Embed Size (px)
Citation preview

Role of penicillin-binding protein1b in competitive stationary-phase survival ofEscherichia coliEvan D. Pepper, Michael J. Farrell & Steven E. Finkel
Molecular and Computational Biology Program, Department of Biological Sciences, University of Southern California, Los Angeles, CA, USA
Correspondence: Steven E. Finkel,
Molecular and Computational Biology
Program, MCB 201, University of Southern
California, Los Angeles, CA 90089-2910,
USA. Tel.: 11 213 821 1498; fax: 11 213 740
8631; e-mail: [email protected]
Received 4 May 2006; revised 7 July 2006;
accepted 10 July 2006.
First published online 15 August 2006.
DOI:10.1111/j.1574-6968.2006.00418.x
Editor: Robert Gunsalus
Keywords
penicillin-binding proteins; stationary phase-
specific competition defect; osmotic stress.
Abstract
The penicillin-binding proteins (PBPs) catalyze the synthesis and modification of
bacterial cell wall peptidoglycan. Although the biochemical activities of these
proteins have been determined in Escherichia coli, the physiological roles of many
PBPs remain enigmatic. Previous studies have cast doubt on the individual
importance of the majority of PBPs during log phase growth. We show here that
PBP1b is vital for competitive survival of E. coli during extended stationary phase,
but the other nine PBPs studied are dispensable. Loss of PBP1b leads to the
stationary phase-specific competition defective phenotype and causes cells to
become more sensitive to osmotic stress. Additionally, we present evidence that this
protein, as well as AmpC, may assist in cellular resistance to b-lactam antibiotics.
Introduction
Twelve penicillin-binding proteins (PBPs) have been identi-
fied in Escherichia coli: five high molecular weight PBPs (1a,
1b, 1c, 2, and 3) and seven low molecular weight PBPs (4, 5,
6, 7, AmpC, AmpH, and DacD). Two of these proteins,
PBP2 (mrdA) and PBP3 (ftsI), are essential for normal
growth (Spratt, 1975) and are not further characterized in
this study. Although the physiological functions of PBPs 1a,
1b, and 5 are fairly well understood (Denome et al., 1999;
Nelson & Young, 2000), the biological roles of the remaining
PBPs are not fully determined, even though most of their
specific in vitro biochemical activities have been elucidated.
Briefly, PBPs 1a (mrcA) and 1b (mrcB) are transglycosylases
and transpeptidases (Ghuysen & Dive, 1994); PBP1c is a
close homolog to PBPs 1a and 1b but has not been
completely characterized (Schiffer & Holtje, 1999); PBPs 4,
5, 6, and DacD are carboxypeptidases (Matsuhashi et al.,
1979; Amanuma & Strominger, 1980; Korat et al., 1991;
Baquero et al., 1996); PBPs 4 and 7 are endopeptidases
(Korat et al., 1991; Romeis & Holtje, 1994); AmpC is a
b-lactamase, and AmpH binds many b-lactams, though it
has not been shown to possess demonstrable b-lactamase
activity (Henderson et al., 1997).
PBPs are found in all free-living bacteria. Their ubiquity
suggests an important role in cell physiology. They are known
to synthesize and remodel cell wall peptidoglycan, which gives
the cell its rigidity and shape, prevents osmotic lysis and resists
toxins. PBPs are also the target of many antibiotics. Although
E. coli maintains 12 PBP genes, many of the gene products have
overlapping biochemical functions and seem to be nonessential
for exponential phase growth. In fact, an E. coli mutant lacking
eight of 12 PBPs is viable (Denome et al., 1999). Perhaps
individual PBPs play more important roles in natural environ-
ments, where cells compete for limited carbon and energy
sources, encounter antimicrobial agents, and are subjected to a
variety of other stresses. Escherichia coli faces similarly stressful
environmental challenges during long-term stationary-phase
batch culture incubation, where nutrients are depleted and the
cells encounter alkaline and oxidative stress conditions (Farrell
& Finkel, 2003; Finkel, 2006). Thus, in light of previous
experiments casting doubt on their importance during log
phase, we sought to broaden the understanding of the physio-
logical functions of the PBPs using stationary-phase survival
assays conducted under a variety of laboratory conditions.
Materials and methods
Bacterial strains
Strains used in this study are derived from E. coli K-12 strain
ZK126 (W3110 DlacU169 tna-2) and are listed in Table 1.
FEMS Microbiol Lett 263 (2006) 61–67 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
Mutants of 10 PBP genes were constructed by homologous
recombination and gene replacement with a chlorampheni-
col-resistance cassette (CamR) flanked by FLP recombinase
(FRT) sites (see Datsenko & Wanner, 2000). A tnaA mutant
was also constructed in the same manner to serve as a
control strain. This strain shows the same growth, survival,
and competition phenotypes as the parent strain. The CamR
mutations were transduced by bacteriophage P1 into the
genetically marked strain ZK1142 (NalR) (see Zambrano
et al., 1993; Finkel & Kolter, 1999). Removal of CamR
resistance cassettes was achieved by transformation of each
strain with a temperature-sensitive plasmid expressing FLP
recombinase (Datsenko & Wanner, 2000). For complemen-
tation experiments, PBP mutant SF2341 (mrcB) was trans-
formed with plasmid pSAD426-11, which expresses mrcB, or
the plasmid pSAD444-1, which carries a mutant variant of
mrcB (Denome et al., 1999).
Culture conditions and titering assays
Five milliliter cultures were grown in 18� 150 mm test tubes
at 37 1C with aeration in a TC-7 roller drum (New Bruns-
wick Scientific, Edison, NJ). Luria–Bertani (LB) medium
was prepared according to manufacturer’s instructions (Dif-
co). For all titering assays, viable counts were determined by
serial dilution of cells removed periodically from the cul-
tures, followed by plating on LB agar containing appropriate
antibiotics at the following concentrations: nalidixic acid
(20 mg mL�1), streptomycin (25mg mL�1), or chlorampheni-
col (30mg mL�1). To measure long-term survival of indivi-
dual strains in monoculture, 5 mL cultures were inoculated
1 : 1000 (v/v) from fresh overnight cultures started from
frozen LB–glycerol stocks. To ascertain the relative fitness of
mutant strains, wild-type and PBP mutants were either
coinoculated 1 : 1000 as described above, or 2.5 mL of each
strain from overnight cultures were mixed, followed by
periodic sampling, serial dilution, and plating on selective
media that allowed quantification of individual subpopula-
tions. Where appropriate, cultures were buffered at pH 5
with 100 mM MES, pH 7 with 100 mM HEPES, or at pH 9
with 100 mM TAPS (Sigma, Aldrich). To determine dou-
bling time, c. 103 CFU mL�1 from overnight cultures were
inoculated into either fresh LB or conditioned medium and
quantified hourly during exponential growth phase.
Table 1. Strains used in this study
Strain Relevant phenotype Gene Sequence of knockout primer pair� or Reference
ZK126 Wild type – Zambrano et al. (1993); Finkel & Kolter (1999).
ZK1142 ZK126 NalR – Zambrano et al. (1993); Finkel & Kolter (1999).
ZK1143 ZK126 StrR – Finkel & Kolter (1999).
SF2338 PBP1a�NalR mrcA H1P1: gcctataccgctacatcgagccacaactgccggatggtgtaggctggagctgcttc
H2P2: ctgacgtaagcgaattggaccagcatactgcggtggcatatgaatatcctcctta
SF2341 PBP1b�NalR mrcB H1P1: gacgattatgatgactatgaggatgaagaaccgatggtgtaggctggagctgcttc
H2P2: catatccttgatccaaccggctacaccgtcgctgtccatatgaatatcctccttag
SF2365 PBP1c�NalR pbpC H1P1: cactgtcgcgatttgccccacgtcatccatcaccaggtgtaggctggagctgcttc
H2P2: gagcagataaactctggcctctaccgctgcatgaagcatatgaatatcctcctta
SF2344 PBP4�NalR dacB H1P1: gatttatcatcggattgaccagctgtatagcgttcagtgtgtaggctggagctgcttc
H2P2: gctttcaaaacgcactaacggaatacggcgattacgctgcatatgaatatcctccttag
SF2347 PBP5�NalR dacA H1P1: cgctcaccacggctctttgcacagcctttatctctggtgtaggctggagctgcttc
H2P2: cacaaccagcgggcgttgctcgatcgttttgccatccatatgaatatcctccttag
SF2350 PBP6�NalR dacC H1P1: ctcctctctccttcgtggtcttgcagcgggttctgcgtgtaggctggagctgcttc
H2P2: gaaatcccacacccgaccaaagaatccgccctcttccatatgaatatcctccttag
SF2362 PBP6b�NalR dacD H1P1: ggtcagatcctcaccgcgggtaatgagcatcaacaggtgtaggctggagctgcttc
H2P2: cactcaggcgagaaaacatgctgccttccccgacagcatatgaatatcctcctta
SF2353 PBP7�NalR pbpG H1P1: gcctggccctgatgctggctgtgccttttgcaccgcgtgtaggctggagctgcttc
H2P2: cgttctgtgccgtctgccccgccgccgccatttgggcatatgaatatcctccttag
SF2356 AmpC�NalR ampC H1P1: ccacatttgctgcccctcaacaaatcaacgatattggtgtaggctggagctgcttc
H2P2: cgttaagaatctgccaggcggcgtcgactctcgctgcatatgaatatcctccttag
SF2359 AmpH�NalR ampH H1P1: ccgatcactgaaccggagtttgcctctgatattgtcgtgtaggctggagctgcttc
H2P2: caacggtttattcccgcttagctcggtcaccaggtccatatgaatatcctccttag
SF2336 TnaA�NalR tnaA H1P1: actttaaacatctccctgaaccgttccgcattcgtgtgtgtaggctggagctgcttc
H2P2: acttctttaagttttgcggtgaagtgacgcaatactttcatatgaatatcctcctta
SF2370 SF2341 pSAD426-11 mrcB1 This strain carries a plasmid expressing PBP1b. Denome et al. (1999)
SF2371 SF2341 pSAD444-1 mrcB This is the vector-only control strain for SF2370. Denome et al. (1999)
�The sequences of P1 and P2 are the same for each primer, corresponding to regions flanking the chloramphenicol-resistance cassette used to replace
each gene (Datsenko & Wanner, 2000). H1 and H2 correspond to sequences near the beginning (H1) or end (H2) of each gene.
FEMS Microbiol Lett 263 (2006) 61–67c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
62 E.D. Pepper et al.
Antibiotic sensitivity assays
In these experiments, 200mL of cells from overnight cultures
were spread onto 25 mL LB agar (in standard 15 cm dia-
meter Petri dishes). Sterile filter discs (13 mm diameter)
infused with 50 mL of antibiotic solution [ampicillin, cefox-
itin, moxalactam, kanamycin, chloramphenicol, or strepto-
mycin (Sigma-Aldrich) at either 1.5 or 7.5 mg mL�1] were
then placed on the inoculated agar. After 16–20 h of incuba-
tion at 37 1C, the diameter of the zone of inhibition was
measured. For minimum inhibitory concentration (MIC)
assays, overnight cultures were inoculated 1 : 1000 into fresh
LB broth containing an appropriate range of antibiotic
concentrations (see Results), then incubated for 18–20 h
and quantified to determine the lowest concentration that
inhibited the growth of microorganisms.
Osmotic stress assays
For osmotic stress experiments, LB medium was supple-
mented with sucrose to a final concentration of 0.5–2 M.
Hypertonic cultures were incubated as described above and
viable counts were determined periodically.
Conditioned medium assays
To prepare conditioned medium, cultures were inoculated
with either wild-type or mutant bacteria and incubated for
1–5 days, followed by filter sterilization (0.22mm filters;
Nalgene) to remove bacterial cells from the medium. Cells
from freshly grown overnight cultures were then inoculated
1 : 1000 into conditioned media and monoculture assays
were performed as described to quantify viable cells. For
long-term survival assays, conditioned medium from 1- or
5-day-old cultures was used. For outgrowth experiments
used to determine doubling time in conditioned medium,
medium from 3-day-old cultures was used.
Results and discussion
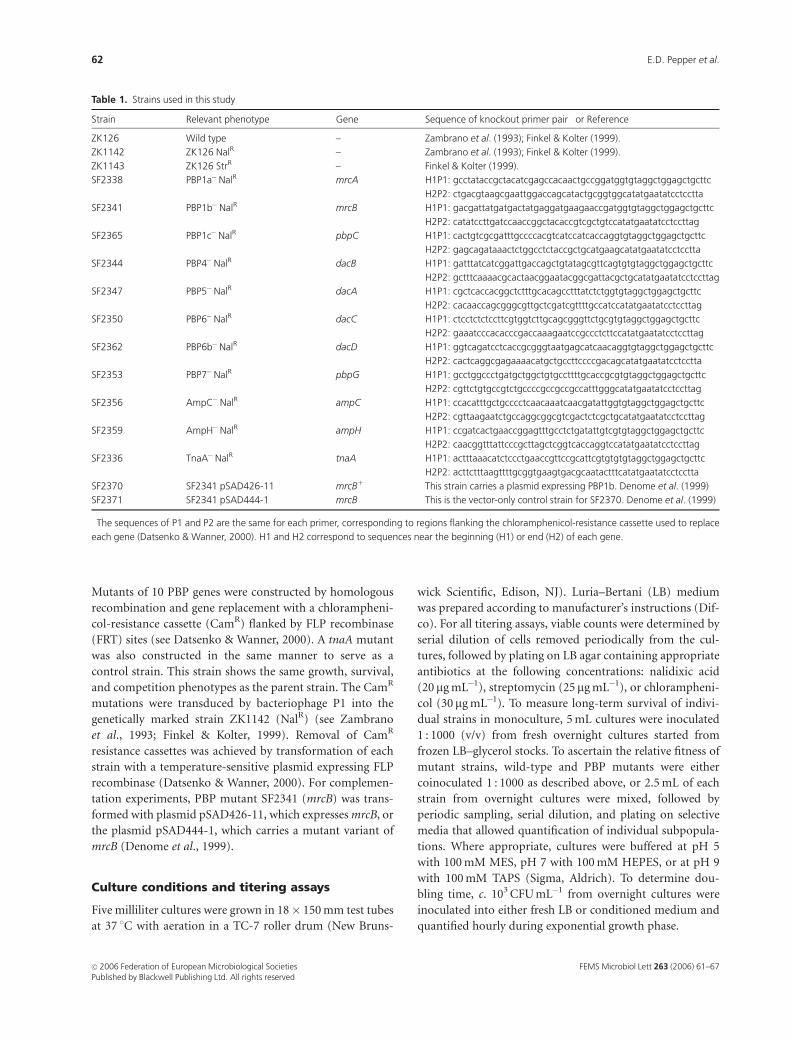
Cultured in LB under standard laboratory conditions, single
mutations in each PBP gene tested had no statistically
significant effect on bacterial growth yields (Fig. 1a). How-
ever, PBP1b� strains show an increase in average doubling
time during exponential phase compared with wild-type cells
(discussed below). All PBP mutants were viable during long-
term monoculture for more than 10 days, exhibiting similar
survival patterns to the parent and control strains (Fig. 1a).
To determine whether any PBP mutants display the
stationary phase-specific competition defective (SPCD)
phenotype (Finkel & Kolter, 2001; R. Yalamanchili and
S. Finkel, manuscript in preparation), we competed PBP
mutants against wild-type strains in LB batch cultures. The
SPCD phenotype is defined by the inability of a mutant
strain to coexist with its parental strain during long-term
14121086420
4
6
8
10
PBP1B−
log
CF
U m
L−1
*
1086420
*
4
6
8
10
2
PBP1B−
Day
4
6
8
10
21086420
***
PBP1B−
(a)
(b)
(c)
Fig. 1. Long-term survival and competition of wild-type and PBP mutant
strains. (a) Composite of representative long-term survival patterns when
wild-type or PBP mutant strains are grown individually in monoculture:
wild-type and control strains (&), the PBP1b� strain (n), and all other
PBP mutant strains (�). (b) Composite of representative survival patterns
when strains are co-inoculated (1 : 1000) in competition with the wild-
type ZK1143, the StrR wild-type parent (—). The squares and triangles
indicate competition pairs: ZK1143 ( ) vs. either the Tna� control
strain ( ) or the PBP1b� mutant strain ( ). (c) Composite of
survival patterns when wild-type (—) and PBP1b�mutant strains (- - - -)
are mixed at a 1 : 1 ratio from overnight cultures. Circles and squares
indicate competition pairs. Asterisks indicate cell titers are below the limit
of detection (o 1000 CFU mL�1). PBP1b� strains are indicated. Represen-
tative data is shown (nZ3 for all experiments).
FEMS Microbiol Lett 263 (2006) 61–67 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
63PBP1b is required for competitive stationary-phase survival
coculture in stationary phase. In contrast, when cultured
alone, the mutant strain shows no long-term growth or
survival defects. In competition assays, only the PBP1b
mutant showed a significant SPCD phenotype (Fig. 1b).
The competitive fitness of this mutant is sharply reduced,
leading to its extinction from the population within 1 week
of coculture. When transformed with a plasmid expressing
mrcB (pSAD426-11), long-term fitness of PBP1b mutants
increased by at least two orders-of-magnitude over a 10-day
period compared with strains transformed with the vector
plasmid pSAD444-1 (data not shown). None of the other
PBP mutant strains tested showed a significant decrease in
fitness or viability compared with the tna control strain
(data not shown). The SPCD phenotype is also apparent
when mrcB mutants are competed against parental strains in
conditioned media (data not shown) or when both strains
are grown individually to stationary phase and then mixed
such that the initial population of both strains is approxi-
mately equal (initial density �109 CFU mL�1; see Fig. 1c).
Thus, even though mrcA mrcB double mutants, lacking both
PBP1a and PBP1b, are lethal (Suzuki et al., 1977) and single
mutations in either gene are not (indicating overlapping
function), the PBP1b mutant displayed the SPCD pheno-
type (Fig. 1b), whereas the PBP1a mutant exhibited normal
competitive ability. This suggests that PBP1b plays an
important role in E. coli during stationary phase and
correlates well with previous studies proposing that PBP1b
possesses a greater biosynthetic capacity than PBP1a (Garcia
del Portillo & de Pedro, 1990), in spite of the fact that both
proteins are transglycosylases and transpeptidases. It is also
in agreement with studies that conclude that PBP1b is the
major high-molecular PBP in E. coli (Chalut et al., 2001).
Possessing multiple PBPs may also assist the cell in
resisting the effects of certain antibiotics that target cell wall
synthesis. PBPs interact with b-lactam antibiotics, such as
ampicillin, in differing manners. Some form stable serine
esters, while others have weak to strong b-lactam hydrolyz-
ing activity (Livermore, 1995). Thus we expected that
individual PBP mutants might display differential sensitivity
and/or resistance to b-lactams. The wild-type and all PBP
mutant strains were tested for b-lactam sensitivity using
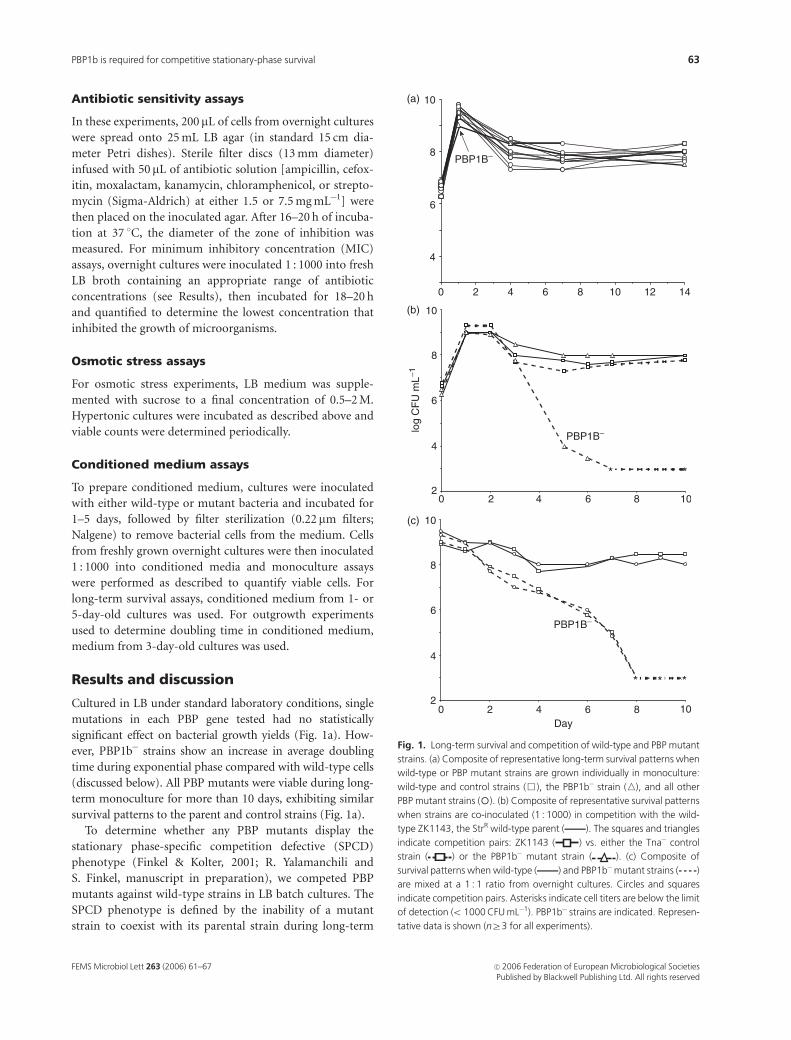
zone of inhibition and MIC assays. Our results indicate that
mutations in mrcB cause E. coli to become more sensitive to
ampicillin (Fig. 2) as well as the b-lactams cefoxitin and
moxalactam (data not shown). In zone of inhibition assays,
this effect was completely reversed by complementation
with a plasmid carrying mrcB1 (Fig. 2). Importantly, PBP1b
mutants were not more sensitive than wild-type cells to
other antibiotics that target protein synthesis, including
kanamycin, chloramphenicol, and streptomycin (data not
shown). Mutations in ampC, as expected, also increased
ampicillin sensitivity (Fig 2; also see Henderson et al., 1997),
though mutants of ampH, a b-lactamase homolog, and all
other PBP mutants tested had responses similar to the wild-
type (data not shown). In MIC assays, mrcB mutants were
also more susceptible than wild-type strains to the effects of
ampicillin (MIC = 3.0 mg mL�1 for mrcB vs. 6.0mg mL�1 for
wild-type), cefoxitin (1.5 vs. 2.5mg mL�1), and moxalactam
(0.15 vs. 0.30 mg mL�1).
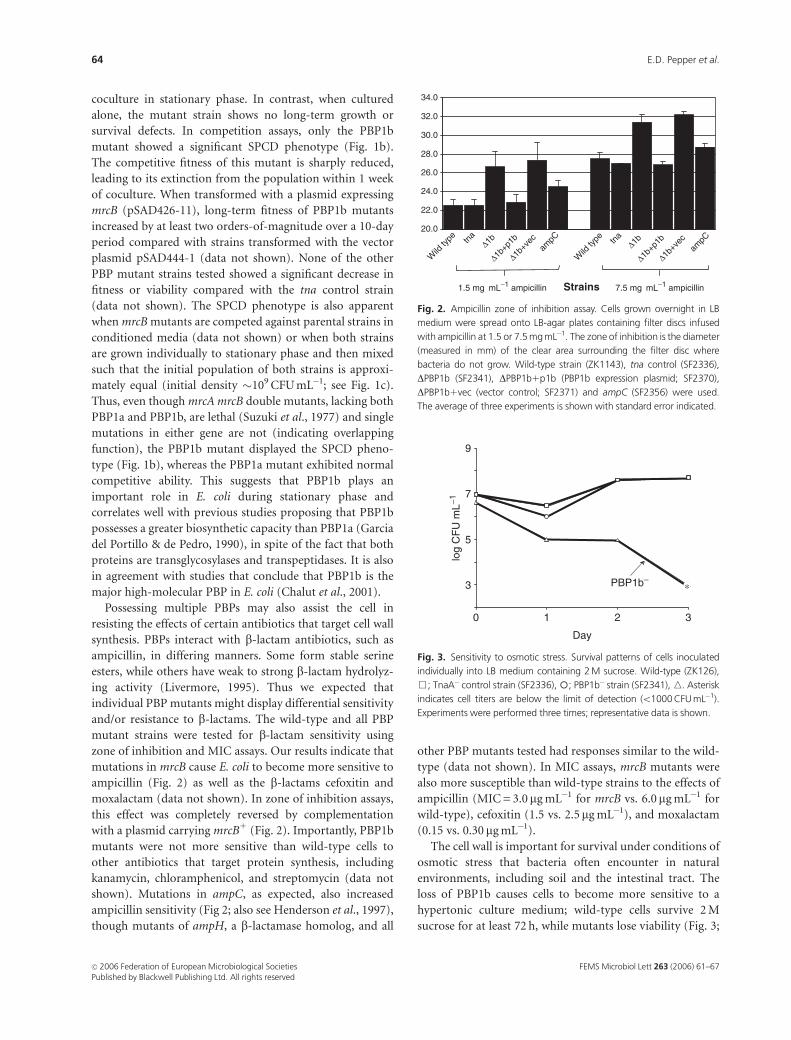
The cell wall is important for survival under conditions of
osmotic stress that bacteria often encounter in natural
environments, including soil and the intestinal tract. The
loss of PBP1b causes cells to become more sensitive to a
hypertonic culture medium; wild-type cells survive 2 M
sucrose for at least 72 h, while mutants lose viability (Fig. 3;
20.0
22.0
24.0
26.0
28.0
30.0
32.0
34.0
Wild
type tn
a∆1b ∆1b
∆1b+p
1b
∆1b+p
1b
∆1b+v
ec
∆1b+v
ecam
pCam
pC
Wild
type tn
a
Strains
Fig. 2. Ampicillin zone of inhibition assay. Cells grown overnight in LB
medium were spread onto LB-agar plates containing filter discs infused
with ampicillin at 1.5 or 7.5 mg mL�1. The zone of inhibition is the diameter
(measured in mm) of the clear area surrounding the filter disc where
bacteria do not grow. Wild-type strain (ZK1143), tna control (SF2336),
DPBP1b (SF2341), DPBP1b1p1b (PBP1b expression plasmid; SF2370),
DPBP1b1vec (vector control; SF2371) and ampC (SF2356) were used.
The average of three experiments is shown with standard error indicated.
Day
log
CF
U m
L−1
3210
3
5
7
9
PBP1b−*
Fig. 3. Sensitivity to osmotic stress. Survival patterns of cells inoculated
individually into LB medium containing 2 M sucrose. Wild-type (ZK126),
&; TnaA� control strain (SF2336),�; PBP1b� strain (SF2341), n. Asterisk
indicates cell titers are below the limit of detection (o1000 CFU mL�1).
Experiments were performed three times; representative data is shown.
FEMS Microbiol Lett 263 (2006) 61–67c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
64 E.D. Pepper et al.
compare with Fig 1a). At concentrations below 2 M sucrose
both wild-type and mrcB cells have similar survival character-
istics (data not shown). All other PBP mutants survived
osmotic stress as well as the parent and control strains (data
not shown). The sensitivity of the PBP1b mutant to osmotic
stress under laboratory conditions suggests that PBP1b may less
efficiently build or maintain its cell wall, a factor that may
contribute to the SPCD phenotype displayed by the mutant.
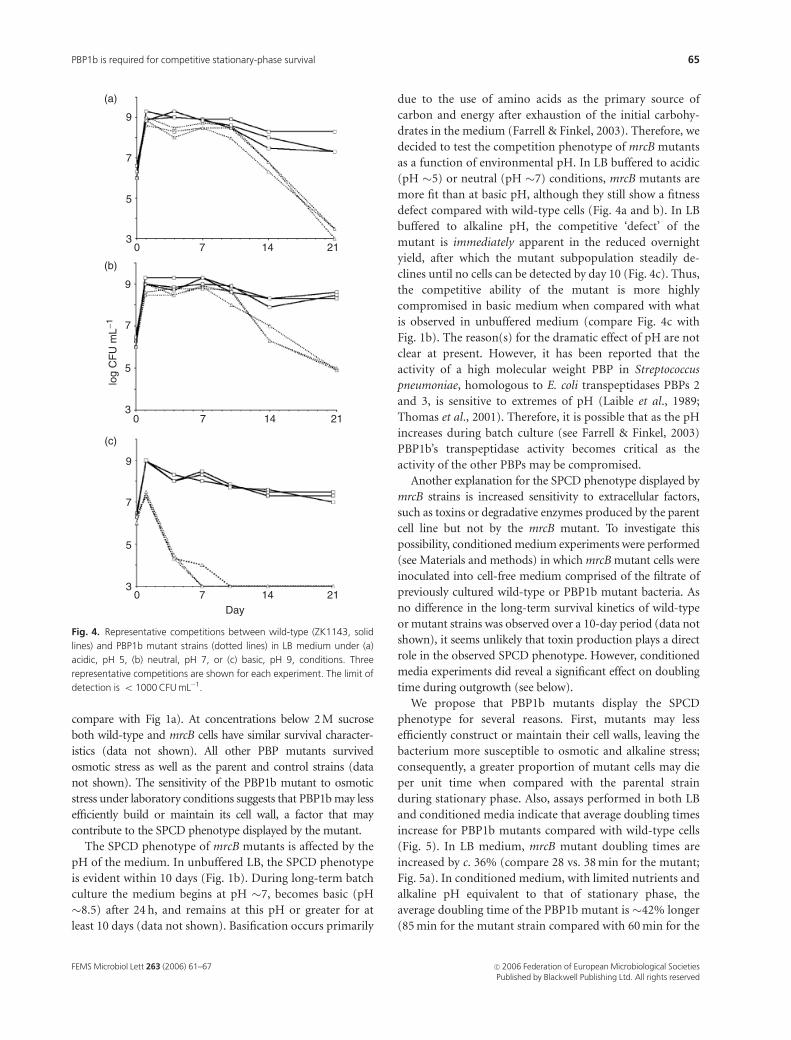
The SPCD phenotype of mrcB mutants is affected by the
pH of the medium. In unbuffered LB, the SPCD phenotype
is evident within 10 days (Fig. 1b). During long-term batch
culture the medium begins at pH �7, becomes basic (pH
�8.5) after 24 h, and remains at this pH or greater for at
least 10 days (data not shown). Basification occurs primarily
due to the use of amino acids as the primary source of
carbon and energy after exhaustion of the initial carbohy-
drates in the medium (Farrell & Finkel, 2003). Therefore, we
decided to test the competition phenotype of mrcB mutants
as a function of environmental pH. In LB buffered to acidic
(pH �5) or neutral (pH �7) conditions, mrcB mutants are
more fit than at basic pH, although they still show a fitness
defect compared with wild-type cells (Fig. 4a and b). In LB
buffered to alkaline pH, the competitive ‘defect’ of the
mutant is immediately apparent in the reduced overnight
yield, after which the mutant subpopulation steadily de-
clines until no cells can be detected by day 10 (Fig. 4c). Thus,
the competitive ability of the mutant is more highly
compromised in basic medium when compared with what
is observed in unbuffered medium (compare Fig. 4c with
Fig. 1b). The reason(s) for the dramatic effect of pH are not
clear at present. However, it has been reported that the
activity of a high molecular weight PBP in Streptococcus
pneumoniae, homologous to E. coli transpeptidases PBPs 2
and 3, is sensitive to extremes of pH (Laible et al., 1989;
Thomas et al., 2001). Therefore, it is possible that as the pH
increases during batch culture (see Farrell & Finkel, 2003)
PBP1b’s transpeptidase activity becomes critical as the
activity of the other PBPs may be compromised.
Another explanation for the SPCD phenotype displayed by
mrcB strains is increased sensitivity to extracellular factors,
such as toxins or degradative enzymes produced by the parent
cell line but not by the mrcB mutant. To investigate this
possibility, conditioned medium experiments were performed
(see Materials and methods) in which mrcB mutant cells were
inoculated into cell-free medium comprised of the filtrate of
previously cultured wild-type or PBP1b mutant bacteria. As
no difference in the long-term survival kinetics of wild-type
or mutant strains was observed over a 10-day period (data not
shown), it seems unlikely that toxin production plays a direct
role in the observed SPCD phenotype. However, conditioned
media experiments did reveal a significant effect on doubling
time during outgrowth (see below).
We propose that PBP1b mutants display the SPCD
phenotype for several reasons. First, mutants may less
efficiently construct or maintain their cell walls, leaving the
bacterium more susceptible to osmotic and alkaline stress;
consequently, a greater proportion of mutant cells may die
per unit time when compared with the parental strain
during stationary phase. Also, assays performed in both LB
and conditioned media indicate that average doubling times
increase for PBP1b mutants compared with wild-type cells
(Fig. 5). In LB medium, mrcB mutant doubling times are
increased by c. 36% (compare 28 vs. 38 min for the mutant;
Fig. 5a). In conditioned medium, with limited nutrients and
alkaline pH equivalent to that of stationary phase, the
average doubling time of the PBP1b mutant is �42% longer
(85 min for the mutant strain compared with 60 min for the
log
CF
U m
L−1
3
5
7
9
3
5
7
9
3
5
7
9
Day
0 7 14 21
0 7 14 21
0 7 14 21
(a)
(b)
(c)
Fig. 4. Representative competitions between wild-type (ZK1143, solid
lines) and PBP1b mutant strains (dotted lines) in LB medium under (a)
acidic, pH 5, (b) neutral, pH 7, or (c) basic, pH 9, conditions. Three
representative competitions are shown for each experiment. The limit of
detection is o 1000 CFU mL�1.
FEMS Microbiol Lett 263 (2006) 61–67 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
65PBP1b is required for competitive stationary-phase survival

wild-type). If extracellular toxin production were contribut-
ing to the SPCD phenotype of mrcB mutants, we would
expect a greater difference in relative growth rates in condi-
tioned medium as compared with fresh LB due to accumu-
lation of potentially toxic extracellular compounds during
long-term stationary-phase incubation. However, in both
LB and conditioned media the PBP1b mutant strain shows
an �40% increase in doubling time compared with wild-
type cells, suggesting no such role for extracellular factors.
Both a decreased rate of cell division (especially during
stationary phase) and an increased death rate due to altered
sensitivity to environmental stressors can affect the ability of
cells to compete and may lead to the eventual extinction of
PBP1b mutants when cocultured with wild-type strains. In
general, functional redundancy may be a reason that most
PBP mutants display no obvious phenotype (Young, 2001).
A physiological function for PBP5 was realized only when
other PBP genes had also been deleted (Nelson & Young,
2000). Additional studies employing strains harboring mul-
tiple PBP mutations may reveal other in vivo functions of
this important group of bacterial proteins.
Acknowledgements
We thank Vyacheslav Palchevskiy, Sudha Nair, and Gary
Nord for technical assistance, Alison Alcott and two re-
viewers for comments on the manuscript, and Dr Kevin
Young for generously providing plasmids. This research was
supported in part by the USC/Norris Comprehensive Can-
cer Center. E.D.P. is a recipient of a Fletcher Jones Founda-
tion Fellowship in the Biological Sciences and a National
Institute on Aging Predoctoral Training Grant
(#5T32AG00093-22).
References
Amanuma H & Strominger JL (1980) Purification and properties
of penicillin-binding proteins 5 and 6 from Escherichia coli
membranes. J Biol Chem 255: 11173–11180.
Baquero MR, Bouzon M, Quintela JC, Ayala JA & Moreno F
(1996) dacD, an Escherichia coli gene encoding a novel
penicillin-binding protein (PBP6b) with DD-
carboxypeptidase activity. J Bacteriol 178: 7106–7111.
Chalut C, Charpentier X, Remy MH & Masson JM (2001)
Differential responses of Escherichia coli cells expressing
cytoplasmic domain mutants of penicillin-binding protein 1b
after impairment of penicillin-binding proteins 1a and 3.
J Bacteriol 183: 200–206.
Datsenko KA & Wanner BL (2000) One-step inactivation of
chromosomal genes in Escherichia coli K-12 using PCR
products. Proc Natl Acad Sci USA 97: 6640–6645.
Denome SA, Elf PK, Henderson TA, Nelson DE & Young KD
(1999) Escherichia coli mutants lacking all possible
combinations of eight penicillin binding proteins: viability,
characteristics, and implications for peptidoglycan synthesis.
J Bacteriol 181: 3981–3993.
Farrell MJ & Finkel SE (2003) The growth advantage in
stationary-phase phenotype conferred by rpoS mutations is
dependent on the pH and nutrient environment. J Bacteriol
185: 7044–7052.
Finkel SE (2006) Long-term survival during stationary phase:
evolution and the GASP phenotype. Nat Rev Microbiol 4:
113–120.
Finkel SE & Kolter R (1999) Evolution of microbial diversity
during prolonged starvation. Proc Natl Acad Sci USA 96:
4023–4027.
Finkel SE & Kolter R (2001) DNA as a nutrient: novel role for
bacterial competence gene homologs. J Bacteriol 183: 6288–6293.
Garcia del Portillo F & de Pedro MA (1990) Differential effect of
mutational impairment of penicillin-binding-proteins 1A and
1B on Escherichia coli strains harboring thermosensitive
mutations in the cell division genes ftsA, ftsQ, ftsZ, pbpB.
J Bacteriol 172: 5863–5870.
Ghuysen JM & Dive G (1994) Biochemistry of the penicilloyl
serine transferases. Bacterial Cell Wall (Ghuysen JM &
Hakenbeck R, eds), pp. 103–129. Elsevier Science B.V.,
Amsterdam.
Henderson TA, Young KD, Denome SA & Elf PK (1997) AmpC
and AmpH, proteins related to the class C b-lactamases, bind
penicillin and contribute to the normal morphology of
Escherichia coli. J Bacteriol 179: 6112–6121.
Korat B, Mottl H & Keck W (1991) Penicillin-binding protein 4 of
Escherichia coli: molecular cloning of the dacB gene, controlled
overexpression, and alterations in murein composition. Mol
Microbiol 5: 675–684.
Laible G, Hakenbeck R, Sicard MA, Joris B & Ghuysen JM (1989)
Nucleotide sequences of the pbpX genes encoding the
penicillin-binding-proteins 2x from Streptococcus pneumoniae
R6 and a cefotaxime-resistant mutant, C506. Mol Microbiol 3:
1337–1348.
Livermore DM (1995) b-Lactamases in laboratory and clinical
resistance. Clin Microbiol Rev 8: 557–584.
Matsuhashi M, Tamaki S, Curtis SJ & Strominger JL (1979)
Mutational evidence for identity of penicillin-binding protein
Outgrowth
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00
100.00
WT/LB PBP1b−/LB WT/C.M. PBP1b−/C.M.
Strain/Media
Fig. 5. Comparison of wild-type (ZK1143) and PBP1b mutant growth
rates (measured in min) in LB or 3-day-old conditioned medium (CM).
The average of six trials is shown with standard error indicated.
FEMS Microbiol Lett 263 (2006) 61–67c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
66 E.D. Pepper et al.

5 in Escherichia coli with the major D-alanine carboxypeptidase
IA activity. J Bacteriol 137: 644–647.
Nelson DE & Young KD (2000) Penicillin binding protein 5
affects cell diameter, contour, and morphology of Escherichia
coli. J Bacteriol 182: 1714–1721.
Romeis T & Holtje JV (1994) Penicillin-binding protein 7/8 of
Escherichia coli is a DD-endopeptidase. Eur J Biochem 224:
597–604.
Schiffer G & Holtje JV (1999) Cloning and characterization of
PBP 1C, a third member of the multimodular class A
penicillin-binding proteins of Escherichia coli. J Biol Chem 274:
32031–32039.
Spratt BG (1975) Distinct penicillin binding proteins involved in
the division, elongation, and shape of Escherichia coli K12. Proc
Natl Acad Sci USA 72: 2999–3003.
Suzuki H, Nishimura Y & Hirota Y (1977) On the process of
cellular division in Escherichia coli: a series of mutants of E. coli
altered in the penicillin-binding proteins. Proc Natl Acad Sci
USA 75: 664–668.
Thomas B, Wang Y & Stein RL (2001) Kinetic and
mechanistic studies of penicillin-binding protein 2x
from Streptococcus pneumoniae. Biochemistry 40:
15811–15823.
Young KD (2001) Approaching the physiological functions of
penicillin-binding proteins in Escherichia coli. Biochimie 83:
9–102.
Zambrano MM, Siegele DA, Almiron M, Tormo A & Kolter R
(1993) Microbial competition: Escherichia coli mutants
that take over stationary phase cultures. Science 259:
1757–1760.
FEMS Microbiol Lett 263 (2006) 61–67 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
67PBP1b is required for competitive stationary-phase survival




























