Embed Size (px)
Citation preview

41
CHAPTER 4
STANDARDIZATION OF PROTOCOLS FOR CALLUS INDUCTION, SOMATIC
EMBRYOGENESIS AND ORGANOGENESIS IN AQUILARIA MALACCENSIS LAM.
4.1. Introduction
Different techniques in plant tissue culture offer certain advantages over
traditional methods of propagation, such as, production of exact copies of plants that
produce particularly good flowers, fruits, or have other desirable traits. Production of
secondary metabolites, genotype modification, germplasm conservation, etc. may also
be achieved through this technique. Basically the technique consists of taking a piece of
a plant (such as a stem tip, node, meristem, embryo, or even a seed/leaf) and placing it
on a sterile, (usually gel -based) nutrient medium where it multiplies freely without
interference by any other organism. The formulation of the growth medium is changed
depending on trial and error basis to get the plant that produce undifferentiated cell
mass, multiplication of the number of plantlets, development of roots, or multiplication
of embryos for artificial seed. Plant Growth regulators (PGR) play an important role
over all other factors. Plant tissue culture has been widely employed as an experimental
technique for studying somatic embryogenesis and organogenesis. Plant regeneration
via somatic embryogenesis may be of immense commercial interest for developing
automated clonal mass propagation system as it provides almost nil or considerably low
somaclonal variation among regenerates and may be exploited for cryopreservation
(Bajaj, 1983). Inducing and developing callus tissue is also one of the aims of plant
tissue culture experiments. Callus tissues arise as all unorganized proliferation of cells
from segments of plant organs. Callus culture is usually grown on a solid medium.
Callus tissues can be initiated from explants taken from any part like root, hypocotyls,
leaf, stem, embryo, seed, anthers etc. Callus mediated somatic embryos with dependable
growth characteristics and plant regeneration are beneficial for raising improved
varieties through in-vitro selection from high yielding callus lines and possibly also for

42
genetic transformation. Organogenesis is a method of rapid clonal propagation of
genetically uniform plants from callus cells.
Formation of callus tissue is the outcome of cell expansion and cell division of
the cells of the explants. The whole plant can be regenerated in large numbers from
callus tissues through manipulation of the nutrient and hormonal constituents in the
culture media. Callus tissues are a good source of genetic variability, so it may be
possible to regenerate a plant from genetically variable cells. Callus culture is very
useful to obtain commercially important secondary metabolites. Several biochemical
assays can be performed from callus culture.
The agarwood tree (Aquilaria malaccensis Lam. belongs to the family
Thymelaeaceae) is a commercially important and critically endangered forest tree
species of northeast India. The plants are continuously exploited due to its heartwood to
be used in distilling aromatic oil. In addition to overexploitation, high mortality and low
survival rate of Aquilaria seeds are the two major constraints leading to its declined
population in the wild. Naturally, Aquilaria species regenerate through seeds. The seeds
of the plant have physiological limitations such as short dormancy period, rapid loss of
moisture and rapid decline in seed weight which causes poor seed viability prior to
germination. Moreover, the seeds exhibit a very short life of few days to a week only
and rapid loss of seeds moisture during the first few hours to few days leads to rapid
loss of viability; adversely affecting the germination rate. Furthermore, insect attack and
insect pests infestation often inhibits the growth of the seedlings and saplings in the
early phase of establishment (Okugawa et al., 1996). Because of the above facts
coupled with unauthorized overexploitation of this tree for commercial extraction of
agar oil and wood, the population status of this species has become rare in the wild
habitats hence included in International Union for Conservation of Nature (IUCN) red
data list as vulnerable and reported to be at the verge of extinction from the natural
forests (www.iucnredlist.org).
Recently, there was noticeable decrease in the population of A. malaccensis in
natural forests of north east India mainly due to unlimited harvesting of its mature trees
for agarwood trade. As a result, the species has been declared as critically endangered in

43
India from conservation point of view. Therefore, there is an urgent need to develop
suitable regeneration strategies to supplement its natural regeneration and conservation
programmes with sustainable exploitation scheme (Saikia and Khan, 2013). Application
of biotechnological tools and techniques, particularly plant tissue culture in
regeneration, mass propagation and in-vitro conservation of rare and endangered
medicinal and economically important plants have shown a rapid success in the recent
past (Rai, 2010; Verma et al., 2012). In-vitro plant regeneration technique has become a
popular tool and useful method to overcome such problems, if applied skillfully. By
using tissue cultured agarwood plantlets, growers may obtain more oleoresin than by
growing from seeds. The uniform growth rate and enhanced physiological function of
tissue culture raised plantlets may reveal the mechanism of inducing oleoresin
production and predictable volume of oleoresin. Hence, the present research
investigations propose to develop a reproducible and fast protocol(s) for large scale
production of callus of A. malaccensis which could be utilized for organogenesis,
embryogenesis and direct enhanced oil production from this commercially important
tree species of north east India.
4.2. Material and methods
The nodal and leaf explants of A. malaccensis Lam. were taken from young
seedlings of 3-6 month old plants growing in nursery of the Department of Forestry,
NERIST, Arunachal Pradesh. Collected explants were used for cultures immediately
after collection. MS basal medium (Murashige and Skoog, 1962), B5 basal medium
(Gamborg et al., 1968) and Woody Plant Medium (Lloyd and McCown, 1980) with
different hormonal combinations and sucrose concentrations were tested for the present
studies. Auxins play important role in callus induction and multiplication, hence auxin
alone or in combination with cytokinins were tested. The detailed hormonal
combinations and concentrations used for callus initiation and growth in three nutrient
media used in present studies are presented in the Table 4.1, Table 4.2, Table 4.3 &
Table 4.4 respectively. The detailed hormonal combinations with different sucrose
concentrations used for callus growth studies are given in Table 4.9.

44
4.2.1. Incubation conditions
For callus induction, culture vials were incubated at 25º± 2ºC under complete
dark conditions. After callus formation, a set of flasks/tubes were also kept in light
illumination (light intensity ranging from 20-40µE/m2 /S1, fluorescent tubes, Bajaj, 40
Watts) to check the effect of light on callus growth.
4.2.2. Maintenance of callus
Callus initiated in the media were regularly sub-cultured at an interval of 15
days during the whole study period under the same conditions. The initiated calli were
maintained in the fresh culture media with same composition of hormones or with
different treatment of auxin and cytokinin according to the experimental design.
4.2.3. Growth studies of callus
Callus growth studies were carried out to standardize the protocol(s) for its fast
growth rate and production of quality callus mass under optimum conditions. After
initiation, callus was subjected to its growth studies. Growth of callus was monitored in
different sucrose concentrations by determining the fresh and dry weight of the same.
Fresh weight and dry weight of callus that originated from various explants were
measured to the nearest weight in gram.
Under aseptic conditions 400mg of callus was weighted from stock culture,
transferred to the conical flask containing 50ml medium, and then the flasks were kept
in the culture room for incubation in dark. The production percentage of callus biomass
on fresh weight (FW) and dry weight (DW) basis was recorded after the period of 15-
30, 45-60 and 75-90 days respectively. After removing adhered agar and superficial
moisture, the callus tissues were weighted by placing it on pre-weighed aluminum
foil/glass vial. The fresh weight and dry weight of callus were taken after two
subcultures. For determination of dry weight, the callus tissues were kept in a hot air
oven at 60ºC for 6 hours. The weight were recorded and calculated with mean value of
5 replicates.

45
4.2.4. Embryogenic test
4.2.4.1. Preparation of aceto-carmine stain
Callus tissues were studied for the presence of embryos/ embryogenic cells by
aceto-carmine stain test (Sharma and Sharma, 1965). 45% (v/v) acetic acid solution was
prepared and boiled by heating in a water bath. 0.5gm carmine was added slowly to the
boiling solution and mixed gently stirring using glass rod until the dyes dissolved
completely. The solution was cooled down to room temperature, filtered and used for
embryogenic test or stored at 4°C in refrigerator until used. For staining, callus tissues
were transferred to aceto-carmine solution for 30 minutes and then watched under high
magnification microscope.
4.2.4.2. Procedure:
Calli that responded for somatic embryogenesis were extracted from culture and
macerated gently to break down in to small pieces. Small amount of calli were taken on
a clear glass slide and treated with 1-2 drops of aceto-carmine stain by smearing with
the help of a needle. The smear was fixed by warming the slide for 1-2 minutes over the
flame. The slide was then rinsed with warm water for 2-3 times, squashed under a cover
slip by very firm downward pressure and examined under the microscope for
appearance of crimson red embryogenic cells, if any.
4.2.5. Embryogenesis/organogenesis studies of callus tissue
The fresh callus tissues were used and tested for regeneration of shoots. The
calli cells were placed on embryogenic or organogenic callus proliferation media
supplemented with BAP (1-5mg/l) and NAA (0.5-2mg/l) alone or in combination with
NAA (0.1-1mg/l) or IAA (0.1-1mg/l) for regeneration through embryogenesis and
organogenesis (Table 4.11). Twenty replicates were taken for each treatment and media
without hormone were treated as control.
4.2.6. Statistical analysis
Callus induction percentage (Deka, 2002) was calculated for each hormonal
treatment following the formula:

46
Total number of explants with callus induction
Callus induction %= X 100
Total number of explants inoculated for each treatment
The callus growth was recorded on per day basis of callus induction and graded
by comparison to group into four categories as very good (++++), good (+++), poor
(++) and very poor (+). The callus morphology such as color, appearance and texture
etc. were also studied and recorded as per hormonal treatments. The effect of various
growth regulators and time period on callus induction were analyzed statistically using
one way ANOVA and mean differences were tested using Duncan’s multiple range test
at p < 0.05 level.
4.2.7. Microscopy
Microscopic observations were carried out using Semi DV4 stereomicroscope
and ZEISS Axiostar plus compound microscope. Images were taken using ZEISS
Axiostar plus compound microscope and were measured in 40X using the software
ProgRes CapturePro 2.6.
4.3. Results
4.3.1. Comparative study on three different modified media (MS, B5 & WPM)
tested for callus induction of A. malaccensis Lam.
The callus could be induced using callus induction media (MS and WPM & B5)
from both young fresh leaves as well as nodal explants. Hormone(s) played an
important role in callus induction rate with varying results in different hormonal
treatments and combinations. Rapid callus induction was obtained in case of leaf
explants as compare to stem nodal explants irrespective of culture media used or growth
regulators treated. In the present investigations, earliest initiation of callus was recorded
on 12th day using MS medium with leaves as explants. The hormonal treatments MCI11
(MS+NAA 3mg/l + BAP 0.5 mg/l) and MCI13 (MS+2, 4D 2mg/l + kinetin 0.5 mg/l)
(Table 4.1) could induce callus at a faster rate with an average period of 14.66 and
15.77 days respectively (Table 4.5). On the contrary in B5 and WPM, callus induction
took place very late i. e. on 31 and 28th days with the average periods of 33 and 31 days

47
respectively after incubation of all replicates. The grading of callus with ‘good’ (+++)
was observed in treatment number BCI4 (B5+2, 4D 2.5mg/l + BAP 0.5 mg/l) & WCI4
(WPM+NAA 2.5mg/l + BAP 0.5 mg/l) respectively (Table 4.7 & Table 4.8). After
callus induction, callus could be further sub-cultured for 5-6 times after an interval of
every 15 days.
The maximum callus induction percentage (73.33%) was recorded through leaf
explants in the treatment number MCI12 (MS+2, 4D 2mg/l + kinetin 0.1mg/l) followed
by treatments MCI11 and MCI13 with 60% each (Table 4.5). MS medium
supplemented with CW showed slow and poor callus initiation; however showed fast
growth with profuse callus production when it was transferred onto the fresh medium
containing CW.
In case of nodal explants, callus induction took place on 14th day with an
average period of 17.85 days in treatment number MCI12 (MS+2, 4D 2mg/l + kinetin
0.1mg/l) (Table 4.6) using MS medium. The best induction percentage (70 %) was
recorded in hormonal treatment MCI12 followed by MCI13 (MS+2, 4D 4mg/l + kinetin
0.5 mg/l) and MCI10 (MS+NAA 2mg/l + BAP 0.5mg/l) i.e. 50 % each (Table 4.6).
The hormonal combinations MCI12 (MS+ 2, 4D 2mg/l + kinetin 0.1mg/l) may
be considered as most advantageous for callus induction for agarwood species as it
provided highest callus induction percentage (73.33% and 70%; r value = 0.45 and 0.40
respectively at 5% significant level) in both leaf and nodal explants. The minimum
induction percentage were recorded (20%) in MCI25 (MS+2, 4D 8mg/l + Kinetin
3mg/l) and MCI28 (MS+15% CW) (r value= 0.35 at 5% significant level) (Table 4.4)
with leaf explants. In case of nodal explants, the lowest was recorded 30% in treatment
number MCI6 (MS+2, 4D 4mg/l) and MCI9 (MS+NAA 1mg/l + BAP 0.5mg/l) (r value
= 0.36 at 5% significant level) (Table 4.6).
When the observation of callus induction was made for all treatments on 18th
day after inoculation on the culture media, it was found that as maximum as 90% callus
induction could have occurred in MCI11 & MCI13 with that of leaf explants (Fig 4.1
A). The minimum induction percentage of callus for leaf explants was recorded
as11.5% in MCI25 (Fig 4.1 A).

48
In nodal explants, multiple shoots were initiated in MS medium supplemented
with low concentration of auxin (NAA, 0-0.5mg/l) and high concentration of cytokinin
(BAP, 1-2mg/l). Induction of multiple shoots was stopped when the explants were
grown in the same medium with higher cytokinin concentration (≥ 2.5mg/l BAP) with
or without NAA which increased initiation and formation of callus at the base of the
explants.
In B5 medium, maximum callus induction percentage (46.7%) was recorded in
BCI7 (2, 4D 4mg/l + Kn 0.5mg/l) (average of 33.57 days) and least induction
percentage (20%) was recorded in BCI5 (2,4D 1mg/l + Kn 0.5mg/l) (Table 4.7). In
WPM, WCI4 and WCI5 (2, 4D 1mg/l + Kn 0.5mg/l) showed highest callus percentage,
i.e. 50% in 31 and 37.17 average days respectively (Table 4.8). Least induction
percentage (30%) was observed in WCI3 (NAA 1mg/l + BAP 0.5mg/l) (Table 4.8).
A well grown callus could be maintained in all three induction media as well as
in MS basal medium however; induction medium supported long term culturing of
white and friable callus with faster multiplication rate. There was no significant
induction of callus in the MS, B5 and WPM media when 2, 4D, NAA and kinetin were
supplemented individually otherwise initiation started lately with negligible amount of
callus tissue formation for both leaf as well as nodal explants. No explants responded
for callus initiation on basal MS, B5 and WPM media i. e. in control. The callus growth
was graded as ‘very good’ (++++) in four point scale in the treatments MCI11, MCI12
with rapid initiation of callus within 16 and 12 days (average 15.77 & 14.7 days)
respectively in leaf explants and in 14 days (average 17.85 days) in nodal explants in
the treatment MCI12.
The effect of exposure of Aquilaria callus to light and darkness were also
studied simultaneously for its morphology, multiplication and growth. The results
revealed that the callus remained creamish yellow or white in color if maintained in
dark conditions (Plate 4.1 E & G) which on exposure to light (20-40µE/m2 /S1
intensity) developed pigmentation and turned green in color (Plate 4.1 F). Four distinct
types and color of calli were obtained from the explants of A. malaccensis. i. e. White
callus, Yellow white to cream, Blackish white and Yellowish brown colored callus

49
(Plate 4.2). Loose and fragile white callus was obtained when MS or B5 media
supplemented with high concentrations of 2, 4D with low concentrations of Kinetin
(Plate 4.1 D, E & H) were used and placed in dark. It turned green within 2-3 weeks
after transferring to light conditions (Plate 4.1 F). When media supplemented with high
concentrations of NAA and less concentration of BAP, yellow white to cream colored
and compact callus was obtained (Plate 4.1 I). MS medium supplemented with CW
produced blackish white and compact callus (Plate 4.1 C); however, the callus obtained
on WPM was pale yellow or brown in color, compact and globular in nature (Plate 4.1
G). In nodal explants, callus induction was slow and took more time than that of leaf
explants (Plate 4.1 A- B).
4.3.2. Effect of various growth regulators and sucrose concentration on the growth
of callus
The different concentration of sugars also effects callus induction and growth. It
is the most common source of carbohydrate needed in callus induction and regeneration
of plants (Thompson and Thrope, 1987). The maximum callus biomass (7.457gm) was
measured with 4% sucrose concentration using MS medium supplemented with NAA
3mg/l+ BAP 0.5 mg/l after 45-60 days of incubation (Table 4.10; Plate 4.3 A). In
general, the dry cell biomass of callus was found to be proportional to their fresh cell
biomass (Table 4.10). The callus growth declined with increase of time period in all
treatments except in case of hormonal combination 2, 4D 2mg/l + Kn 0.1mg/l, where
slight increase or less decrease in fresh as well as dry cell biomass was noticed.
It was recorded that higher auxin: cytokinin ratio with 3-4% sucrose
concentration in MS medium resulted with better callus biomass production. Among
three ratios (20: 1, 6:1 & 4:1) tested, best result was obtained with 20:1 (2, 4D 2mg/l +
Kn 0.1mg/l) followed by 1:6 (NAA 3mg/l+ BAP 0.5 mg/l) and lowest was recorded in
1:4 (2, 4D 2mg/l + BAP 0.5mg/l).
The maximum fresh callus biomass (7.457gm) and dry callus biomass
(2.302gm) was recorded in the MS medium with hormone combination B at 4%
concentration (MCG7) of sucrose after about 45-60 days of incubation (Table 4.10,

50
Plate 4.3 A). The callus so obtained, could be multiplied at the same rate on sub-
culturing (5-6 times) hence, this protocol may be used for mass callus production. The
lowest callus cell biomass was observed in MCG9 at 2% sucrose concentration with cell
biomass of 0.931gm FW and 0.229gm DW after 45-60 days.
Using hormonal Combination A, the maximum callus biomass was obtained in
MCG3 (7.368gm FW) in 45-60 days old culture with 4% sucrose whereas the lowest
was recorded in MCG1 with 2% sucrose concentration (1.661gm FW). In hormonal
Combination B, the maximum callus biomass (7.457gm) was obtained in 4% sucrose
(MCG7) and the lowest (0.493gm) was observed in 2% sucrose concentration (MCG5)
after 45-60 days of incubation. Callus growth in hormone combination C i.e. 2, 4D
2mg/l + BAP 0.5mg/l (MCG10) depicted that, 3% sucrose concentration supported the
highest callus cell biomass (7.210gm) and the lowest cell biomass was measured in case
of 2% sucrose concentration (MCG9) (0.931gm) after 45-60 days of incubation. In 75-
90 days incubation time, it was interesting to note that there was decrease in fresh cell
biomass however, the dry cell biomass increased. The older cultures appeared dull and
darken in color (Plate 4.3 B).
Sucrose had various effects on callus initiation and growth. The callus growth
was recorded faster in case of media treated with 4% sucrose irrespective of hormonal
treatments after 45-60 days of incubation. The biomass of callus was universally
proportional to the concentration of sucrose up to 4% and declined beyond it. From the
present investigations, it was revealed that prolonged culturing period of callus has
direct impact on its growth as weight of the callus declined sharply after 60 days of
incubation. Therefore, the period of 45-60 days was optimized as ideal period for mass
callus production.
In all the treatments, 2% sucrose showed least growth (0.931gm) as compared to
other sucrose concentrations. In 15-30 days old callus, the highest growth was recorded
in hormonal Combination B (MCG7) (2.390gm FW) followed by Combination A
(MCG3) (1.281gm FW) and the lowest growth (0.259gm FW) in 4% sucrose
concentration was observed in Combination C (MCG12) in 5% sucrose. In 75-90 days
old culture, callus weight was declined in all treatments except in hormone Combination

51
A with 3% sucrose and Combination B with 2% sucrose where very little amount of
callus amount increased. At 5 % sucrose concentration, reduced growth of callus was
recorded in all cases. The highest growth was noted in the hormonal Combination A.
Callus tissues of A. malaccensis grown and maintained under dark condition remained
whitish or creamish yellow in color up to approximately 60 days. They usually turned
green when transferred to light illuminations (intensity 20-40µE/m2 /S1) due to
development of chloroplastids.
4.3.3. Embryogenic test
Aceto-carmine staining of young and matured callus tissues revealed the
presence of embryogenic cells which appeared crimson red in colour (Plate 4.4 A-F);
however, non-embryogenic cells appeared unstained, distorted and irregular in shape or
with lighter shade due to lack of protoplasm (Plate 4.4 G & H). Squash preparation of
the callus tissues of A. malaccensis revealed heterogeneous cellular composition
ranging from small rounded cells with dense cytoplasm to large irregular shaped cells
with vacuolated cytoplasm in case of young or friable callus. The friable callus cells
were loose, with entire margin, full of protoplasm and mostly globular in shape with
average diameter of 31.5µm. The other shapes like oval, slightly elongated, beaked or
comma shaped ranging between 28-45 µm in length and 18-35µm in breath were also
observed.
4.3.4. Effect of various growth regulators on embryogenic and organogenic callus
tissue development of A. malaccensis
The callus initiated in the modified culture media were aseptically transferred to
different embryogenic/organogenic callus proliferation media (EAM) supplemented
with varied concentration of growth regulators alone or in combination. Pre-
embryogenic calli developed in MS media treatments EAM4 (3mg/l Activated Charcoal
+ 2mg/l BAP), EAM11 (2mg/l BAP) and in EAM14 (2mg/l BAP + 0.3mg/l NAA).
Embryogenic/organogenic callus were observed in all the treatments (Plate 4.7 A-C) of
MS medium except in EAM2 (0.5 mg/l BAP & 2,4D each + 3mg/l Activated charcoal).
This is for the first time ever in A. malaccensis plant; organogenesis and somatic

52
embryogenesis have been reported. Opposite to our expectation, somatic embryo
formation was also took place in control i.e. MS basal medium without hormone
(EAM1) (Plate 4.5 F) and in treatment number EAM8 (BAP 0.5mg/l + IAA 0.3mg/l) in
65 and 43 days of incubation respectively but they did not regenerate shoots. Among the
media used, MS medium supplemented with 0.75mg/l BAP + 0.3mg/l IAA (EAM9) and
1mg/l BAP + 0.3mg/l NAA (EAM13) could be able to induce caulogenesis or shoot
formation (Plate 4.5A-E). Some shoots were seemed to be stressed and ultimately died
but others survived and eventually regenerated into healthy shoots. Aquilaria required
approximately 6months time for multiplication of shoots from callus as 3-4 months time
required for shoot induction and two more months for shoot elongation. Rhizogenesis
was observed with highest rooting percentage (90%) when callus cells were grown in
EAM6 (half MS medium supplemented with 1mg/l NAA) followed by 85% rooting in
EAM7 (2mg/l NAA) and 50% in EAM5 (0.5mg/l NAA) but further shoot formation
was not observed (Plate 4.6 B). B5 medium, embryogenic callus development were
observed in treatment numbers EAM19 & EAM20 (Plate 4.7 D) and only rhizogenesis
was observed in EAM21, EAM22 & EAM23 treatments when supplemented with the
combination of 1-3mg/l 2, 4D and 0.1mg/l kinetin (Plate 4.6 A). In WPM, only green
callus were formed in EAM25, EAM26 & EAM27 (Plate 4.7 E & F) but rhizogenesis
or caulogenesis could not be induced.
4.4. Discussion
The organogenic and embryogenic potentials of woody plants are strongly
affected by the maturity phase. As tree grow and attain maturity, the ability of
vegetative tissues to differentiate plant organs declines. In addition, explants derived
from mature trees can often cause difficulties in establishing in vitro cultures and inhibit
proliferation of axillary buds by phenolic exudations into the medium (Trigano et al.,
1992). The experiment with micropropagation of Acer caudatifolium showed response
to 2-year old seedlings (Durkovic, 2003) but not in 5 year old seedling (Durkovic and
Misalova, 2008). Depending on the plant species, only a limited number of cells in an
explant show the organogenic response (Declerk et al., 1997). Therefore juvenile

53
explants from nodal and leaves of A. malaccensis were used for callus induction. The
leaf explants showed best callus induction in MS medium. In-vitro plant regeneration
from stem and leaf calluses of Aquilaria sinensis was carried out by Ye et al. (1998).
The induction and growth characteristics of callus of A. malaccensis were also studied
earlier using leaves, shoot tips and stems as explants and best results were obtained
from leaves (Talukdar and Ahmed, 2001). This supports the present findings about
callus induction. Combinations of 2, 4D and Kinetin for initiation of callus in some
woody plant species viz. Phoenix dactylifera, Sesamum indicum and Ceratozamia
hildae were established earlier by several workers (Omar and Novak, 1990; Rao and
Vaidyanath, 1997). MS medium supplemented with lower levels of 2, 4D (2mg/l) and
Kinetin (0.5mg/l) was recorded to initiate the callus in Havea brasiliensis (Jayashree et
al., 1999). Our studies report fast callus induction and high multiplication rate in
Aquilaria species at very low hormonal (MS + 2,4D 2mg/l + Kinetin 0.1mg/l) and
sucrose (3%) concentrations. Enhanced rate of callusing in Aquilaria species has been
reported earlier using various carbohydrate sources such as fructose, maltose, sucrose
etc. at their different concentrations (Talukdar and Ahmed, 2001). They have studied
induction and growth characteristics of callus of A. malaccensis using leaves, stems and
shoot tips as explants where best results were obtained from leaves. The effect of varied
sucrose concentrations on callus induction was also investigated in A. malaccensis Lam.
using leaf explants (Talukdar and Ahmed, 2004) and maximum callus biomass was
obtained in MS medium supplemented with NAA 3mg/l + BAP 0.5 mg/l at 4% sucrose
concentration. In vitro response, growth and maintenance of callus of A. malaccensis
was examined recently by Debnath, (2013) who resulted that only 2-4 mg/l 2, 4D in MS
medium induced callus. CW is known to serve as a source of natural growth regulator
influencing in-vitro growth of plant tissues (George, 1993); however, in the present
studies, CW did not influence early initiation of callus but it had enhanced further callus
multiplication. No clarification could be made for this. The maximum callus was
obtained only after 30-45 days when comparatively higher amount of growth regulators
and sucrose (4%) were used in MS medium + 2, 4D, 6mg/l + Kinetin, 2mg/l
combination (Talukdar and Ahmed, 2001). Similarly, highest growth of Aquilaria

54
agallocha callus was obtained using fructose as source of carbohydrate as a treatment
constant through leaf explants (Talukdar and Ahmed, 2004). It was reported that 30gm/l
fructose concentration resulted with highest growth rate (547%) of fresh callus biomass
as compared to maltose (60 g/l) which showed least growth rate (212%) after 45 days.
In contrast to these findings, the results of present investigation were very encouraging
since early induction and rapid proliferation of calli could be achieved at comparatively
low concentrations of growth hormones. The callus growth was graded as ‘very good’
(++++) in four point scale in two treatments (MSI11 & MSI12). In these treatments,
fastest callus initiation was recorded within 12 - 16 days in leaf and within 14 days in
nodal explants. B5 and WPM were found not suitable for callus induction and growth of
A. malaccensis as compare to MS medium. This may be due to different composition
and quantity of compounds present in B5 and WPM unable to fulfill physiological
needs and provide favorable cellular metabolic environment.
No notable callus was observed or very less amount of callus initiated lately in
all three media when supplemented with only auxins in both leaf and nodal explants of
A. malaccensis. All three media without growth hormones did not form callus.
Therefore, it can be concluded that, for callus formation of A. malaccensis, growth
hormones with combination of both auxin and cytokinin are the essential requirements.
The light and dark period showed effects on callus morphology. Four distinct
types and color of calli i. e. White callus, Yellow white to cream, Blackish white and
Yellowish brown were obtained from the explants of A. malaccensis. The texture and
type of callus depended on the type of growth regulators and explants used. Similar
types of callus initiation have been reported on A. malaccensis by earlier workers also
((Talukdar and Ahmed, 2001) and confirm our findings.
The effect of sucrose concentrations (2-5%) on callus induction was also studied
and recorded. MS medium supplemented with BAP 0.5 mg/l + NAA 3mg/l with 4%
sucrose concentration was recorded with maximum callus biomass (7.457gm) after 45-
60 days of incubation. Overall, 3%-4% sucrose concentration with higher
auxin:cytokinin ratio may be considered most suitable and may be utilized for mass
callus biomass production. The dry cell biomass of callus was found to be proportional

55
to their fresh cell biomass except in older callus. The biomass of callus was universally
proportional to the concentration of sucrose up to 4% and declined beyond it. The
results of present studies indicate that the optimum incubation period required is about
45-60 days in contrast to Talukdar and Ahmed (2001; 2004) who obtained maximum
callus after 45 days; however they used quite higher amount of growth hormones (2, 4D
6mg/l + Kn 2mg/l) at 4% sucrose concentration which support our findings. From the
present experiment, it can be concluded that after 45-60 days of incubation, hormonal
combination of NAA 3mg/l + BAP 0.5mg/l and 4% sucrose level were best for rapid
callus growth in A. malaccensis. Overall, 2, 4D 2mg/l + Kn 0.1mg/l i.e. Combination A
(MCG1-MCG4) was found more suitable in terms of callus biomass at 3-5% sucrose
concentrations as compare to others. The prolonged culturing period of callus had direct
impact on its growth as weight of the callus declined sharply after 60 days of
incubation. The fresh weight and dry weight ratio considerably decreased when
incubation period was prolonged (75-90 days) and color of callus turned darker and
blackish. This may be due to the fact that lignifications in callus tissues might have
taken place as a result of cyto-differentiation in older callus tissues and made them
harder. It was also evident by the changes in the color and appearance of 75-90 days old
callus which looked dull and darker. Callus morphology of A. malaccensis was studied
earlier by Talukdar and Ahmed (2004) and recorded that, after 45 days callus cells were
darkened due to onset of senescence. Only two types of callus i.e. green callus and fast
growing white callus were reported by them.
Embryogenic callus were distinguished easily by acetocarmine staining method.
The embryogenic cells were appeared crimson red in colour however non-embryogenic
cells appeared unstained. Identification of embryogenic callus by acetocarmine stain
was also used in Gerbera jamesonii (Hasbullah et al., 2007) and Onobrychis sativa
(Mohajer et al., 2012) plants. Embryogenic/organigenic callus were observed in all
treatments of MS medium but treatment EAM2 neither responded to embryogenesis nor
to organogenesis. Somatic embryos were also formed in some treatments but they could
not regenerate shoots. This may be due to imbalance of nutrition in the culture medium
which has blocked the further growth.

56
Although several investigations have compared the embryonic or organogenic
potential of calli produced in different culture media, there has not been a general
standard for all the vegetal species. For example, evaluating different nutritive medium
in calli cultures from leaf segments of Morus indica L., it was found that the MS media
does not induce caulogenesis (Sahoo et al., 1997); Nevertheless, in studies with adult
leaves of Paulownia elongate, a higher proportion of callus with morphogenic response
in the MS media than in the WPM media (Castellanos et al., 2006) was obtained.
Organogenesis from stem and leaf calluses of Aquilaria sinensis was successful in MS
medium (Ye et al., 1998).
There are differences in the organogenic potential between plant families,
genera, species as well as genotypes, and different genotypes of a species may show
widely different responses (George, 1996). For example, Capsicum species that belong
to the Solanaceae have been shown to be recalcitrant to differentiation and plant
regeneration under in-vitro conditions compared to Nicotiana tabacum, Lycopersicum
esculentum, and Solanum tuberosum, which belong to the same family (Ochoa-Alejo
and Ramirez- Malagon, 2001). Organogenesis from A. malaccensis callus tissue was not
successful in earlier studies (Tiengtum, 1995). The earlier studies on growth
characteristics of callus tissue of A. malaccensis also showed that callus tissue did not
regenerate into whole plant (Talukdar and Ahmed, 2001). Rhizogenesis in callus tissue of
A. malaccensis was observed in the present investigations but caulogenesis in the same
sample did not take place. Organogenic connection between shoot and root primordia is
essential for the regeneration of complete plantlets from the same culture. Shoot
formation followed by rooting is the general characteristics of organogenesis. During
organogenesis, if the roots are first formed, then it is very difficult to induce shoot bud
formation from the same callus tissue (De, 1992). This was confirmed by our findings.
MS medium supplemented with 0.75mg/l BAP + 0.3mg/l IAA (EAM9) and 1mg/l BAP
+ 0.3mg/l NAA could be able to form shoots from callus tissue. This is the first report
on the regeneration of A. malaccensis from callus tissue through organogenesis
(rhizogenesis & caulogenesis) from leaf and nodal explants; however, more elaborated
and specific studies may be required to induce organogenesis in callus of A.

57
malaccensis at a faster rate. Our experimental results may provide specific insight to
potential investigators to design their research. This would also help in conservation of
valuable species for future. Direct and successful extraction of the agar oil has been
achieved from calli of A. malaccensis (Talukdar et al., 2002). Evaluation of few
biochemical parameters of the in-vitro grown callus tissue of A. malaccensis was also
studied by Talukdar (2012). The important biochemical constituents such as
sesquiterpenoids may be synthesized in living cells using calli and chromone derivatives
which are the major components of agarwood may be produced from debris of dying
cells of callus (Okudera and Ito, 2009).
Therefore, such large scale and rapid callus production protocols could be
employed for in-vitro biosynthesis of important biochemical principles in laboratory
conditions. This may help in reducing uncontrolled extraction and exploitation of this
threatened tree species for commercial purposes while conserving in the wild natural
habitats of region in particular and other habitats in general in the near future.
By these findings, it may be accomplished that the growth and morphogenesis of
plant tissue culture in vitro are largely governed by the composition of the culture
medium. Media composition is therefore formulated considering specific requirements
of particular plant species. A standard and reproducible tissue culture protocol for rapid
induction of callus has been developed using leaf and nodal explants of A. malaccensis
Lam. Use of young fresh leaves on MS medium supplemented with low concentration
of auxin and cytokinin was found suitable for rapid callus initiation in A. malaccensis as
compared to nodal explants and WPM or B5 as alternate source of culture medium.
Overall, the hormonal combination of 2, 4D 2mg/l + kinetin 0.1mg/l in MS medium
with 3% sucrose concentration may be recommended for rapid callus induction and
production of good callus biomass in A. malaccensis. The callus biomass may be
maximized by increasing sucrose concentration (4%) with MS medium supplemented
with BAP (0.5 mg/l) + NAA (3mg/l). The same could be utilized for direct agar oil
extraction from this commercially important but critically endangered tree species. Four
morphologically different types of callus were observed as influenced by different
hormonal combinations and light conditions. Another interesting area of research may

58
be to investigate effect of introduction of endophytic fungi on agar oil quality and
quantity analysis. Organogenesis from callus tissues were achieved in MS medium but
further optimization is needed to increase the efficiency and frequency of regeneration
and to reduce the regeneration period.
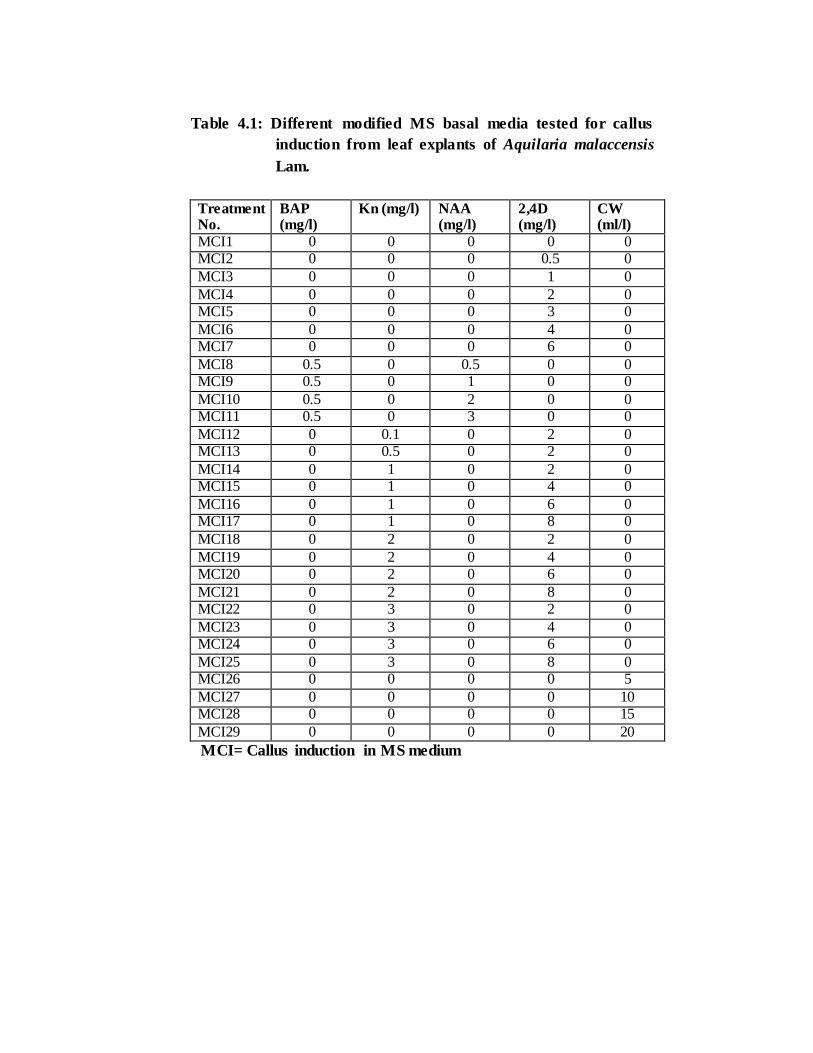
Table 4.1: Different modified MS basal media tested for callus
induction from leaf explants of Aquilaria malaccensis
Lam.
Treatment No.
BAP (mg/l)
Kn (mg/l) NAA (mg/l)
2,4D (mg/l)
CW (ml/l)
MCI1 0 0 0 0 0 MCI2 0 0 0 0.5 0
MCI3 0 0 0 1 0
MCI4 0 0 0 2 0 MCI5 0 0 0 3 0
MCI6 0 0 0 4 0 MCI7 0 0 0 6 0
MCI8 0.5 0 0.5 0 0 MCI9 0.5 0 1 0 0
MCI10 0.5 0 2 0 0 MCI11 0.5 0 3 0 0
MCI12 0 0.1 0 2 0 MCI13 0 0.5 0 2 0
MCI14 0 1 0 2 0 MCI15 0 1 0 4 0
MCI16 0 1 0 6 0 MCI17 0 1 0 8 0
MCI18 0 2 0 2 0
MCI19 0 2 0 4 0 MCI20 0 2 0 6 0
MCI21 0 2 0 8 0 MCI22 0 3 0 2 0
MCI23 0 3 0 4 0 MCI24 0 3 0 6 0
MCI25 0 3 0 8 0 MCI26 0 0 0 0 5
MCI27 0 0 0 0 10 MCI28 0 0 0 0 15
MCI29 0 0 0 0 20
MCI= Callus induction in MS medium
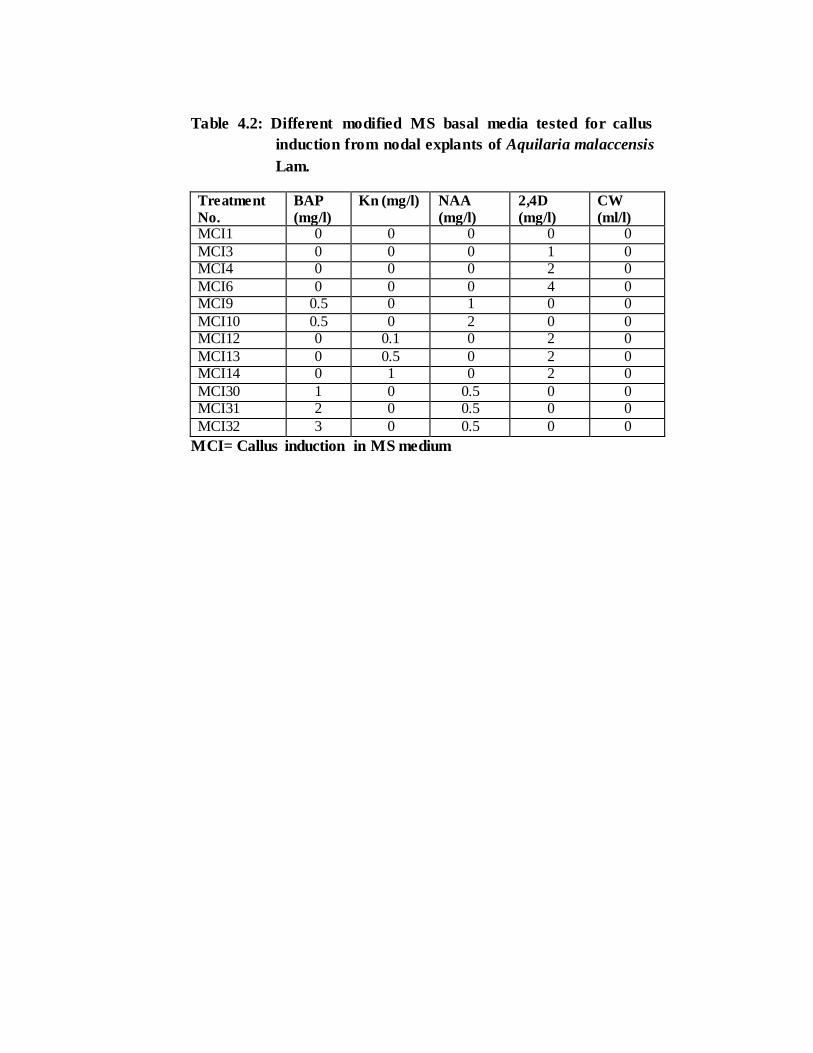
Table 4.2: Different modified MS basal media tested for callus
induction from nodal explants of Aquilaria malaccensis
Lam.
Treatment
No.
BAP
(mg/l)
Kn (mg/l) NAA
(mg/l)
2,4D
(mg/l)
CW
(ml/l) MCI1 0 0 0 0 0
MCI3 0 0 0 1 0 MCI4 0 0 0 2 0
MCI6 0 0 0 4 0 MCI9 0.5 0 1 0 0
MCI10 0.5 0 2 0 0 MCI12 0 0.1 0 2 0
MCI13 0 0.5 0 2 0 MCI14 0 1 0 2 0
MCI30 1 0 0.5 0 0 MCI31 2 0 0.5 0 0
MCI32 3 0 0.5 0 0
MCI= Callus induction in MS medium
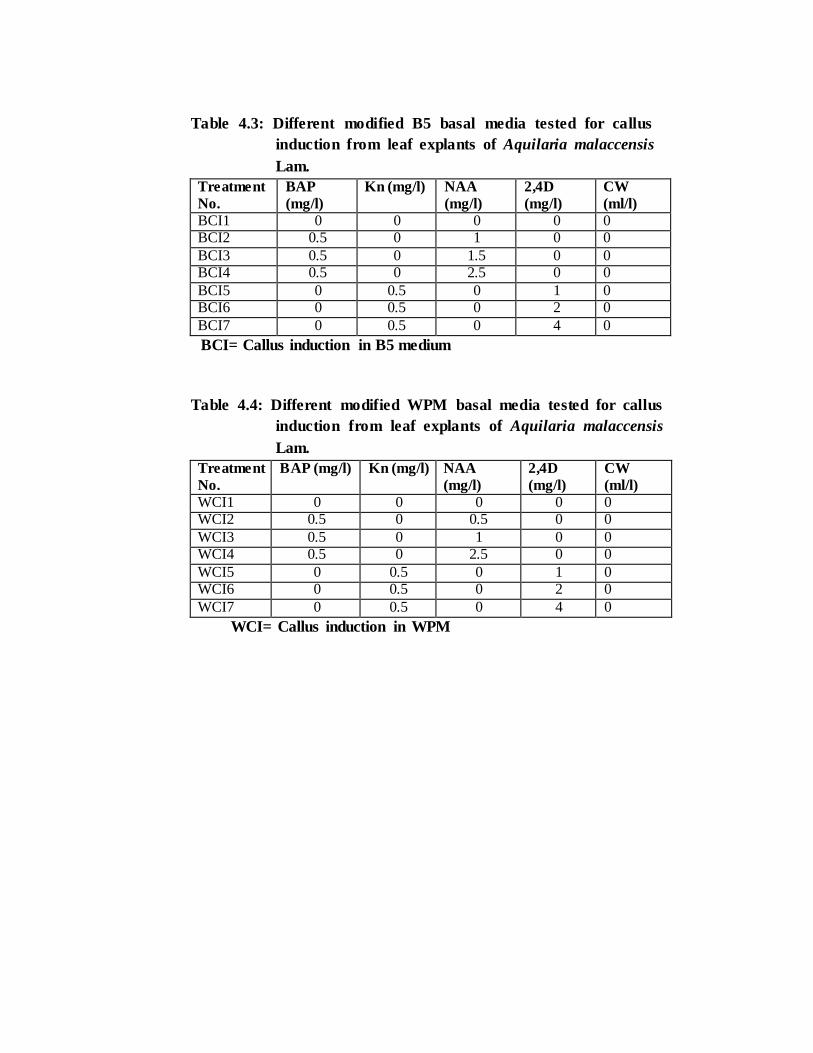
Table 4.3: Different modified B5 basal media tested for callus
induction from leaf explants of Aquilaria malaccensis
Lam.
Treatment
No.
BAP
(mg/l)
Kn (mg/l) NAA
(mg/l)
2,4D
(mg/l)
CW
(ml/l)
BCI1 0 0 0 0 0 BCI2 0.5 0 1 0 0
BCI3 0.5 0 1.5 0 0 BCI4 0.5 0 2.5 0 0
BCI5 0 0.5 0 1 0 BCI6 0 0.5 0 2 0
BCI7 0 0.5 0 4 0
BCI= Callus induction in B5 medium
Table 4.4: Different modified WPM basal media tested for callus
induction from leaf explants of Aquilaria malaccensis
Lam.
Treatment
No.
BAP (mg/l) Kn (mg/l) NAA
(mg/l)
2,4D
(mg/l)
CW
(ml/l)
WCI1 0 0 0 0 0 WCI2 0.5 0 0.5 0 0
WCI3 0.5 0 1 0 0 WCI4 0.5 0 2.5 0 0
WCI5 0 0.5 0 1 0 WCI6 0 0.5 0 2 0
WCI7 0 0.5 0 4 0
WCI= Callus induction in WPM
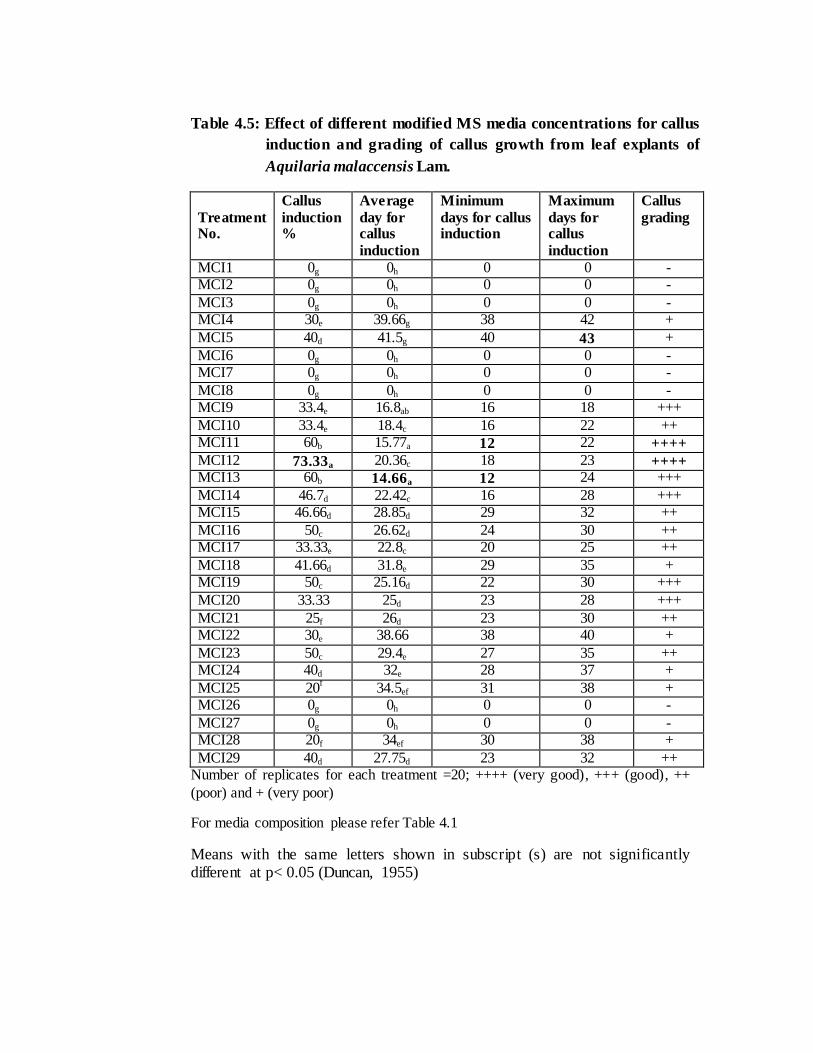
Table 4.5: Effect of different modified MS media concentrations for callus
induction and grading of callus growth from leaf explants of
Aquilaria malaccensis Lam.
Treatment No.
Callus
induction %
Average
day for callus
induction
Minimum
days for callus induction
Maximum
days for callus
induction
Callus
grading
MCI1 0g 0h 0 0 - MCI2 0g 0h 0 0 -
MCI3 0g 0h 0 0 - MCI4 30e 39.66g 38 42 +
MCI5 40d 41.5g 40 43 +
MCI6 0g 0h 0 0 - MCI7 0g 0h 0 0 -
MCI8 0g 0h 0 0 - MCI9 33.4e 16.8ab 16 18 +++
MCI10 33.4e 18.4c 16 22 ++ MCI11 60b 15.77a 12 22 ++++
MCI12 73.33a 20.36c 18 23 ++++ MCI13 60b 14.66a 12 24 +++
MCI14 46.7d 22.42c 16 28 +++ MCI15 46.66d 28.85d 29 32 ++
MCI16 50c 26.62d 24 30 ++ MCI17 33.33e 22.8c 20 25 ++
MCI18 41.66d 31.8e 29 35 + MCI19 50c 25.16d 22 30 +++
MCI20 33.33 25d 23 28 +++
MCI21 25f 26d 23 30 ++ MCI22 30e 38.66 38 40 +
MCI23 50c 29.4e 27 35 ++ MCI24 40d 32e 28 37 +
MCI25 20f
34.5ef 31 38 + MCI26 0g 0h 0 0 -
MCI27 0g 0h 0 0 - MCI28 20f 34ef 30 38 +
MCI29 40d 27.75d 23 32 ++ Number of replicates for each treatment =20; ++++ (very good), +++ (good), ++
(poor) and + (very poor)
For media composition please refer Table 4.1
Means with the same letters shown in subscript (s) are not significantly different at p< 0.05 (Duncan, 1955)
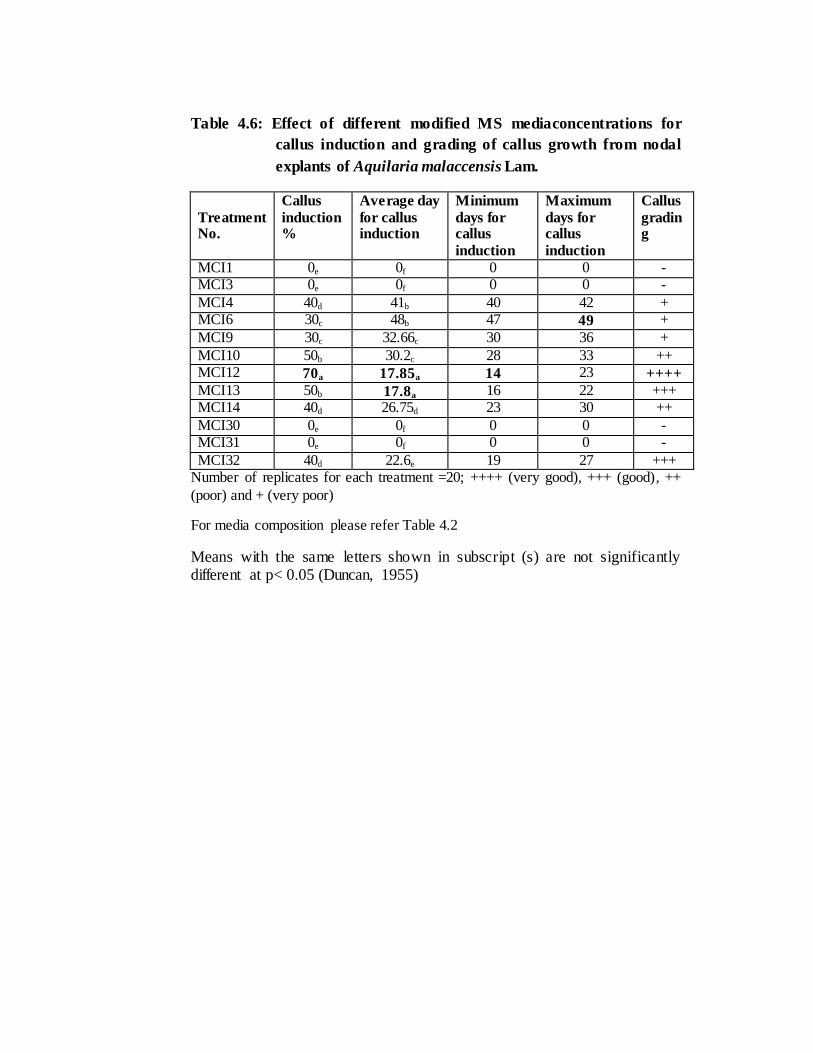
Table 4.6: Effect of different modified MS mediaconcentrations for
callus induction and grading of callus growth from nodal
explants of Aquilaria malaccensis Lam.
Treatment No.
Callus
induction %
Average day
for callus induction
Minimum
days for callus
induction
Maximum
days for callus
induction
Callus
grading
MCI1 0e 0f 0 0 - MCI3 0e 0f 0 0 -
MCI4 40d 41b 40 42 + MCI6 30c 48b 47 49 +
MCI9 30c 32.66c 30 36 +
MCI10 50b 30.2c 28 33 ++ MCI12 70a 17.85a 14 23 ++++
MCI13 50b 17.8a 16 22 +++ MCI14 40d 26.75d 23 30 ++
MCI30 0e 0f 0 0 - MCI31 0e 0f 0 0 -
MCI32 40d 22.6e 19 27 +++ Number of replicates for each treatment =20; ++++ (very good), +++ (good), ++
(poor) and + (very poor)
For media composition please refer Table 4.2
Means with the same letters shown in subscript (s) are not significantly different at p< 0.05 (Duncan, 1955)
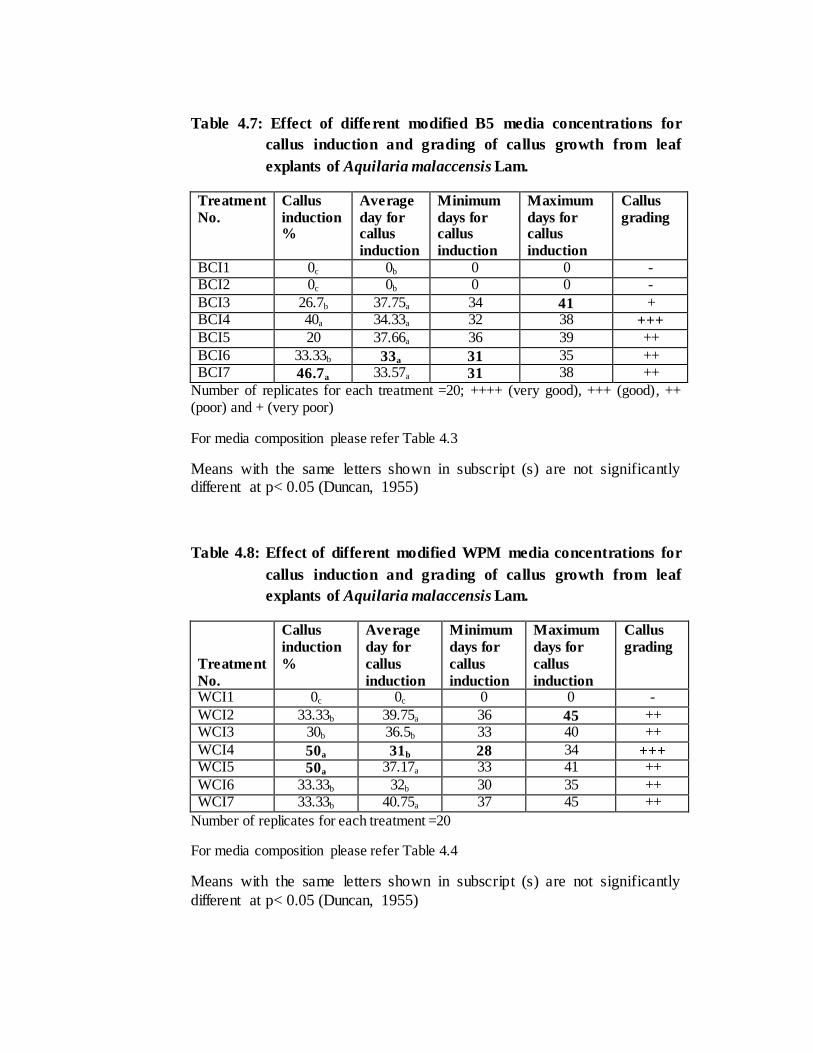
Table 4.7: Effect of different modified B5 media concentrations for
callus induction and grading of callus growth from leaf
explants of Aquilaria malaccensis Lam.
Treatment
No.
Callus
induction %
Average
day for callus
induction
Minimum
days for callus
induction
Maximum
days for callus
induction
Callus
grading
BCI1 0c 0b 0 0 - BCI2 0c 0b 0 0 -
BCI3 26.7b 37.75a 34 41 + BCI4 40a 34.33a 32 38 +++
BCI5 20 37.66a 36 39 ++
BCI6 33.33b 33a 31 35 ++ BCI7 46.7a 33.57a 31 38 ++
Number of replicates for each treatment =20; ++++ (very good), +++ (good), ++ (poor) and + (very poor)
For media composition please refer Table 4.3
Means with the same letters shown in subscript (s) are not significantly different at p< 0.05 (Duncan, 1955)
Table 4.8: Effect of different modified WPM media concentrations for
callus induction and grading of callus growth from leaf
explants of Aquilaria malaccensis Lam.
Treatment
No.
Callus
induction
%
Average
day for
callus
induction
Minimum
days for
callus
induction
Maximum
days for
callus
induction
Callus
grading
WCI1 0c 0c 0 0 -
WCI2 33.33b 39.75a 36 45 ++ WCI3 30b 36.5b 33 40 ++
WCI4 50a 31b 28 34 +++ WCI5 50a 37.17a 33 41 ++
WCI6 33.33b 32b 30 35 ++ WCI7 33.33b 40.75a 37 45 ++
Number of replicates for each treatment =20
For media composition please refer Table 4.4
Means with the same letters shown in subscript (s) are not significantly
different at p< 0.05 (Duncan, 1955)
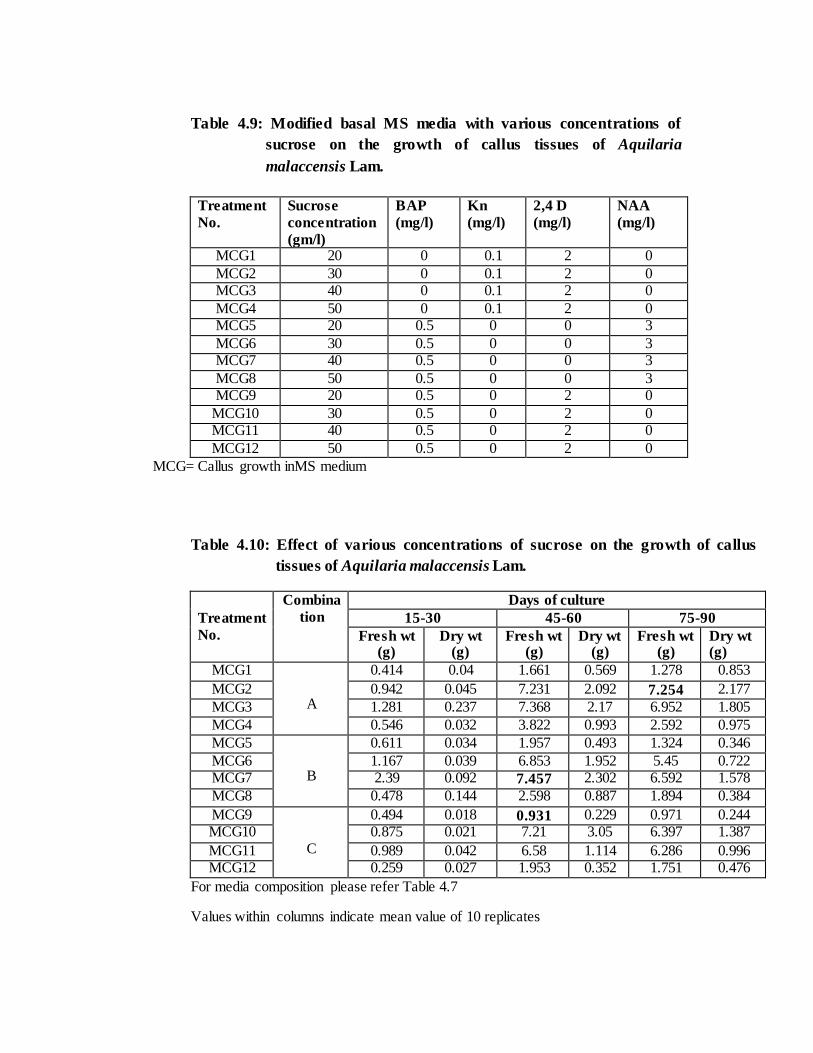
Table 4.9: Modified basal MS media with various concentrations of
sucrose on the growth of callus tissues of Aquilaria
malaccensis Lam.
Treatment
No.
Sucrose
concentration
(gm/l)
BAP
(mg/l)
Kn
(mg/l)
2,4 D
(mg/l)
NAA
(mg/l)
MCG1 20 0 0.1 2 0
MCG2 30 0 0.1 2 0 MCG3 40 0 0.1 2 0
MCG4 50 0 0.1 2 0 MCG5 20 0.5 0 0 3
MCG6 30 0.5 0 0 3 MCG7 40 0.5 0 0 3
MCG8 50 0.5 0 0 3 MCG9 20 0.5 0 2 0
MCG10 30 0.5 0 2 0 MCG11 40 0.5 0 2 0
MCG12 50 0.5 0 2 0
MCG= Callus growth inMS medium
Table 4.10: Effect of various concentrations of sucrose on the growth of callus
tissues of Aquilaria malaccensis Lam.
Treatment
No.
Combina
tion
Days of culture
15-30 45-60 75-90
Fresh wt (g)
Dry wt (g)
Fresh wt (g)
Dry wt (g)
Fresh wt (g)
Dry wt (g)
MCG1
A
0.414 0.04 1.661 0.569 1.278 0.853
MCG2 0.942 0.045 7.231 2.092 7.254 2.177
MCG3 1.281 0.237 7.368 2.17 6.952 1.805
MCG4 0.546 0.032 3.822 0.993 2.592 0.975
MCG5
B
0.611 0.034 1.957 0.493 1.324 0.346
MCG6 1.167 0.039 6.853 1.952 5.45 0.722 MCG7 2.39 0.092 7.457 2.302 6.592 1.578
MCG8 0.478 0.144 2.598 0.887 1.894 0.384
MCG9
C
0.494 0.018 0.931 0.229 0.971 0.244 MCG10 0.875 0.021 7.21 3.05 6.397 1.387
MCG11 0.989 0.042 6.58 1.114 6.286 0.996 MCG12 0.259 0.027 1.953 0.352 1.751 0.476
For media composition please refer Table 4.7
Values within columns indicate mean value of 10 replicates
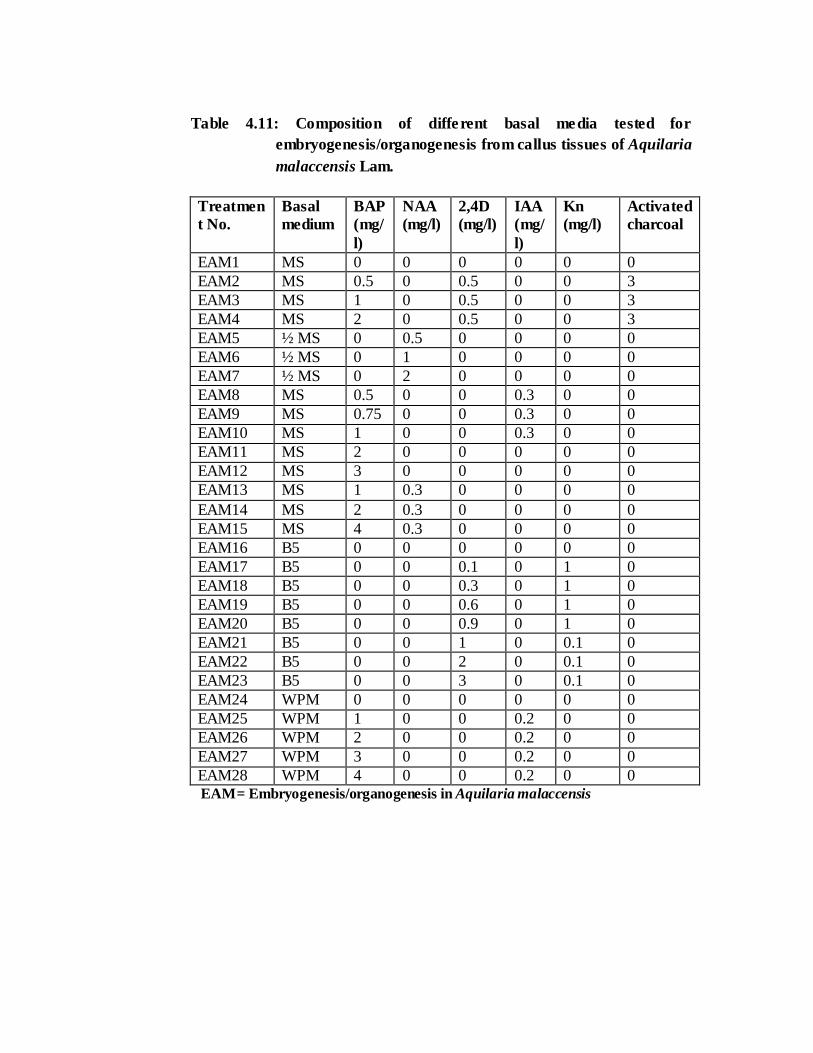
Table 4.11: Composition of different basal media tested for
embryogenesis/organogenesis from callus tissues of Aquilaria
malaccensis Lam.
Treatmen
t No.
Basal
medium
BAP
(mg/
l)
NAA
(mg/l)
2,4D
(mg/l)
IAA
(mg/
l)
Kn
(mg/l)
Activated
charcoal
EAM1 MS 0 0 0 0 0 0
EAM2 MS 0.5 0 0.5 0 0 3
EAM3 MS 1 0 0.5 0 0 3
EAM4 MS 2 0 0.5 0 0 3
EAM5 ½ MS 0 0.5 0 0 0 0
EAM6 ½ MS 0 1 0 0 0 0
EAM7 ½ MS 0 2 0 0 0 0
EAM8 MS 0.5 0 0 0.3 0 0
EAM9 MS 0.75 0 0 0.3 0 0
EAM10 MS 1 0 0 0.3 0 0
EAM11 MS 2 0 0 0 0 0
EAM12 MS 3 0 0 0 0 0
EAM13 MS 1 0.3 0 0 0 0
EAM14 MS 2 0.3 0 0 0 0
EAM15 MS 4 0.3 0 0 0 0
EAM16 B5 0 0 0 0 0 0
EAM17 B5 0 0 0.1 0 1 0
EAM18 B5 0 0 0.3 0 1 0
EAM19 B5 0 0 0.6 0 1 0
EAM20 B5 0 0 0.9 0 1 0
EAM21 B5 0 0 1 0 0.1 0
EAM22 B5 0 0 2 0 0.1 0
EAM23 B5 0 0 3 0 0.1 0
EAM24 WPM 0 0 0 0 0 0
EAM25 WPM 1 0 0 0.2 0 0
EAM26 WPM 2 0 0 0.2 0 0
EAM27 WPM 3 0 0 0.2 0 0
EAM28 WPM 4 0 0 0.2 0 0 EAM= Embryogenesis/organogenesis in Aquilaria malaccensis
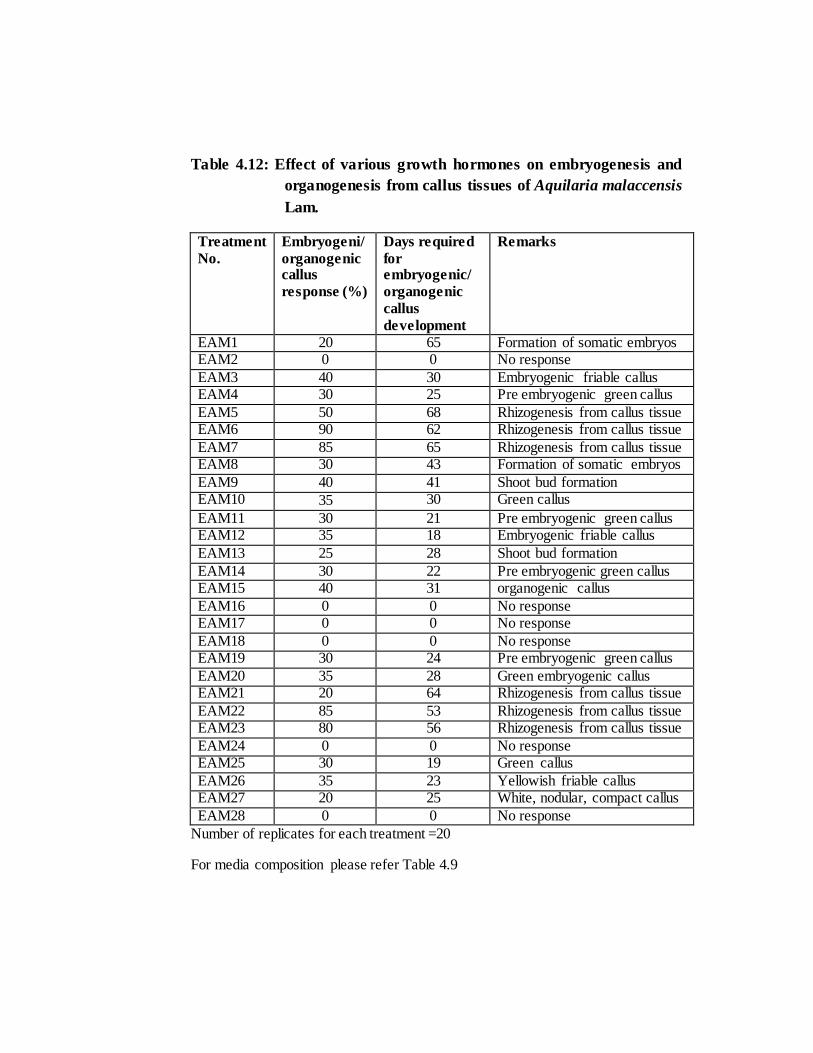
Table 4.12: Effect of various growth hormones on embryogenesis and
organogenesis from callus tissues of Aquilaria malaccensis
Lam.
Treatment
No.
Embryogeni/
organogenic callus
response (%)
Days required
for embryogenic/
organogenic
callus
development
Remarks
EAM1 20 65 Formation of somatic embryos EAM2 0 0 No response
EAM3 40 30 Embryogenic friable callus EAM4 30 25 Pre embryogenic green callus
EAM5 50 68 Rhizogenesis from callus tissue EAM6 90 62 Rhizogenesis from callus tissue
EAM7 85 65 Rhizogenesis from callus tissue EAM8 30 43 Formation of somatic embryos
EAM9 40 41 Shoot bud formation EAM10 35 30 Green callus
EAM11 30 21 Pre embryogenic green callus EAM12 35 18 Embryogenic friable callus
EAM13 25 28 Shoot bud formation
EAM14 30 22 Pre embryogenic green callus EAM15 40 31 organogenic callus
EAM16 0 0 No response EAM17 0 0 No response
EAM18 0 0 No response EAM19 30 24 Pre embryogenic green callus
EAM20 35 28 Green embryogenic callus EAM21 20 64 Rhizogenesis from callus tissue
EAM22 85 53 Rhizogenesis from callus tissue EAM23 80 56 Rhizogenesis from callus tissue
EAM24 0 0 No response EAM25 30 19 Green callus
EAM26 35 23 Yellowish friable callus EAM27 20 25 White, nodular, compact callus
EAM28 0 0 No response
Number of replicates for each treatment =20
For media composition please refer Table 4.9
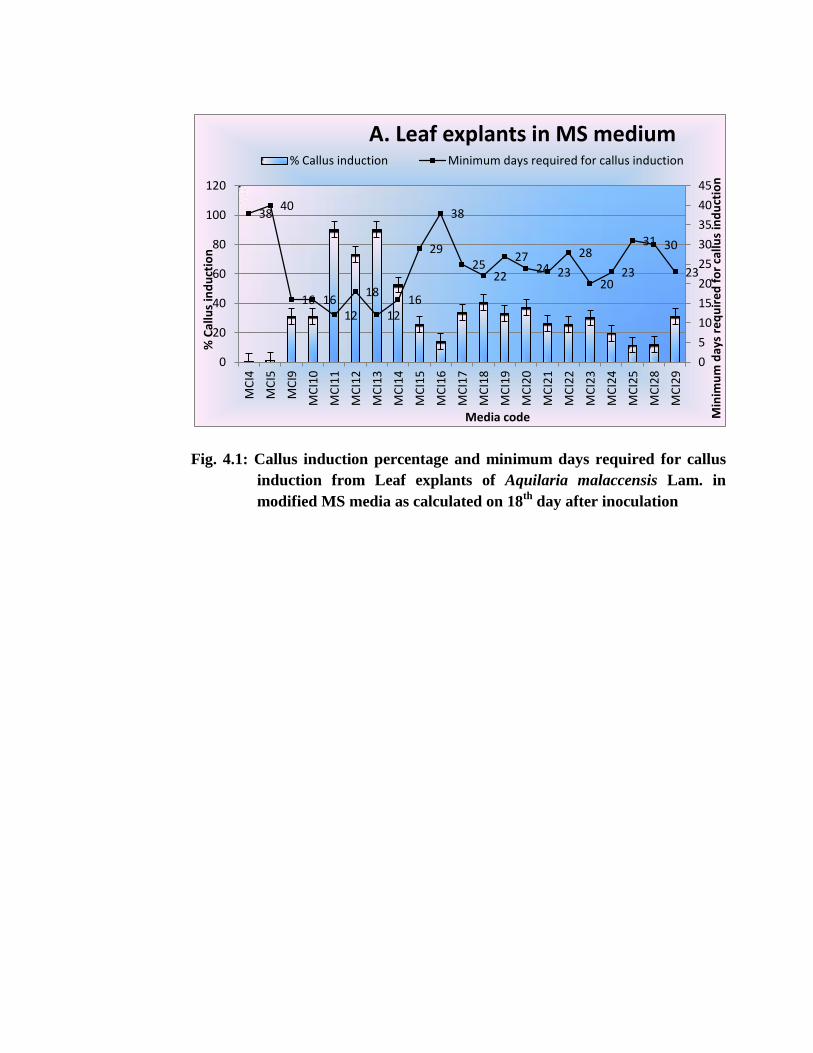
Fig. 4.1: Callus induction percentage and minimum days required for callus
induction from Leaf explants of Aquilaria malaccensis Lam. in
modified MS media as calculated on 18th
day after inoculation
38 40
16 16 12
18
12 16
29
38
25 22
27 24 23
28
20 23
31 30
23
0
5
10
15
20
25
30
35
40
45
0
20
40
60
80
100
120
MC
I4
MC
I5
MC
I9
MC
I10
MC
I11
MC
I12
MC
I13
MC
I14
MC
I15
MC
I16
MC
I17
MC
I18
MC
I19
MC
I20
MC
I21
MC
I22
MC
I23
MC
I24
MC
I25
MC
I28
MC
I29
Min
imu
m d
ays
req
uir
ed
fo
r ca
llus
ind
uct
ion
Media code
% C
allu
s in
du
ctio
n
A. Leaf explants in MS medium % Callus induction Minimum days required for callus induction
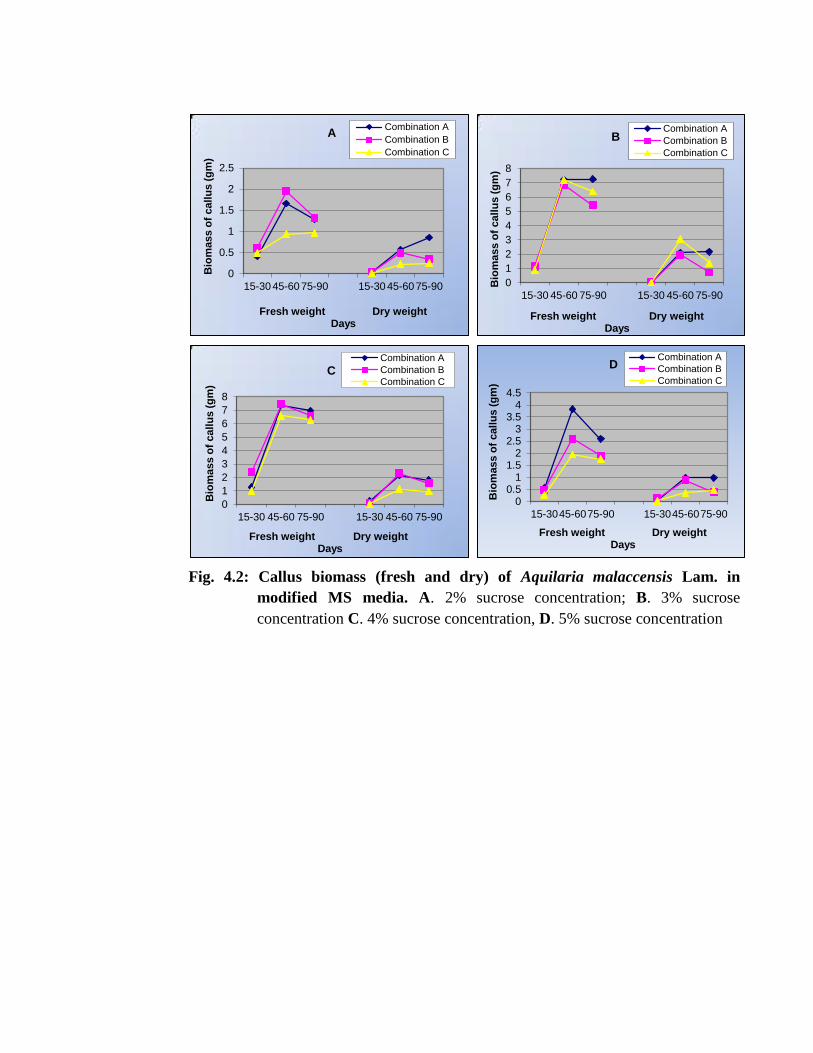
Fig. 4.2: Callus biomass (fresh and dry) of Aquilaria malaccensis Lam. in
modified MS media. A. 2% sucrose concentration; B. 3% sucrose
concentration C. 4% sucrose concentration, D. 5% sucrose concentration
0
0.5
1
1.5
2
2.5
15-3045-6075-90 15-3045-6075-90
Bio
mass o
f call
us
(g
m)
Fresh weight Dry weight Days
A Combination A
Combination B
Combination C
0
1
2
3
4
5
6
7
8
15-30 45-60 75-90 15-30 45-60 75-90
Bio
mass o
f call
us
(g
m)
Fresh weight Dry weight Days
B Combination A
Combination B
Combination C
0
1
2
3
4
5
6
7
8
15-30 45-60 75-90 15-30 45-60 75-90
Bio
mass o
f call
us
(g
m)
Fresh weight Dry weight Days
C Combination A
Combination B
Combination C
00.5
11.5
22.5
33.5
44.5
15-3045-6075-90 15-3045-6075-90
Bio
mass o
f call
us
(g
m)
Fresh weight Dry weight Days
D Combination A
Combination B
Combination C

Plate 4.1: Callus growth from leaf and nodal explants as influenced by the
treatment of different plant growth hormone(s): A & B. Callus
induction from nodal explants in MS medium, C. Callus from leaf
explants in MS + Coconut water, D-F. Callus growth from leaf
explants in MS +2,4D+Kinetin; G. Callus growth in MS+NAA+ BAP;
H. Callus induction in B5 medium; I. Callus induction from leaf
explants in WPM
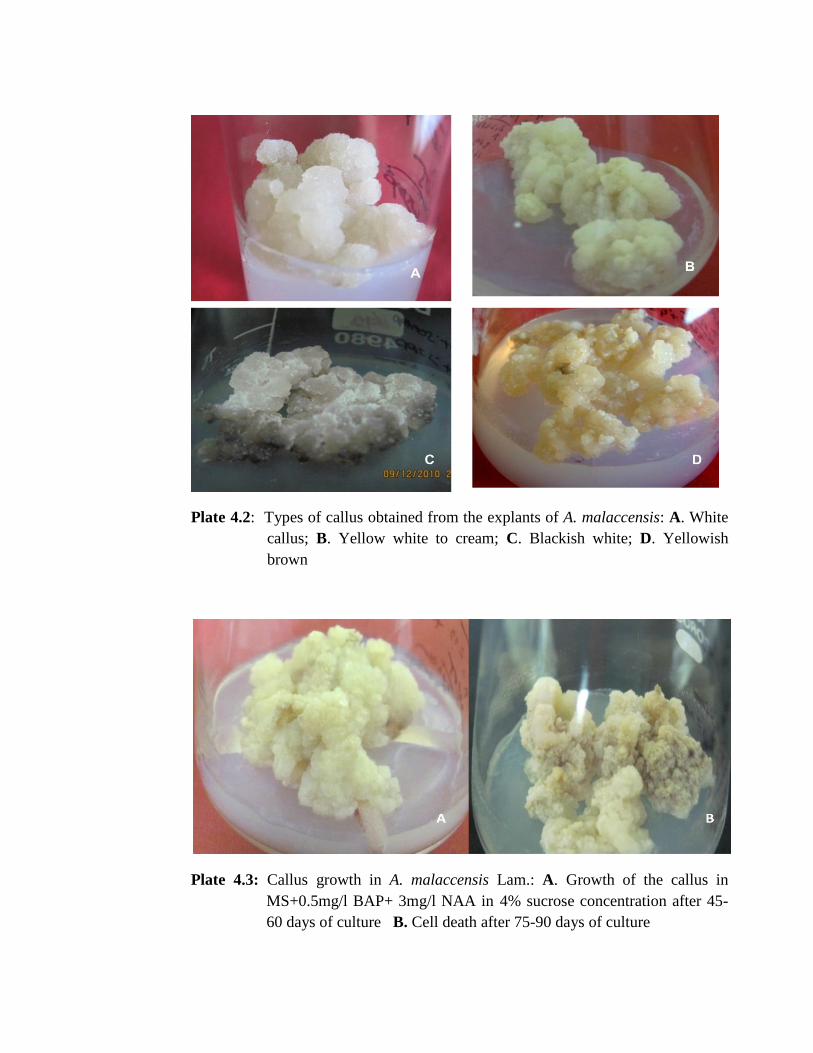
Plate 4.2: Types of callus obtained from the explants of A. malaccensis: A. White
callus; B. Yellow white to cream; C. Blackish white; D. Yellowish
brown
Plate 4.3: Callus growth in A. malaccensis Lam.: A. Growth of the callus in
MS+0.5mg/l BAP+ 3mg/l NAA in 4% sucrose concentration after 45-
60 days of culture B. Cell death after 75-90 days of culture
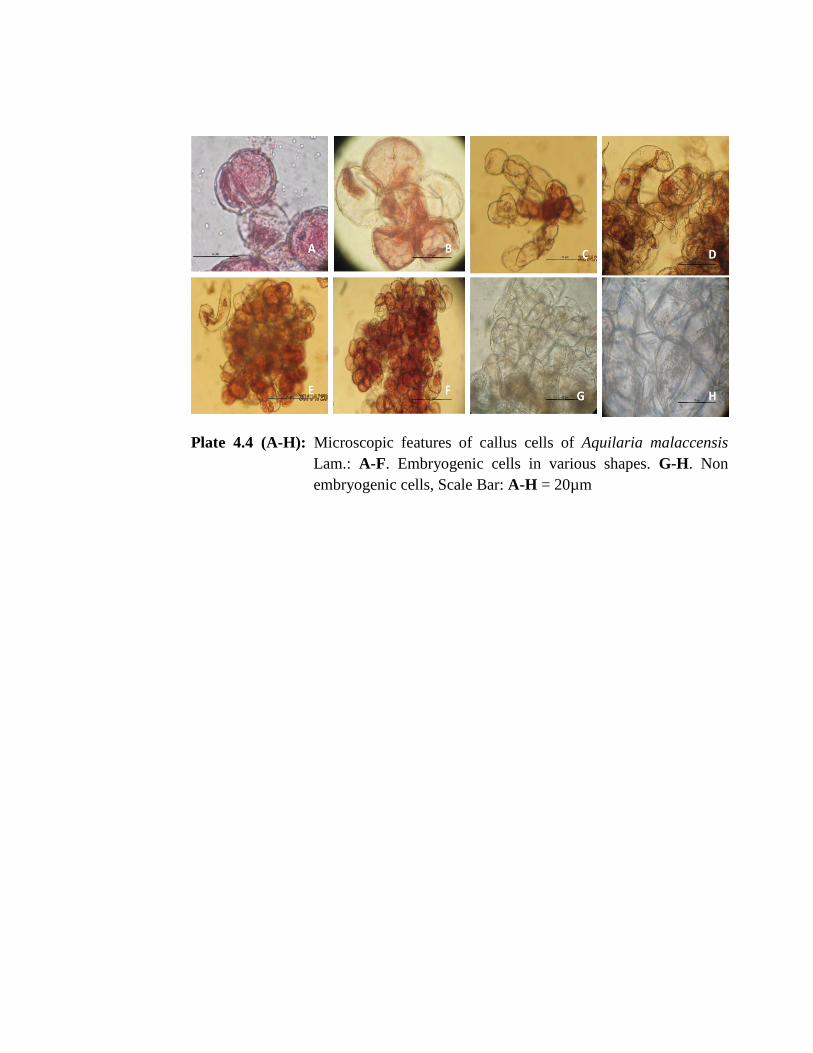
Plate 4.4 (A-H): Microscopic features of callus cells of Aquilaria malaccensis
Lam.: A-F. Embryogenic cells in various shapes. G-H. Non
embryogenic cells, Scale Bar: A-H = 20µm
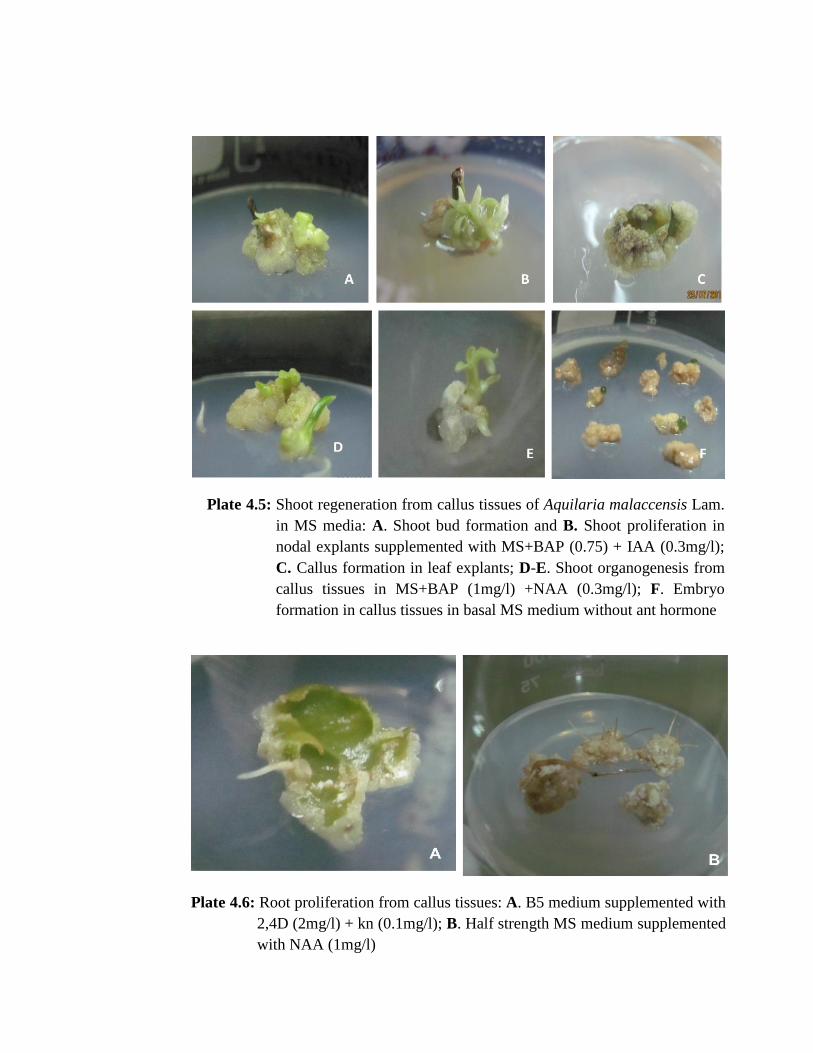
Plate 4.5: Shoot regeneration from callus tissues of Aquilaria malaccensis Lam.
in MS media: A. Shoot bud formation and B. Shoot proliferation in
nodal explants supplemented with MS+BAP (0.75) + IAA (0.3mg/l);
C. Callus formation in leaf explants; D-E. Shoot organogenesis from
callus tissues in MS+BAP (1mg/l) +NAA (0.3mg/l); F. Embryo
formation in callus tissues in basal MS medium without ant hormone
Plate 4.6: Root proliferation from callus tissues: A. B5 medium supplemented with
2,4D (2mg/l) + kn (0.1mg/l); B. Half strength MS medium supplemented
with NAA (1mg/l)
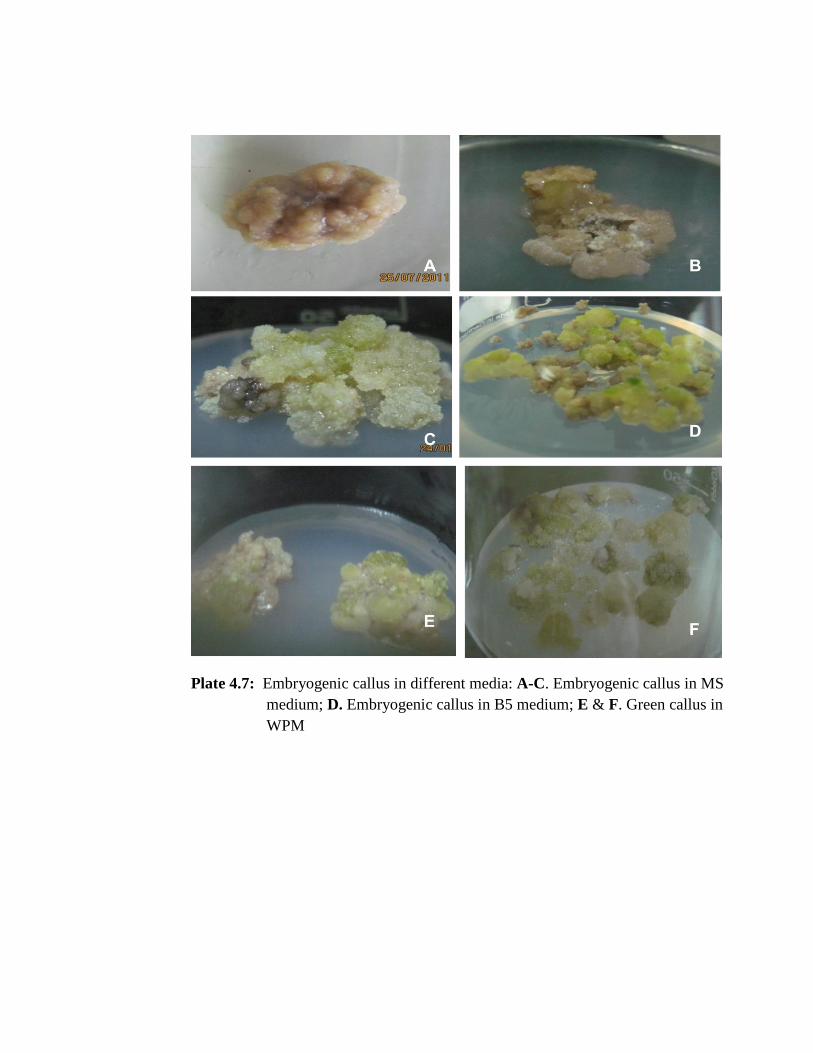
Plate 4.7: Embryogenic callus in different media: A-C. Embryogenic callus in MS
medium; D. Embryogenic callus in B5 medium; E & F. Green callus in
WPM

![Direct Organogenesis from Cotyledonary Node Explants of ... · shoot organogenesis in C. peporeported [19] direct organogenesis in Cucumis sativus [20] and reported L. cy-lindrica](https://img.pdfslide.us/doc/110x75/5fac27dc76c37d66627b9b5d/direct-organogenesis-from-cotyledonary-node-explants-of-shoot-organogenesis.jpg)