Embed Size (px)
Citation preview

Bulow, E.S. and Ferdinand, T.J. 2013
08 Fall
The Effect of Consumptive Waste on Mangrove Functionality: A Comparative Analysis
M c G i l l U n i v e r s i t y P a n a m a F i e l d S t u d y S e m e s t e r C e n t r o d e I n c i d e n c i a A m b i e n t a l ( C I A M )

2 | P a g e
CIAM
Centro de Incidencia Ambiental
Urb. Los Ángeles, Calle Los Periodistas,
Casa G-14 Planta Alta.
Teléfono: (507) 236-0866 / (507) 236-0868
Tele-fax: (507) 236-0872
Naos Marine Laboratories
Bldg. 356, Amador Causeway, Naos Island
Panama, Rep. de Panamà
McGill University
845 Sherbrooke Street West, Montreal
QC, Canada
H3A 0G4
(514) 398-4455

3 | P a g e
Table of Contents
Host Information
Background 5
Contact Information 6
Project Personnel 6
Project Information
Background and Basis 7
The Mangrove 7
Mangroves in Panama 8
Crabs: Mangrove Engineers 11
Mangroves and Salinity 12
The Black Mangrove Ecosystem 13
Waste Management in Panama City 14
Juan Diaz: Study Site #1 15
Punta Chame: Study Site #2 17
The Question 18
Methodology 19
Site Locations 19
Sampling Crabs, Garbage and Trees 19
Sampling and Testing Salinity 20
Results 21
Discussion 25
Limitations and Obstacles Encountered 28

4 | P a g e
Looking Forward 28
Logistical Information
Work Log 29
Financial Expenditures 29
Official Recognition Requests 29
Additional Acknowledgments 30
Bibliography 31

5 | P a g e
Host Information
Background
CIAM, el Centro de Incidencia Ambiental or the Environmental Advocacy
Center, is a non-profit organization that is dedicated to legally defending the lands, rivers,
oceans and cultural heritages of Panama. Before the creation of CIAM in 2007 Panama
lacked a major environmental group containing activists that initiated legislative
processes. The Board of Directors consists of members independently recognized for
their expertise in their respective fields and their resolute integrity and commitment to
environmental and human rights. Since its creation, the NGO has had enormous
successes in such areas as: climate change, open-pit mining, hydroelectric plants and,
more recently, mangrove and wetland protection.
In all of their endeavors CIAM aims to provide citizens with public information,
promote and facilitate civic participation, advocate sound environmental public policy
and legislate a healthy environment as a human right.
CIAM relies on consensus building across all sectors and scales of society in
order to produce greater influential leverage and construct a productive common vision.
Besides working with individual citizens and communities, CIAM finds partnership in
ANCON, Fundación Almanaque Azul, Sociedad Audubon de Panamá, Fundación
Natura, Fundación Albatros Media, Fundación ProMar, Asociación Oceánica de Panamá
and Fundación MarViva. On an International level CIAM has made alliances with such
groups as the Environmental Law Alliance Worldwide (ELAW), the Inter-American
Association for the Defense of the Environment (AIDA), the Center for International
Environmental Law (CIEL), and Mining Watch, along with major U.S. and Canadian
universities.

6 | P a g e
Contact Information
CIAM
Centro de Incidencia Ambiental
Urb. Los Ángeles, Calle Los Periodistas,
Casa G-14 Planta Alta.
Teléfono: (507) 236-0866 / (507) 236-0868
Tele-fax: (507) 236-0872
Correo electrónico: [email protected]
Project Personnel
McGill University Staff
Tyler Ferdinand (Internship Student)
Email: [email protected]
Telephone: 6585-6552
Ella (Elise) Bulow (Internship Student)
Email: [email protected]
Roberto Ibanez (Internship Supervisor)
Victor Frankel (Internship Teaching
Assistant)
CIAM Staff
Lourdes Lozano
Private Consultant and Internship
Coordinator
Email: [email protected]
Telephone: 6676-5797
Sonia Montenegro
Deputy Director
Email: [email protected]
Telephone: 6675-8021
Tania Arosemena
Director of Legal Affairs
Email: [email protected]
Telephone: (507) 6781-4278
Nathalie Vásquez Cabrera
Project Assistant
Email: [email protected]
Telephone: 6780-8481
Gustavo Cárdenas
Universidad de Panamá Intern
Email: [email protected]

7 | P a g e
Project Information
Background and Basis
The Mangrove
Mangroves are among the most productive and biologically complex ecosystems
on earth, scattering intertidal regions within 30 degrees of the equator (Warne, n.d).
These vital ecosystems are critically important for ecological and social communities
around the world. Mangroves play an important role guarding the low-lying coastal land
by forming a protective barrier which reduces damage caused by storms, limiting wave
energy and preventing the land from being flooded and eroded. This has become much
more globally recognized after the 2004 Asian tsunami that left areas with previously
destroyed mangroves with a much higher rate of casualties and destruction than areas
with intact, healthy mangrove forests (Danielsen et al.2005; Radhika, 2006).
Fig 1 - (http://www.flmnh.ufl.edu)
Their physical presence not only serves as a barrier to protect human populations
but also provides shelter and a food resource for estuarine and coastal fishery food
chains. Many fish, shellfish, birds and other wildlife adopt mangrove ecosystems as
breeding, feeding and nursery habitats. This habitat allows the growth of numerous

8 | P a g e
seafood species that make up a significant portion of many countries economies; for
example it contributes $86 million a year to Panama’s economy (Audubon de Panamá,
n.d). Additionally, these wetlands act as an essential filter for hazardous agricultural
effluent as well as urban pollution, safeguarding marine trophic systems.
While these forests have proven to be essential to animals and humans alike, they
have been severely threatened by anthropogenic exploitations. More than 35% of the area
of mangrove forests has been destroyed in the past two decades. This is equivalent to an
annual loss of 3,000 km2 of mangrove forests along shorelines around the world since the
1980’s (EcoLogic, n.d). This alarming loss of mangrove ecosystems plays an important
role in the global climate crisis, as mangroves are one of the most carbon rich of all
tropical forest biomes. Although this particular ecosystem only accounts for 0.7% of all
tropical forests, their
degradation is responsible for
10% of the global carbon
emissions caused by
deforestation. These
ecosystems are among the
most carbon-rich forests in the
tropics not necessarily because
of the tree’s high wood
biomass, but because of the
high organic-rich soils that
account for 49-98% of the
biome’s carbon storage (Fig 2,
Donato et al., 2011).
Mangroves in Panama
Since its geography includes extensive coastlines, Panama has become an
important focus in Central America for mangrove conservation and management. The
topography, climate and varying oceanic conditions across the isthmus gives rise to a
variety of mangrove species on both the Pacific and Caribbean coasts, with the majority
Fig 2 - (Donato et al., 2011)

9 | P a g e
of mangroves found on the Pacific perimeter. Out of 172, 000 hectares of mangrove
forest in Panama, around 166,000 hectares are found on the Pacific coast. Of the thirteen
mangrove species present in the Americas, eleven species are native to Panama that fall
under the genera of Laguncularia, Avicennia, Rhizophora, or Pelliciera. The most
common species are the red and white mangroves, Rhizophora mangle y Rhizophora
racemosa respectively. Although the different species may have extremely different
phenotypes, they do share common traits: propagule dispersal by seawater, vivipary, and
their capability to easily adapt. Each mangrove has some form of a salinity filtration
system and a complex root system that allows it to survive in the intertidal zone. Some
have snorkel-like roots called pneumatophores that stick out of the mud to help them take
in air; others use prop roots or buttresses to keep their trunks upright in the soft sediments
at the tide's edge. (Warne, n.d; Betts, 2006).
Although mangroves only account for 5.6% of the total forest cover in Panama,
they are being destroyed at a distressing rate. Between the years of 1980 and 1990
Panama lost 75% of it’s mangrove forests (Ecologic, n,d). Even though some
conservation frameworks have been developed, rampant development continues to
eradicate mangroves from certain regions. Land clearing, aquaculture expansion, over-
harvesting, and private and corporate development are main drivers for mangrove
decline.
In 2003 Panama signed the Ramsar convention declaring the Panama Bay area as
a “wetland of international importance” and in 2005 the bay was declared as a “Site of
Hemispheric Importance” due to its recognition as the most important shorebird site in
Central America. Between one and two million shorebirds of over 30 different species
use Panama during their migration route. For certain bird species, as much as 50% of
their global population depend specifically on Panama Bay for their migration.
(Audubon, n.d).
On top of these significant international recognitions, the Panama Bay Wetlands,
consisting of 80,000 ha, became a National Protected Area in 2009. However, on August
16th
, 2010 Panamanian courts began annulment trials, which ended in the revocation of
the protective law. This impugnment resulted from a dispute over a matter of “prior and
informed consent” inspired by the pressure for urban and resort development (AIDA,

10 | P a g e
2011). Under pressure from urban and resort developers, the government suspended this
protected area clause in May 2012 under Article AG-0072. Not only were developers
allowed to take advantage of the protected mangroves, but the Panama’s Aquatic
Resources Authority also reduced the fees for the use of mangroves and the fines for
illegal mangrove clearance, greatly facilitating the destruction of this vital ecosystem.
The fines for illegal cutting of mangroves went from $300,000 per hectare to $40,000 per
hectare, and the fee to get a permit to legally clear cut mangroves went from $150,000
per hectare to $10,000 per hectare (Jackson, 2012). Panama has no large -scale mangrove
reforestation program and these tidal zone trees are difficult to successfully germinate
and replant. At the current state of science and given the cost of the skilled labor required,
$10,000 is not enough to restore a hectare of mangroves. Thus the concept of a developer
replacing trees that are destroyed by construction by planting trees elsewhere becomes
immaterial when corporations discuss their project logistics (Jackson, 2012). Not only
will the continued destruction of the mangroves in Panama release vast amounts of
detrimental carbon into the atmosphere, but will also have drastic deleterious effects on
numerous species that depend on the mangroves for survival. Despite the direct loss of
several at risk mangrove tree species, many globally threatened species like Crocodylus
acutus, Caretta caretta, Tapirus terrestris and Ateles geoffroyi will also be negatively
affected (Audobon, nd).
Something that has seen international attention recently is the severe flooding of
Panama’s main international airport, Tocumen. The Panama Bay mangroves are a major
shield for flooding and extreme weather events like storms and tsunamis by providing a
sink for mass amounts of incoming water. Panama’s major airport has been flooding in
the past years due to the intense destruction of mangroves in Panama Bay, and instead
putting efforts towards restoring the mangroves, the government is building a new airport
further inland.
11 | P a g e
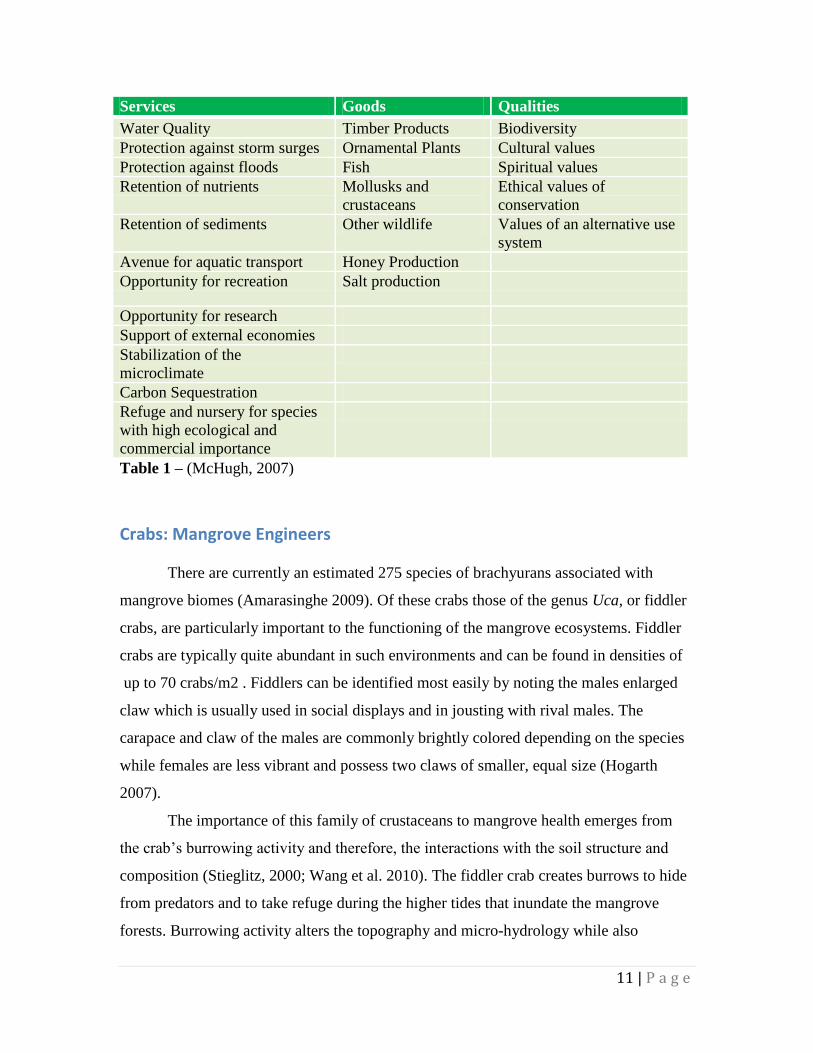
Services Goods Qualities
Water Quality Timber Products Biodiversity
Protection against storm surges Ornamental Plants Cultural values
Protection against floods Fish Spiritual values
Retention of nutrients Mollusks and
crustaceans
Ethical values of
conservation
Retention of sediments Other wildlife Values of an alternative use
system
Avenue for aquatic transport Honey Production
Opportunity for recreation Salt production
Opportunity for research
Support of external economies
Stabilization of the
microclimate
Carbon Sequestration
Refuge and nursery for species
with high ecological and
commercial importance
Table 1 – (McHugh, 2007)
Crabs: Mangrove Engineers
There are currently an estimated 275 species of brachyurans associated with
mangrove biomes (Amarasinghe 2009). Of these crabs those of the genus Uca, or fiddler
crabs, are particularly important to the functioning of the mangrove ecosystems. Fiddler
crabs are typically quite abundant in such environments and can be found in densities of
up to 70 crabs/m2 . Fiddlers can be identified most easily by noting the males enlarged
claw which is usually used in social displays and in jousting with rival males. The
carapace and claw of the males are commonly brightly colored depending on the species
while females are less vibrant and possess two claws of smaller, equal size (Hogarth
2007).
The importance of this family of crustaceans to mangrove health emerges from
the crab’s burrowing activity and therefore, the interactions with the soil structure and
composition (Stieglitz, 2000; Wang et al. 2010). The fiddler crab creates burrows to hide
from predators and to take refuge during the higher tides that inundate the mangrove
forests. Burrowing activity alters the topography and micro-hydrology while also

12 | P a g e
functioning as a system of channels that carry water, dissolved nutrients and oxygen to
the anaerobic soils around mangrove roots. This bioturbation, the process of changing the
physical, chemical and biological nature of the ecosystem via burrowing, increases the
reactive anaerobic area of the mangrove soils helping to facilitate the decomposition and
mineralization provision of nutrients to improve mangrove viability. The absence of
crabs, and therefore their burrows, could result in plant nutrient deficiencies and
unfavorable soil conditions that would reduce mangrove primary production and damage
the overall ecosystem functionality (Amarasinghe 2009).
Particularly relevant to this study, the presence of crab burrows helps to reduce
soil salinity when, during tidal events, the lining of burrows are flushed and cycled
(Amarasinghe 2009). Although these changes in groundwater and soil composition may
only represent small fluctuations in salinity, without crab burrows a chronic occurrence
of elevated salinity contrary to normal dynamics can hinder not only tree fitness but also
the survivorship of crab populations. In one study conducted in a Kenyan mangrove
ecosystem, two crab species could osmoregulate for a certain period of time in elevated
salinity conditions but could not withstand long-term hypersalinity (Gillikin et al. 2004).
Mangroves and Salinity
The uniqueness of the mangrove is especially evident when discussing its
relationship with the ocean and therefore, an existence in a terrestrially adverse
environment in which vegetation must cope with higher levels of salinity. Mangrove
species are obligated halophytes, growing in habitats with a salinity between that of
freshwater and seawater, seawater being approximately 35 g/l salt (Hogarth 2007,
Suaarez and Medina 2005). Due to these conditions there is generally an osmotic
potential of -2.5 MPa meaning that desalinated water must be taken in against this
pressure (Hogarth 2007). As salinity increases, the ability for the mangrove to counteract
this pressure becomes increasingly difficult. However, evolutionary adaptations to salty
environments have allowed mangroves to thrive along global coastlines.
The main mechanisms of managing salinity are preferential assimilation by roots,
a tolerance of high tissue salt concentrations and elimination of excess salt by secretion

13 | P a g e
(Khan and Aziz 2001; Lovelock et al. 2010). The genus Avicennia is capable of
excluding 90-97% of salt at the root surface. Like other halophytes, this is possible by
generating a negative hydrostatic pressure that overcomes the negative osmotic pressure
in the root environment allowing water to be drawn in while excluding potentially
harmful ions (Khan and Aziz 2001, Hogarth 2007). This form of salt exclusion can result
in higher salinity measurements in areas surrounding the root system (Hogarth 2007).
This can become problematic if the available soil is not cycled and redistributed, resulting
in decreased stomatal conductance, lower water potential and accumulation of inorganic
ions (Khan and Aziz 2001).
In some mangrove species, including Avicennia, sodium chloride is deposited in
the bark of stems, roots and also in senescent leaves. When salt deposits accumulate
within the tree’s leaves they are eventually shed, most likely as a method to remove salt
from metabolic tissue (Hogarth 2007). Under chronic salinity stress, leaf mortality is
accompanied by a notable decrease in leaf production rate, which in most cases can lead
to the tree’s death. The ratio of leaf production to leaf death, as well as leaf life span is
extremely important to overall plant fitness due to the leaf surface area being directly
related to the tree’s carbon, nutrient and photosynthetic economy (Suaarez 2005, Sobrado
2000).
Mangrove seedling success is also determined by certain salinity ratios, each
species thriving at a particular salinity ratio (Basak et al. 2004). Avicennia propagules
generally grow best in 50% seawater, but at later stages the optimal salinity is lower (Ball
1988a). This may result in a forest structure in which tree demography is skewed towards
younger individuals when salinity is too high for mature individuals to survive.
The Black Mangrove Ecosystem
The black mangrove, Avicennia germinans, is probably the most widespread
neotropical mangrove. Black mangrove populations are located globally, including the
eastern tropical coasts of North and South America, the Caribbean Islands and parts of
Africa (Houck & Neill, 2009; Ellison et al., 2010). It is generally found at higher tidal
elevations than Rhizophora mangle, the red mangrove, which colonizes the ocean-shore
interface. Black mangroves can only persist where there is adequate protection from

14 | P a g e
wave action, and is adapted to sandy, silty clay loam, and muck soils (NRCS, 2005). The
spatial distribution across the intertidal zone for black mangrove, red mangrove and white
mangrove (Laguncularia racemosa) suggests differential flooding tolerance among these
species (Fig 3).
Fig 3 - http://www.dcbiodata.net/explorer/info/habitats
This species is characterized by its opposite leaf pattern and leaves which are
narrow and elliptical in shape, often found encrusted with salt. The root system of
A.germinans consists of long underground cable roots that produce hundreds of thin,
upright pneumatophores that penetrate the soil´s superficial layer to allow root respiration
in anaerobic, waterlogged soils.
On average they grow to around 3-12 m in height, in areas with tidal amplitude
ranges of 2-6 meters (WWF, 2010). It is widely believed that the flowers of Avicennia
germinans are pollinated by insects, principally bees. Reproductive adaptations enable
seedlings to germinate while still attached to the parent tree, a phenomenon known as
cryptovivipary, in which the embryo emerges from the seed coat, but remains in the fruit
before falling from the parent plant. The seedlings, lima bean-shaped propagules, sprout
into 1 inch (2-3 cm), and eventually fall from the parent plant and are able, in the absence
of suitable location, to float for an indefinite period in salt water without rooting.
(FLMNH, n.d)
Waste Management in Panama City
Currently in Panama there is limited legislation in regards to waste management.
Approximately 1,300 tons of garbage is produced daily in the capital city, a number that

15 | P a g e
is rapidly rising as the population grows and tourism increases. Public media estimates
that 70% of Panamanian’s waste is collected in urban areas and moved to local dumps to
be burned. The remaining 30% is accumulated in public areas including beaches, forests
and waterways. That is approximately 390 tons of garbage a day that is discarded into
non-managed environmental systems.
In 2010 the national government established the Autoridad de Aseo Urbano y
Domiciliario (AAUD) via Act 51. This institution was designed to take charge of
administering, directing, planning, and developing a waste management framework while
investigating, inspecting and auditing institutions in relation to urban sanitation.
However, after several years there is still the lack of a provision dealing with the
treatment of waste and no prominent education campaigns aimed at redirecting
consumption trends and sparking recycling initiatives (Pereira 2012).
With the existence of the AAUD, the Ministry of Education and Health and the
Autoridad Nacional de Ambiente (ANAM) there is enough political organization to
create legislation but there appears to be no political initiative. A presentation given by
ANAM in 2007 listed problems in regards to waste management ranging from
institutional issues such as the lack of training for the municipal staff to economic issues
such as the lack of availability of loans and investment for the management sector
(Hernandez 2007).
Until the creation of legislation, perhaps fueled by a shift in the cultural paradigm,
garbage will rapidly continue to fill natural niches, negatively affecting ecosystem
functionality and services along with threatening the incredible biodiversity of the
isthmus of Panama.
Juan Diaz: Study Site #1
The Juan Diaz basin, found among the 150 km extent of wetlands that comprises
Panama Bay, has recently received a lot of attention from environmental advocacy
groups. A large portion of Panama Bay, including the bay of Juan Diaz, was part of the
territory declared a Wetland of International Importance in 2003 under the Ramsar
Convention (Ramsar, 2000) and designated a Site of Hemispheric Importance in 2005 as

16 | P a g e
the most important shorebird site in Central America (Audubon, n.d). This section was
also part of the protected area that was recently suspended and is now the location of a
series of new construction projects.
The main three developers that spurred the legal challenge to lift the protected
area status are Santa Maria Golf and Country Club, Panama Bay Development and
Compañia Lefevre, the former of which is responsible for a huge portion of mangrove
clearance in the Juan Diaz basin. In the same location Odebrecht has also built a large
water treatment facility to help mitigate Panama City’s water management issues. The
first of three installments will be operational this year, and they are expanding to three
times the current size in the coming years (Fig 4).
Fig 4 – Study Site #1 in the Juan Diaz Basin (Google Maps)
In addition to deforestation, the intact mangrove forest in the Juan Diaz basin is
facing pollution issues stemming from the inefficient waste management system in
17 | P a g e
Panama City. Scattered across the forest floor are large amounts of consumptive debris
left behind from fluctuations of the Juan Diaz river level and ocean tides. During the
rainy season, all the street garbage in the local area gets washed into the Juan Diaz river
which discharges into the basin and the mangrove forests.
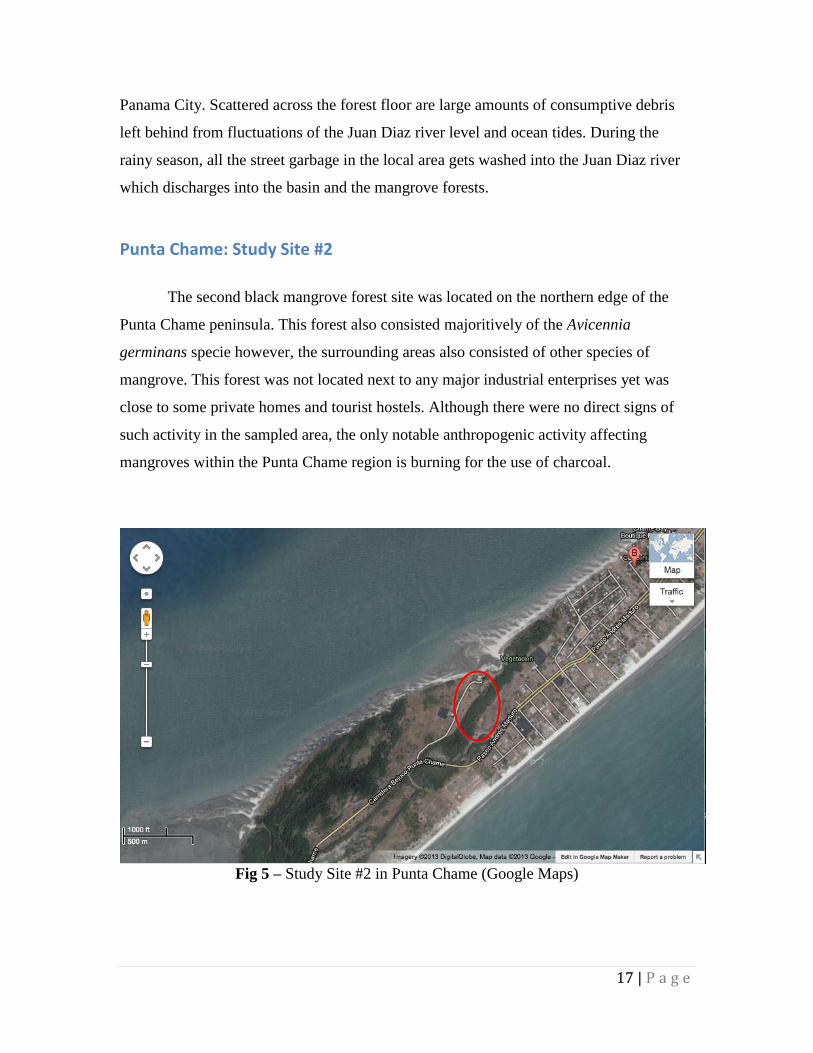
Punta Chame: Study Site #2
The second black mangrove forest site was located on the northern edge of the
Punta Chame peninsula. This forest also consisted majoritively of the Avicennia
germinans specie however, the surrounding areas also consisted of other species of
mangrove. This forest was not located next to any major industrial enterprises yet was
close to some private homes and tourist hostels. Although there were no direct signs of
such activity in the sampled area, the only notable anthropogenic activity affecting
mangroves within the Punta Chame region is burning for the use of charcoal.
Fig 5 – Study Site #2 in Punta Chame (Google Maps)
18 | P a g e
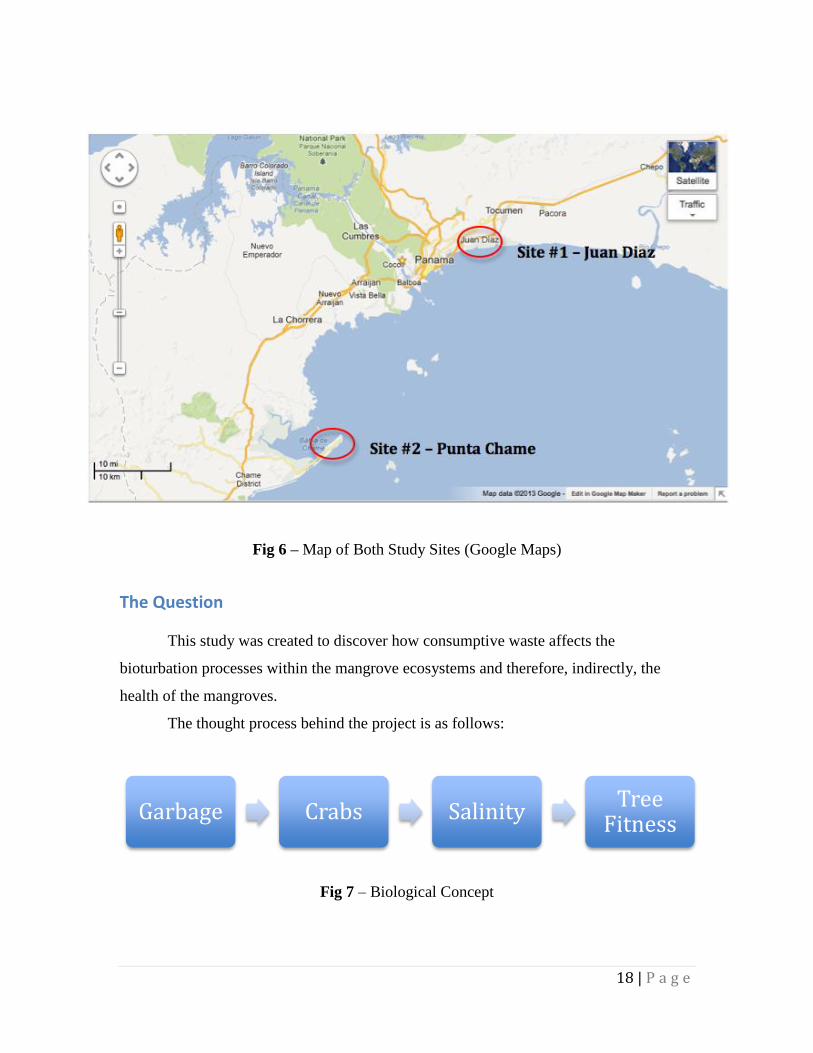
Fig 6 – Map of Both Study Sites (Google Maps)
The Question
This study was created to discover how consumptive waste affects the
bioturbation processes within the mangrove ecosystems and therefore, indirectly, the
health of the mangroves.
The thought process behind the project is as follows:
Fig 7 – Biological Concept
Garbage Crabs Salinity Tree
Fitness

19 | P a g e
This simple graphic entails garbage limiting the physical space for crabs to create
burrows. Without the existence of burrows there is limited bioturbation occurring
resulting in an accumulation of salt content and decrease in available nutrients. The
measurement of salinity was used to verify if the waste debris was actually affecting the
crab’s ability to cycle and redistribute soil contents. Salinity was selected as a good
indicator because it also has a very important relationship with the health of the trees.
With elevated salinity ratios there will be consequences for tree functionality and
therefore fitness. We are quantifying fitness, or tree health, through total aboveground
biomass calculated in the quadrats.
With this in mind the resulting question is: Will consumer trash inhibit crab
bioturbation therefore creating higher ratios of salinity which will affect the biomass and
health of the forest?
Methodology
Site Locations
Two site locations were selected for similar climatic conditions and forest
composition, both being in Pacific coast bays and consisting of majoritively Avicennia
germinans. The Juan Diaz location was selected for its condition of being heavily
polluted by consumptive waste, such as plastic and styrofoam debris. The Punta Chame
location was selected for its condition of being pristine and garbage free.
Sampling Crabs, Garbage and Trees
At both locations a 4 transect by 5 quadrat sampling design was utilized, transects
being spaced 10 meters apart and quadrats being spaced 10 meters apart along those
transects. Quadrats were 1 m2. Within quadrats the number of crab holes and percent
garbage cover were calculated. Percent garbage cover was calculated by laying a 1 m2
piece of plastic fencing with 127 holes over the quadrat and counting how many holes
contained garbage. This was converted in percentages by dividing the number of garbage
containing holes by 127 and multiplying it by 100. Note that the Punta Chame location

20 | P a g e
was selected due to its garbage free condition and therefore garbage cover was always
zero percent. From the center of each quadrat a 2.5 meter radius was used to incorporate
all surrounding trees. The circumference at breast height of each tree within the 2.5-meter
radius was measured and later converted to diameter at breast height (DBH). Many
researchers have used the following formulas to calculate biomass based on the work
done by Fromard et al. (1998):
If 1<dbh<4 (cm); then y = 200.4*DBH^2.1 (g)
dbh> 4 (cm); then y = b(DBH)^a; where a = 2.4 and b = 0.14 (kg)
Sampling and Testing Salinity
In every quadrat a soil sample was taken. This was done by inserting a spade in a
randomized spot within the quadrat until it reached a depth of 10 cm. Soil was then
collected from the bottom of the remaining hole and stored in ziplock bags to be later
tested in the lab.
Testing of the soil samples followed procedures outlined by the Salt Action
initiative of the government of New South Wales (NSW). Soil samples were left to dry in
open containers in room temperature (~27 C) for a period of 2 days. In the lab dried
samples were crushed to eliminate any large aggregates. Five grams of each soil sample
were separately weighed and combined with 25 ml of distilled water in glass beakers.
Each soil solution was mixed until any noticeable clumps were dissolved and allowed to
settle for at least two minutes. The top layer liquid was then transferred to a separate
beaker where an OysterTM Portable pH/Conductivity/TDS/ORP Salinity kit
conductometer (accuracy +/- 2%FS) was used to measure the salinities of the solutions
by placing the meter electrodes directly into the solution. Readings were recorded and
equipment was washed with distilled water before proceeding to the next sample. Once
all samples were measured the salinity meter readings were converted to soil salinity
(ECe) by multiplying the value by the conversion factor based on the texture of the soil
sample (Table 2).
21 | P a g e
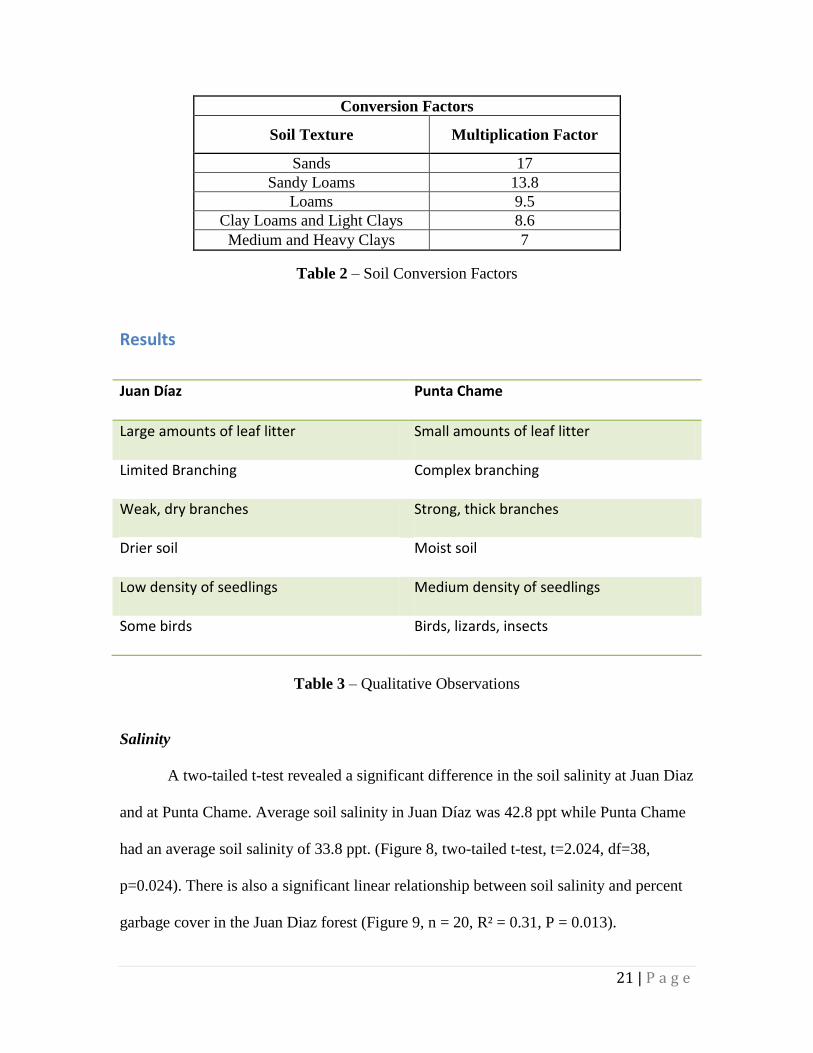
Conversion Factors
Soil Texture Multiplication Factor
Sands 17
Sandy Loams 13.8
Loams 9.5
Clay Loams and Light Clays 8.6
Medium and Heavy Clays 7
Table 2 – Soil Conversion Factors
Results
Table 3 – Qualitative Observations
Salinity
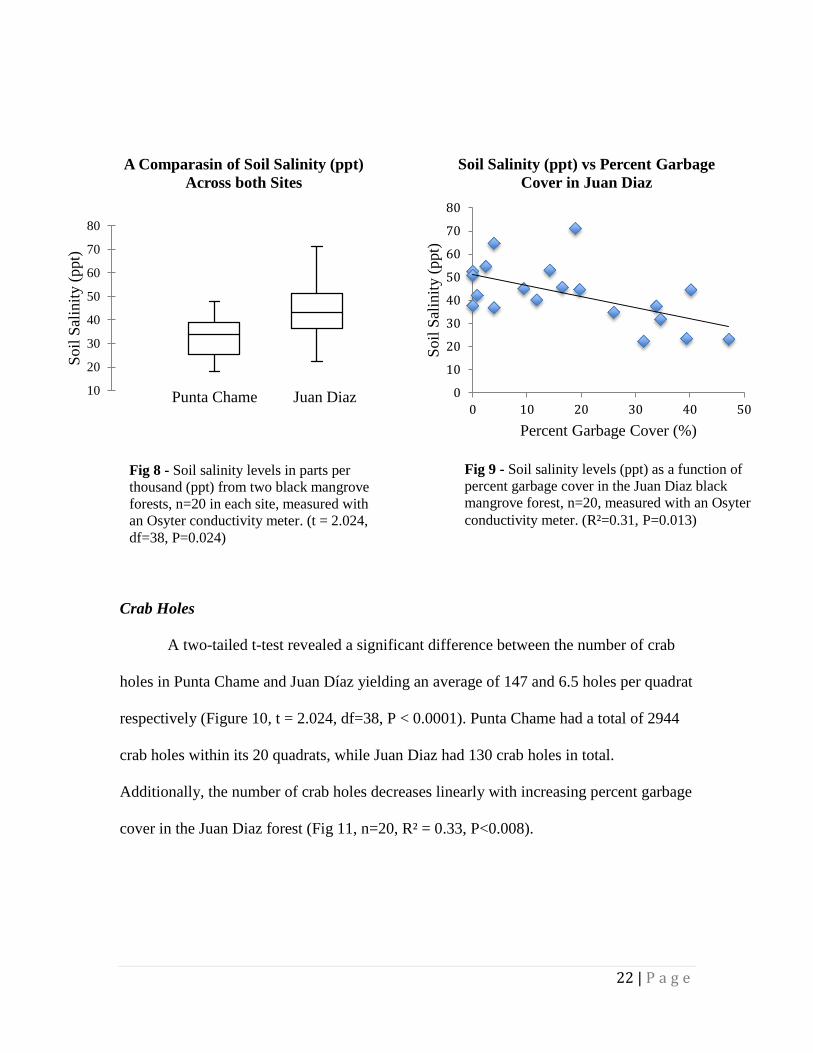
A two-tailed t-test revealed a significant difference in the soil salinity at Juan Diaz
and at Punta Chame. Average soil salinity in Juan Díaz was 42.8 ppt while Punta Chame
had an average soil salinity of 33.8 ppt. (Figure 8, two-tailed t-test, t=2.024, df=38,
p=0.024). There is also a significant linear relationship between soil salinity and percent
garbage cover in the Juan Diaz forest (Figure 9, n = 20, R² = 0.31, P = 0.013).
Juan Díaz Punta Chame
Large amounts of leaf litter Small amounts of leaf litter
Limited Branching Complex branching
Weak, dry branches Strong, thick branches
Drier soil Moist soil
Low density of seedlings Medium density of seedlings
Some birds Birds, lizards, insects
22 | P a g e
Crab Holes
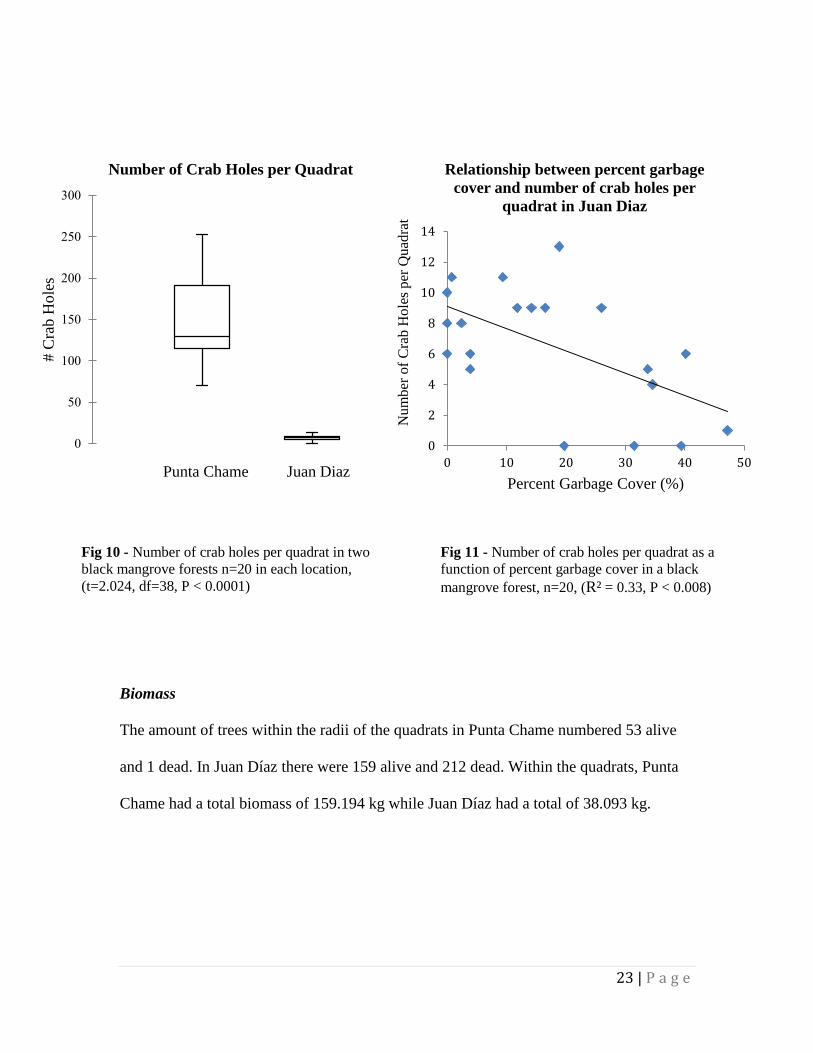
A two-tailed t-test revealed a significant difference between the number of crab
holes in Punta Chame and Juan Díaz yielding an average of 147 and 6.5 holes per quadrat
respectively (Figure 10, t = 2.024, df=38, P < 0.0001). Punta Chame had a total of 2944
crab holes within its 20 quadrats, while Juan Diaz had 130 crab holes in total.
Additionally, the number of crab holes decreases linearly with increasing percent garbage
cover in the Juan Diaz forest (Fig 11, n=20, R² = 0.33, P<0.008).
10
20
30
40
50
60
70
80
Soil
Sal
init
y (
ppt)
Punta Chame Juan Diaz
A Comparasin of Soil Salinity (ppt)
Across both Sites
Fig 8 - Soil salinity levels in parts per
thousand (ppt) from two black mangrove
forests, n=20 in each site, measured with
an Osyter conductivity meter. (t = 2.024, df=38, P=0.024)
0
10
20
30
40
50
60
70
80
0 10 20 30 40 50
Soil
Sal
init
y (
ppt)
Percent Garbage Cover (%)
Soil Salinity (ppt) vs Percent Garbage
Cover in Juan Diaz
Fig 9 - Soil salinity levels (ppt) as a function of
percent garbage cover in the Juan Diaz black
mangrove forest, n=20, measured with an Osyter
conductivity meter. (R²=0.31, P=0.013)
23 | P a g e
Biomass
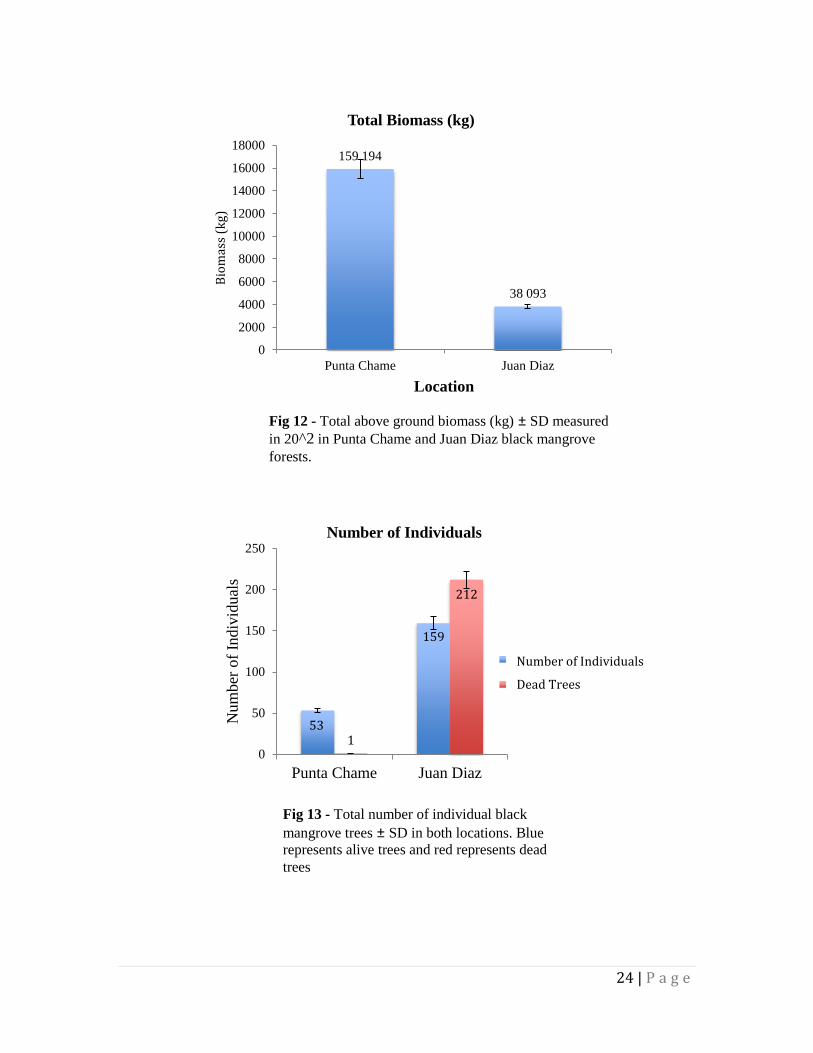
The amount of trees within the radii of the quadrats in Punta Chame numbered 53 alive
and 1 dead. In Juan Díaz there were 159 alive and 212 dead. Within the quadrats, Punta
Chame had a total biomass of 159.194 kg while Juan Díaz had a total of 38.093 kg.
0
50
100
150
200
250
300
# C
rab H
ole
s
Punta Chame Juan Diaz
Number of Crab Holes per Quadrat
0
2
4
6
8
10
12
14
0 10 20 30 40 50
Num
ber
of
Cra
b H
ole
s p
er Q
uad
rat
Percent Garbage Cover (%)
Relationship between percent garbage
cover and number of crab holes per
quadrat in Juan Diaz
Fig 10 - Number of crab holes per quadrat in two
black mangrove forests n=20 in each location,
(t=2.024, df=38, P < 0.0001)
Fig 11 - Number of crab holes per quadrat as a
function of percent garbage cover in a black
mangrove forest, n=20, (R² = 0.33, P < 0.008)
24 | P a g e
53
159
1
212
0
50
100
150
200
250
Punta Chame Juan Diaz
Num
ber
of
Indiv
idual
s
Number of Individuals
Species Density
Dead Trees
159 194
38 093
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
Punta Chame Juan Diaz
Bio
mas
s (k
g)
Location
Total Biomass (kg)
Fig 13 - Total number of individual black
mangrove trees ± SD in both locations. Blue
represents alive trees and red represents dead
trees
Fig 12 - Total above ground biomass (kg) ± SD measured
in 20^2 in Punta Chame and Juan Diaz black mangrove
forests.
Number of Individuals
Dead Trees

25 | P a g e
Discussion
Simple qualitative observations indicate that Juan Diaz, in comparison to Punta
Chame, is in a degraded state. While the ground cover in Punta Chame mostly consisted
of moist soil, crab activity, sapling growth and minimal leaf litter, the Juan Díaz forest
was composed of garbage, large amounts of leaf litter and dead tree matter. Although this
study does not explicitly prove that the rampant quantity of trash is the main cause of the
degraded condition of the Juan Diaz’s, it does suggest that there is a quantifiable
difference between the two forests despite their abiotic and biotic similarities. These
results support our initial model where garbage decreases crab burrowing capabilities
leading to a change in soil quality that in turn affects the health of the mangrove (Fig 7).
There was a significant difference between the number of crab holes per quadrat
in both forests (P < 0.0001) suggesting that less soil was subject to bioturbation processes
(Fig 10). As mentioned earlier, a lack of bioturbation processes reduces the provision of
nutrients to improve mangrove viability and may also promote the accumulation of
harmful proportions of certain soil compounds. Literature suggests that a large absence of
crabs could result in plant nutrient deficiencies and unfavorable soil conditions that
would reduce mangrove primary production and damage the overall ecosystem
functionality (Amarasinghe 2009).
Our results also yielded a significant difference between forests in terms of soil
salinity (P = 0.024) potentially indicating a lower soil quality in Juan Diaz (Fig 8 ).
Another aspect keeping the level of salinity high, could be in part due to the fact that one
of the main mechanisms of managing salinity is excess salt secretion by roots (Khazn and

26 | P a g e
Aziz 2001; Lovelock et al. 2010) Due to the large amount of garbage in the Juan Diaz
forest, salt secreted by roots and pneumatophores may be accumulating in the
surrounding soil since garbage could be inhibiting tidal flushing. In Avicennia, sodium
chloride is also deposited in senescent leaves, and when accumulated, the tree’s leaves
eventually shed, most likely as a method to remove salt from metabolic tissue (Hogarth
2007). Therefore, if the roots cannot successfully excrete the salt, then the leaves must
regulate salinity levels. Previous studies show that under chronic salinity stress, leaf
mortality is accompanied by a notable decrease in leaf production rate, which in most
cases can lead to the tree’s death. Our data and recorded observations coincide very well
with the literature as we observed a high quantity of leaves on the forest floor and many
dead trees indicating that it is quite possible that higher salinity levels are in part
responsible for the forest’s degradation.
Contrary to the original hypothesis, salinity was lower in areas with higher
garabage cover. The initial thought process behind this hypothesis was that garbage
would inhibit crabs from being able to cycle salt within the soil. However, based on the
results it may be the case that salt is settling on garbage and leaf litter instead of on the
superficial layer of the soil. This being said, the average salinity being higher in Juan
Diaz rather than Punta Chame could still be explained by a limited amount of
bioturbation. Although salt may be settling on garbage and leaf litter, the salt being
excreted or excluded from the mangroves may be accumulating around the root systems
due to the lack of bioturbation. Therefore, this study still demonstrates a negative impact
of elevated salinity levels.

27 | P a g e
One way we have come to this conclusion is based on the forest composition and
biomass. Not only was there a large reduction in biomass in Juan Diaz despite the much
higher number of individuals, but there were more dead trees present than alive trees,
inferring a damaged ecosystem (Figure 13).
Our results reveal a significant difference between Juan Diaz and Punta Chame in
terms of their crab populations and their soil salinity levels giving rise to a large
difference in biomass and tree composition. The presence of garbage could be
influencing the deterioration of the Juan Diaz black mangrove forest, since it significantly
influenced the salinity (p = 0.013) and number of crab holes (p < 0.008). It is hard to
know exactly what factors are influencing mangrove health and to what magnitude. There
are numerous indicators of health that this study could have utilized which could have
potentially offered different results. With this realization it is recommended that more
studies are designed to identify the key sources of degradation and their importance.
Despite the degradation of the Juan Díaz mangroves, conservation action may
reverse existing degradation trends. This study was not only designed to bring insight to
mangrove dynamics but was also carried out to encourage political action towards
mangrove preservation and environmentalism. This study suggests that Panama needs a
more effective waste management system to keep consumptive waste out of natural
systems. This may entail the formation of new legislation but may also require a shift in
the Panamanian cultural paradigm in which the common citizen is more conscious of
daily consumption habits.

28 | P a g e
Limitations and Obstacles Encountered
Throughout the course of our internship and research project we encountered
several obstacles and limiting factors. Of all of these the three that were the most
impacting were transport, contacts and funding. Transport was extremely difficult
considering the amount of equipment that we needed to transfer to our study sites. With
the lack of a personal vehicle, much time, effort and money was spent carrying pounds of
equipment via public transportation in crowded conditions. Access to both of our study
sites were also very difficult to get to in general and always dependent on other people.
These study site contacts were difficult to find, maintain and utilize. Access to Juan Diaz
was granted by a stroke of good fortune while access to a control site took almost two
months of contact searching, communicating and planning. Finally, funding may have
been the greatest obstacle and limitation. This is due to the fact that the original study
included measuring nitrogen soil content rather than salinity. Almost $400 was spent to
design an experiment that looked at nitrogen on a temporal scale. However, after the
necessary chemicals ran out, additional expenses were deemed too costly to purchase the
remaining chemicals needed to complete the experiment. Therefore, the original
equipment purchased had no final value and approximately 2 months of work was
nullified.
Additional minor obstacles included:
-Access to technical expertise
-Time constraints when balancing course load and contact availability
-Manpower

29 | P a g e
Looking Forward
The main objective of supplying this study to the Centro de Incidencia Ambiental
was to add to an index of citable knowledge in hopes that they will be able to utilize it in
some shape or form when suggesting conservation legislation or educating the greater
public. Additionally, the research team would like to see this study encourage additional
and similar investigations that may in the long run contribute to sparking political
initiatives that will aid in formulating environmental policy and frameworks. Research,
like the type conducted in this project, will help to elucidate biological details that in turn
will support effective environmentalism.
Logistical Information
Work Log
Throughout our internship approximately 35 days were dedicated to work either in the
CIAM office, performing literature review and research or attending CIAM coordinated
events. Additionally, 7 full days were spent in the field.
Financial Expenditures
Hardware 352.43
Taxis ~$58
Bus ~$40
Lodging $30
Total $480.43

30 | P a g e
Table 4 - Expenses
Official Recognition Requests
Official acknowledgement letters should be sent to the following individuals:
1. Lourdes Lozano
2. Tania Arosemena
3. Natty Vasquez
4. Rodrigo Noriega
5. Sonia Montenegro
6. Gustavo Cárdenas
The above six individuals can have their letter addressed to:
Urb. Los Ángeles, Calle Los Periodistas
Casa G-14 Planta Alta.
Panamá, Rep. de Panamá
Additionally:
7. Ivania Cerno
Naos Marine Laboratories
Bldg. 356, Amador Causeway, Naos Island
Panama, Rep. de Panamà
8. Carlos Manuel Peralta
Additional Acknowledgments
We would like to thank:
Rose Marie and Gilles for their hospitality and guidance.
César Rodríguez for his assitance in Punta Chame.
Elana Evans and Samuel Bulow for their assitance in Juan Diaz.
Catherine Lovelock for all the information she provided.

31 | P a g e
Bibliography
AIDA - Interamerican Association for Environmental Defense: Human Rights and
Environmental. (2011) Developers challenge Panama Bay’s protected status.
Retrieved from http://www.aida-americas.org/en/project/developers-challenge-
panama-bay%E2%80%99s-protected-status
Amarasinghe, M. (2009). Mangrove crabs: the ecosystem engineers and keystone species.
Network of Aquaculture Centres in Asia-Pacific. Retrieved from
http://www.enaca.org/modules/news/article.php?storyid=1855
Angehr, George R. (2003). Directorio de areas importantes para aves en Panama.
Directory of important bird areas in Panama. Panama: Sociedad Audubon de
Panama
Audubon de Panamá. (n.d.). Birdlife Data Zone. BirdLife International - conserving the
world's birds. Retrieved from
http://www.birdlife.org/datazone/sitefactsheet.php?id=19322
Basak, U. C., Gupta, N., Rautaray, S., & Das, P. (January 01, 2004). Effects of salinity on
the growth of mangrove seedlings. Journal of Tropical Forest Science, 16, 4, 437-
443.
Benfield. S. (2002). An assessment of mangrove cover and forest structure in Punta Mala
Bay, Panama city, Panama by means of field study and GIS analysis of aerial
photographs. Masters Thesis, Heriot Watt University, Edinburgh, Scotland.
Betts, T. (2006). An Assessment of Mangrove Cover and Forest Structure in Las Perlas,
Panama. School of Life Sciences. Heriot-Watt University, Edinburgh.
http://www.stri.si.edu/sites/darwin_initiative/PDFs/TaniaBetts.pdf
BirdLife International (2013. Important Bird Areas factsheet: Upper Bay of Panamá.
Retrieved from http://www.birdlife.org
Danielsen, F. et al. 2005. The Asian tsunami: A protective role for coastal vegetation.
Science, 310, 643-644.
Donato D. et al. (2011). Mangroves among the most carbon-rich forests in the tropics
Nature Geoscience, 4, 293–297 doi:10.1038/ngeo1123
Ecologic Development Fund. (n.d.). Mangroves: Unique Forests by the Sea. EcoLogic
Development Fund. Retrieved from http://www.ecologic.org/en/issues/mangroves
Ellison, A., Farnsworth, E. & Moore, G. (2010). Avicennia germinans. In: IUCN 2012.
IUCN Red List of Threatened Species. Version 2012.2. <www.iucnredlist.org>.

32 | P a g e
FLMNH Ichthyology Department: South Florida Aquatic Environments. (n.d.). Home :
Florida Museum of Natural History. Retrieved April 18, 2013, from
http://www.flmnh.ufl.edu/fish/southflorida/mangrove/profiles.html#black
Gillikin, David Paul, De Wachter, Bart and Tack, Jurgen F., (2004). Physiological
responses of two ecologically important Kenyan mangrove crabs exposed to
altered salinity regimes Journal of Experimental Marine Biology and Ecology,
301(1), 93-109.
Harnandez, C. (2007) Gestión de desechos sólidos en Panamá. Autoridad Nacional del
Ambient. Retrieved from www.epa.gov/.../Gestion_desechos_panama.pdf
Houck, M., Neill, R. (2009). Plant fact sheet for black mangrove (Avicennia Germinans
(L.) L.). USDA - Natural Resources Conservation Service, Louisiana Plant
Materials Center, Galliano, Louisiana.
Jackson, E. (2012). High court and ANAM lift mangrove protections, environmentalists
unite to protest. The Panama News, Panama's English Language Newspaper.
Retrieved from http://www.thepanamanews.com/pn/v_18/issue_05/news_05.html
Khan, M. A., & Aziz, I. (January 01, 2001). Salinity tolerance in some mangrove species
from Pakistan. Wetlands Ecology and Management, 9, 3, 229-233.
Lovelock, C. E., Ball, M. C., Feller, I. C., Engelbrecht, B. M. J., & Ling, E. M. (July 01,
2006). Variation in hydraulic conductivity of mangroves: influence of species,
salinity, and nitrogen and phosphorus availability. Physiologia Plantarum, 127, 3,
457-464.
Marine habitats. (n.d.). Dutch Caribbean Biodiversity Explorer. Retrieved from
www.dcbiodata.net/explorer/info/habitats
McGinley m., & Hogan, M. C., (2008). Gulf of Panama mangroves. Encyclopedia of
Earth. Retrieved from
<http://www.eoearth.org/article/Gulf_of_Panama_mangroves?topic=49597>
McHugh, A. J. (2007). Threatened mangrove populations on the Pacific coast of
Panama: what environmental public policy strategies can be employed to deal
with this threat. Panama : Smithsonian Tropical Research Institute.
Pereira, D.R. (2012). Un país que no sabe reciclar. La Estrella. Retrieved from
http://www.laestrella.com.pa/online/impreso/2012/09/30/un-pais-que-no
sabereciclar.asp
Radhika, D. (2006). Mangrove Ecosystems of Southwest Madagascar: An
Ecological,Human Impact, and Subsistence Value Assessment. Tropical Resources
Bulletin, 25.

33 | P a g e
Ramsar Convention - Briefing Notes. (2000, January 31). Ramsar Convention. Retrieved
March 20, 2013, from http://www.ramsar.org/cda/en/ramsar-pubs-notes
annotated-ramsar-16127/main/ramsar/1-30-168%5E16127_4000_0__
Suaarez, N., & Medina, E. (January 01, 2005). Salinity effect on plant growth and leaf
demography of the mangrove, Avicennia germinans L. Trees, 19, 6, 722-728.
Sobrado, M. A. (January 01, 2000). Leaf Photosynthesis of the Mangrove Avicennia
Germinans as Affected by NaCl. Photosynthetica, 36, 4, 547-555
Stieglitz, T., Ridd, P. and Muller, P. (2000) Passive irrigation and functional morphology
of crustacean burrows in a tropical mangrove swamp. Hydrobiologia, 421:69-76.
Warne, K. (n.d.). Mangroves - National Geographic Magazine. National Geographic
Magazine. Retrieved from
http://ngm.nationalgeographic.com/2007/02/mangroves/warne-text/2
Wang, J. Q., Zhang, X. D., Jiang, L. F., Bertness, M. D., Fang, C. M., Chen, J. K., Hara,
T., ... Li, B. (2010). Bioturbation of Burrowing Crabs Promotes Sediment
Turnover and Carbon and Nitrogen Movements in an Estuarine Salt
Marsh. Ecosystems,13, 4, 586-599.





























