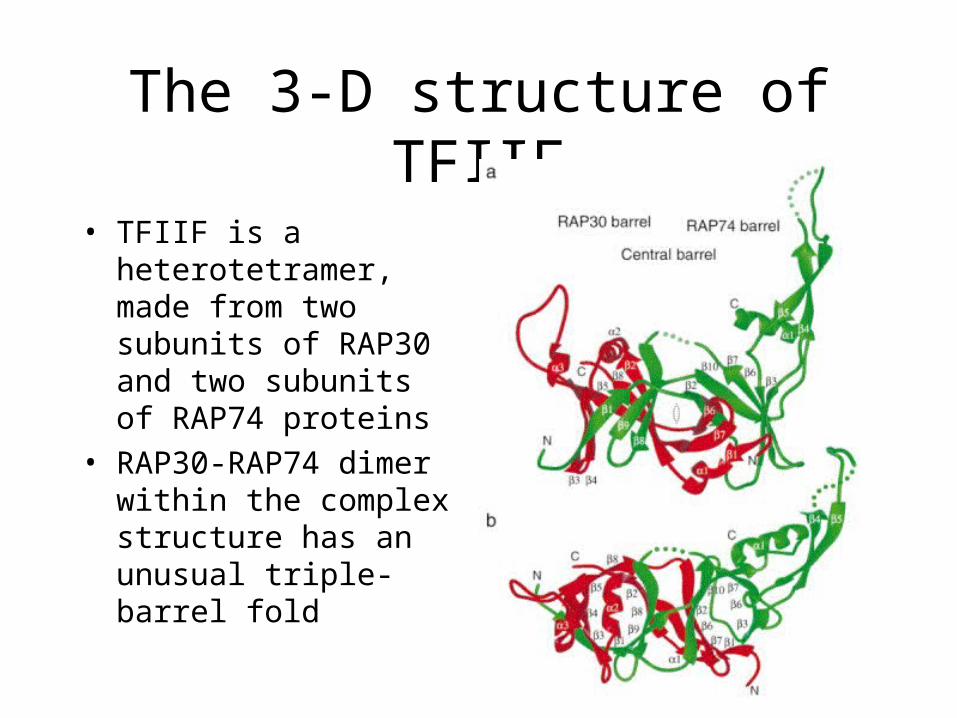
The 3-D structure of TFIIF
• TFIIF is a heterotetramer, made from two subunits of RAP30 and two subunits of RAP74 proteins
• RAP30-RAP74 dimer within the complex structure has an unusual triple-barrel fold
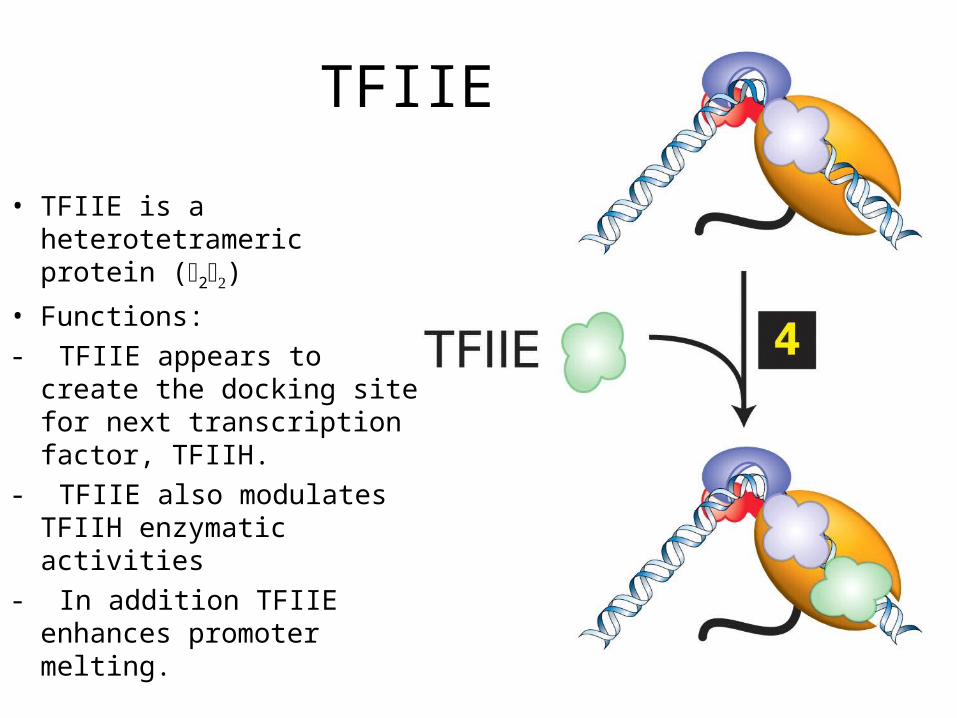
TFIIE
• TFIIE is a heterotetrameric protein (2)
• Functions:
- TFIIE appears to create the docking site for next transcription factor, TFIIH.
- TFIIE also modulates TFIIH enzymatic activities
- In addition TFIIE enhances promoter melting.

TFIIH• TFIIH is a multimeric protein,
composed of 9 subunits, some of them with distinct enzymatic activities
• Functions:
- TFIIH has a helicase activity, which unwinds the DNA duplex at a start site, allowing Pol II to bind to the template strand.
- TFIIH also has a kinase ativity, it phosphorylates PolII in the begining of elongation
- Other TFIIH subunits have been shown to recruit DNA-repairing enzymes if polymerase reaches damaged region in DNA and gets stalled

• As Pol II transcribes away from the start site subunit of TFIIH phosphorylates the Pol II CTD, which results in promoter escape.
• General factors get released
Early events in elongation

TFIIA
• For transcription in vivo, another factor TFIIA is required
• The function of TFIIA is somewhat unclear, but it might help the other factors to bind.
• TFIIA has also shown to have some anti-repressor functions
• TFIIA is not required for transcription in vitro.

TAFs
• Apart from TBP, TFIID has 13 TAF ( TBP associated factors) subunits
• Some of them seem to be necessary for transcription initiation from promoters, lacking the TATA box
• Other TAFs have been shown to be tissue-specific coactivators
• TAF subunits also interact with other GTFs therefore stabilyzing the complex.
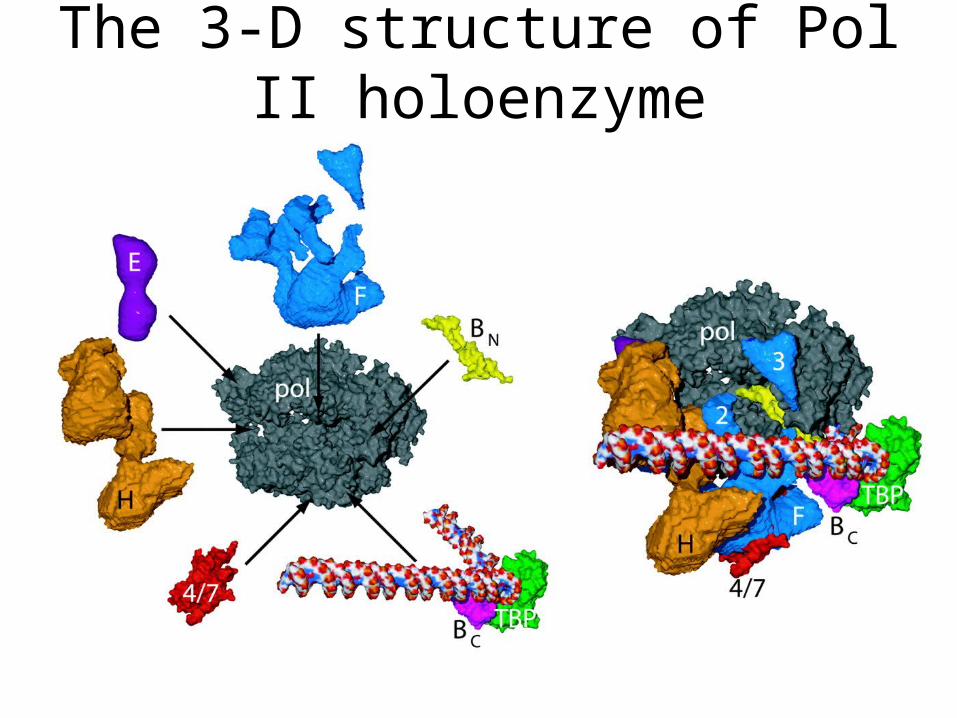
The 3-D structure of Pol II holoenzyme
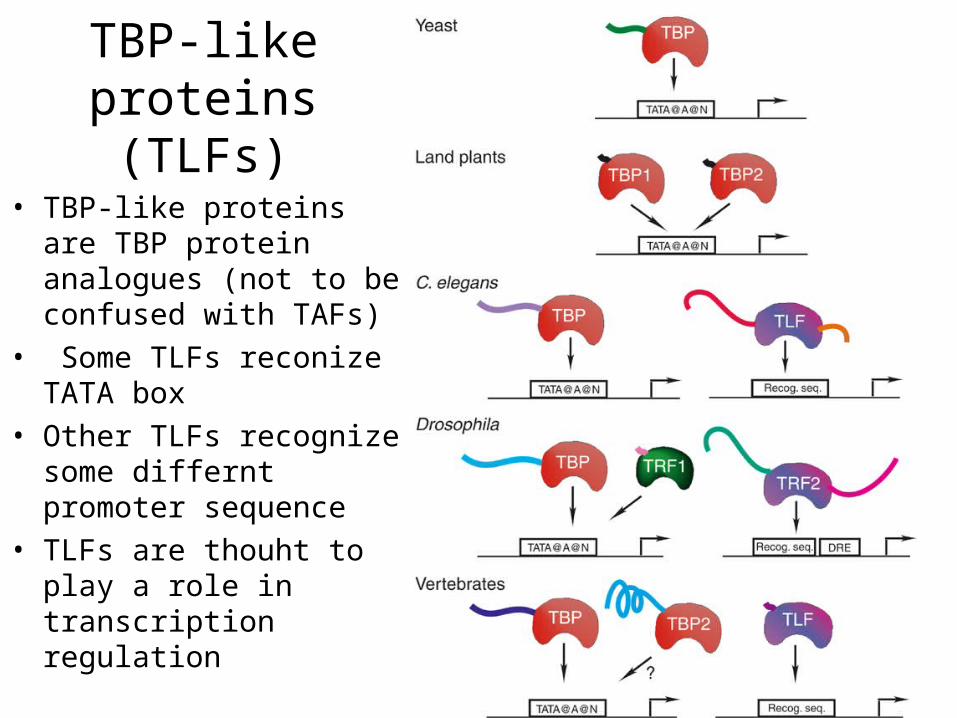
TBP-like proteins (TLFs)
• TBP-like proteins are TBP protein analogues (not to be confused with TAFs)
• Some TLFs reconize TATA box
• Other TLFs recognize some differnt promoter sequence
• TLFs are thouht to play a role in transcription regulation
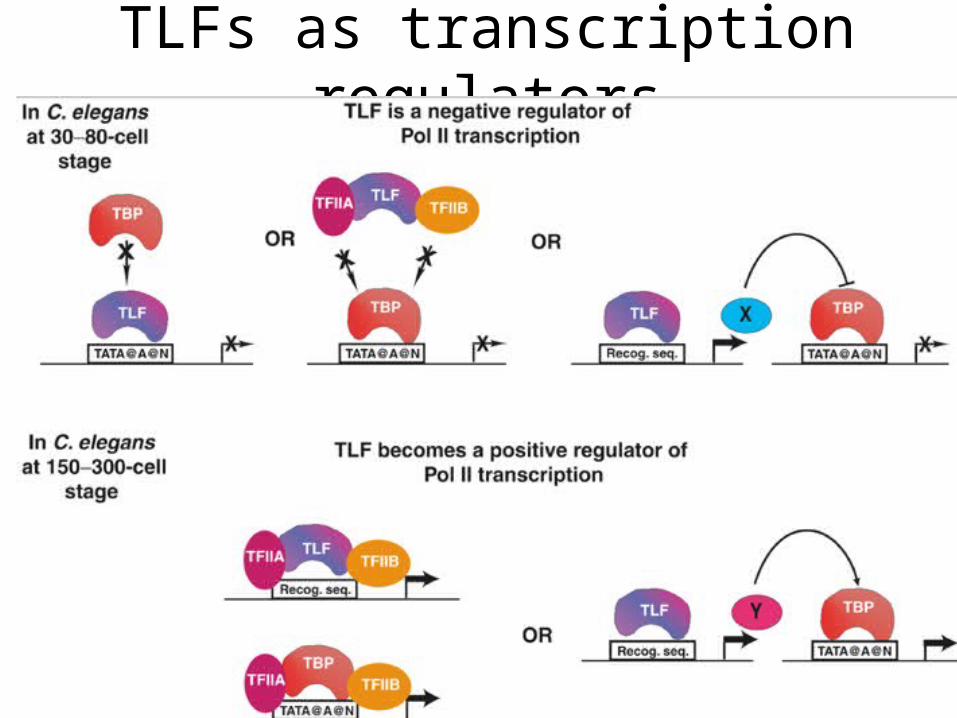
TLFs as transcription regulators

What is Pol II backsliding, pausing and arrest?
• Backsliding is an event when RNA polymerase moves backwards and newly made RNA gets inserted in the funnel. It can be caused by incorporation of wrong NTP or when 3’OH looses contact with active site Mg2+
• If the backsliding RNA piece is 2-4 nt long, this a reversible process and RNA polymerase can recover by itself. This is called pausing.
• If the backsliding RNA gets longer (7-10 nt), it gets trapped in funnel pore and RNA polymerase can not recover by itself. This is caused arrest.
• Arrest can be overcome by specific elongation factors, which help RNA Pol II to cleave the arrested RNA
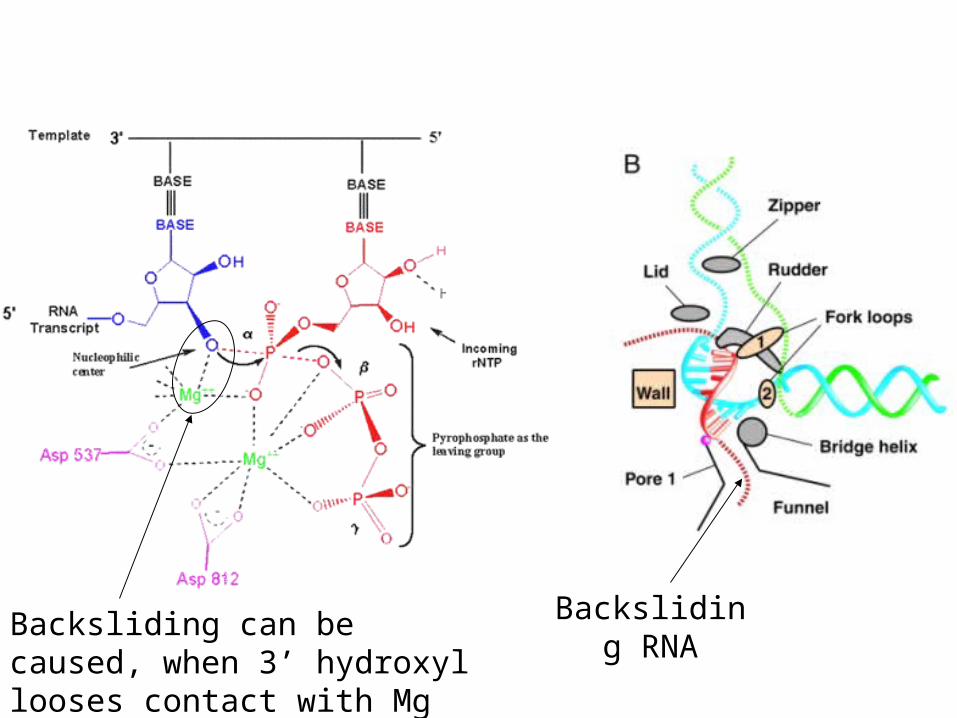
Backsliding RNA
Backsliding can be caused, when 3’ hydroxyl looses contact with Mg
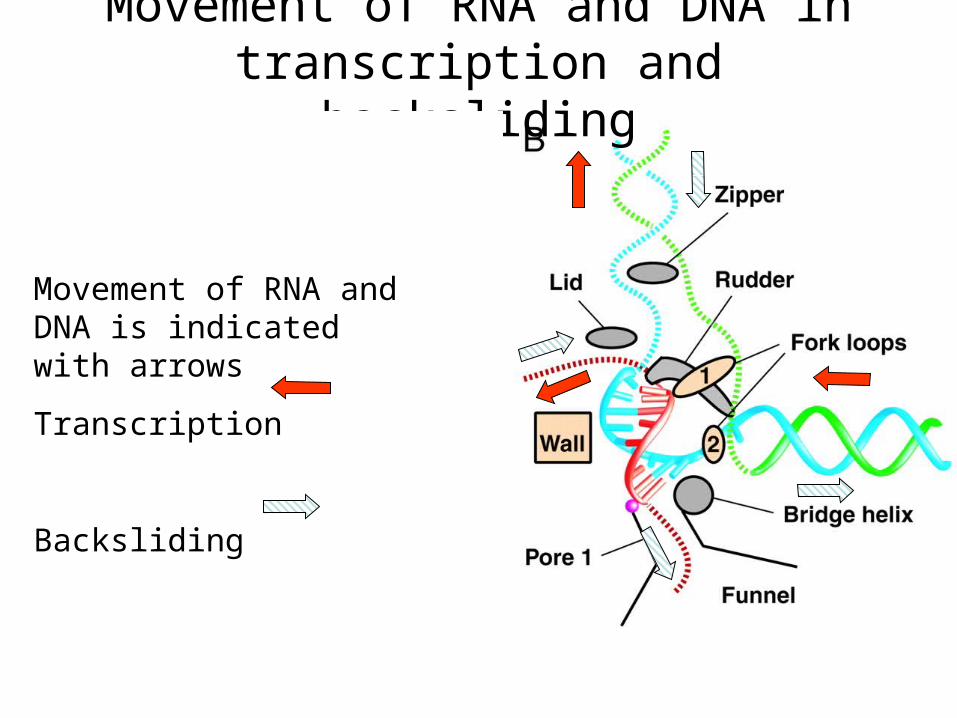
Movement of RNA and DNA in transcription and backsliding
Movement of RNA and DNA is indicated with arrows
Transcription
Backsliding

Transcription elongation factors
In vivo rates of transcription = 1200-2000 nucleotides/min.
Baseline in vitro rates = 100-300 nucleotides/min.
In vitro frequent transcription pausing and arrests occur even with chromatin-free DNA
Large number of proteins and protein complexes exist that regulate elongation of transcription (elongation factors)
Elongation factors display following activities:
• Enable elongation through chromatin• Suppress pausing• Overcome transcription arrest

Transcription elongation factor IIS
• Elongated shape –why?
• TFIIS gets inserted in active site of PolII
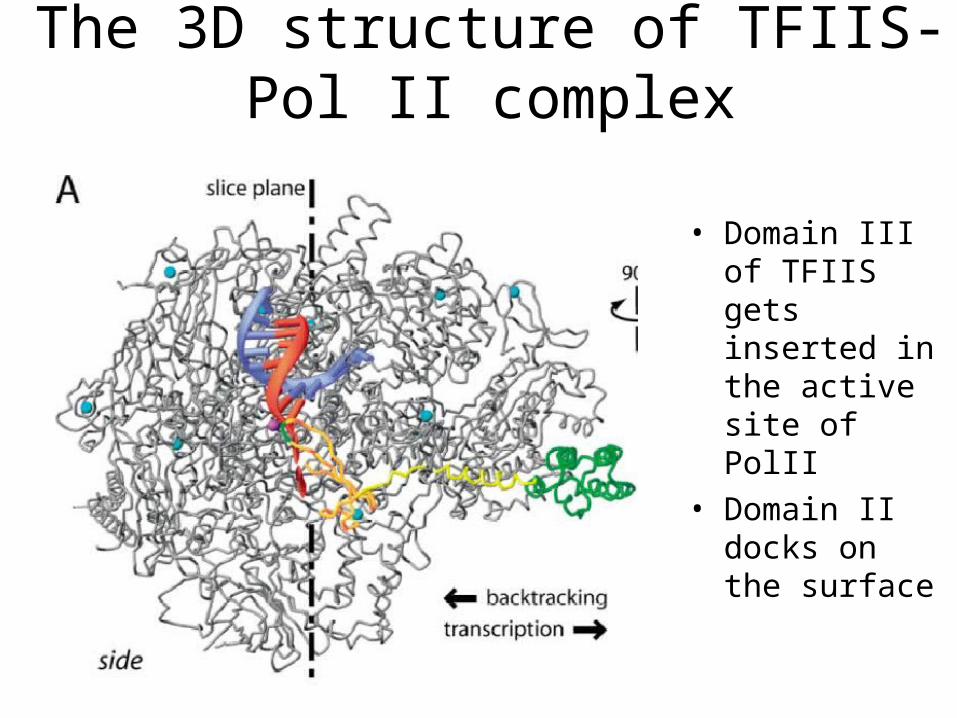
The 3D structure of TFIIS-Pol II complex
• Domain III of TFIIS gets inserted in the active site of PolII
• Domain II docks on the surface
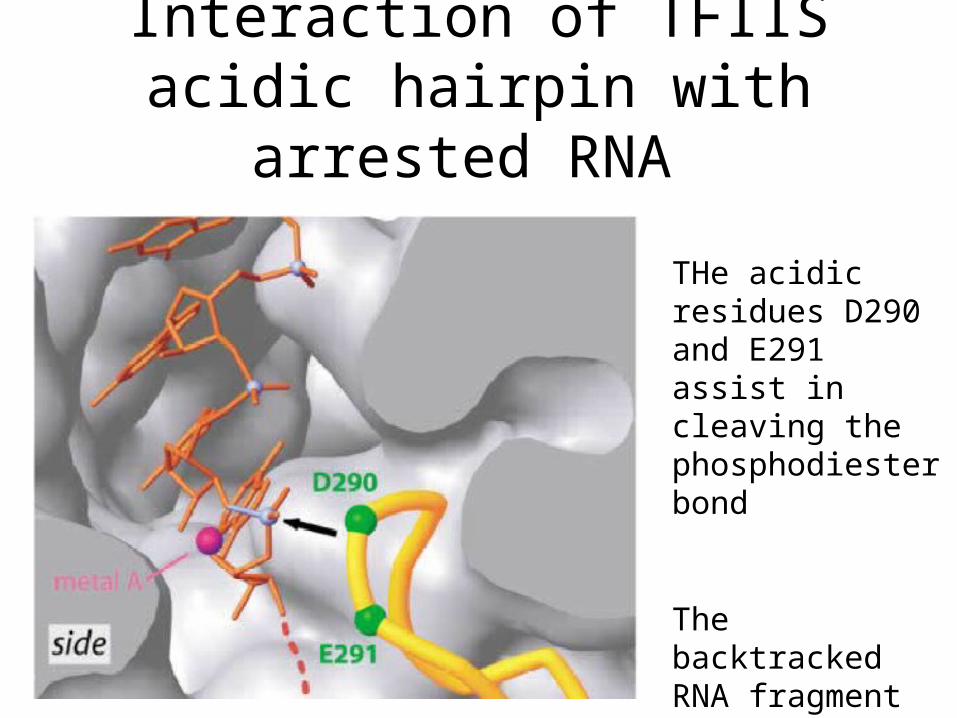
Interaction of TFIIS acidic hairpin with arrested RNA
THe acidic residues D290 and E291 assist in cleaving the phosphodiester bond
The backtracked RNA fragment gets released and RNA Pol II is rescued from arrest
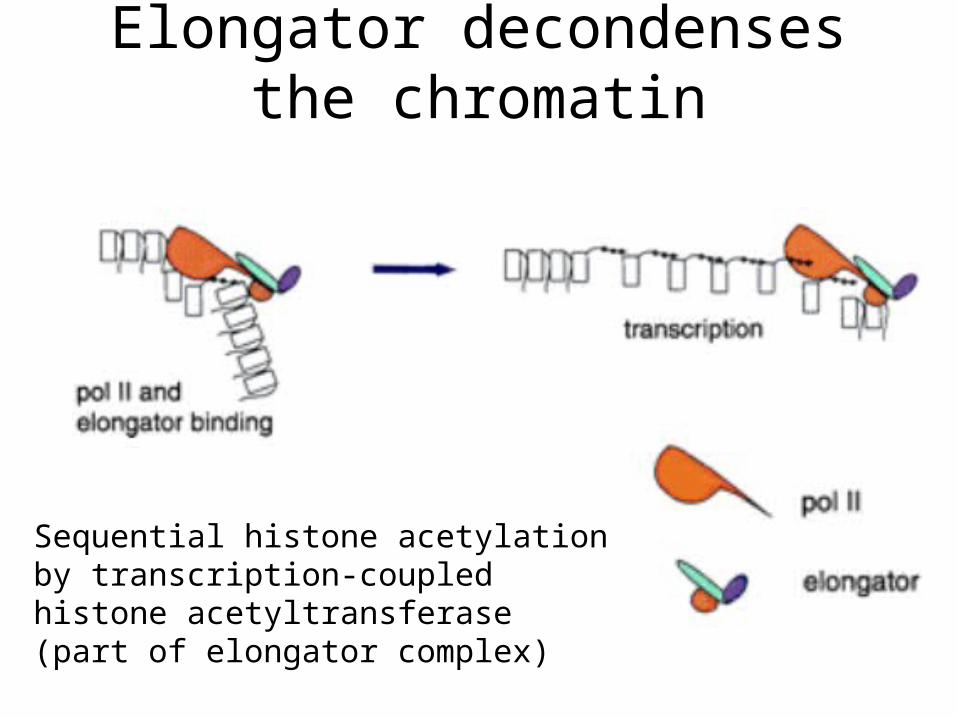
Elongator decondenses the chromatin
Sequential histone acetylation by transcription-coupled histone acetyltransferase (part of elongator complex)
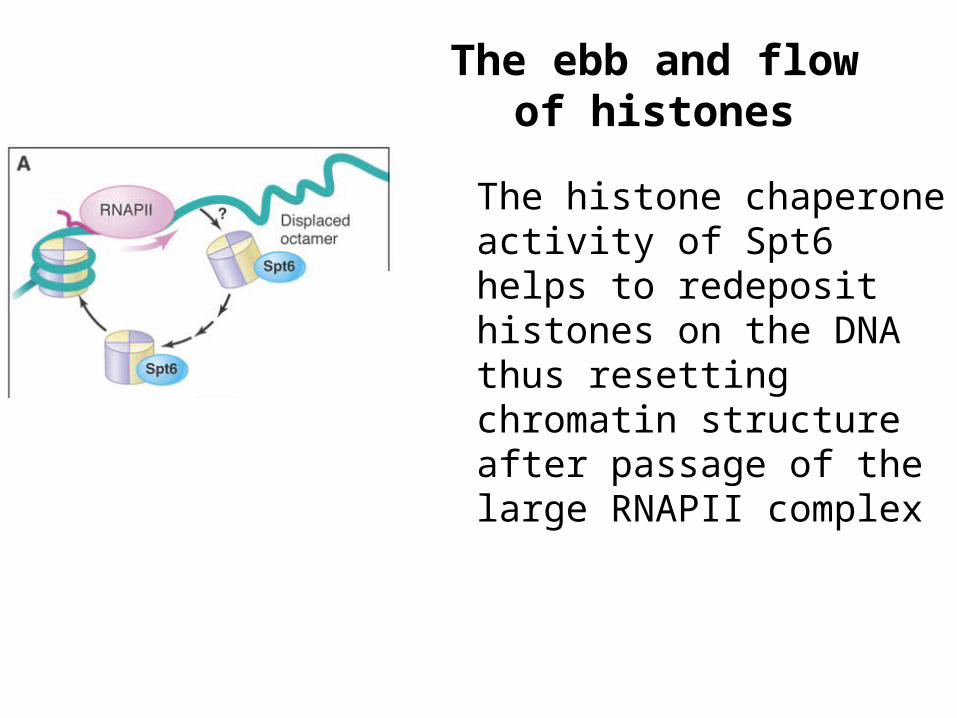
The ebb and flow of histones
The histone chaperone activity of Spt6 helps to redeposit histones on the DNA thus resetting chromatin structure after passage of the large RNAPII complex

Other elongation factors
• P-TEFb (Positive Transcription Elongation Factor) phosphorylates additional residues in CTD of PolII (some are already phosphorylated by TFIIH).
• ELL family of proteins also stimulates transcription elongation through a poorly understood mechanism. There is some evidence, that ELLs target RNA polymerase for degradation upon reaching damaged regions in DNA. Degradation of PolII is followed by recruitment of DNA repairing enzymes.
• Negative factors NELF and DSIF supress elongation by direct interaction with Pol II
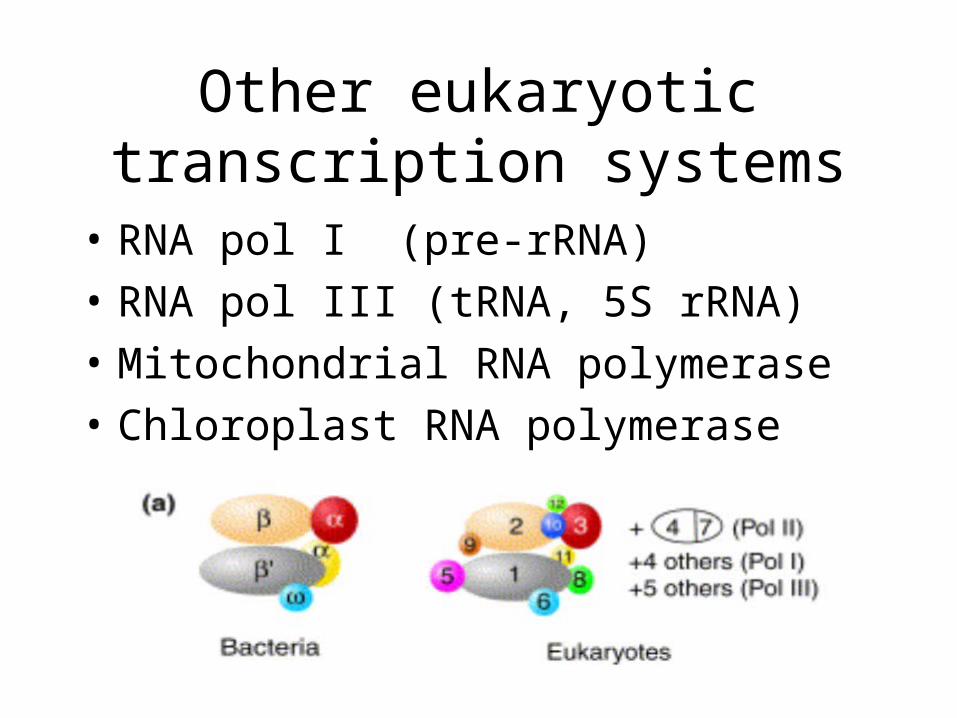
Other eukaryotic transcription systems
• RNA pol I (pre-rRNA)
• RNA pol III (tRNA, 5S rRNA)
• Mitochondrial RNA polymerase
• Chloroplast RNA polymerase
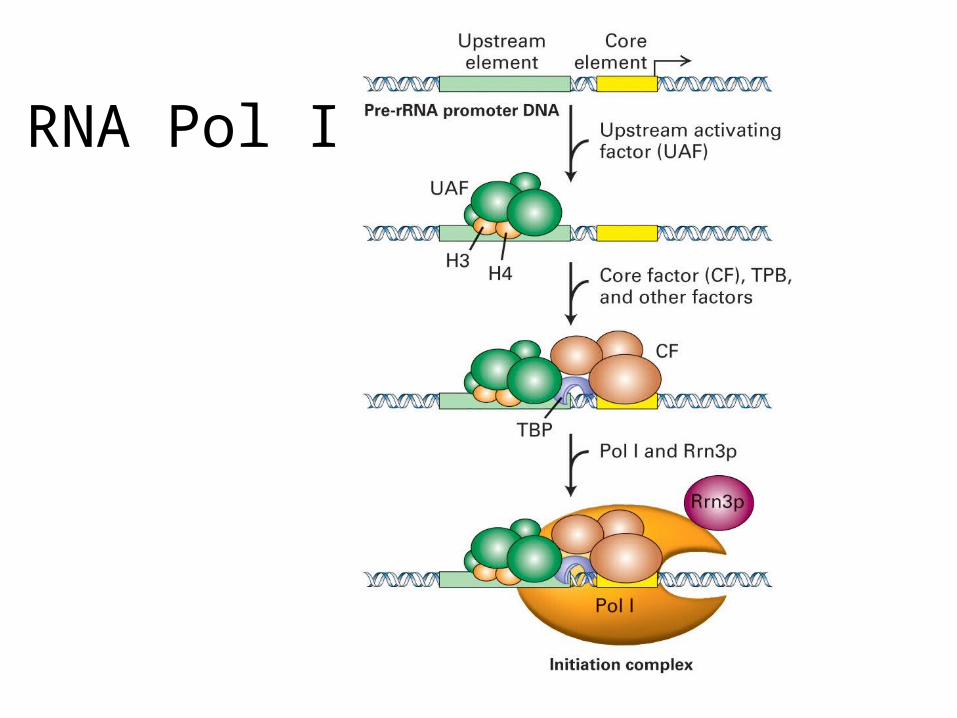
RNA Pol I
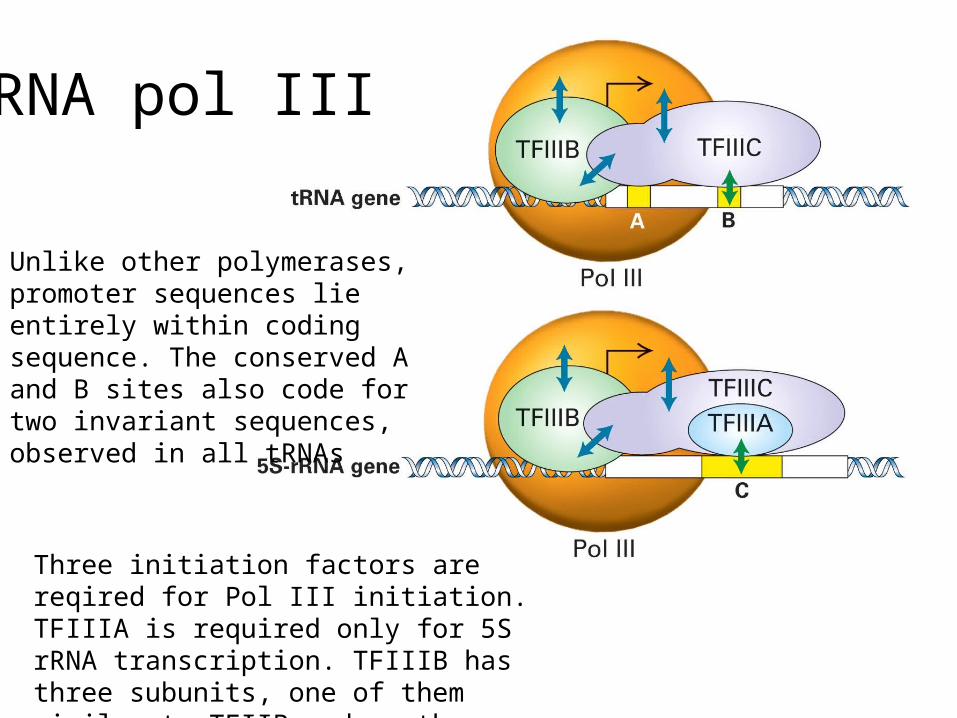
RNA pol III
Unlike other polymerases, promoter sequences lie entirely within coding sequence. The conserved A and B sites also code for two invariant sequences, observed in all tRNAs
Three initiation factors are reqired for Pol III initiation. TFIIIA is required only for 5S rRNA transcription. TFIIIB has three subunits, one of them similar to TFIIB and another one being TBP (same as for Pol I and II)

Mitochondrial RNA polymerase
• Encoded in nuclear RNA and transported to mitochondrial matrix
• Contains only two subunits
• One subunit similar to bacteriophage T7 RNA polymerase
• Other subunit similar to bacterial factor
-like
’
T7-like
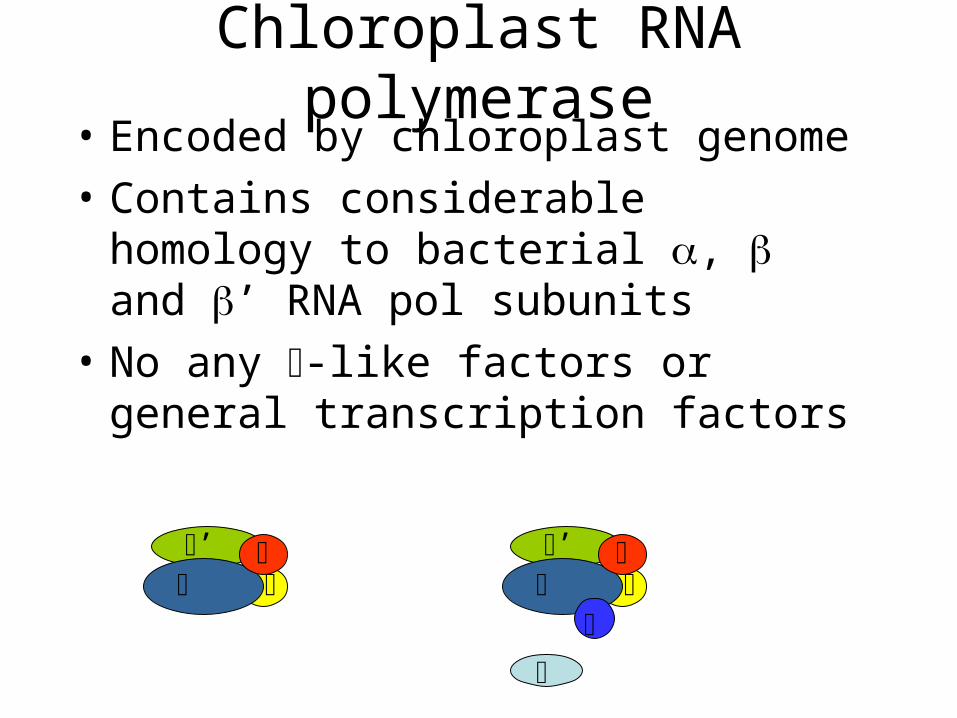
Chloroplast RNA polymerase• Encoded by chloroplast genome
• Contains considerable homology to bacterial , and ’ RNA pol subunits
• No any -like factors or general transcription factors
’
’