Embed Size (px)
Citation preview

Severe abnormalities in the oral mucosa induced by suprabasal
expression of epidermal keratin K10 in transgenic mice.
Mirentxu Santos(1), Ana Bravo(2), Ceferino López (2), Jesús M. Paramio (1,3)
and José L. Jorcano(1)
1.- Project on Cell and Molecular Biology and Gene Therapy, CIEMAT
Av. Complutense 22, E-28040 Madrid, (SPAIN).
2.- Dept. of Animal Pathology, Veterinary School, University of Santiago de
Compostela, Lugo (SPAIN).
3.-To whom correspondence should be addressed.
Phone: 34+ 91 3466438
Fax 34+91346484
e-mail: [email protected]
Running title. Expression of bK6βK10 in transgenic mice
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on July 15, 2002 as Manuscript M205143200 by guest on A
ugust 10, 2019http://w
ww
.jbc.org/D
ownloaded from

ABSTRACT
Previous studies have demonstrated that keratin K10 plays an important role in
mediating cell signaling processes, since the ectopic expression of this keratin induces
cell cycle arrest in proliferating cells in vitro and in vivo. However, apart from its well-
known function of providing epithelial cells with resilience to mechanical trauma, little is
known about its possible roles in non-dividing cells. To investigate what these might
be, transgenic mice were generated in which the expression of K10 was driven by
bovine K6β gene control elements (bK6βhK10). The transgenic mice displayed severe
abnormalities in the tongue and palate, but not in other K6 expressing cells such as
those of the esophagus, nails and hair follicles. The lesions in the tongue and palate
included the cytolysis of epithelial suprabasal cells associated with an acute
inflammatory response and lymphocyte infiltration. The alterations in the oral mucosa
caused the death of transgenic pups soon after birth, probably because suckling was
impaired. These anomalies, together with others found in the teeth, are reminiscent of
the lesions observed in some patients with pachyonychia congenita, an inherited
epithelial fragility associated with mutations in keratins K6 and K16. Though no
epithelial fragility was observed in the bK6βhK10 oral epithelia of the experimental
mice, necrotic processes were seen. Collectively, these data show that the carefully
regulated tissue- and differentiation-specific patterns displayed by the keratin genes
have dramatic consequences on the biological behavior of epithelial cells, and that
changes in the specific composition of the keratin intermediate filament cytoskeleton
can affect their physiology, in particular those of the oral mucosa.
Santos et al., 2
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

INTRODUCTION
Keratin intermediate filaments (KIF) are present in the cytoplasm of all epithelial
cells as heteropolymers of type I and type II keratin polypeptides. Type I and type II
keratin genes display highly regulated expression patterns in a pairwise and
differentiation-specific fashion (1-3). The role of KIF in epithelial cells and tissues
remained elusive until the discovery, through studies with transgenic mice and the
finding of mutations affecting keratin proteins in dominantly inherited epithelial fragility
syndromes (4-8), that keratins impart mechanical resilience to cells. This appears to
be a function shared by the majority of the keratin family. Therefore, the changes in
keratin expression observed during differentiation - or in certain situations such as in
tumor growth or wound healing involving stratified epithelia - probably indicate subtle,
cell type-specific differences in function among these polypeptides. Further keratin-
specific functions should not be discarded.
Previous studies have demonstrated that K10 has specific functions. This
keratin replaces K14 as skin keratinocytes enter the terminal differentiation program
and become postmitotic (9). In addition, K10 expression is severely reduced under
hyperproliferative situations, such as in wound healing and epidermal tumors. We have
previously demonstrated that forced K10 expression in cultured cells induces cell cycle
arrest through a mechanism that requires a functional retinoblastoma gene (10). This
process seems to take place by impairing the activation of Akt and PKCζ, and leads to
reduced cyclin D1 expression (10, 11). Moreover, ectopic human K10 (hK10)
expression in the basal layer of the epidermis of transgenic mice (making use of the
bovine basal keratin bK5 promoter [bK5hK10 mice]), also inhibits cell proliferation and
dramatically impairs tumor development (12). Collectively, these results indicate that
K10 may play a role in the induction and/or maintenance of postmitotic status of
Santos et al., 3
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

suprabasal epidermal cells (10-12). However, no evidence was provided of other
functions of K10 in cells that normally do not divide in vivo, but where this protein is
normally expressed (9). A way of exploring these possible functions is to express K10
ectopically in postmitotic suprabasal cells of transgenic mice.
Keratin 6 (K6) is a type II keratin under elaborate control. It displays constitutive
and inducible expression in various types of complex epithelia. K6 is constitutively
expressed in the suprabasal cells of the paw pad and sole of the foot, the nail bed,
esophagus, trachea, oral cavity, and the outer root sheath of the hair follicles (13-16).
In addition, K6 is induced after injury and in diseases involving altered proliferation or
differentiation in humans and mouse skin epidermis (15, 17-22). The complexity of K6
expression is further increased by the fact that there are six functional K6 genes in
man, three in the mouse (13, 17, 18), and putatively three in cow (J.L.J. unpublished
results). The significance of this diversity is unclear. Finally, inherited mutations
affecting the K6 genes in humans are associated with type I (23) and type II (24)
pachyonychia congenita. These genetic disorders are characterized by severe
dystrophy of the nail plate, and differ principally in the involvement of other stratified
epithelia (6, 19).
The expression pattern of the K6 genes makes them their regulatory regions
appropriate to direct the expression of selected transgenes to stratified epithelia in
transgenic mice (15, 20, 21). We have previously reported two lines of transgenic mice
expressing human keratin K10 (hK10) under the control of the bovine keratin K6β
promoter (bK6βhK10 mice) (25). Although they displayed no overt phenotype, a clear
delay was found in tumor development when these mice were subjected to skin
chemical carcinogenesis protocols (25). However, this is a relatively minor effect
compared to that observed in bK5hK10 transgenic mice, which are almost completely
resistant to tumor development (12). This difference is probably attributable to the
Santos et al., 4
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

expression of hK10 in different cell compartments. The three mK6 genes are normally
absent from interfollicular epidermis, but they are rapidly induced upon
hyperproliferative stimuli in suprabasal keratinocytes (13, 15, 16). Only one, namely
mK6a, is expressed in the basal layer of the hyperproliferative epidermis (13, 16),
where bK5 is expressed (26). In contrast, the bovine bK6β regulatory elements drive
the expression of the transgene, similarly to the endogenous mK6b gene, in the
suprabasal layers of the hyperproliferative epidermis (13, 15, 16). In this compartment,
the keratinocytes display a very limited proliferative activity compared to the basal layer
cells. In addition, differences in the level of K10 expression may also contribute towards
explaining the observed differences in tumorigenic susceptibility between bK5hK10 and
bK6βhK10 transgenic mice. In support of this, heterozygous bK5hK10 mice do not
display overt epidermal abnormalities, whereas hypoplasic and hyperkeratotic
epidermises have been observed in homozygous bK5hK10 transgenic mice in parallel
with increased expression of the transgene (12). This is also in agreement with our
observations demonstrating that the effects of keratin K10 are clearly related to its
expression level (10, 11).
In this work we have tried to address the possible functions of K10 in non-
proliferative cells by studying the consequences of hK10 expression in tissues normally
expressing K6. As previously reported, bK6βhK10 animals display no obvious
phenotype even in homozygosis, we have thus generated new bK6βhK10 transgenic
mice lines bearing a higher copy number of the transgene in order to increase the
expression of hK10 in those cells in which bK6β is active. In this context, it is important
to point out that the bK6β promoter has an expression pattern very similar to the
endogenous mouse keratin K6b (mK6b) (13-16).
All these high copy number transgenic mice display a clear phenotype that
affects the oral mucosa and is characterized by the necrosis of the suprabasal cells of
Santos et al., 5
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

the tongue, palate and gingival epithelium, in association with acute inflammation. This
leads to severe shedding of the epithelium, causing perinatal death by impeding
suckling. Alterations were also seen in the incisors, but no significant anomalies were
observed in nails or hair. Our results indicate that the ectopic expression of K10 in
postmitotic suprabasal cells provokes dramatic alterations in their biological behavior
and indicate that alterations of the specific expression pattern of keratins in a given
epithelium affect the physiological status of the tissue, providing clear evidence of the
functional diversity of these proteins.
Santos et al., 6
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

MATERIALS AND METHODS
Transgene construction and generation of transgenic mice. To generate the
bK6βhK10 construct (13, Fig. 1A), the 9kb fragment containing the bovine bK6β gene
regulatory sequence (15) was inserted, using Asp718 digestion, 5’ of the human k10
gene in the plasmid hK10-MC (25). This plasmid contains the full-length hK10 gene
including the ATG site and the polyadenylation site 0.5 kb downstream (27). The
specificity of the construct was monitored by transfection into K6-expressing and non-
expressing cells (see below and Fig. 1B). Transgenic mice were produced by
microinjection into (C57Bl/10x BALB/c)F2 mouse embryos as previously described (12,
15, 25, 26). For genotyping and identification of the transgenic mice, genomic DNA was
extracted from mouse tails, digested with BamHI, electrophoresed, and transferred
onto nylon membrane (Gene Screen Plus; NEN Life Science Products) for Southern
blotting. The number of copies of the transgene (see Fig 1D) was estimated using
Phosphorimager scanner (Bio-Rad) after normalization. A randomly primed labeled
specific probe was generated using 1 kb of the 3’ untranslated fragment of the hK10
gene.
Cell culture and transfection. Bovine mammary gland BMGE+H cells and
mouse MCA3D keratinocytes were cultured in DMEM containing 10% FBS as
previously reported (10, 11, 28). Transfections using the calcium phosphate method
were performed using cells cultured on glass coverslips as previously described (10,
11, 28). The expression of the transfected bK6bhK10 construct and the endogenous
keratin K6 protein were analyzed by double immunofluorescence using mouse mAb
K8.60 antibody (Sigma; 1/40 diluted) against K10, and a rabbit polyclonal against K6
(Covance; 1/600 diluted). Secondary antibodies and immunofluorescence visualization
Santos et al., 7
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

were as previously described (10, 11, 28).
Histological and immunohistochemical analysis. Freshly collected tissues were
fixed immediately in 10% formaldehyde and left for at least 24 h before being
embedded in paraffin prior to sectioning. 4µm sections were stained with hematoxylin-
eosin. The antibodies used for immunohistochemistry were K8.60 mAb (Sigma), which
recognizes mouse and human K10, and a rabbit polyclonal anti K6 (Covance, CA).
Sections were incubated with a biotinylated anti-mouse or anti-rabbit antibody, and
then with streptavidin conjugated to horseradish peroxidase (DAB kit, Vector). Control
immunostainings using the secondary antibody in the absence of the primary antibody
were routinely performed. Antibody localization was determined using 3,3-
diaminobenzidine (DAB) as the chromogenic substrate for peroxidase (DAB kit,
Vector).
Transmission Electron Microscope (TEM) Analysis. Tongues from 0 - 3 day old
transgenic and control mice were fixed in 2.5% gluteraldehyde in 0.1 M phosphate
buffer (pH 7.5), and postfixed in 1% osmium tetroxide prior to dehydration and
embedding in Epon 812 resin. Semithin sections were stained with 1% toluidine blue
for field selection. Ultrathin sections were stained with uranyl acetate and lead citrate
and analyzed as previously described (12).
Western Blot Analysis. Newborn mice were sacrificed and excised tongues
were snap-frozen in liquid nitrogen. Total protein extracts were prepared as previously
reported (12, 25). Protein concentrations were determined using a modified Bradford
Assay Kit (BioRad). Equal amounts of protein were electrophoresed in 8.5% SDS-
PAGE gels and electroblotted onto nitrocellulose. The membranes were incubated with
the primary antibody, followed by donkey anti–mouse HRP or donkey anti–rabbit HRP
(1:2,000 Jackson Immunoresearch Laboratories). WestPicoSignal (Pierce, Rockford,
Santos et al., 8
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

IL) was used to detect the bands according to the manufacturer’s recommendations.
The primary antibodies used included rabbit polyclonal antiserum directed against K6,
K5, K13 and K10 (BabCo Covance CA, USA), and RCK107 mAb to react with K14
(29).
Santos et al., 9
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

RESULTS AND DISCUSSION
Generation of transgenic mice expressing bK6βhK10.
To monitor the proper expression of the bK6βhK10 construct (Fig. 1 A),
transient transfection experiments were performed in MCA3D mouse keratinocytes
which express K6. Synthesis and incorporation of hK10 into the endogenous keratin
cytoskeleton was observed in all transfection experiments, with no sign of cytoskeletal
disruption (arrows in Fig. 1B, B’). Similar results were obtained using bovine BMGE+H
cells. No expression of the construct was detected when using cell lines that did not
express K6 (VeroC, PtK2 and NIH3T3, not shown). Since the absence of keratin
clumping and the proper incorporation of hK10 into the keratin cytoskeleton was
observed in all experiments, these results indicate that K10 does not cause the
collapse of the endogenous cytoskeleton, contrary to the reported effects of increased
hK16 (30-32) and mutant mK6 (14) expression.
The linearized bK6βhK10 construct was subsequently injected into fertilized
oocytes. We have previously reported that in animals with 15 or less copies of the
bK6βhK10 transgene, no overt phenotype (besides a delay in tumor formation) is observed
(25). Given that the expression levels of transgenes under the control of the bK5 and
bK6 promoter regions are frequently proportional to the copy number of the transgene
integrated (12, 25, 33 and data not shown), attempts were made to generate transgenic
mice with more than 25 copies of the transgene. Two founders were obtained bearing
50 and 75 copies respectively (see Fig 1D). These animals displayed no overt
phenotype. However, further analysis indicated that this was attributable to mosaicism
in the expression of the transgene, as reported in other transgenic mice models (data
not shown, see 33 for a careful discussion). As expected, however, the offspring
derived from these founders displayed no such mosaicism in K10 expression (see
Santos et al., 10
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

below) and showed severe phenotypic alterations, which finally lead to death between
days 3 and 5 after birth. Both transgenic mice lines had similar phenotypic alterations
and are collectively described as high copy number transgenics.
Overexpression of bK6βhK10 leads to severe necrosis and acute inflammation of the
oral mucosa
Transgenic bK6βhK10 pups appeared normal at birth but, within a few days,
they were smaller, weaker, and had less milk in their stomachs (sometimes none) in
comparison with their control littermates (Fig. 1C arrows). These pups generally died
between 3-5 days postpartum, weighing about half that of their littermate controls (1.4
g and 2.9 g respectively, on average). At death, the skin and nails of these mice
appeared normal (Fig. 1C, and data not shown) and showed no obvious developmental
anomalies other than reduced size and frail appearance.
Since the bK6β transgene is constitutively expressed in several oral epithelia
(15), and since one possible reason for the observed mortality of the bK6βhK10
transgenics could be poorly feeding, the anatomy of the oral cavity of these mice was
examined. The dorsal surface of the tongue, and to a lesser extent the ventral surface
of the upper palate, were covered with white plaques from the mid region to the
pharynx (denoted by lp in Fig. 1D, and data not shown). Non-transgenic littermates
(Fig. 1D’) showed no signs of these lesions. Similar features have been reported for
mice lacking keratins mK6a and mK6b (13, 16), and are similar to the oral leukoplakia
seen in some pachyonychia congenita patients (23, 34).
Histological studies of sections from the tongue of bK6βhK10 mice
demonstrated severe damage from the mid to the posterior region of the dorsal
epithelium. Extensive areas with consistent features of coagulative necrotic changes
were seen, with loss of cell cytoplasm and degenerative nuclei. Some tissue
Santos et al., 11
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

organization was still present, however, as seen by the remaining papillae profiles,
although these were heavily infiltrated by neutrophils (Fig. 2A´´ and 3C; compare with
controls in 2A and 3A, and no-lesion tongue in Fig. 3B). Similar features were also
observed in the palate (Figs. 2B´ and 3D; compare with controls in 2B and 3A) and in
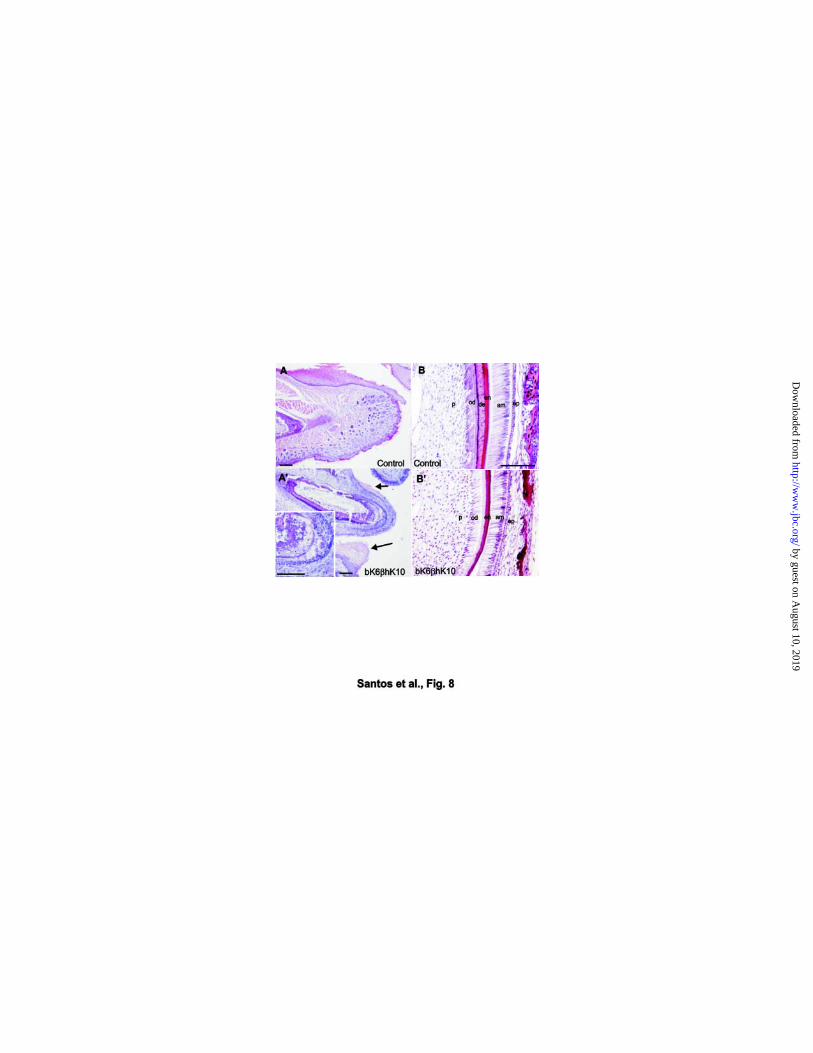
the gingival epithelium of the incisors (Fig. 8A´; compare with control in 8A). In all cases
the apparent thickening of the non-affected epithelial cell compartments was observed,
giving rise to a moderate hyperplasic phenotype, probably caused by inflammation and
infiltration of the tissue. The lesions of the tongue primarily affected the filiform papillae,
which are the most abundant papillary type in mice. In these, three distinct programs of
epithelial differentiation exist (Fig. 2A’), giving rise to the anterior (ac), posterior (pc)
and buttress (bc) columns (35, 36). Interestingly, the data obtained with mK6a/b-null
mice suggest that these filiform papillae are particularly sensitive to mechanical stress
and alterations of the keratin cytoskeleton (13, 16). The bK6β promoter is active in the
anterior and buttress columns (Fig. 3B), as previously demonstrated by β-Gal staining
in bK6βLacZ transgenic mice (15). In agreement, the immunohistochemical analysis of
the tongue of newborn transgenic mice showed a coincident expression of mK6 (Fig.
3A, wild type tongue denoted by t) and the transgene (Fig. 3B, non-lesional region of
the tongue). Consistently, K10 expression was also observed in the necrotic
suprabasal regions of the epithelium in the tongue (Fig. 3C) and palate (Fig. 3D).
The injured superficial areas expanded the tongue and palate epithelia with a
mixture of neutrophils and cell debris disrupted from the several unaffected rows of
basaloid cells normally attached to the basement membrane (Fig. 2A’’, Fig. 4B,
semithin section). This shedding process of the necrotic suprabasal areas was
probably induced by the intensive discharge of gelatinase and other proteases present
in the granules of the abundant neutrophils. No major alterations were observed in the
subjacent muscle of the tongue (Fig. 4B). Most of the pathological changes in the
Santos et al., 12
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

lingual, gingival and palate epithelia are similar to those described for mice lacking
mK6a and mK6b genes (13, 16). In these deficient mice, the lesions are associated with a
decrease in - or even the absence of - KIF in the anterior compartment, which induces
an important increase in the size of intercellular spaces (13, 16). Although a similar
absence of KIF was not expected, it could not be ruled out a priori that other changes in
the keratin cytoskeleton might occur in the bK6βhK10 mice. To investigate such
possible changes, ultrastructural analyses were performed.
Ultrastructural pathology of the tongue of bK6βhK10 transgenic mice
The dorsal epithelium of the tongue in control and transgenic mice was studied
by transmission electron microscopy. In wild-type animals, differentiating keratinocytes
in the anterior column (AC) showed electron dense granules similar to those found in
the granular layer of the epidermis (37). The buttress column (BC) keratinocytes had a
flattened shape and large bundles of densely packed keratin IF, but no keratohyalin
granules. Finally, the keratinocytes of the posterior column (PC) were rounded and had
a well-organized keratin cytoskeleton, although less bundled than in BC keratinocytes.
An example of the different regions of a control filiform papilla, as observed with the
electron microscope, is provided in Fig. 4A. Samples taken from the same region in
bK6ßhK10 transgenic mice (semithin section in Fig. 4B) showed dramatic alterations in
the AC and BC keratinocytes, readily visible through their cytolytic and distended
appearance and unusually clear and swollen cytoplasm (asterisks in Fig. 4D and 5A).
The AC cells appeared to have a normal complement of keratohyalin granules, but
which were larger than those of wt. mice (Fig. 4C; wild type Fig. 4A). Desmosomes and
also keratin IF were found in these cytolytic swollen cells (data not shown and Fig. 5A
arrows). Similar cytolytic events were observed in BC keratinocytes (Fig. 4C), which
also displayed normally arranged keratin IF bundles and desmosomes (Fig. 5B). No
Santos et al., 13
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

major abnormalities were observed in the PC or basal keratinocytes (Fig. 4D and Fig.
5A), which do not express the transgene (Fig. 3B, C). Neutrophil leukocytes were seen
infiltrating the cytolytic areas of the AC and BC (arrows in Fig. 4D, pmn in Fig. 5A and
data not shown). Despite the severe cytolytic changes observed in these regions, the
affected cells maintained intact desmosomes (white arrows in Fig. 5B) and the
intercellular spaces were not enlarged (Fig. 4C, D and Fig. 5A). As the hK10
expression is restricted to the AC and BC in agreement with the expression of
endogenous mK6 (compare Fig. 3B with Fig. 3A), these findings suggest that these
cells are particularly susceptible to changes in the expression, either quantitative or
qualitative, of keratin polypeptides. In this regard, the blisters occurring in mK6a/b-
deficient mice have been attributed to the absence of keratin IF in these cells (13, 16),
whereas keratin IF and desmosomes normally arranged were clearly observed even in
cytolytic cells (Fig. 5A, B). Data obtained with mK6a/b-deficient mice have been
interpreted as an indication that the cells in the AC region of the filiform papillae are
particularly sensitive to the mechanical stress of suckling (13, 16). However, some
animals were maintained by parenteral feeding and similar lesions still occurred (not
shown), indicating that mechanical stress is not the only origin of the observed tongue
and palate anomalies. Nonetheless, it is worth mentioning that some differences exist
between the lesions observed in mK6a/b null mice and those of our transgenic animals.
In particular, desmosomes and keratin IF are normally arranged and intercellular
edema is absent from the tongue lesions in bK6βhK10 mice, whereas in K6a/b-
deficient mice keratin IF is decreased or absent in the AC, and the intercellular spaces
are enlarged (13, 16). This clearly points to a different etiology in these apparently
similar lesions: in the deficient mice there is a clear epithelial fragility process, whereas
in the present case the defects are more related to an acute inflammatory response.
Santos et al., 14
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

The expression of other keratins is not altered in the tongue of bK6βhK10 transgenic
mice.
The above results do not rule out the possibility that the ectopic expression of
hK10 can affect the composition of the keratin cytoskeleton of the tongue keratinocytes.
To investigate this possibility, protein extracts prepared from whole tongues of low copy
number (15 copies) non-transgenic and high copy number (75 copies) transgenic mice
(Fig. 6) were studied by Western blotting. As a control, protein extracts from human
skin were included. The anti K10 antibody detects a single polypeptide in the extracts
from human skin and transgenic tongues. In addition, the level of expression of hK10
was increased in those extracts from transgenic animals with high transgene copy
number. The amounts of the characteristic keratin polypeptides of the suprabasal
tongue keratinocytes, namely K13 and K6 (36), were not significantly different between
the transgenic and control samples. Finally, compared with the controls, the protein
levels of the basal keratins K5 and K14 were not altered in transgenic samples.
Collectively, these results indicate that the expression of hK10 does not produce major
alterations in the keratin expression profile. It is worth mentioning that the hK10
expression levels in high copy number transgenic mice are very similar to those of
endogenous hK10 in human epidermis, indicating that the phenotype found in these
mice cannot be attributed to an aberrant excessive overexpression of the transgenic
protein.
bK6bhK10 transgenic mice have no hair or nail abnormalities.
Keratin K6 is constitutively expressed in the outer root sheath of the hair follicles
and nail bed epithelium. Possible alterations in these cutaneous structures were
therefore studied. No abnormalities were found between control and transgenic mice in
the hair or interfollicular skin by day 4 after birth (Fig. 7A, A’, respectively). To see
Santos et al., 15
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

whether the absence of such defects could be due to a lack of hK10 transgene
expression, the expression of K6 and K10 were studied in samples from wt and
transgenic mice. The results confirmed the expression of hK10 in the outer root sheath
of transgenic mouse hairs (Fig. 7C), coincident with endogenous mK6 expression (Fig.
7B). No expression of K10 was detected in the hair follicles of control mice (Fig. 7D).
No defects were seen in the nails of the transgenic mice (data not shown). Neither
were defects seen in hair follicles or nails of K6a/b-deficient mice (13, 16). This can be
explained in that these mice die too early to develop any possible alterations. In
agreement with this hypothesis is the appearance of hair and nail lesions in
pachyonychia congenita patients several months after birth. However, in the mK6a/b-
deficient mice, the absence of hair and nail abnormalities might also be attributed to the
expression of the recently described mK6hf gene (13), which appears to be specifically
expressed in the hair follicles and nail beds and may compensate for mK6a/b
deficiency (13). The present results also point to the possibility that quantitative and/or
qualitative alterations in the normal characteristic keratin expression pattern may cause
functional anomalies in certain keratinocytes (e.g., AC and BC tongue keratinocytes),
but not in others (e.g., outer root sheath keratinocytes).
As with the hair follicles, no alterations were detected in the esophagus or forestomach (not
shown). Again, the absence of abnormalities may be due to the early death of bK6βhK10 transgenic
mice, which precludes the detection of lesions that might occur later. Supporting the
early lethality hypothesis is the appearance of esophageal alterations only 2 months
after birth in K4-deficient mice (38), in clear contrast with the absence of alterations in
the tongue of these null mice. However, it cannot be ruled out that the levels of hK10
transgene expression may vary among the different K6-expressing cells in the different
tissues, thus promoting phenotypic changes (or not) or that these cells may display
different intrinsic susceptibility to possible K10-induced effects.
Santos et al., 16
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Tooth abnormalities in bK6bhK10 transgenic mice.
Different tissues from bK6βhK10 mice were analyzed and no ectopic expression
of K10 besides that observed in K6-expressing cells was seen. The findings in oral
mucosa closely resemble those observed in patients suffering from pachyonychia
congenita. One of the variants of this disease, the Jackson-Lawler form or
pachyonychia congenita type II, is characterized by the presence of neonatal teeth (24,
39-41). We therefore investigated the presence of abnormalities in the teeth of the
transgenic mice. These animals displayed severe teeth anomalies such as defects in
position and precocious eruption of the incisors, in some cases associated with
microdontia (Fig. 8A´ and data not shown; wild type in Fig. 8A). Hematoxylin-eosin
sections revealed a clear decrease (even absence) of the thickness of the dentine layer
(de in Fig. 8B, wild type; transgenic in 8B’) associated with degenerative changes in
the ameloblast and odontoblast layers (am and od in Fig. 8B, B’, wild type and
transgenic respectively). In addition, some degenerative changes were also observed
(see inset in Fig. 8A’). Collectively, these abnormalities are clearly suggestive of
specific roles of keratin expressing cells in the process of tooth growth. In this regard, it
has been demonstrated that keratins participate in the assembly of amelogenin during
amelogenesis, supporting a possible involvement of these proteins in tooth
development (42). This important and complex issue is the subject of future studies.
Many of the phenotypic features observed in the bK6βhK10 mice resemble
those characterized in pachyonychia congenita patients. This disease is an autosomal
dominant ectodermal dysplasia caused by mutations in keratins K6, K16 and K17,
whose major hallmark is hypertrophic nail dystrophy (6). Two main types of this
disorder have been characterized, type I or the Jadassohn-Lewandowsky form, in
which pachyonychia occurs in conjunction with palmoplantar keratoses and oral
leukokeratoses (6, 23), and type II or the Jackson-Lawler form, characterized by hair
Santos et al., 17
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

abnormalities and neonatal teeth, but no oral lesions (6, 24). The phenotype reported
here displays some characteristics coincident with type I (tongue and palate lesions),
and others with type II (incisor anomalies). However, no alterations in nails (a common
feature of type I and II), the palmoplantar epidermis (type I) or hair abnormalities (type
II) were seen, suggesting that the alterations found in bK6βhK10 transgenics is not a
pachyonychia-like phenotype.
Similar to other keratins, K10 is involved in providing mechanical resilience to
epidermal cells, as demonstrated by the presence of disruptive K10 mutations in
epidermolytic hyperkeratosis (or BCIE, see 6 and references therein) and reinforced by
the phenotype displayed by newborn K10-deficient mice (43). Therefore, it is difficult to
explain why the ectopic expression of this keratin in oral epithelia leads to alterations
reminiscent of those due to the changes in the keratin cytoskeleton associated with
epithelial fragility. One possible explanation is that the abnormal KIF cytoskeleton
observed in pachyonychia congenita keratinocytes, and characterized by densely
aggregated keratin filament bundles (41), might also be produced by increased
expression of K10. In this regard, we have previously reported that K10 forms highly
bundled filaments in transfected cells and in transgenic mouse epidermis (12).
However, we did not detect altered KIF in the affected cells of the AC and BC
compartments of the tongue of bK6βhK10 transgenic mice (Fig. 4 and 5). Further, the
phenotypic alterations in these mice are frequently associated with an acute
inflammatory response, with high leukocyte infiltration (Fig. 2 and 3). No such
alterations have been reported in affected tissues of pachyonychia congenita patients,
and only mild infiltration was found in mK6a/b-deficient mice (16). Determining whether
this inflammatory response is a cause or a consequence of the disruption of the tongue
epithelia requires further analysis. However, the implication of K10 in the modulation of
Akt-dependent signaling (11, 12) suggests that the presence of ectopic K10 might also
Santos et al., 18
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

affect the expression or activity of certain inflammatory cytokines in the affected
epithelia. Among them, TNF-α is of particular interest, since it has recently been
demonstrated that the alterations in the keratin cytoskeleton may modulate the activity
and response to this molecule (44-46). These aspects will be addressed in future
experiments using bK5hK10 and bK6βhK10 transgenic mice.
Santos et al., 19
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

REFERENCES
1 Moll, R., Franke, W.W., Schiller, D.L., Geiger, B., and Krepler, R. (1982)
Cell. 31, 11-24.
2 Quinlan, R.A., Schiller, D.L., Hatzfeld, M., Achtstatter, T., Moll, R.,
Jorcano, J.L., Magin, T.M. and Franke, W.W. (1985). Ann N Y Acad Sci. 455, 282-306.
3 O’Guin, W.M., Schermer, A., Lynch, M., and Sun, T.-T. (1990). In
Goldman R.D., Steinert P.M., eds. Cellular and Molecular Biology of Intermediate
Filaments. London, Plenum Publishing Co, 301-334.
4 Fuchs, E., and Weber, K. (1994). Ann. Rev. Biochem. 63, 345-382.
5 Fuchs, E., and Cleveland, D.W. (1998). Science. 279, 514-519
6 Irvine, A.D., and McLean, W.H.I. (1999). Br. J. Dermatol. 140, 815-828.
7 Takahashi, K., Coulombe, P.A., and Miyachi, Y. (1999). J. Dermatol. Sci.
21, 73-95.
8 Arin. M.J. and Roop, D.R. (2001) Trends Mol. Med. 7, 422-424.
9 Fuchs, E.V. and H. Green. (1980). Cell. 19, 1033-1042.
10 Paramio, J.M., Casanova, M.Ll., Segrelles, C., Mittnacht, S., Lane, E.B.
and Jorcano, J.L. (1999). Mol. Cell Biol. 19, 3086-3094.
11 Paramio, J.M., Segrelles,C., Ruiz, S., and Jorcano, J.L. (2001) Mol. Cell
Biol. 21, 7449-7459
12 Santos M, Paramio JM, Bravo A, Ramirez A, Jorcano JL.(2002) J Biol
Chem. 277, 19122-19130
13 Wojcik, S.M., Longley, M.A. and Roop, D.R. (2001) J Cell Biol. 154,
619-630.
14 Wojcik, S.M., Imakado, S., Seki, T., Longley, M.A., Petherbridge, L.,
Bundman, D.S., Bickenbach, J.R., Rothnagel, J.A., and Roop, D.R (1999)
Differentiation. 65, 97-112.
Santos et al., 20
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

15 Ramirez, A., Vidal, M., Bravo, A., Larcher, F., and Jorcano, J.L. (1995).
Proc. Natl. Acad. Sci. USA. 92, 4783-4787.
16 Wong, P., Colucci-Guyon, E., Takahashi, K., Gu, C., Babinet, C., and
Coulombe, P.A. (2000) J Cell Biol. 150, 921-928.
17 Rothnagel, J.A., Seki, T., Ogo, M., Longley, M.A., Wojcik, S.M.,
Rundman, D., Bickenbach, J.R., and Roop, D.R. (1999). Differentiation. 65, 119-130.
18 Takahashi, K., Yan, B., Yamanishi, K., Imamura, S., and Coulombe, P.A.
(1998). Genomics. 53, 170-183.
19 McGowan, K.M., and Coulombe, P.A. (1998). In Harris J.R., Herrmann
H., eds. Subcellular Biochemistry, Intermediate Filaments. London, Plenum Publishing
Co, 141-165.
20 Takahashi K, Coulombe PA. (1997) J Biol Chem. 272, 11979-11985.
21 Takahashi K, Coulombe PA. (1996) Proc Natl Acad Sci U S A. 93,
14776-14781.
22 Weiss, R.A.R., Eichner, R. and Sun, T.T. (1984). J. Cell Biol. 98, 1397-
1406
23 Bowden, P.E., Haley, J.L., Kansky, A., Rothnagel, J.A., Jones, D., and
Turner, R.J. (1995). Nat. Genet. 10, 363-365
24 Smith, F.J., Jonkman, M.F., van Goor, H., Coleman, C.M., Covello, S.P.,
Uitto, J., and McLean, W.H. (1998). Hum. Mol. Genetics. 7, 1143-1148.
25 Santos, M., Ballestín, C., García-Martín, R. and Jorcano, J.L. (1997).
Mol. Carcinogenesis 20, 3-9
26 Ramirez A, Bravo A, Jorcano JL, Vidal M.(1994) Differentiation. 58, 53-
64.
27 Blessing, M., Rüther U., and Franke, W.W. (1993). J. Cell Biol. 120, 743-
755.
Santos et al., 21
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

28 Paramio, J.M. and Jorcano, J.L. (1994). Exp. Cell Res. 215, 319-331
29 Purkis, P.E., Steel, J.B., Mackenzie, I.C., Nathrath, W.B., Leigh, I.M.,
and Lane E.B. (1990) J Cell Sci. 97, 39-50.
30 Paladini, R.D. and Coulombe, P.A. (1998). J. Cell Biol. 142, 1035-1051
31 Wawersik, M., and Coulombe, P.A. (2000) Mol Biol Cell. 11, 3315-3327
32 Wawersik, M.J., Mazzalupo, S., Nguyen, D., and Coulombe, P.A. (2001)
Mol Biol Cell. 12, 3439-3450.
33 Ramírez, A., Milot, E., Ponsa, I., Marcos-Gutierrez, C., Page, A.,
Santos, M., Jorcano, J., and Vidal, M. (2001). Genetics 158, 341-350.
34 Lim, T.W., Paik, J.H., and Kim, N.I. (1999) J Dermatol. 26, 677-681.
35 Hume, W.J., and Potten, C.S. (1976). J. Cell Sci. 22, 149-160
36 Lindberg, K., and Rheinwald, J.G. (1990) Differentiation. 45, 230-241.
37 Manabe, M., and O’Guin, W.M. (1994). Differentiation. 58, 65-75.
38 Ness, S.L., Edelmann, W., Jenkins, T.D., Liedtke, W., Rustgi, A.K., and
Kucherlapati, R. (1998). J. Biol. Chem. 273, 23904-23911.
39 Gorlin, R.J., Cohen, M.M., and Levin, L.S. (1990). In Gorlin R.J., Cohen
M.M., Levin L.S., eds. Oxford Monographs on Medical Genetics No. 19, Syndromes of
the Head and Neck. New York, Oxford University Press, 445-448.
40 Smith, F.J., Corden, L.D., Rugg, E.L., Ratnavel, R., Leigh, I.M., Moss,
C., Tidman, M.J., Hohl, D., Huber, M., Kunkeler, L., Munro, C.S., Lane, E.B., and
McLean, W.H. (1997) J Invest Dermatol. 108, 220-223.
41 McLean, W.H., Rugg, E.L., Lunny, D.P., Morley, S.M., Lane, E.B.,
Swensson, O., Dopping-Hepenstal, P.J., Griffiths, W.A., Eady, R.A., Higgins, C., et al.
Nat Genet. 9, 273-278.
42 Ravindranath, R.M., Tam, W.Y., Bringas, P. Jr., Santos, V., and
Fincham, A.G. (2001) J Biol Chem. 276, 36586-36597.
Santos et al., 22
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

43 Reichelt, J., Büssow, H., Gurnd, C., Magin, T.M. (2001) Mol. Biol. Cell
12, 1557-1568
44 Caulin, C., Ware, C.F., Magin, T.M., Oshima, R.G. (2000) J. Cell Biol.
149, 17-22.
45 Gilbert, S., Loranger, A., Daigle, N., Marceau, N. (2001) J. Cell Biol. 154,
763-773
46 Inada, H., Izawa, I., Nishizawa, M., Fujita, E., Kiyono, T., Takahashi, T.,
Momoi, T., Inagaki, M. (2001) J. Cell Biol. 155, 415-425.
Santos et al., 23
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE LEGENDS
Fig. 1. Structure of the transgene, expression in cultured cells and gross abnormalities
in bK6βhK10 transgenic mice. A) Structure of the bK6βhK10 transgene used in these
studies, including the bK6β gene regulatory region (shadowed) and the hK10 gene
showing the exons and introns. B) Expression of bK6βhK10 in MCA3D mouse
keratinocyte cells 48 hours after transfection. Transfected cells were analyzed by
double immunofluorescence for hK10 (B) and endogenous K6 (B’) expression. Note
that in all cases K10 is integrated into the endogenous keratin cytoskeleton (at least
200 cells were scored per experiment; experiments were performed in triplicate). C)
Example of Southern blot showing the identification of the transgenic animals and the
estimated copy number of the transgene after normalization and Phosphorimager scan.
D) Appearance of transgenic (right) and wild type (left) 3 day-old littermate. Note the
runt appearance of the transgenic animal. The arrows indicate the empty stomach in
the transgenic compared with the milk-containing stomach in its wild type littermate. E)
Abnormalities in transgenic tongue, note the leukoplakia lesions observed from the mid
to the posterior region of the dorsal tongue in the transgenics (denoted by lp). E´)
Normal tongue in wild type littermate.
Fig. 2. Histological abnormalities in the epithelium of the tongue and palate of
bK6βhK10 transgenic mice. Hematoxylin-eosin stained sections of the tongue (A, A´, A´´) and
palate (B, B´) in wild type (A, A´, B) and transgenic mice (A´´, B´). See the common
filiform papillae arrangement of the dorsal epithelium in the tongue of the wild type (A)
compartmentalized in the anterior column, posterior column and buttress column (ac,
pc and bc respectively in A´). The transgenic suprabasal layers of the epithelium in the
tongue (A´´) and palate (B´) appeared necrotic and infiltrated by neutrophil leukocytes,
Santos et al., 24
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

while the basal layers remained unaffected. A moderate increase in the number of cell
layers in this region can also be observed in the transgenic samples. Dashed lines
denote the epithelial boundaries in A’’, B’. Bar = 100µm.
Fig. 3. Immunohistochemical analysis of K6 and K10 expression in the epithelium of the
tongue and palate of bK6βhK10 transgenic mice. Peroxidase-hematoxylin stained
sections of the tongue (A, B, C) and palate (A, D) in wild type (A) and transgenic mice
(B, C, D). The mK6 expression is restricted to the suprabasal epithelium of the palate
(p in A) and the anterior and buttress column of the tongue (t in A). K10 in the
transgenics is strongly expressed in the same regions of the non-lesional regions of
the tongue (B), as well as in the necrotic plaque of the tongue (C) and the palate (D).
Dashed lines denote the epithelial boundaries Bars = 100µm.
Fig. 4. Ultrastructural analysis of the tongue of bK6βhK10 transgenic mice. Semithin
toluidine-blue stained section (B) and electron microscope appearance (A, C, D) of the
tongue epithelium in wild type (A) and transgenic mice (B, C, D). A) Note the
characteristic features of the filiform papillae in the wild type, organized in three
compartments, anterior column with abundant keratohialin granules (AC), buttress
column with heavily packed bundles of keratin IF (BC) and posterior column (PC). SC
denotes the stratum corneum. B) Toluidine blue-stained semithin section of the
lesional tongue of the bK6bhK10 transgenic mice. The big arrow denotes the shedding
region. C) High magnification obtained by electron microscopy of the superficial region
marked by the upper rectangle in B. Note that this necrotic plaque still preserves part of
the papillary structure and that anterior column cells (AC) have enlarged keratohialin
granules, whereas buttress column cells (BC) display abundant keratin bundles. D)
High magnification obtained by electron microscopy of the region marked by the lower
Santos et al., 25
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

rectangle in (B). Asterisks indicate the abnormally swollen and clear cytoplasm in AC
cells. Open arrows point to the keratohialin granules characteristic of this compartment.
Black arrows show numerous neutrophil leukocytes infiltrating the cytolytic AC areas.
Note that posterior column and basal cells are unaffected (PC and BC, respectively); C
= conjunctive cells in the submucosa. In B m marks the muscular layer. Dashed lines in
B D denote the epithelial boundaries and the basement membrane.
Fig. 5. Presence of keratin filaments and normal desmosomes in altered AC and BC
cells of bK6βhK10 transgenic mice. Electron microscopy analysis of the epithelium of
the tongue in transgenic mice. A, Cytolytic cells in the anterior column showing swollen
and clear cytoplasm (asterisks), keratohialin granules (kh) and KIF (arrows); N =
nucleus, pmn = neutrophils. Note that the underlying posterior column cells remain
unaffected. B, Intercellular space between two BC cells showing a close cellular union
and desmosomes (white arrows); f = keratin IF bundles, m = mitochondria.
Fig. 6. Absence of major alterations in the keratin expression pattern of tongue
epithelium. Analysis of keratin polypeptides in tongue epithelium. Western blots were
performed using 10 µg of total protein extracts from human skin, non-transgenic and
transgenic mice, with low and high copy numbers. The antibodies specifically reacting
with the indicated keratins were used. Note that no significant changes were observed
in the levels of mK13, mK14, mK5 or mK6 keratins among the different murine tongue
samples.
Fig. 7. Absence of alterations in interfollicular epidermis and hair follicles of bK6βhK10
transgenic mice. Hematoxylin-eosin (A, A´) or Peroxidase-hematoxylin stained (B, C,
D) dorsal skin sections from wild type (A, B, D) and transgenic mice (A´, C). The
Santos et al., 26
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

morphology of the epithelium and anagen hair follicles is similar in the transgenic (A´)
and wild type (A) animals. K10 in the transgenic skin is expressed in the suprabasal
layer of the epidermis (C) and the outer root sheath of hair follicles (arrows in C)
coinciding with the expression of K10 (D) and mK6 (B) in the wild type. Bar = 100µm.
Fig. 8. Alterations in the incisors of bK6βhK10 transgenic mice. Hematoxylin-eosin
stained sections of the lower jaw (A, A´) and incisor (B, B´) in newborn wild type (A, B)
and transgenic mice (A´, B´). Note the precocious eruption of the incisor in the
transgenic (A´) in comparison with the wild type (A). The gingival epithelium displays
similar necrotic changes in suprabasal layers as described for the tongue and palate
epithelium (arrows in A´). Inset in A’ shows the degenerative region in the transgenic
incisor. Higher magnification of these degenerative areas of the transgenic incisor (B´),
compared to wild type littermate (B). Note the affected ameloblast (denoted by am) and
odontoblast (denoted by od) layers as well as the absence of the dentine layer
(denoted by de in B). p = dental pulp, od = odontoblast layer, de = dentine, en =
enamel, am = ameloblast layer, ep = enamel pulp. Bars = 50µm.
Santos et al., 27
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

ACKNOWLEDGEMENTS
We are greatly indebted to J. Martínez for excellent animal care, I. de los Santos for
assistance in histology preparations, and S. Moreno for help in photography. This work
was partially funded by grants PB94-1230 from the Spanish MCYT and 08.1/0054/2001.1
from the CAM.
Santos et al., 28
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Mirentxu Santos, Ana Bravo, Ceferino Lopez, Jesus M. Paramio and Jose L. Jorcanoepidermal keratin K10 in transgenic mice
Severe abnormalities in the oral mucosa induced by suprabasal expression of
published online July 15, 2002J. Biol. Chem.
10.1074/jbc.M205143200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on August 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from