Embed Size (px)
Citation preview

Structural plasticity of an aminoacyl-tRNAsynthetase active siteJames M. Turner*, James Graziano*, Glen Spraggon†‡, and Peter G. Schultz*‡
*Department of Chemistry and The Skaggs Institute for Chemical Biology, The Scripps Research Institute, 10550 North Torrey Pines Road,La Jolla, CA 92037; and †Genomics Institute of the Novartis Research Foundation, 10675 John Jay Hopkins Drive, San Diego, CA 92121
Contributed by Peter G. Schultz, March 3, 2006
Recently, tRNA aminoacyl-tRNA synthetase pairs have been evolvedthat allow one to genetically encode a large array of unnatural aminoacids in both prokaryotic and eukaryotic organisms. We have deter-mined the crystal structures of two substrate-bound Methanococcusjannaschii tyrosyl aminoacyl-tRNA synthetases that charge the un-natural amino acids p-bromophenylalanine and 3-(2-naphthyl)ala-nine (NpAla). A comparison of these structures with the substrate-bound WT synthetase, as well as a mutant synthetase that chargesp-acetylphenylalanine, shows that altered specificity is due to bothside-chain and backbone rearrangements within the active site thatmodify hydrogen bonds and packing interactions with substrate, aswell as disrupt the �8-helix, which spans the WT active site. The highdegree of structural plasticity that is observed in these aminoacyl-tRNA synthetases is rarely found in other mutant enzymes withaltered specificities and provides an explanation for the surprisingadaptability of the genetic code to novel amino acids.
x-ray crystal structure � unnatural amino acids � expanded genetic code �molecular evolution
W ith the rare exceptions of selenocysteine (1) and pyr-rolysine (2, 3), the common 20 amino acids are conserved
across all known organisms. However, there does not appear tobe an inherent limit to the size or chemical nature of the geneticcode, since it has been shown that additional amino acids can begenetically encoded in both prokaryotic and eukaryotic organ-isms in response to nonsense or frameshift codons (4, 5). Thisrequires a unique codon-suppressor tRNA pair and the corre-sponding aminoacyl-tRNA synthetase, which do not crossreactwith the amino acids, tRNAs, or synthetases of the host organism(4, 5). The specificity of the aminoacyl tRNA synthetase is thenaltered by generating large libraries of active-site mutants andpassing them through positive and negative selections to identifysynthetases that selectively acylate the cognate tRNA with theunnatural amino acid but not any of the common amino acids.This approach has been used to add �30 unnatural amino acidsto the genetic codes of bacteria, yeast, and mammalian cells withhigh fidelity and efficiencies.
In Escherichia coli, a tyrosyl aminoacyl-tRNA synthetase(TyrRS) tRNACUA
Tyr pair from the archea Methanococcus jann-aschii (Mj) was used as an orthogonal tRNA synthetase pair (6).Mutant synthetases have been evolved that selectively amino-acylate their cognate suppressor tRNAs with glycosylated (7),photoreactive (8), and metal-binding amino acids, as well asamino acids with unique functional groups (9–11) . To establishthe molecular basis for the surprising adaptability of this syn-thetase, we solved (12) the structure of a mutant Mj TyrRS thatis selective for p-acetylphenylalanine (p-AcPhe). The x-ray crys-tal structure revealed significant structural changes within theenzyme active site that result from the mutations Y32L, D158G,I159C, and L162R. The Y32L and D158G mutations remove twohydrogen bonds (H-bonds) with the Tyr hydroxyl group todisfavor recognition of the natural substrate. The Y32L mutationalso forms van der Waals interactions with the methyl group ofthe p-AcPhe amino acid, and D158G enlarges the binding pocketto sterically accommodate the acetyl group, as well as allow
formation of a H-bond between the keto oxygen and Q109. TheD158G mutation in the p-AcPhe synthetase also prematurelyterminates the �8-helix. As a result, several residues in thep-AcPhe TyrRS (including H160, Y161, and L162R) move up to4 Å away from their position in the structure of the WTTyrRS–Tyr complex to optimize binding interactions with boundp-AcPhe.
Because the mutations in the Mj p-AcPhe TyrRS active siteresult in significant alterations in both side-chain and backboneconformations, we sought to determine the extent to which thisstructural plasticity can be observed in other mutants of the MjTyrRS. Thus, the structures of Mj TyrRSs specific for 3-(2-naphthyl)Ala (NpAla) (13) and p-bromophenylalanine (p-BrPhe) (11) were solved and compared with structures of theWT and p-AcPhe-specific Mj TyrRS. This analysis suggests thataltered amino acid recognition by the Mj TyrRS mutants maygenerally result from a large degree of structural plasticity in thebackbone and side-chain conformations in this active site.
Results and DiscussionExpression and Crystallization of Mutant Mj TyrRSs. The specificityof the Mj TyrRS was altered (14) by generating two synthetaselibraries, each of which is formed by randomizing five residueswithin the synthetase active site (Library 1: Tyr-32, Glu-107,Asp-158, Ile-159, and Leu-162; Library 2: Tyr-32, Asp-158,Ile-159, Leu-162, and Ala-167) based on the crystal structure ofthe homologous Bacillus stearothermophilus TyrRS–tyrosyl ad-enylate complex. These libraries were separately subjected tosubsequent rounds of positive and negative selection to identifyaminoacyl tRNA synthetases that, together with a cognate MjtRNACUA
Tyr , selectively incorporate into proteins in E. coli theunnatural amino acids p-BrPhe (Library 1) or NpAla (Library 2),in response to the amber nonsense codon. The positive selectionwas based on the suppression of an amber codon at a permissivesite in the chloramphenicol acetyl transferase gene (CmR) in thepresence of the unnatural amino acid; the negative selectioninvolved suppression of amber codons at permissive sites in thebarnase gene in the absence of the unnatural amino acid. A finalround of shuffling with subsequent positive and negative selec-tions was carried out also for the NpAla TyrRS (by using fouractive-site mutants as templates), but no additional mutationsbeyond the randomized five amino acids were found. The mutantMj TyrRS that resulted from selections with NpAla had thefollowing mutations: Y32L, D158P, I159A, L162Q, and A167V;the mutant Mj TyrRS that resulted from selections with p-BrPhehad the following mutations: Y32L, E107S, D158P, I159L, and
Conflict of interest statement: No conflicts declared.
Abbreviations: p-BrPhe, p-bromophenylalanine; NpAla, 3-(2-naphthyl)alanine; p-AcPhe,p-acetylphenylalanine; TyrRS, tyrosyl aminoacyl-tRNA synthetase; Mj, Methanococcusjannaschii.
Data deposition: The atomic coordinates for NpAla and p-BrPhe TyrRS have been depositedin the Protein Data Bank, www.pdb.org (PDB ID codes 1ZH0 and 2AG6, respectively).
‡To whom correspondence may be addressed. E-mail: [email protected] or [email protected].
© 2006 by The National Academy of Sciences of the USA
www.pnas.org�cgi�doi�10.1073�pnas.0601756103 PNAS � April 25, 2006 � vol. 103 � no. 17 � 6483–6488
BIO
CHEM
ISTR
Y

L162E. Subsequent experiments showed that the latter syn-thetase also recognizes the unnatural amino acid p-iodoPhe,which is not surprising given the similarity in van der Waals radiibetween bromine and iodine (difference � 0.2 Å). Both mutantsynthetases incorporate their respective amino acids with goodefficiency and high fidelity (�0.2% incorporation of 1 of the 20common amino acids). The p-BrPhe and NpAla Mj aminoacyl-tRNA synthetases were expressed with a C-terminal His-6 fusiontag in E. coli and purified by affinity chromatography overNi-NTA resin. Subsequent purification by cation ion-exchangechromatography provided protein for crystallization. Crystalswere grown either in the presence of 2 mM p-BrPhe or NpAlaand belong to space group P43212, containing one molecule perasymmetric unit.
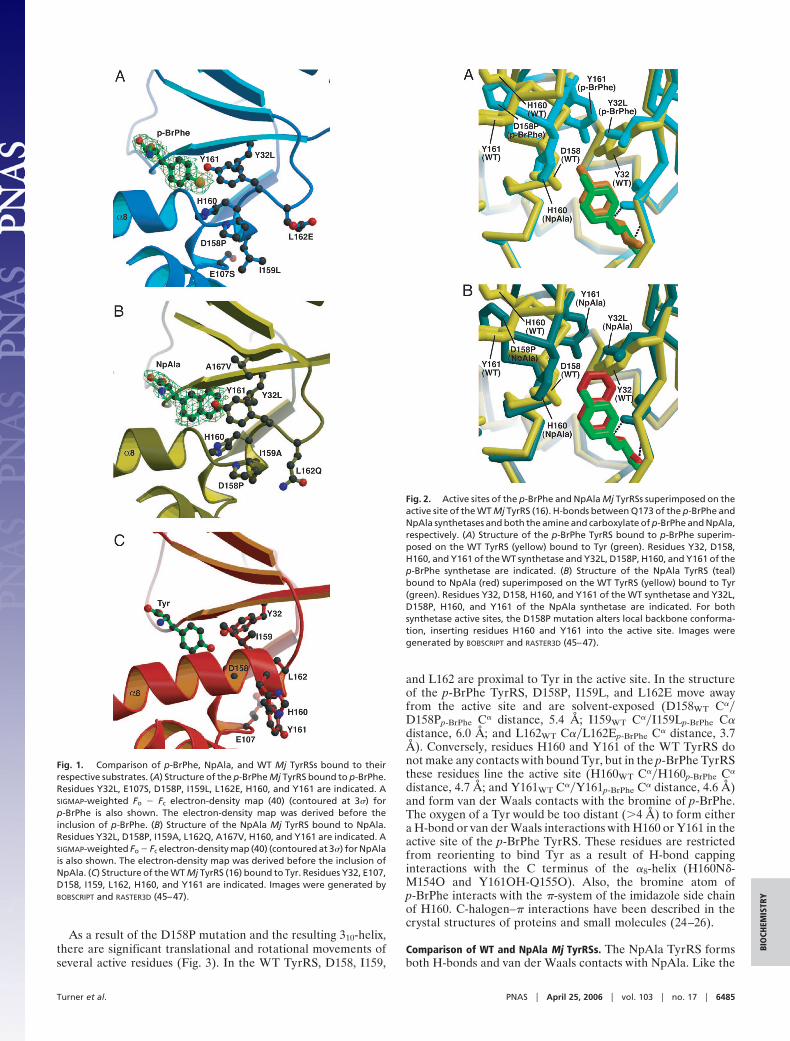
Structure Determination of Mutant Mj TyrRSs. The structure of thep-BrPhe TyrRS–p-BrPhe complex was solved to 1.9 Å, withRcryst � 0.21 and Rfree � 0.25. The structure of the NpAlaTyrRS–NpAla complex was solved to 1.9 Å, with Rcryst � 0.22and Rfree � 0.28 (Table 1). The final models for both structureshad no residues in disallowed regions of the Ramachandran plotas well as a strong difference electron density for the boundamino acid. Like the WT Mj TyrRS (15, 16), the NpAla andp-BrPhe Mj synthetases are divided into five regions (the Ross-mann-fold catalytic domain; the short N-terminal region; theconnective polypeptide 1 domain, which forms the dimer inter-face; the C-terminal domain; and the KMSKS loop, which linksthe Rossmann-fold domain to the C-terminal domain). Bothmutant synthetases superimpose well with the WT TyrRS–Tyrcomplex in all regions except the active site (rms deviations wereas follows: WT�pBrPhe, 0.83 Å, over 299 aligned C� atoms; andWT�NpAla, 0.77 Å, over 294 aligned C� atoms), where themutations in the p-BrPhe TyrRS and NpAla TyrRS significantlyreconfigure the Tyr-binding pocket to selectively bind p-BrPheor NpAla, respectively (Fig. 1).
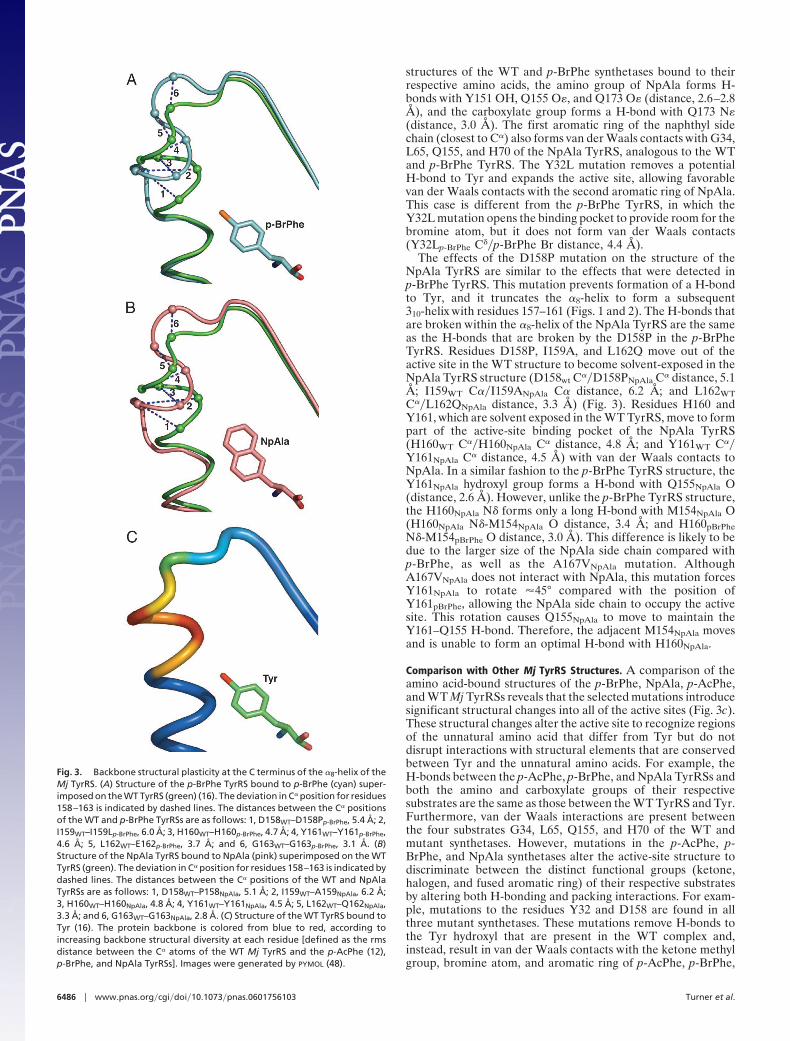
Comparison of WT and p-BrPhe Mj TyrRSs. In the WT structure (16),recognition of Tyr involves a combination of H-bonding inter-actions and van der Waals contacts. The amino group of Tyrforms H-bonds with Y151 OH, Q155 O�, and Q173 O�; theTyr carboxylate group forms a H-bond with Q173 N�; andthe Tyr hydroxyl group forms H-bonds with Y32 OH and D158O�. There are van der Waals contacts between the Tyr side chainand G34, L65, Q155, and H70. The mutations to the p-BrPhe MjTyrRS alter the active-site structure to specifically recognizep-BrPhe vs. Tyr while maintaining contacts to the conservedregions of the two amino acid substrates. For example, the aminoand carboxylate groups of p-BrPhe form the same H-bonds as inthe WT complex, and the aromatic ring of p-BrPhe again formsvan der Waals contacts with G34, L65, Q155, and H70. However,the D158P and Y32L mutations in the p-BrPhe TyrRS enlargethe binding site to accommodate p-BrPhe and remove the twoH-bonds to the hydroxyl oxygen of Tyr that are present in theWT structure (Fig. 2) (16). The D158P mutation also terminatesthe �8-helix, resulting in formation of a short 310-helix consistingof residues 157–161. The H-bonds that are disrupted within the�8-helix (Met154O–Asp158NH, Gln155O–Ile159NH, Val156O–His160NH, Asn157O–Tyr161NH, and Ile159O–Gly163NH) arepartially compensated by H-bond capping interactions (17–20)from H160 and Y161 (His160N�–Met154O, Tyr161OH–Gln155O), as well as an additional side-chain H-bond(Tyr114OH–Met154O). The short 310-helix (after �8-helix) isformed by two H-bonds (Asn157O–His160NH and Pro158O–Tyr161NH) and is capped at the N and C termini by Asn157O�–Leu159NH and Tyr161O–Val164NH, respectively. Consistentwith this observation, it is known that there is a 3.5:1 preferencefor Asn as the N-capping residue and a 2.6:1 preference for Proas the subsequent residue of an �-helix (17); studies of 310-heliceshave shown similar results (21). Other examples of proteinscontaining a 310-helix sequence that is contiguous with an �-helixsequence (21) are hemoglobin (22) and Arthromyces ramosusperoxidase (23).
Table 1. Data and refinement statistics for the p-BrPhe Mj TyrRS–p-BrPhe and NpAla MjTyrRS–NpAla complexes
Parameter p-BrPhe NpAla
Space group P43212 P43212Unit-cell parameters, Å a � b � 102.98, c � 70.71 a � b � 103.0, c � 71.4Wavelength, Å 1.0 1.0Resolution range, Å 70.0–1.9 70.0–1.9Rsymm (highest-resolution shell) 0.072 (0.74) 0.053 (0.67)Unique reflections (observed) 30,607 (166,252) 31,188 (91,993)Completeness (highest-resolution shell), % 99.4 (98.3) 97.1 (93.2)Highest-resolution shell, Å 1.97–1.9 2.03–1.9Mean I��(I) 39.9 (2.6) 32.8 (2.0)Refinement parameters
Reflections, total 28,644 28,143Reflections, test 1,538 1,507Rcryst* (Rfree
†) 0.21 (0.25) 0.22 (0.28)No. of protein atoms 2,464 2,461No. of heterogen atoms 13 24No. of water atoms 142 115rmsd bonds, Å 0.013 0.015rmsd angles, ° 1.47 1.57Average isotropic B value, Å2 41.5 50.7
Protein 40.9 55.0Amino acid 33.2 44.0Waters 45.9 57.9
*Rcryst � ���Ii � �Ii�����Ii�, where Ii is the scaled intensity of the ith measurement, and �Ii� is the mean intensity forthat reflection.
†As for Rcryst, but for 5.0% of the total reflections chosen at random and omitted from refinement.
6484 � www.pnas.org�cgi�doi�10.1073�pnas.0601756103 Turner et al.
As a result of the D158P mutation and the resulting 310-helix,there are significant translational and rotational movements ofseveral active residues (Fig. 3). In the WT TyrRS, D158, I159,
and L162 are proximal to Tyr in the active site. In the structureof the p-BrPhe TyrRS, D158P, I159L, and L162E move awayfrom the active site and are solvent-exposed (D158WT C��D158Pp-BrPhe C� distance, 5.4 Å; I159WT C��I159Lp-BrPhe C�distance, 6.0 Å; and L162WT C��L162Ep-BrPhe C� distance, 3.7Å). Conversely, residues H160 and Y161 of the WT TyrRS donot make any contacts with bound Tyr, but in the p-BrPhe TyrRSthese residues line the active site (H160WT C��H160p-BrPhe C�
distance, 4.7 Å; and Y161WT C��Y161p-BrPhe C� distance, 4.6 Å)and form van der Waals contacts with the bromine of p-BrPhe.The oxygen of a Tyr would be too distant (�4 Å) to form eithera H-bond or van der Waals interactions with H160 or Y161 in theactive site of the p-BrPhe TyrRS. These residues are restrictedfrom reorienting to bind Tyr as a result of H-bond cappinginteractions with the C terminus of the �8-helix (H160N�-M154O and Y161OH-Q155O). Also, the bromine atom ofp-BrPhe interacts with the �-system of the imidazole side chainof H160. C-halogen–� interactions have been described in thecrystal structures of proteins and small molecules (24–26).
Comparison of WT and NpAla Mj TyrRSs. The NpAla TyrRS formsboth H-bonds and van der Waals contacts with NpAla. Like the
Fig. 1. Comparison of p-BrPhe, NpAla, and WT Mj TyrRSs bound to theirrespective substrates. (A) Structure of the p-BrPhe Mj TyrRS bound to p-BrPhe.Residues Y32L, E107S, D158P, I159L, L162E, H160, and Y161 are indicated. ASIGMAP-weighted Fo � Fc electron-density map (40) (contoured at 3�) forp-BrPhe is also shown. The electron-density map was derived before theinclusion of p-BrPhe. (B) Structure of the NpAla Mj TyrRS bound to NpAla.Residues Y32L, D158P, I159A, L162Q, A167V, H160, and Y161 are indicated. ASIGMAP-weighted Fo � Fc electron-density map (40) (contoured at 3�) for NpAlais also shown. The electron-density map was derived before the inclusion ofNpAla. (C) Structure of the WT Mj TyrRS (16) bound to Tyr. Residues Y32, E107,D158, I159, L162, H160, and Y161 are indicated. Images were generated byBOBSCRIPT and RASTER3D (45–47).
Fig. 2. Active sites of the p-BrPhe and NpAla Mj TyrRSs superimposed on theactive site of the WT Mj TyrRS (16). H-bonds between Q173 of the p-BrPhe andNpAla synthetases and both the amine and carboxylate of p-BrPhe and NpAla,respectively. (A) Structure of the p-BrPhe TyrRS bound to p-BrPhe superim-posed on the WT TyrRS (yellow) bound to Tyr (green). Residues Y32, D158,H160, and Y161 of the WT synthetase and Y32L, D158P, H160, and Y161 of thep-BrPhe synthetase are indicated. (B) Structure of the NpAla TyrRS (teal)bound to NpAla (red) superimposed on the WT TyrRS (yellow) bound to Tyr(green). Residues Y32, D158, H160, and Y161 of the WT synthetase and Y32L,D158P, H160, and Y161 of the NpAla synthetase are indicated. For bothsynthetase active sites, the D158P mutation alters local backbone conforma-tion, inserting residues H160 and Y161 into the active site. Images weregenerated by BOBSCRIPT and RASTER3D (45–47).
Turner et al. PNAS � April 25, 2006 � vol. 103 � no. 17 � 6485
BIO
CHEM
ISTR
Y
structures of the WT and p-BrPhe synthetases bound to theirrespective amino acids, the amino group of NpAla forms H-bonds with Y151 OH, Q155 O�, and Q173 O� (distance, 2.6–2.8Å), and the carboxylate group forms a H-bond with Q173 N�(distance, 3.0 Å). The first aromatic ring of the naphthyl sidechain (closest to C�) also forms van der Waals contacts with G34,L65, Q155, and H70 of the NpAla TyrRS, analogous to the WTand p-BrPhe TyrRS. The Y32L mutation removes a potentialH-bond to Tyr and expands the active site, allowing favorablevan der Waals contacts with the second aromatic ring of NpAla.This case is different from the p-BrPhe TyrRS, in which theY32L mutation opens the binding pocket to provide room for thebromine atom, but it does not form van der Waals contacts(Y32Lp-BrPhe C��p-BrPhe Br distance, 4.4 Å).
The effects of the D158P mutation on the structure of theNpAla TyrRS are similar to the effects that were detected inp-BrPhe TyrRS. This mutation prevents formation of a H-bondto Tyr, and it truncates the �8-helix to form a subsequent310-helix with residues 157–161 (Figs. 1 and 2). The H-bonds thatare broken within the �8-helix of the NpAla TyrRS are the sameas the H-bonds that are broken by the D158P in the p-BrPheTyrRS. Residues D158P, I159A, and L162Q move out of theactive site in the WT structure to become solvent-exposed in theNpAla TyrRS structure (D158wt C��D158PNpAla C� distance, 5.1Å; I159WT C��I159ANpAla C� distance, 6.2 Å; and L162WTC��L162QNpAla distance, 3.3 Å) (Fig. 3). Residues H160 andY161, which are solvent exposed in the WT TyrRS, move to formpart of the active-site binding pocket of the NpAla TyrRS(H160WT C��H160NpAla C� distance, 4.8 Å; and Y161WT C��Y161NpAla C� distance, 4.5 Å) with van der Waals contacts toNpAla. In a similar fashion to the p-BrPhe TyrRS structure, theY161NpAla hydroxyl group forms a H-bond with Q155NpAla O(distance, 2.6 Å). However, unlike the p-BrPhe TyrRS structure,the H160NpAla N� forms only a long H-bond with M154NpAla O(H160NpAla N�-M154NpAla O distance, 3.4 Å; and H160pBrPheN�-M154pBrPhe O distance, 3.0 Å). This difference is likely to bedue to the larger size of the NpAla side chain compared withp-BrPhe, as well as the A167VNpAla mutation. AlthoughA167VNpAla does not interact with NpAla, this mutation forcesY161NpAla to rotate �45° compared with the position ofY161pBrPhe, allowing the NpAla side chain to occupy the activesite. This rotation causes Q155NpAla to move to maintain theY161–Q155 H-bond. Therefore, the adjacent M154NpAla movesand is unable to form an optimal H-bond with H160NpAla.
Comparison with Other Mj TyrRS Structures. A comparison of theamino acid-bound structures of the p-BrPhe, NpAla, p-AcPhe,and WT Mj TyrRSs reveals that the selected mutations introducesignificant structural changes into all of the active sites (Fig. 3c).These structural changes alter the active site to recognize regionsof the unnatural amino acid that differ from Tyr but do notdisrupt interactions with structural elements that are conservedbetween Tyr and the unnatural amino acids. For example, theH-bonds between the p-AcPhe, p-BrPhe, and NpAla TyrRSs andboth the amino and carboxylate groups of their respectivesubstrates are the same as those between the WT TyrRS and Tyr.Furthermore, van der Waals interactions are present betweenthe four substrates G34, L65, Q155, and H70 of the WT andmutant synthetases. However, mutations in the p-AcPhe, p-BrPhe, and NpAla synthetases alter the active-site structure todiscriminate between the distinct functional groups (ketone,halogen, and fused aromatic ring) of their respective substratesby altering both H-bonding and packing interactions. For exam-ple, mutations to the residues Y32 and D158 are found in allthree mutant synthetases. These mutations remove H-bonds tothe Tyr hydroxyl that are present in the WT complex and,instead, result in van der Waals contacts with the ketone methylgroup, bromine atom, and aromatic ring of p-AcPhe, p-BrPhe,
Fig. 3. Backbone structural plasticity at the C terminus of the �8-helix of theMj TyrRS. (A) Structure of the p-BrPhe TyrRS bound to p-BrPhe (cyan) super-imposed on the WT TyrRS (green) (16). The deviation in C� position for residues158–163 is indicated by dashed lines. The distances between the C� positionsof the WT and p-BrPhe TyrRSs are as follows: 1, D158WT–D158Pp-BrPhe, 5.4 Å; 2,I159WT–I159Lp-BrPhe, 6.0 Å; 3, H160WT–H160p-BrPhe, 4.7 Å; 4, Y161WT–Y161p-BrPhe,4.6 Å; 5, L162WT–E162p-BrPhe, 3.7 Å; and 6, G163WT–G163p-BrPhe, 3.1 Å. (B)Structure of the NpAla TyrRS bound to NpAla (pink) superimposed on the WTTyrRS (green). The deviation in C� position for residues 158–163 is indicated bydashed lines. The distances between the C� positions of the WT and NpAlaTyrRSs are as follows: 1, D158WT–P158NpAla, 5.1 Å; 2, I159WT–A159NpAla, 6.2 Å;3, H160WT–H160NpAla, 4.8 Å; 4, Y161WT–Y161NpAla, 4.5 Å; 5, L162WT–Q162NpAla,3.3 Å; and 6, G163WT–G163NpAla, 2.8 Å. (C) Structure of the WT TyrRS bound toTyr (16). The protein backbone is colored from blue to red, according toincreasing backbone structural diversity at each residue [defined as the rmsdistance between the C� atoms of the WT Mj TyrRS and the p-AcPhe (12),p-BrPhe, and NpAla TyrRSs]. Images were generated by PYMOL (48).
6486 � www.pnas.org�cgi�doi�10.1073�pnas.0601756103 Turner et al.

and NpAla, respectively. The D158G mutation in the p-AcPheTyrRS and the D158P mutation in the p-BrPhe TyrRS andNpAla TyrRS also remove the second H-bond to Tyr. Also, thesemutations change the backbone structure and side-chain con-formations in the active site. The backbone conformationalchanges that are due to the D158P mutations in both the p-BrPheTyrRS and NpAla TyrRS are more prominent than the changesthat are due to the D158G mutation in the p-AcPhe TyrRS.Although the p-AcPhe D158G mutation allows the formation ofa H-bond between Q109 N� and the p-AcPhe carbonyl oxygenby removing the intervening side chain of D158, residues H160and Y161 of the p-AcPhe TyrRS do not move to form part of thenew active site (as they do in the p-BrPhe TyrRS and NpAlaTyrRS). The C� of H160p-AcPhe moves 2.7 Å, and the C� ofY161p-AcPhe moves 3.6 Å relative to the WT enzyme; for com-parison, H160pBrPhe C� and H160NpAla C� move 4.7 and 4.8 Å,respectively, and Y161p-BrPhe C� and Y161NpAla C� move 4.6 and4.5 Å, respectively. This difference between the p-AcPhe TyrRSand the p-BrPhe and NpAla TyrRS may be a consequence ofdifferent mechanisms of recognition. Wheras the WT andp-AcPhe Mj TyrRSs recognize their substrates by H-bonding aswell as van der Waals interactions, the hydrophobic nature ofp-BrPhe and NpAla may require larger changes in both side-chain and backbone structure in the active site to form comple-mentary van der Waals surfaces as well as a C-halogen–�interaction between H160 and the bromine atom (in the case ofthe p-BrPhe TyrRS).
Another mutation that changes amino acid specificity withoutmaking any contacts with the amino acid is A167V of the NpAlaTyrRS. The larger side chain of this mutation causes the Y161side chain to rotate, which affords additional space for NpAla.Without this mutation, residues H160 and Y161 of the NpAlaTyrRS would be likely to occupy the same positions as in thep-BrPhe TyrRS. Superimposition of these two structures revealsa steric clash between NpAla and Y161p-BrPhe but not Y161NpAla.Therefore, even though A167VNpAla does not interact withNpAla, it is important for binding because it expands the activesite by rotating Y161NpAla. I159L and L162E of the p-BrPheTyrRS, L162R of p-AcPhe TyrRS, and I159A and L162Q of theNpAla TyrRS are all distal to the active site, and these mutationswere likely to have been chosen to maximize physical propertiesof the protein.
Role of Structural Plasticity in Biomolecular Recognition. Previousstructural studies of aminoacyl-tRNA synthetases that recognizeunnatural amino acids have shown little change in the backbonestructures compared with the WT structure. For example, boththe substrate-bound and Tyr-bound structures of a mutant E. colityrosyl synthetase that recognizes 3-iodo-L-Tyr have very similarbackbone traces as the WT synthetase bound to Tyr (27). Thebackbone structure of tryptophanyl tRNA synthetase fromDeinococcus radiodurans bound to 5-hydroxy-Trp (5HT) is sim-ilar to the Trp-bound structure, although the orientation of 5HTin the active site is rotated by almost 180° compared with Trp(28). Indeed, most structurally characterized mutant enzymeswith altered substrate specificity [including mutants of isocitratedehydrogenase, isopropylmalate dehydrogenase (29, 30), andAsp aminotransferase (AspAT) (31)] do not involve significantmain-chain conformational changes. For example, the specificityof the WT AspAT was changed from Asp to Val by directedevolution, but a comparison of the backbone structures of theWT and mutant AspAT shows that only one residue (S363G)deviates by �2 Å, which is likely to be due at least in part to thismutation occurring next to an adjacent Gly residue (31).
However, it is clear that mutations and modifications cansignificantly alter protein backbone conformation and, as aconsequence, protein function. For example, Thr phosphoryla-tion activates yeast glycogen phosphorylase by refolding the
N-terminal loop into an �-helix (32), and several studies haveidentified mutations that cause changes in the topological fold ofthe RNA-binding protein Rop (33, 34). Another example inwhich both backbone and side-chain conformational changeshave a significant role in binding specificity is the affinitymaturation of germ-line antibodies (35, 36). In this case, somaticpoint mutations can act to lock significant conformationalchanges induced by binding of ligands to the germ-line antibody.
There are two factors (12) that likely lead to the observedstructural plasticity of the p-AcPhe, p-BrPhe, and NpAla MjTyrRSs. First, because the changes are on a solvent-exposedhelix, the number of requisite compensatory mutations is re-duced, because bulk water can more readily reorganize to solvatevarious new conformations than adjacent protein residues. Sec-ond, the focused library restricts mutations to active-site residuesin local proximity, which increases the probability of favorablecooperative interactions. In contrast, mutations that arise fromDNA shuffling are recombined from independent selectionexperiments, and the resulting set of mutations may not becooperative. Random point mutations also have a low proba-bility of being cooperative because of the limited size of se-quence space that can be explored. In contrast, successive roundsof point mutations such as those that occur during affinitymaturation are more likely to act cooperatively. In one notableexample, iterative rounds of mutations in the Arc repressorhomodimer (guided by modeling, NMR, and thermal denaturingstudies) showed that interchanging the sequence of a hydropho-bic core residue with an adjacent surface polar residue convertedan N-terminal �-sheet into a 310-helix (37).
As in the case of germ-line antibodies, it may be possible thatsubstrate binding induces some of the conformational changesobserved in the p-BrPhe and NpAla Mj synthetase structures.However, preliminary low-resolution studies suggest that thisstructural change is present in the apo forms as well. Also, theWT Mj TyrRS does not show significant backbone changesbetween the free and Tyr-bound form of the enzyme. High-resolution studies with the apo form of the p-BrPhe and NpAlaMj TyrRSs, as well as other synthetases specific for glycosylatedand photoreactive amino acids, may provide additional insight.
The structures of the p-BrPhe, NpAla, and p-AcPhe MjTyrRSs show that mutation of a few residues within the activesite can induce very significant conformational changes in theside chains and the backbone of the enzyme. These changes canalter interactions with amino acid directly, as well as modulatethe interactions between the amino acid and other residues.Because these changes are currently difficult to predict, struc-tural characterization of other mutant Mj aminoacyl-tRNAsynthetases may reveal unexpected structural motifs that suggestmethods to improve and�or expand library design. For example,we have generated a library with a D158P mutation and ran-domized active-site residues (Y32, H160, and Y161) to takeadvantage of the scaffold that is present in the p-BrPhe TyrRSand NpAla Mj TyrRS.
Materials and MethodsExpression, Purification, and Crystallization of Mutant Mj TyrRSs.DNA fragments encoding the p-BrPhe and NpAla TyrRSs wereamplified by PCR and cloned into the NdeI�XhoI sites of theexpression vector pET22b. This vector was transformed inBL21(DE3) cells (Novagen) and grown to an OD of 0.8–1.0.After induction overnight with 500 mM isopropyl-�-D-thiogalactopyranoside, cells were pelleted by centrifugation andresuspended in lysis buffer [50 mM Hepes, pH 7.9 (Sigma)�500mM NaCl (Fisher)�10 mM 2-mercaptoethanol (Sigma)�10%glycerol (Sigma)�0.1% Triton X-100 (Sigma)�5 mM imidazole(Sigma)] and frozen overnight at �80°C. After thawing on ice,cells were sonicated, and insoluble cell debris was pelleted bycentrifugation. The lysate was incubated with Ni-NTA agarose
Turner et al. PNAS � April 25, 2006 � vol. 103 � no. 17 � 6487
BIO
CHEM
ISTR
Y

(Qiagen, Valencia, CA) for 1 h at 4°C, filtered, and washed withwash buffer (50 mM Hepes, pH 7.9�500 mM NaCl�10 mM�-mercaptoethanol�5 mM imidazole). The synthetase waseluted in wash buffer with 250 mM imidazole, dialyzed in bufferA (25 mM Tris, pH 8.5�25 mM NaCl�10 mM 2-mercaptoetha-nol�1 mM EDTA); purified by anion exchange (MonoQ; Am-ersham Biosciences) by using buffer A with gradient elutionfrom 25 mM to 1 M NaCl; dialyzed in 20 mM Tris, pH 8.5�50mM NaCl�10 mM 2-mercaptoethanol; and concentrated to20–30 mg�ml. Crystals of the mutant TyrRS–amino acid com-plex were grown at 22°C or 4°C by using the sitting-dropvapor-diffusion technique against a mother liquor composed of20–16% polyethylene glycol (PEG) 300, 5–3% PEG 8000, 100mM Tris (pH 8.8–8.2), and 10% glycerol by using a 1:1 mixtureof concentrated protein (15 mg�ml) with 2 mM amino acid(p-BrPhe or NpAla) and mother liquor.
Data Collection, Refinement, and Structure Determination. For theNpAla TyrRS and p-BrPhe Mj TyrRS, data were collected atbeamline 5.0.3 of the Advanced Light Source at a temperatureof 100 K and a wavelength of 1.0 Å to a maximum Bragg spacingof 1.9 Å. All data were reduced and scaled by using the HKL2000package (38). The NpAla Mj TyrRS structure was solved by usingMOLREP (39). We used all data with a resolution of 50–4.0 Å, andthe WT TyrRS [Protein Data Bank (PDB) ID code 1J1U] (16)was used as a probe model. The p-BrPhe Mj TyrRS structure was
solved by rigid body refinement with the NpAla Mj TyrRS.Differences between the WT structure and both the NpAla andp-BrPhe structures were clearly evident in SIGMAA weightedelectron-density maps (40) at this point. Model-building andrefinement were then carried out with O (41) and REFMAC (42).Automated water-building was carried out with ARP/WARP (43),and manual inspection of the waters was then performed. Allother crystallographic manipulations were carried out with theCCP4 program suite (44). The final model of the p-BrPhe MjTyrRS had an Rcryst and Rfree of 0.21 and 0.25, respectively,whereas the final model of the NpAla Mj TyrRS had an Rcryst andRfree of 0.22 and 0.28, respectively. Both models have no residuesin disallowed regions of the Ramachandran plot. Coordinates forthe NpAla and p-BrPhe synthetases have been deposited in thePDB (PDB ID codes 1ZH0 and 2AG6, respectively).
We thank the staff of the Lawrence Berkeley National Laboratory(Berkeley) Advanced Light Source beamlines for their continued sup-port; Eileen Ambing for technical assistance in looping the crystals; YanZhang for helpful discussions; and Andreas Kreusch, Christian Lee,Michael Didonato, Phillip Chamberlain, and Scott Lesley for datacollection. This work is supported by National Institutes of Health GrantGM62159, Department of Energy Grant DE-FG03-00ER46051, and TheSkaggs Institute for Chemical Biology. J.M.T. was supported by aNational Institutes of Health postdoctoral fellowship. The AdvancedLight Source at Lawrence Berkeley National Laboratory is supported byDepartment of Energy Material Sciences Division Contract DE-AC03-76SF00098.
1. Bock, A., Forchhammer, K., Heider, J., Leinfelder, W., Sawers, G., Veprek, B.& Zinoni, F. (1991) Mol. Microbiol. 5, 515–520.
2. Srinivasan, G., James, C. M. & Krzycki, J. A. (2002) Science 296, 1459–1462.3. Polycarpo, C., Ambrogelly, A., Berube, A., Winbush, S. M., McCloskey, J. A.,
Crain, P. F., Wood, J. L. & Soll, D. (2004) Proc. Natl. Acad. Sci. USA 101,12450–12454.
4. Wang, L. & Schultz, P. G. (2004) Angew. Chem. Int. Ed. 44, 34–66.5. Wang, L., Xie, J. & Schultz, P. G. (2006) Annu. Rev. Biophys. Biomol. Struct.
35, 225–249.6. Wang, L., Brock, A., Herberich, B. & Schultz, P. G. (2001) Science 292,
498–500.7. Zhang, Z., Gildersleeve, J., Yang, Y.-Y., Xu, R., Loo, J. A., Uryu, S., Wong,
C.-H. & Schultz, P. G. (2004) Science 303, 371–373.8. Chin, J. W., Martin, A. B., King, D. S., Wang, L. & Schultz, P. G. (2002) Proc.
Natl. Acad. Sci. USA 99, 11020–11024.9. Wang, L., Zhang, Z., Brock, A. & Schultz, P. G. (2003) Proc. Natl. Acad. Sci.
USA 100, 56–61.10. Chin, J. W., Santoro, S. W., Martin, A. B., King, D. S., Wang, L. & Schultz, P. G.
(2002) J. Am. Chem. Soc. 124, 9026–9027.11. Xie, J., Wang, L., Wu, N., Brock, A., Spraggon, G. & Schultz, P. G. (2004) Nat.
Biotechnol. 22, 1297–1301.12. Turner, J. M., Graziano, J., Spraggon, G. & Schultz, P. G. (2005) J. Am. Chem.
Soc. 127, 14976–14977.13. Wang, L., Brock, A. & Schultz, P. G. (2002) J. Am. Chem. Soc. 124, 1836–1837.14. Brick, P., Bhat, T. N. & Blow, D. M. (1989) J. Mol. Biol. 208, 83–98.15. Zhang, Y., Wang, L., Schultz, P. G. & Wilson, I. A. (2005) Protein Sci. 14,
1340–1349.16. Kobayashi, T., Nureki, O., Ishitani, R., Yaremchuk, A., Tukalo, M., Cusack, S.,
Sakamoto, K. & Yokoyama, S. (2003) Nat. Struct. Biol. 10, 425–432.17. Richardson, J. S. & Richardson, D. C. (1988) Science 240, 1648–1652.18. Zhou, H. X., Lyu, P., Wemmer, D. E. & Kallenbach, N. R. (1994) Proteins
Struct. Funct. Genet. 18, 1–7.19. Lyu, P. C., Wemmer, D. E., Zhou, H. X., Pinker, R. J. & Kallenbach, N. R.
(1993) Biochemistry 32, 421–425.20. Presta, L. G. & Rose, G. D. (1988) Science 240, 1632–1641.21. Pal, L. & Basu, G. (1999) Protein Eng. 12, 811–814.22. Vasseur, C., Blouquit, Y., Kister, J., Prome, D., Kavanaugh, J. S., Rogers, P. H.,
Guillemin, C., Arnone, A., Galacteros, F., Poyart, C., et al. (1992) J. Biol. Chem.267, 12682–12691.
23. Fukuyama, K., Kunishima, N., Amada, F., Kubota, T. & Matsubara, H. (1995)J. Biol. Chem. 270, 21884–21892.
24. Saraogi, I., Vijay, V. G., Das, S., Sekar, K. & Row, T. N. G. (2003) Crystal Eng.6, 69–77.
25. Nagaraj, B., Narasimhamurthy, T., Yathirajan, H. S., Nagaraja, P., Naraseg-owda, R. S. & Rathore, R. S. (2005) Acta Crystallogr. C 61, o177–o180.
26. Prusiner, P. & Sundaralingam, M. (1973) Acta Crystallogr. B 29, 2328–2334.27. Kobayashi, T., Sakamoto, K., Takimura, T., Sekine, R., Vincent, K., Kamata,
K., Nishimura, S. & Yokoyama, S. (2005) Proc. Natl. Acad. Sci. USA 102,1366–1371.
28. Buddha, M. R. & Crane, B. R. (2005) Nat. Struct. Mol. Biol. 12, 274–275.29. Hurley, J. H., Chen, R. & Dean, A. M. (1996) Biochemistry 35, 5670–5678.30. Chen, R., Greer, A. & Dean, A. M. (1996) Proc. Natl. Acad. Sci. USA 93,
12171–12176.31. Oue, S., Okamoto, A., Yano, T. & Kagamiyama, H. (1999) J. Biol. Chem. 274,
2344–2349.32. Lin, K., Rath, V. L., Dai, S. C., Fletterick, R. J. & Hwang, P. K. A. (1996)
Science 273, 1539–1541.33. Willis, M. A., Bishop, B., Regan, L. & Brunger, A. T. (2000) Structure (London)
8, 1319–1328.34. Glykos, N. M., Cesareni, G. & Kokkinidis, M. (1999) Structure (London) 7,
597–603.35. Yin, J., Beuscher, A. E., IV, Andryski, S. E., Stevens, R. C. & Schultz, P. G.
(2003) J. Mol. Biol. 330, 651–656.36. Wedemayer, G. J., Patten, P. A., Wang, L. H., Schultz, P. G. & Stevens, R. C.
(1997) Science 276, 1665–1669.37. Cordes, M. H. J., Walsh, N. P., McKnight, C. J. & Sauer, R. T. (2003) J. Mol.
Biol. 326, 899–909.38. Otwinowski, Z. & Minor, W. (1997) Methods Enzymol. 276, 307–326.39. Vagin, A. & Teplyakov, A. (1997) J. Appl. Cryst. 30, 1022–1025.40. Read, R. J. (1986) Acta Crystallogr. A 42, 140–149.41. Jones, T. A., Zou, J.-Y., Cowan, S. W. & Kjeldgaard, M. (1991) Acta Crystallogr.
A 47, 110–119.42. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. (1997) Acta Crystallogr. D 53,
240–255.43. Lamzin, V. S. & Wilson, K. S. (1993) Acta Crystallogr. D 49, 129–147.44. Project, C. C. (1994) Acta Crystallogr. D 50, 760–763.45. Esnouf, R. M. (1997) J. Mol. Graphics Model. 15, 132–134.46. Kraulis, P. J. (1991) J. Appl. Crystallogr. 24, 946–950.47. Merritt, E. A. & Murphy, M. E. P. (1994) Acta Crystallogr. D 50, 869–873.48. DeLano, W. L. (2002) The PYMOL User’s Manual (DeLano Scientific, San
Carlos, CA).
6488 � www.pnas.org�cgi�doi�10.1073�pnas.0601756103 Turner et al.




![An Aminoacyl-tRNA Synthetase Complex Escherichia coli(iii) Freezepress. Cellpellets weesuspendedin 2volumes ofbuffer Aandaddedto afreeze press (Eatonmodification oftheHughespress[7]),](https://img.pdfslide.us/doc/110x75/5e4dfce8f29b5d54b52a0e06/an-aminoacyl-trna-synthetase-complex-escherichia-coli-iii-freezepress-cellpellets.jpg)
























