Embed Size (px)
Citation preview

lable at ScienceDirect
Estuarine, Coastal and Shelf Science 79 (2008) 701–706
Contents lists avai
Estuarine, Coastal and Shelf Science
journal homepage: www.elsevier .com/locate/ecss
Seasonal variability of leaf litter removal by crabs in a Kandelia candelmangrove forest in Jiulongjiang Estuary, China
G.C. Chen, Y. Ye*, C.Y. LuState Key Laboratory of Marine Environmental Science, Xiamen University, Simingnan Road 422, Xiamen, Fujian 361005, China
a r t i c l e i n f o
Article history:Received 22 February 2008Accepted 18 June 2008Available online 26 June 2008
Keywords:Kandelia candelmangrovescrableaf litterremovaltemperature
* Corresponding author.E-mail address: [email protected] (Y. Ye).
0272-7714/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.ecss.2008.06.012
a b s t r a c t
The seasonal variability of leaf litter removal by crabs was observed from May 2006 to April 2007 ina Kandelia candel mangrove forest in Jiulongjiang Estuary, China. Daily average quantities of leaf fallranged 0.85–3.86 gDW m�2 d�1, with high values in May, August, October and November. The whole-year’s leaf fall was 6.48 t ha�1 yr�1 (1.81 gDW m�2 d�1). The standing stock of leaf litter on the forest floorwas 7.78 gDW m�2 averaged from the whole year’s data, with the lowest value in December (1.23 gDW m�2)and the highest in April (16.18 gDW m�2). Annually averaged removal (consumption on mangrovefloorþ burial in burrows) rate of leaf litter by crabs was 0.59 gDW m�2 d�1. High seasonal variability wasobserved in the removal rates of leaf litter by crabs. Removal rates in the winter months (December,January and February) were 0.07–0.09 gDW m�2 d�1, much lower than those in other months with valuesof 0.59–1.18 gDW m�2 d�1. Annually averaged percentage of leaf fall removed by crabs was 33%, with thehighest values in September (reached 76%) and the lowest values in winter months. Of leaf litter removedby crabs, a large proportion was buried by crabs, and only 12% was consumed by crabs on the forest floor.Leaf litter removal rate, consumption rate on the forest floor, percentages of leaf fall and standing stockremoved on the forest floor were significantly positively correlated with air temperature, indicating thatleaf removal ability by crabs was higher in warm months than in cold months.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Litter fall is one of the important components of mangroveprimary production, especially in view of its contribution to theestuarine system (Hossain et al., 2007). Although litter fall is notequivalent to total net primary production, it is important to assessthe productivity of a mangrove ecosystem and its relevance to foodwebs in coastal environments (Dawes et al., 1999; Lee, 1999). Directgrazing of leaf litter by herbivorous crabs is an important trophicpathway in mangrove forests, preventing tidal export of valuabledetritus from the mangrove ecosystem (Robertson, 1986; Lee, 1998;Nordhaus et al., 2006). Crabs also initiate and enhance the break-down of mangrove detritus and recycling of nutrients (Thongthamand Kristensen, 2005).
Leaf litter removal activity of herbivorous sesarmid crabs hasbeen studied frequently in the past (e.g. Robertson, 1986; Camilleri,1989; Emmerson and McGwynne, 1992; Micheli, 1993; Ashton,2002; Macintosh et al., 2002). Sesarmid crabs are important for theecological functions of mangrove ecosystems because they con-sume a large proportion of annual primary production (Micheli
All rights reserved.
et al., 1991; Lee, 1998; Nordhaus et al., 2006). For most of thesestudies, however, estimations of leaf litter removal by crabs werebased on short-term experiments in the field or laboratory.
The role of crabs in mangrove litter removal and processing maybe different in different areas (Micheli et al., 1991; Emmerson andMcGwynne, 1992; Slim et al., 1997). Most studies on leaf litterremoval by crabs have been conducted in tropical environmentsand only few have examined sub-tropical mangrove forests (e.g.Robertson, 1986; Camilleri, 1989; Emmerson and McGwynne, 1992;Micheli, 1993; Ashton, 2002; Imgraben and Dittmann, 2007;Mfilinge and Tsuchiya, 2008). Compared with their tropical coun-terparts, sub-tropical mangroves are subjected to large seasonalvariations in weather. Air temperatures in mangrove areas in Chinasometimes fall below 10 �C in winter (Lu and Lin, 1988). Lowtemperatures slow down important ecological processes such aslitter production (Mfilinge et al., 2005).
Information on feeding ecology of mangrove crabs in China islimited and largely descriptive. Kandelia candel is a mangrovespecies which is widely distributed in China and is commonly usedin mangrove restoration (Chen et al., 2007), but little is knownabout leaf removal of this species by herbivorous crabs. In K. candelforests in Jiulongjiang Estuary, one typical and well-managedmangrove area in China, dynamics of litter fall has been observedfor over 10 years and leaf decomposition is well documented

G.C. Chen et al. / Estuarine, Coastal and Shelf Science 79 (2008) 701–706702
(Zheng et al., 2000), but leaf litter removal by crabs has not beenquantified, resulting in an incomplete understanding of food websand biogeochemical cycles. In Jiulongjiang Estuary, two sesarmidcrab species, Sesarma plicata and Sesarma biden, are common in K.candel forests, implying a significant contribution to the mangroveleaf litter processes (Chen et al., 2007). Based on one whole year’sobservations, this study aims to investigate seasonal variability ofleaf litter removal by crabs in a K. candel forest in JiulongjiangEstuary.
2. Methods
2.1. Study area
Field experiments were carried out from May 2006 to April 2007in a mangrove forest (24�240N, 117�550E) in Jiulongjiang Estuarynear Caoputou Village, Longhai County, Zhangzhou City, FujianProvince, China (Fig. 1). This area is sub-tropical, with most of theannual rainfall (1284 mm) derived from summer typhoons and theannual mean air temperature of this area is 21.0 �C, averaged over20 years’ data from the local meteorological station. The meansalinity (2004–2005) of open water adjacent to the forest is 17.1 ppt(Chen and Ye, 2008). Tides are semi-diurnal with an average rangeof 4 m.
The Kandelia candel mangrove forest at Caoputou was re-habilitated in the intertidal zone in 1962. At present, adult trees are5–6 m high, with diameter-at-breast-height of 7–8 cm and densityof 21,600 ind ha�1. The lowest branch heights of trees are about2 m. The width (from river edge to bank edge) of the forest is about50 m with about 40 m even floor and the forest sea edge and landedge are about 5 m, respectively. The forest is about 1500 m long,with the downriver forest edge adjacent to a 500 m2 non-vegetatedflat at the same tidal elevation as the forest. The sediment is neutral,with pH values of about 7.1, and the content of silt and clay insediment is higher than 95% (Chen and Ye, 2008). In 1 month, theforest was inundated by high tides for only 6–8 days.
Air temperature and rainfall data during this study wereobtained from the local weather bureau of Longhai County about17 km from the mangrove site (Table 1). The highest monthly meantemperature was recorded in July and August (>28 �C) and thelowest in January (14.5 �C). The highest monthly rainfall was601.2 mm in May, resulted from the typhoon Chanchu. Rainfall in
Fig. 1. Map of the study area show
July was also high. Though another typhoon Saomai influencedZhangzhou in August, no significant rainfall was recorded. Rainfallin winter was low.
2.2. Leaf fall production
Litter fall samples were collected using metal framed litter traps(70 cm� 70 cm surface area, 30 cm depth) with 2 mm mesh. Fivetraps were placed in the middle zone (about 20 m wide) of theforest. Care was taken to place traps randomly under trees withsimilar height. The traps were tied to tree branches and suspended1.5 m above the sediment, which was above the maximum tidelevel. Trap contents were collected monthly and sorted into thecategories of leaves, wood, propagules, then dried at 60 �C toconstant weight.
2.3. Standing stock of leaf litter on the forest floor
Standing stock of leaf litter on the forest floor was determinedmonthly on the same date when the leaf litter removal experimentwas carried out. Five plots, each 1 m2 in size and about 3 m fromeach other, were randomly selected within the middle zone of theforest. Leaf litter on the forest floor was collected, washed and driedat 60 �C to constant weight for biomass measurements.
2.4. Leaf litter removal experiment
In each month, leaf removal experiments were carried out onone neap tide day when the forest floor was not flooded during thewhole day, and one flooded day when the forest floor was floodedby high tides. On each experimental date, five plots, each about1 m2 and at 3 m distance from each other, were randomly set up inthe middle zone of the mangrove forest. Leaf litter in the plots wasremoved to eliminate the interference by non-target leaves. Five tofifty pre-weighed newly fallen yellow leaves were laid on one plot,according to its standing stock of leaf number. Water content inthese laid leaves was estimated with other 20 newly fallen yellowleaves. To quantify leaves buried in crab burrows and grazed onforest floor by crabs, leaf petioles were bound to a 1 m long nylonline tied to one prop root of Kandelia candel. In addition, triplicatemesh bags, each containing three newly fallen yellow leaves,inaccessible to crabs were placed near the plots to determine leaf
ing the Jiulongjiang Estuary.
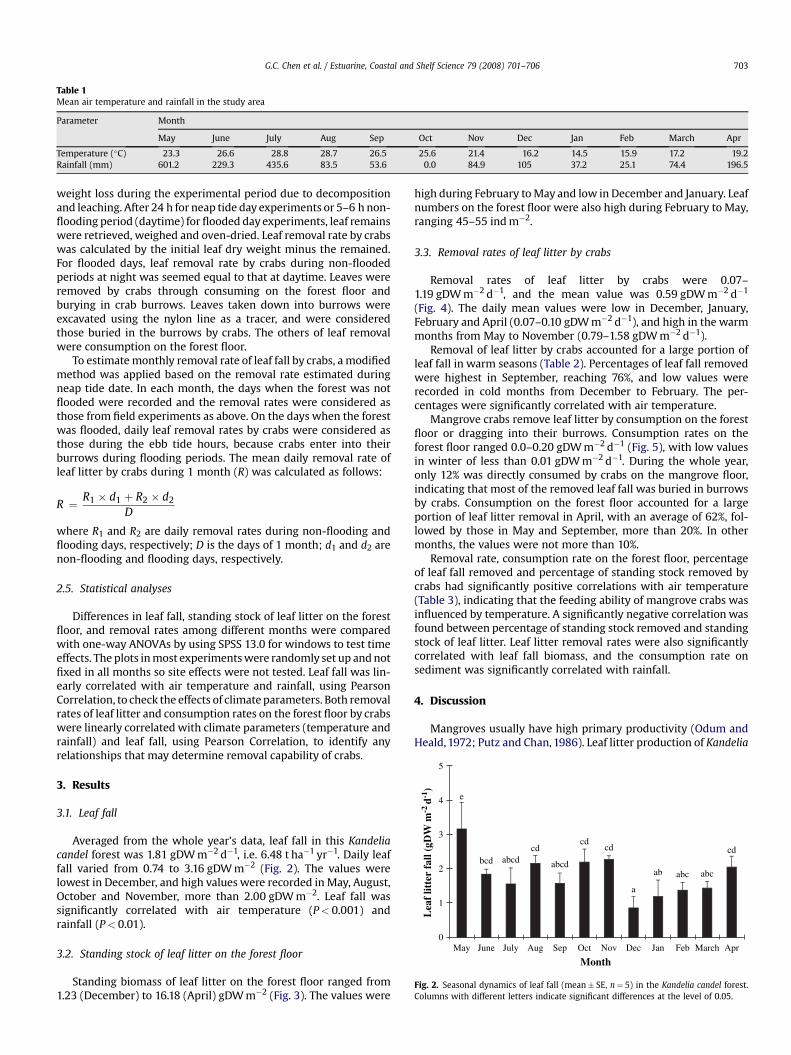
Table 1Mean air temperature and rainfall in the study area
Parameter Month
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Temperature (�C) 23.3 26.6 28.8 28.7 26.5 25.6 21.4 16.2 14.5 15.9 17.2 19.2Rainfall (mm) 601.2 229.3 435.6 83.5 53.6 0.0 84.9 105 37.2 25.1 74.4 196.5
cd
abcabcab
a
cdcd
abcd
cdabcdbcd
e
0
1
2
3
4
5
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Month
Lea
f lit
ter
fall
(gD
W m
-2 d-1
)
Fig. 2. Seasonal dynamics of leaf fall (mean� SE, n¼ 5) in the Kandelia candel forest.Columns with different letters indicate significant differences at the level of 0.05.
G.C. Chen et al. / Estuarine, Coastal and Shelf Science 79 (2008) 701–706 703
weight loss during the experimental period due to decompositionand leaching. After 24 h for neap tide day experiments or 5–6 h non-flooding period (daytime) for flooded day experiments, leaf remainswere retrieved, weighed and oven-dried. Leaf removal rate by crabswas calculated by the initial leaf dry weight minus the remained.For flooded days, leaf removal rate by crabs during non-floodedperiods at night was seemed equal to that at daytime. Leaves wereremoved by crabs through consuming on the forest floor andburying in crab burrows. Leaves taken down into burrows wereexcavated using the nylon line as a tracer, and were consideredthose buried in the burrows by crabs. The others of leaf removalwere consumption on the forest floor.
To estimate monthly removal rate of leaf fall by crabs, a modifiedmethod was applied based on the removal rate estimated duringneap tide date. In each month, the days when the forest was notflooded were recorded and the removal rates were considered asthose from field experiments as above. On the days when the forestwas flooded, daily leaf removal rates by crabs were considered asthose during the ebb tide hours, because crabs enter into theirburrows during flooding periods. The mean daily removal rate ofleaf litter by crabs during 1 month (R) was calculated as follows:
R ¼ R1 � d1 þ R2 � d2
D
where R1 and R2 are daily removal rates during non-flooding andflooding days, respectively; D is the days of 1 month; d1 and d2 arenon-flooding and flooding days, respectively.
2.5. Statistical analyses
Differences in leaf fall, standing stock of leaf litter on the forestfloor, and removal rates among different months were comparedwith one-way ANOVAs by using SPSS 13.0 for windows to test timeeffects. The plots in most experiments were randomly set up and notfixed in all months so site effects were not tested. Leaf fall was lin-early correlated with air temperature and rainfall, using PearsonCorrelation, to check the effects of climate parameters. Both removalrates of leaf litter and consumption rates on the forest floor by crabswere linearly correlated with climate parameters (temperature andrainfall) and leaf fall, using Pearson Correlation, to identify anyrelationships that may determine removal capability of crabs.
3. Results
3.1. Leaf fall
Averaged from the whole year’s data, leaf fall in this Kandeliacandel forest was 1.81 gDW m�2 d�1, i.e. 6.48 t ha�1 yr�1. Daily leaffall varied from 0.74 to 3.16 gDW m�2 (Fig. 2). The values werelowest in December, and high values were recorded in May, August,October and November, more than 2.00 gDW m�2. Leaf fall wassignificantly correlated with air temperature (P< 0.001) andrainfall (P< 0.01).
3.2. Standing stock of leaf litter on the forest floor
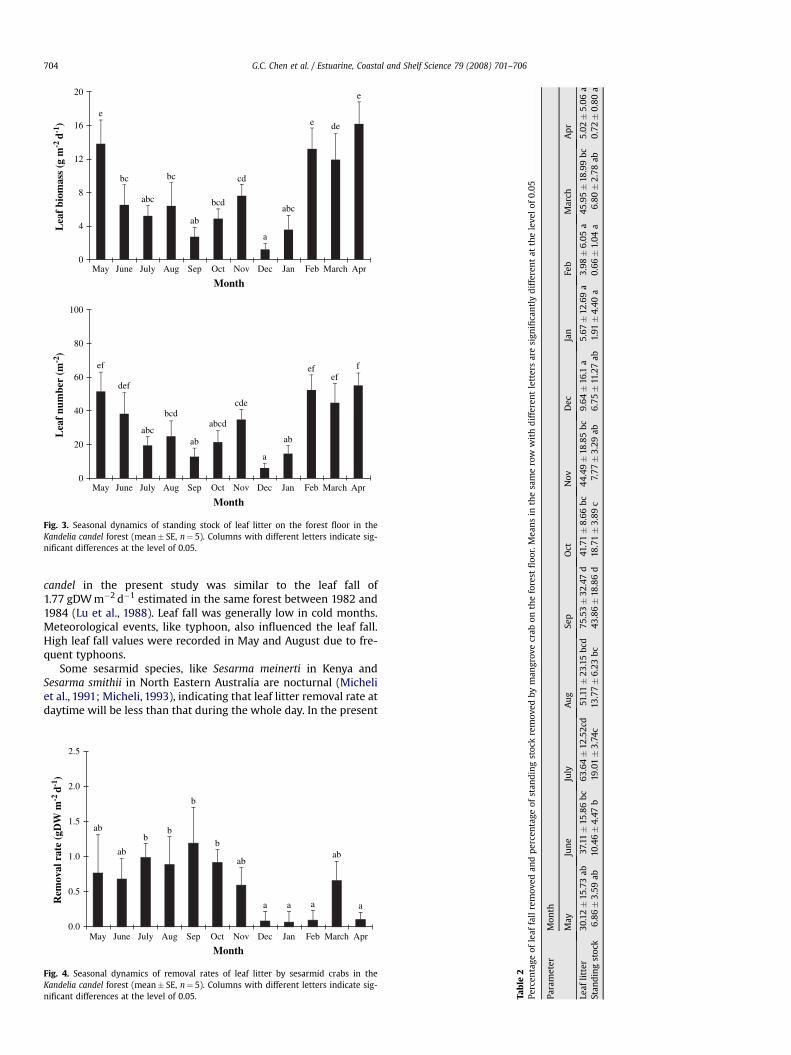
Standing biomass of leaf litter on the forest floor ranged from1.23 (December) to 16.18 (April) gDW m�2 (Fig. 3). The values were
high during February to May and low in December and January. Leafnumbers on the forest floor were also high during February to May,ranging 45–55 ind m�2.
3.3. Removal rates of leaf litter by crabs
Removal rates of leaf litter by crabs were 0.07–1.19 gDW m�2 d�1, and the mean value was 0.59 gDW m�2 d�1
(Fig. 4). The daily mean values were low in December, January,February and April (0.07–0.10 gDW m�2 d�1), and high in the warmmonths from May to November (0.79–1.58 gDW m�2 d�1).
Removal of leaf litter by crabs accounted for a large portion ofleaf fall in warm seasons (Table 2). Percentages of leaf fall removedwere highest in September, reaching 76%, and low values wererecorded in cold months from December to February. The per-centages were significantly correlated with air temperature.
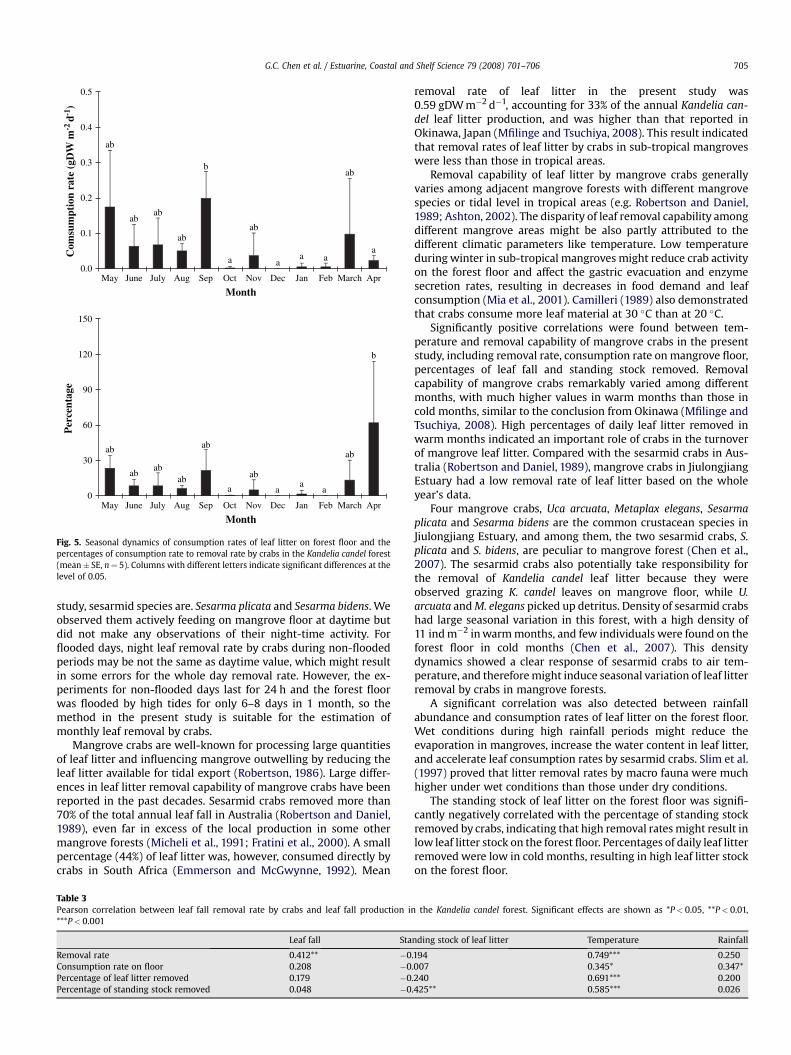
Mangrove crabs remove leaf litter by consumption on the forestfloor or dragging into their burrows. Consumption rates on theforest floor ranged 0.0–0.20 gDW m�2 d�1 (Fig. 5), with low valuesin winter of less than 0.01 gDW m�2 d�1. During the whole year,only 12% was directly consumed by crabs on the mangrove floor,indicating that most of the removed leaf fall was buried in burrowsby crabs. Consumption on the forest floor accounted for a largeportion of leaf litter removal in April, with an average of 62%, fol-lowed by those in May and September, more than 20%. In othermonths, the values were not more than 10%.
Removal rate, consumption rate on the forest floor, percentageof leaf fall removed and percentage of standing stock removed bycrabs had significantly positive correlations with air temperature(Table 3), indicating that the feeding ability of mangrove crabs wasinfluenced by temperature. A significantly negative correlation wasfound between percentage of standing stock removed and standingstock of leaf litter. Leaf litter removal rates were also significantlycorrelated with leaf fall biomass, and the consumption rate onsediment was significantly correlated with rainfall.
4. Discussion
Mangroves usually have high primary productivity (Odum andHeald, 1972; Putz and Chan, 1986). Leaf litter production of Kandelia
e
dee
abc
a
cd
bcd
ab
bc
abc
bc
e
0
4
8
12
16
20
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Month
Lea
f bi
omas
s (g
m-2
d-1)
ef
def
abc
bcd
ab
abcd
cde
a
ab
efef
f
0
20
40
60
80
100
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Month
Lea
f nu
mbe
r (m
-2)
Fig. 3. Seasonal dynamics of standing stock of leaf litter on the forest floor in theKandelia candel forest (mean� SE, n¼ 5). Columns with different letters indicate sig-nificant differences at the level of 0.05.
oved
bym
angr
ove
crab
onth
efo
rest
floo
r.M
ean
sin
the
sam
ero
ww
ith
dif
fere
nt
lett
ers
are
sign
ifica
ntl
yd
iffe
ren
tat
the
leve
lof
0.0
5
Au
gSe
pO
ctN
ovD
ecJa
nFe
bM
arch
Ap
r
51.1
1�
23
.15
bcd
75
.53�
32
.47
d41
.71�
8.6
6bc
44
.49�
18.8
5b
c9
.64�
16.1
a5
.67�
12.6
9a
3.9
8�
6.0
5a
45
.95�
18.9
9b
c5
.02�
5.0
6a
13.7
7�
6.2
3b
c4
3.8
6�
18.8
6d
18.7
1�
3.8
9c
7.7
7�
3.2
9ab
6.7
5�
11.2
7ab
1.91�
4.4
0a
0.6
6�
1.0
4a
6.8
0�
2.7
8ab
0.7
2�
0.8
0a
G.C. Chen et al. / Estuarine, Coastal and Shelf Science 79 (2008) 701–706704
candel in the present study was similar to the leaf fall of1.77 gDW m�2 d�1 estimated in the same forest between 1982 and1984 (Lu et al., 1988). Leaf fall was generally low in cold months.Meteorological events, like typhoon, also influenced the leaf fall.High leaf fall values were recorded in May and August due to fre-quent typhoons.
Some sesarmid species, like Sesarma meinerti in Kenya andSesarma smithii in North Eastern Australia are nocturnal (Micheliet al., 1991; Micheli, 1993), indicating that leaf litter removal rate atdaytime will be less than that during the whole day. In the present
a
ab
aaa
ab
b
b
bb
ab
ab
0.0
0.5
1.0
1.5
2.0
2.5
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Month
Rem
oval
rat
e (g
DW
m-2
d-1)
Fig. 4. Seasonal dynamics of removal rates of leaf litter by sesarmid crabs in theKandelia candel forest (mean� SE, n¼ 5). Columns with different letters indicate sig-nificant differences at the level of 0.05. Ta
ble
2Pe
rcen
tage
ofle
affa
llre
mov
edan
dp
erce
nta
geof
stan
din
gst
ock
rem
Para
met
erM
onth
May
Jun
eJu
ly
Leaf
litt
er3
0.1
2�
15.7
3ab
37.1
1�
15.8
6b
c6
3.6
4�
12.5
2cd
Stan
din
gst
ock
6.8
6�
3.5
9ab
10.4
6�
4.4
7b
19.0
1�
3.7
4c
ab
abab
ab
b
a
ab
aa a
ab
a
0.0
0.1
0.2
0.3
0.4
0.5
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Month
Com
sum
ptio
n ra
te (
gDW
m-2
d-1)
ab
abab
ab
ab
a
ab
aa
a
ab
b
0
30
60
90
120
150
May June July Aug Sep Oct Nov Dec Jan Feb March Apr
Month
Per
cent
age
Fig. 5. Seasonal dynamics of consumption rates of leaf litter on forest floor and thepercentages of consumption rate to removal rate by crabs in the Kandelia candel forest(mean� SE, n¼ 5). Columns with different letters indicate significant differences at thelevel of 0.05.
G.C. Chen et al. / Estuarine, Coastal and Shelf Science 79 (2008) 701–706 705
study, sesarmid species are. Sesarma plicata and Sesarma bidens. Weobserved them actively feeding on mangrove floor at daytime butdid not make any observations of their night-time activity. Forflooded days, night leaf removal rate by crabs during non-floodedperiods may be not the same as daytime value, which might resultin some errors for the whole day removal rate. However, the ex-periments for non-flooded days last for 24 h and the forest floorwas flooded by high tides for only 6–8 days in 1 month, so themethod in the present study is suitable for the estimation ofmonthly leaf removal by crabs.
Mangrove crabs are well-known for processing large quantitiesof leaf litter and influencing mangrove outwelling by reducing theleaf litter available for tidal export (Robertson, 1986). Large differ-ences in leaf litter removal capability of mangrove crabs have beenreported in the past decades. Sesarmid crabs removed more than70% of the total annual leaf fall in Australia (Robertson and Daniel,1989), even far in excess of the local production in some othermangrove forests (Micheli et al., 1991; Fratini et al., 2000). A smallpercentage (44%) of leaf litter was, however, consumed directly bycrabs in South Africa (Emmerson and McGwynne, 1992). Mean
Table 3Pearson correlation between leaf fall removal rate by crabs and leaf fall production in***P< 0.001
Leaf fall Sta
Removal rate 0.412** �0Consumption rate on floor 0.208 �0Percentage of leaf litter removed 0.179 �0Percentage of standing stock removed 0.048 �0
removal rate of leaf litter in the present study was0.59 gDW m�2 d�1, accounting for 33% of the annual Kandelia can-del leaf litter production, and was higher than that reported inOkinawa, Japan (Mfilinge and Tsuchiya, 2008). This result indicatedthat removal rates of leaf litter by crabs in sub-tropical mangroveswere less than those in tropical areas.
Removal capability of leaf litter by mangrove crabs generallyvaries among adjacent mangrove forests with different mangrovespecies or tidal level in tropical areas (e.g. Robertson and Daniel,1989; Ashton, 2002). The disparity of leaf removal capability amongdifferent mangrove areas might be also partly attributed to thedifferent climatic parameters like temperature. Low temperatureduring winter in sub-tropical mangroves might reduce crab activityon the forest floor and affect the gastric evacuation and enzymesecretion rates, resulting in decreases in food demand and leafconsumption (Mia et al., 2001). Camilleri (1989) also demonstratedthat crabs consume more leaf material at 30 �C than at 20 �C.
Significantly positive correlations were found between tem-perature and removal capability of mangrove crabs in the presentstudy, including removal rate, consumption rate on mangrove floor,percentages of leaf fall and standing stock removed. Removalcapability of mangrove crabs remarkably varied among differentmonths, with much higher values in warm months than those incold months, similar to the conclusion from Okinawa (Mfilinge andTsuchiya, 2008). High percentages of daily leaf litter removed inwarm months indicated an important role of crabs in the turnoverof mangrove leaf litter. Compared with the sesarmid crabs in Aus-tralia (Robertson and Daniel, 1989), mangrove crabs in JiulongjiangEstuary had a low removal rate of leaf litter based on the wholeyear’s data.
Four mangrove crabs, Uca arcuata, Metaplax elegans, Sesarmaplicata and Sesarma bidens are the common crustacean species inJiulongjiang Estuary, and among them, the two sesarmid crabs, S.plicata and S. bidens, are peculiar to mangrove forest (Chen et al.,2007). The sesarmid crabs also potentially take responsibility forthe removal of Kandelia candel leaf litter because they wereobserved grazing K. candel leaves on mangrove floor, while U.arcuata and M. elegans picked up detritus. Density of sesarmid crabshad large seasonal variation in this forest, with a high density of11 ind m�2 in warm months, and few individuals were found on theforest floor in cold months (Chen et al., 2007). This densitydynamics showed a clear response of sesarmid crabs to air tem-perature, and therefore might induce seasonal variation of leaf litterremoval by crabs in mangrove forests.
A significant correlation was also detected between rainfallabundance and consumption rates of leaf litter on the forest floor.Wet conditions during high rainfall periods might reduce theevaporation in mangroves, increase the water content in leaf litter,and accelerate leaf consumption rates by sesarmid crabs. Slim et al.(1997) proved that litter removal rates by macro fauna were muchhigher under wet conditions than those under dry conditions.
The standing stock of leaf litter on the forest floor was signifi-cantly negatively correlated with the percentage of standing stockremoved by crabs, indicating that high removal rates might result inlow leaf litter stock on the forest floor. Percentages of daily leaf litterremoved were low in cold months, resulting in high leaf litter stockon the forest floor.
the Kandelia candel forest. Significant effects are shown as *P< 0.05, **P< 0.01,
nding stock of leaf litter Temperature Rainfall
.194 0.749*** 0.250
.007 0.345* 0.347*
.240 0.691*** 0.200
.425** 0.585*** 0.026

G.C. Chen et al. / Estuarine, Coastal and Shelf Science 79 (2008) 701–706706
Stocking leaves in crab burrows and then consuming them is thekey manner in which leaf litter is removed from mangrove floor butretains in the ecosystem. Most of leaf litter materials wereindirectly removed in this manner and less than 25% was directlyconsumed on the forest floor. Leaves dragged into burrows werefound torn into fragments before being consumed. In this way,mangrove crabs might enhance leaf nutrition quality (Giddins et al.,1986), prevent leaf litter from being removed by tidal flushing(Robertson, 1986) and avoid competition and predation when foodor time available for collecting food is limited, and predatorabundance is high (Ashton, 2002).
In conclusion, mangrove crabs in Jiulongjiang Estuary playa significant role in the turnover of Kandelia candel leaf fall. Removalcapability of crabs significantly varied during the whole year, withmuch higher rates in warm months than in cold months. Consid-ering this great temporal variation, when estimating and evaluatingthe removal capability of mangrove crabs, we show the need forlong-time observations, especially in sub-tropical mangroves.Simple studies and straight comparisons of removal rates based ona short time observation may cause erroneous results.
Acknowledgements
This work was supported by Programs for National NaturalScience Foundation of China (40476040) and New CenturyExcellent Talents in University (NCET). We thank F.Z. Zheng, H.Y. Hu,C.Q. Chao, J.S. Guo, Z.Q. Yin, Y.L. Xu, L. Zhang, X.X. Chen and Q.F. Hufor their dedicated work in the field and/or laboratory. We alsothank Y.X. Xu of Longhai Weather Bureau for weather data.
References
Ashton, E.C., 2002. Mangrove sesarmid crab feeding experiments in PeninsularMalaysia. Journal of Experimental Marine Biology and Ecology 273, 97–119.
Camilleri, J.C., 1989. Leaf choice by crustaceans in a mangrove forest in Queensland.Marine Biology 102, 453–459.
Chen, G.C., Ye, Y., Lu, C.Y., 2007. Changes of macro-benthic faunal community withstand age of rehabilitated Kandelia candel mangrove in Jiulongjiang Estuary,China. Ecological Engineering 31, 215–224.
Chen, G.C., Ye, Y., 2008. Leaf consumption by Sesarma plicata in a mangrove forest atJiulongjiang Estuary, China. Marine Biology 154, 997–1007.
Dawes, C., Siar, K., Marlett, D., 1999. Mangrove structure, litter and macroalgalproductivity in a northern-most forest of Florida. Mangroves and Salt Marshes3, 259–267.
Emmerson, W.D., McGwynne, L.E., 1992. Feeding and assimilation of mangroveleaves by the crab Sesarma meineri de Man in relation to leaf-litter productionin Mgazana, a warm-temperate southern African mangrove swamp. Journal ofExperimental Marine Biology and Ecology 157, 41–53.
Fratini, S., Cannicci, S., Vannini, M., 2000. Competition and interaction betweenNeosarmatium smithi (Crustacea: Grapsidae) and Terebralia palustris (Mollusca:Gastropoda) in a Kenyan mangrove. Marine Biology 137, 309–316.
Giddins, R.L., Lucas, J.S., Neilson, M.J., Richards, G.N., 1986. Feeding ecology of themangrove crab Neosarmatium smithi (Crustacea: Decapoda: Sesarmidae).Marine Ecology Progress Series 33, 147–155.
Hossain, M., Othman, S., Sidik, J., Misri Kusnan, B., 2007. Net primary productivity ofBruguiera parviflora (Wight & Arn.) dominated mangrove forest at KualaSelangor, Malaysia. Forest Ecology and Management 255, 179–182.
Imgraben, S., Dittmann, S., 2007. Leaf dynamics and litter consumption in twotemperate South Australian mangrove forest. Journal of Sea Research 59, 83–93.
Lee, S.Y., 1998. Ecological role of grapsid crabs in mangrove ecosystems: a review.Marine and Freshwater Research 49, 335–343.
Lee, S.Y., 1999. Tropical mangrove ecology: physical and biotic factors influencingecosystem structure and function. Australian Journal of Ecology 24, 355–366.
Lu, C.Y., Lin, P., 1988. Litter leaf decomposition of two species of mangrove. Journalof Xiamen University (Nature Science) 27, 679–683.
Lu, C.Y., Zheng, F.Z., Lin, P., 1988. Study of litter fall production of Kandelia candelmangrove community in estuary. Journal of Xiamen University (Nature Science)27, 459–463.
Macintosh, D.J., Ashton, E.C., Havanon, S., 2002. Mangrove rehabilitation andintertidal biodiversity: a study in the Ranong mangrove ecosystem, Thailand.Estuarine, Coastal and Shelf Science 55, 331–345.
Mia, Y., Shokita, S., Watanabe, S., 2001. Stomach content of two grapsid crabs, Heliceformosensis and Helice leachi. Fisheries Science 67, 173–175.
Micheli, F., 1993. Feeding ecology of mangrove crabs in North Eastern Australia:mangrove litter consumption by Sesarma messa and Sesarma smithii. Journal ofExperimental Marine Biology and Ecology 171, 165–186.
Micheli, F., Gherardi, F., Vannini, M., 1991. Feeding and burrow ecology of two EastAfrican mangrove crabs. Marine Biology 111, 247–254.
Mfilinge, P.L., Meziane, T., Bachok, Z., Tsuchiya, M., 2005. Litter dynamics and par-ticulate organic matter outwelling from a subtropical mangrove in OkinawaIsland, South Japan. Estuarine, Coastal and Shelf Science 63, 301–313.
Mfilinge, P.L., Tsuchiya, M., 2008. Effect of temperature on leaf litter consumption bygrapsid crabs in a subtropical mangrove (Okinawa, Japan). Journal of Sea Re-search 59, 94–102.
Nordhaus, I., Wolff, M., Diele, K., 2006. Litter processing and population food intakeof the mangrove crab Ucides cordatus in a high intertidal forest in northernBrazil. Estuarine, Coastal and Shelf Science 67, 239–250.
Odum, W.E., Heald, E.J., 1972. Trophic analyses of an estuarine mangrove commu-nity. Bulletin of Marine Science 22, 671–738.
Putz, F.E., Chan, H.T., 1986. Tree growth, dynamic, and productivity in a matureforest in Malaysia. Forest Ecology and Management 17, 211–230.
Robertson, A.I., 1986. Leaf-burying crabs: their influence on energy flow and exportfrom mixed mangrove forests (Rhizophora spp.) in northeastern Australia.Journal of Experimental Marine Biology and Ecology 102, 237–248.
Robertson, A.I., Daniel, P.A., 1989. The influence of crabs on litter processing in highintertidal mangrove forests in tropical Australia. Oecologia 78, 191–198.
Slim, F.J., Hemminga, M.A., Ochieng, C., Jannink, N.T., Corcheret de la Moriniere, E.,van der Velde, G., 1997. Leaf litter removal by the snail Terebralia palustris(Linnaeus) and sesarmid crabs in an East African mangrove forest (Gazi Bay,Kenya). Journal of Experimental Marine Biology and Ecology 215, 35–48.
Thongtham, N., Kristensen, E., 2005. Carbon and nitrogen balance of leaf-eatingsesarmid crabs (Neoepisesarma versicolor) offered different food sources. Estu-arine, Coastal and Shelf Science 65, 213–222.
Zheng, F.Z., Lu, C.Y., Zheng, W.J., Lin, P., 2000. Seasonal dynamics of litter fall andenergy flow through the leaf litter of Kandelia candel mangrove in JiulongjiangEstuary, Fujian province, China. Journal of Xiamen University (Nature Science)39, 693–698.





























