Embed Size (px)
Citation preview

Revegetation and Nitrate Leaching from Lake States Northern Hardwood ForestsFollowing Harvest
Thomas M. Iseman, Donald R. Zak,* William E. Holmes, and Amy G. Merrill
ABSTRACTThe sugar maple (Acer saccharum Marshall)-rcd oak (Quercus
rubra L.) and sugar maple-basswood (Ti/ia americana L.) ecosystemsare Lake States forests that differ in net nitrification (5 and 15 g Nm"2 yr~', respectively), but experience equivalent rates of NOf leach-ing following clear-cut harvest (=5 g N m 2 yr '). Our objectives wereto determine whether high rates of N leaching are sustained followingharvest and whether ecosystem-specific patterns of biomass accumula-tion influence NO., loss. We studied two stands in each ecosystemand established four research plots in each stand; two plots were clear-cut in 1991 and two were controls. In 1996, we measured soil solutionNOf concentration (1-m depth) and calculated area! losses by a waterbalance method. We used allometric equations to estimate woodybiomass in clearcut plots; herbaceous biomass was clipped. In thesugar maple-red oak ecosystem, NO, leaching from 5-yr-oId clear-cut plots (0.56 g N m 2 yr ') was significantly greater than leachingfrom control plots (0.05 g N m~2 yr"1). In contrast, NOf leaching didnot differ between control (0.41 g N m"2 yr"1) and 5-yr-old clear-cut(0.02 g N m~2 yr~') in the sugar maple-basswood ecosystem; however,loss from these clear-cut plots was significantly lower than that fromclear-cut sugar maple-red oak plots. Five years after harvest, 7.1 Mgha"1 of aboveground biomass accumulated in clear-cut sugar maple-basswood plots, almost twice that of clear-cut sugar maple-red oakplots (3.9 Mg ha"'). Five years after harvest, the highest rates ofNOf loss occurred in the sugar maple-red oak ecosystem, in whichaboveground biomass accumulation was least.
CLEAR-CUT HARVEST can dramatically alter nutrientdynamics in forest ecosystems, particularly in for-
ests where rates of N cycling are rapid. Because of thewide geographic extent of northern hardwood forestsin eastern North America and their rapid rates of Ncycling, the effects of clear-cut harvest on nutrient cy-cling and loss from these ecosystems have been inten-sively studied, especially the leaching of NO3~ followingoverstory harvest (Bormann and Likens, 1979; Vitousekand Melillo, 1979; Hornbeck and Kropelin, 1982). Im-mediately following clear-cut harvest, direct insolationcauses an increase in soil temperature, while a declinein evapotranspiration results in an increase in soil waterstatus. These conditions promote net N mineralizationand nitrification (Matson and Vitousek, 1981), whileplant N uptake is simultaneously reduced by overstoryremoval (Vitousek et al., 1979). In the absence of plantuptake, NO-T accumulating in soil solution can be lostfrom forest ecosystems through leaching below the root-ing zone, a process that is exacerbated by augmentedflux of water resulting from reduced evapotranspiration(Vitousek, 1977; Likens et al., 1978).
Revegetation following clear-cut harvest in northern
School of Natural Resources & Environment, Univ. of Michigan,Ann Arbor, MI, 48109-1115. Received 10 June 1998. ""Correspondingauthor ([email protected]).
Published in Soil Sci. Soc. Am. J. 63:1424-1429 (1999).
hardwood forests functions as a sink for N, diminishingN loss from ecosystems following disturbance (Marksand Bormann, 1972; Vitousek and Reiners, 1975). Earlysuccessional species, such as red raspberry (Rubusideaus L.) and pin cherry (Prunuspensylvanica L.), takeadvantage of reduced competition for light and nutrientsto densely colonize clear-cut areas (Marks, 1974; Whit-ney, 1982; Mou et al., 1993). Loss of N is reduced orminimized by rapid accumulation of biomass, which uti-lizes available soil N (Vitousek, 1977). In addition, theproduction of NO3~ often declines as enhanced rates ofdecomposition reduce labile soil organic matter, thusslowing net N mineralization and nitrification (Likenset al., 1978; Fisk and Fahey, 1990). Bormann and Likens(1979) observed a significant reduction in NO3~ leachingwithin 2 yr following harvest in northern hardwood for-ests of New Hampshire. Subsequent studies in theNortheast confirmed that elevated N loss from northernhardwood forests immediately following harvest de-clines to preharvest levels within 2 to 5 yr (Hornbeckand Kropelin, 1982; Mann et al., 1988).
This study was a continuation of our investigationinto the effects of clear-cut harvest on N dynamics in twofloristically and functionally distinct northern hardwoodecosystems located in northern Lower Michigan. Thesugar maple-red oak/Maianthemum G.H. Weber exWiggers and the sugar maple-basswood/Ctemor/zz'zaRaf. ecosystems differ in rates of net nitrification, pre-senting the possibility that they also could differ in ratesof NOs" leaching following harvest (Zak et al., 1986;Holmes and Zak, 1994). In December 1991, plots withineach ecosystem were clear-cut harvested and N leachingwas measured in the growing season immediately fol-lowing harvest (1992). Despite initial differences in netnitrification, both the sugar maple-red oak/Maian-themum and sugar maple-basswood/Osraor/i/za ecosys-tems had equally high rates of N leaching the first yearfollowing harvest (=5 g m"2 yr"1; Holmes and Zak,1999). Our objectives were (i) to determine the extentto which high rates of N leaching are sustained followingclear-cut harvest in these Lake States northern hard-wood forests and (ii) to assess whether ecosystem-spe-cific differences in aboveground biomass accumulationlead to differences in rates of N leaching followingclear-cut.
METHODSStudy Area
Our study was conducted in two northern hardwood ecosys-tems which are widely distributed throughout northern LowerMichigan: the sugar maple-bassv/ood/Osmorhiza ecosystemand the sugar maple-red oak/Maianthemum ecosystem (here-after referred to as "sugar maple-basswood" and "sugar ma-
1424

ISEMAN ET AL.: REVEGETATION AND NITRATE LEACHING FOLLOWING HARVEST IN NORTHERN FORESTS 1425
pie-red oak"). These ecosystems were classified by a multifac-tor site classification method, using relationships amonglandform, soil, and vegetation to delineate functionally distinctecological units (Barnes et al., 1982). The sugar maple-basswood and sugar maple-red oak ecosystems are primarilydistinguished from one another by differences in overstorycomposition, edaphic properties, and patterns of N cycling(Table 1). Although annual rates of net N mineralization areequivalent between the two ecosystems, annual rates of netnitrification are significantly greater in the sugar maple-basswood ecosystem (14.8 g N m~2 yr^1) than in the sugarmaple-red oak ecosystem (5.4 g N m"2 yr"1; Holmes andZak, 1994).
As part of previous work, two stands representative of eachecosystem were randomly located within the Manistee Na-tional Forest in northwestern Lower Michigan (Zak et al.,1986). The landscape is characteristic of the Interlobate mo-raine. Hilly topography and well-drained, sandy soils predomi-nate in this region. All stands are subject to similar climaticconditions throughout the year: mean annual temperature is6.7°C, growing seasons last =115 d, and annual precipitationaverages 77 cm (Albert et al., 1986). Because of its proximityto Lake Michigan and relatively high elevation (300-450 m),this region accumulates substantial snowpack in winter.
Within each stand, four plots (5 by 30 m) were randomlylocated (Zak and Pregitzer, 1990). Two of the four plots withineach stand were randomly selected for clear-cut harvest; theother two plots within each stand were left intact to serve ascontrols. Clear-cut harvest took place in December 1991, whenstems >10 cm diameter at breast height were harvested andremoved. Branches and stems <10 cm diameter were distrib-uted throughout the clear-cut area. In order to minimize theinfluence of adjacent vegetation on the clear-cut plots, a tree-height buffer strip (22.5 m) surrounding each clear-cut plotwas also harvested, such that the total harvested area was 50by 75 m (0.375 ha). A trench (0.25 m wide and 1.3 m deep)was excavated 15 m from the perimeter of each clear-cutplot to prevent N uptake by roots of adjacent overstory trees(Holmes and Zak, 1999). Natural revegetation of clear-cutplots has been allowed to occur.
Nitrogen LeachingWithin each plot, four ceramic cup lysimeters were installed
in November 1989 (Merrill, 1991). The lysimeters were locatedat 10-m intervals along the length of each plot. They wereinstalled to collect soil water at a depth of 1 m beneath thesoil surface, a depth which is below the majority of fine rootsin many upland forest ecosystems (Gale and Grigal, 1987). Atension of 35 kPa was applied to the lysimeters at each sam-pling date. Soil water samples were collected monthly duringthe snow-free months prior to harvest (1990-1991; Merrill,1991), the year following harvest (1992-1993; Holmes andZak, 1999), and 5 yr after harvest (1996-1997). Concentrationof NO3~-N in soil water was determined using an Alpkem 300Rapid Flow Analyzer (Astoria-Pacific International, Clacka-mas, OR).
A water budget method was used to determine the volumeof water moving below the rooting zone on a monthly basis(Thornthwaite and Mather, 1957). Mean monthly temperatureand latitude were used to calculate monthly potential evapo-transpiration. Actual evapotranspiration was estimated on amonthly basis by budgeting the available water (precipita-tion + soil water storage) vs. potential evapotranspiration.When potential evapotranspiration was satisfied by monthlyprecipitation and soil water storage, excess water was assumedto move below the rooting zone, transporting NO3~-N from
Table 1. Selected overstory, soil, and N-cycling characteristics oftwo northern hardwood forests.t________________
Sugar maple-Sugar maple-red oak/ basswood/
Maianthemum Osmorhiza
OverstoryAge, yr 86 91Density, stems h;i ' 950 642Biomass, Mg ha ' 260 242Annual biomass increment,
Mineral soil (0-10 cm)PHOrganic C, mg g '
N-cycling, g N m~2 yr 'Atmospheric N deposition
Wet + dryNet N mineralizationNitrification
2.8
4.613
1.213.05.4
3.3
6.326
1.214.314.8
t Data are summarized from Zak and Pregitzer (1990) and Holmes andZak (1994,1999).
the ecosystems. Areal leaching losses NO3~-N (g N m"2 mo"1)were calculated on a monthly basis as the product of N concen-tration and volume of water moving below the rooting zone.Climatic data were obtained from the Cadillac weather station,Wexford County, MI, operated by the USDA Forest Service.
Aboveground BiomassAboveground biomass was estimated for each clear-cut plot
5 yr following overstory harvest. All aboveground vegetationwas clipped by hand from four randomly located samplingframes (0.75 by 0.75 m) within each clear-cut plot. Species-specific biomass equations were used to estimate abovegroundbiomass of stump-sprouted black cherry (Prunus serotinaEhrh.) that had grown too large to harvest (Smith and Brand,1983). Due to the highly patchy distribution of vegetation,four additional samples were collected from within each ofthe clear-cut plots in one sugar maple-red oak stand (totalsamples per plot = 8). Biomass samples were oven dried at70°C and weighed in the laboratory. Biomass was expressedin kilograms per hectare and averaged by plot.
Statistical AnalysesDifferences in annual rates of NO3~ leaching (g N m'2 yr"1)
5 yr following clear-cut harvest were analyzed using a partiallycrossed, partially nested analysis of variance. Stands (n = 2per ecosystem) were nested within ecosystems (n = 2), whichwere crossed with treatments (« = 2). Ecosystem and treat-ment were fixed factors and stand was a random factor. Meanswere compared by Fisher's protected LSD. A repeated mea-sures analysis of variance with a similar design was used tocompare temporal changes in annual N leaching rates acrossthree sampling intervals: preharvest (1990-1991), 1 yr follow-ing harvest (1992-1993), and 5 yr postharvest (1996-1997).Differences in aboveground biomass in clear-cut plots werecompared using a nested analysis of variance (stands nestedwithin ecosystems). Statistical results were accepted as signifi-cant at P < 0.05.
RESULTS AND DISCUSSIONNitrogen Leaching
Temporal patterns of NO-T leaching in both ecosys-tems were similar during the 3 yr. The majority ofNO3~ export occurred during fall and early spring (Fig.1 and 2), periods when water inputs exceeded soil water
1426 SOIL SCI. SOC. AM. J., VOL. 63, SEPTEMBER-OCTOBER 1999
15-
coro 10-<DO
8 5-COO
CNI
E
03
ocoCD
co
2-
.1-
Sugar maple-red oakD intact• clear-cut
8 5
J. I
t .i
B
>* c -TO 13 -
.=J 05
>> C ~g D) Q--ft > feC O S ^ D C B X O 9-
1 yr pre-harvest 1 yr post-harvest 5 yr post-harvest1990-91 1992-93 1996-97
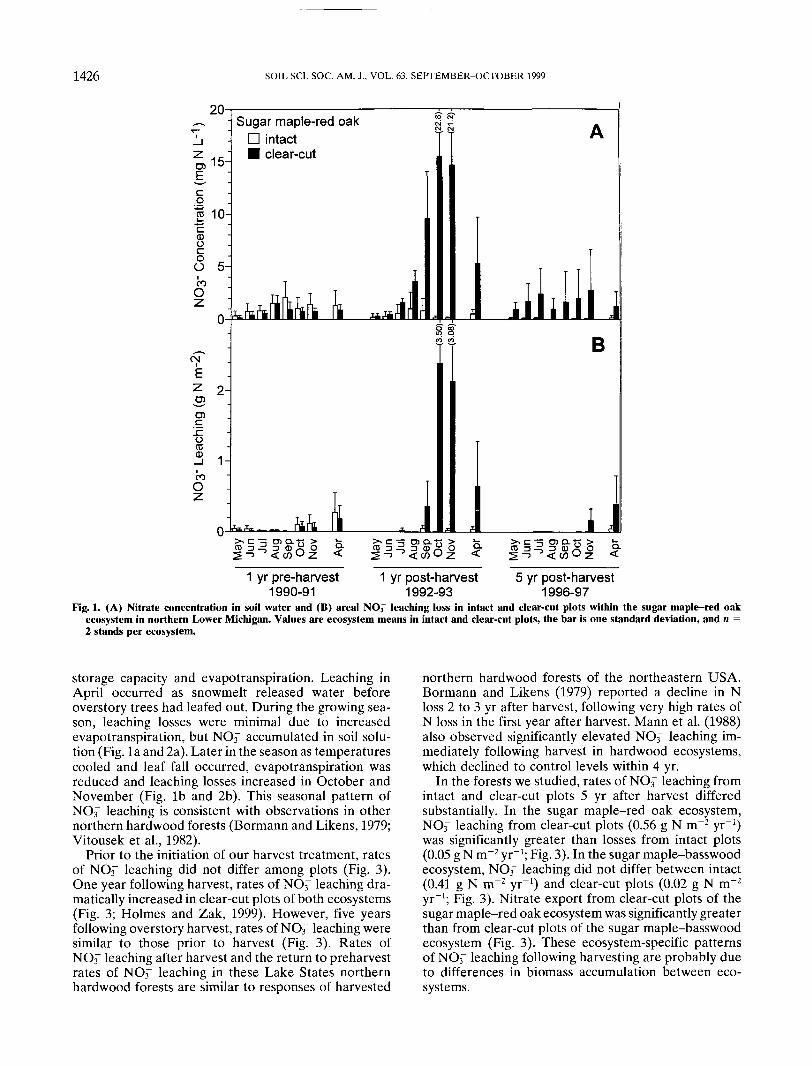
Fig. 1. (A) Nitrate concentration in soil water and (B) area! NOf leaching loss in intact and clear-cut plots within the sugar maple-red oakecosystem in northern Lower Michigan. Values are ecosystem means in intact and clear-cut plots, the bar is one standard deviation, and n =2 stands per ecosystem.
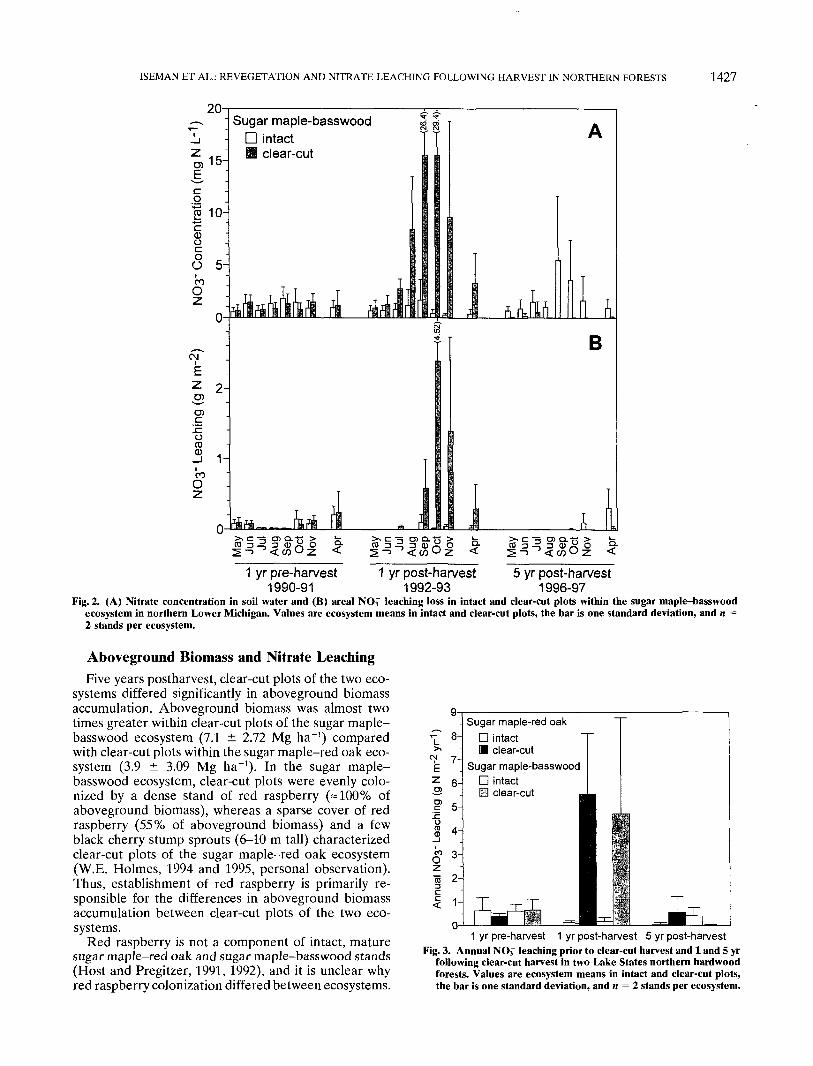
storage capacity and evapotranspiration. Leaching inApril occurred as snowmelt released water beforeoverstory trees had leafed out. During the growing sea-son, leaching losses were minimal due to increasedevapotranspiration, but NOi" accumulated in soil solu-tion (Fig. la and 2a). Later in the season as temperaturescooled and leaf fall occurred, evapotranspiration wasreduced and leaching losses increased in October andNovember (Fig. Ib and 2b). This seasonal pattern ofNO3~ leaching is consistent with observations in othernorthern hardwood forests (Bormann and Likens, 1979;Vitousek et al., 1982).
Prior to the initiation of our harvest treatment, ratesof NO3~ leaching did not differ among plots (Fig. 3).One year following harvest, rates of NO3~ leaching dra-matically increased in clear-cut plots of both ecosystems(Fig. 3; Holmes and Zak, 1999). However, five yearsfollowing overstory harvest, rates of NO3~ leaching weresimilar to those prior to harvest (Fig. 3). Rates ofNO3~ leaching after harvest and the return to preharvestrates of NO-T leaching in these Lake States northernhardwood forests are similar to responses of harvested
northern hardwood forests of the northeastern USA.Bormann and Likens (1979) reported a decline in Nloss 2 to 3 yr after harvest, following very high rates ofN loss in the first year after harvest. Mann et al. (1988)also observed significantly elevated NO3~ leaching im-mediately following harvest in hardwood ecosystems,which declined to control levels within 4 yr.
In the forests we studied, rates of NO3~ leaching fromintact and clear-cut plots 5 yr after harvest differedsubstantially. In the sugar maple-red oak ecosystem,NO-T leaching from clear-cut plots (0.56 g N m~2 yr"1)was significantly greater than losses from intact plots(0.05 g N m~2 yr"1; Fig. 3). In the sugar maple-basswoodecosystem, NOr leaching did not differ between intact(0.41 g N m"2 yr-1) and clear-cut plots (0.02 g N m~2
yr"1; Fig. 3). Nitrate export from clear-cut plots of thesugar maple-red oak ecosystem was significantly greaterthan from clear-cut plots of the sugar maple-basswoodecosystem (Fig. 3). These ecosystem-specific patternsof NO3~ leaching following harvesting are probably dueto differences in biomass accumulation between eco-systems.
ISEMAN ET AL.: REVEGETATION AND NITRATE LEACHING FOLLOWING HARVEST IN NORTHERN FORESTS 1427
o'"TO 10-•«—'
Q)O
3 5HroO
D)
o030)
00O
2-
1-
Sugar maple-basswoodD intact• clear-cut
i
Fifii __
B
1 yr pre-harvest 1 yr post-harvest 5 yr post-harvest1990-91 1992-93 1996-97
Fig. 2. (A) Nitrate concentration in soil water and (B) area! NOf leaching loss in intact and clear-cut plots within the sugar maple-basswoodecosystem in northern Lower Michigan. Values are ecosystem means in intact and clear-cut plots, the bar is one standard deviation, and n =2 stands per ecosystem.
Aboveground Biomass and Nitrate LeachingFive years postharvest, clear-cut plots of the two eco-
systems differed significantly in aboveground biomassaccumulation. Aboveground biomass was almost twotimes greater within clear-cut plots of the sugar maple-basswood ecosystem (7.1 ± 2.72 Mg ha"1) comparedwith clear-cut plots within the sugar maple-red oak eco-system (3.9 ± 3.09 Mg ha"1)- In the sugar maple-basswood ecosystem, clear-cut plots were evenly colo-nized by a dense stand of red raspberry (=100% ofaboveground biomass), whereas a sparse cover of redraspberry (55% of aboveground biomass) and a fewblack cherry stump sprouts (6-10 m tall) characterizedclear-cut plots of the sugar maple-red oak ecosystem(W.E. Holmes, 1994 and 1995, personal observation).Thus, establishment of red raspberry is primarily re-sponsible for the differences in aboveground biomassaccumulation between clear-cut plots of the two eco-systems.
Red raspberry is not a component of intact, mature 1 yr Pre'harvest 1 ̂ P°st-harvest 5 yr post-harvestsiKiar manle rerl oalc anrl <jn»ar manle hasswnnH stands Fig'3' Annual NO' leaching prior to clear-cut harvest and 1 and S yrsugar maple-red oak and sugar maple-basswood stands following clear-cut harvest in two Lake States northern hardwood(Host and Pregltzer, 1991, 1992), and it IS unclear why forests. Values are ecosystem means in intact and clear-cut plots,red raspberry colonization differed between ecosystems. the bar is one standard deviation, and n = 2 stands per ecosystem.
Sugar maple-red oakD intact
clear-cutSugar maple-basswood
D intactclear-cut

1428 SOIL SCI. SOC. AM. J., VOL. 63, SEPTEMBER-OCTOBER 1999
However, several factors may have affected differencesin establishment between clear-cut plots of these ecosys-tems. We know that red raspberry colonization of dis-turbed sites: (i) increases with intensity of disturbance(Roberts and Dong, 1993), (ii) increases with greaterlight availability (Mou et al., 1993; Ricard and Messier,1996), and (iii) depends on germination of buried seeds(Hughes and Fahey, 1991). The sugar maple-red oakecosystem has moderately thick Oi and Oe (4-6 cm)horizons overlying the mineral soil, whereas in the sugarmaple-basswood ecosystem there is little forest floordevelopment (only a thin, discontinuous Oi horizon)and a thorough mixing of organic matter into mineralsoil by fauna. The relatively thick Oi and Oe horizonsin sugar maple-red oak ecosystem, although disturbedsomewhat by harvesting, may have limited the germina-tion and establishment of red raspberry in clear-cut plotsof this ecosystem.
Plant uptake is the primary mechanism of nutrientretention in regenerating forests. Available nutrients areused and retained by accumulating biomass followingdisturbance (Bormann and Likens, 1979). Red rasp-berry is an early-successional species, which rapidly col-onizes disturbed northern hardwood sites, using avail-able nutrients and growing vigorously under conditionsof high soil moisture and light availability soon afterharvest (Marks, 1974; Whitney, 1982). This species typi-cally thrives for 2 to 6 yr following harvest, declining asnutrient and light availability diminishes. Tilman (1987)found that Rubus spp. became more abundant in re-sponse to increased soil N availability, and Truax et al.(1994) report that red raspberry is particularly adaptedto NO3~ uptake. Several studies suggest that red rasp-berry acts to limit nutrient loss, particularly NO3, imme-diately following disturbance, when forests are mostsusceptible to nutrient loss (Bormann et al., 1974; Horn-beck and Kropelin, 1982; Mou et al., 1993). This patternis consistent with our observation that clear-cut plotswith greater biomass had lower rates of NO3~ leaching.The low rates of NO3~ leaching in clear-cut plots of thesugar maple-basswood ecosystem probably resultedfrom high demand for NO-T due to the dense growth ofred raspberry. Rates of NOs" leaching were significantlygreater within clear-cut plots of the sugar maple-redoak ecosystem, where red raspberry density and biomassaccumulation were lower.
In conclusion, the Lake States northern hardwoodforests we studied responded to harvesting in a mannersimilar to that observed in northern hardwood forestsof other regions. Nitrate leaching increased dramaticallyfollowing clear-cut harvest, but rates in intact and clear-cut plots were similar to preharvest rates after 5 yr. Inour study, ecosystem-specific patterns of biomass accu-mulation appear to control rates of NO-T leaching. Fiveyears postharvest, clear-cut plots of the sugar maple-redoak ecosystem had greater rates of NO^ leaching andlower aboveground biomass than clearcut plots of thesugar maple-basswood ecosystem. The density of redraspberry was greater in clear-cut plots of the sugarmaple-basswood ecosystem, which may have contrib-uted to the lower rates of NO-T leaching, since red rasp-berry has a high demand for NO3~.
ACKNOWLEDGMENTSThis manuscript is based on a portion of the thesis submitted
by the senior author in partial fulfillment of the requirementsof the Master of Science degree in the School of NaturalResources & Environment, University of Michigan. Our studywas supported by funds from the Mclntire-Stennis Coopera-tive Forestry Act (P.L. 87-788). We are grateful for the assis-tance of Matt Sands and Rose Ingram (USDA Forest Service,Manistee National Forest) and David Rothstein.

ANDREWS ET AL.: SEPARATION OF ROOT RESPIRATION FROM TOTAL SOIL RESPIRATION 1429
of red raspberry (Rubus ideaus L.) along a natural light gradientin a northern hardwood forest. For. Ecol. Manage. 81:153-160.
Roberts, M.R., and H. Dong. 1993. Effects of soil organic layer re-moval on regeneration after clear-cutting a northern hardwoodstand in New Brunswick. Can. J. For. Res. 23:2093-2100.
Smith, W.B., and G.J. Brand. 1983. Allometric biomass equations for98 species of herbs, shrubs, and small trees. USDA Forest Service:Research Note NC-299.
Thornthwaite, C.W., and J.R. Mather. 1957. Instructions and tablesfor computing potential evapotranspiration and the water balance.Climatology 10:185-311.
Tilman, D. 1987. Secondary succession and the pattern of plant domi-nance along experimental nitrogen gradients. Ecol. Monogr. 57:189-214.
Truax, B., D. Gagnon, F. Lambert, and N. Chevrier. 1994. Nitrateassimilation of raspberry and pin cherry in a recent clear-cut. Can.J. Bot. 72:1343-1348.
Vitousek, P.M. 1977. The regulation of element concentrations inmountain streams in the northeastern United States. Ecol. Mo-nogr. 47:65-87.
Vitousek, P.M., and W.A. Reiners. 1975. Ecosystem succession andnutrient retention: a hypothesis. BioScience 25:376-381.
Vitousek, P.M., and J.M. Melillo. 1979. Nitrate losses from disturbedforests: patterns and mechanisms. For. Sci. 25:605-619.
Vitousek, P.M., J.R. Gosz, C.C. Grier, J.M. Melillo, and W.A. Reiners.1982. A comparative analysis of potential nitrification and nitratemobility in forest ecosystems. Ecol. Monogr. 52:155-177.
Vitousek, P.M., J.R. Gosz, C.C. Grier, J.M. Melillo, W.A. Reiners,and R.L. Todd. 1979. Nitrate losses from disturbed ecosystems.Science 204:469^(74.
Whitney, G.G. 1982. The productivity and carbohydrate economy ofa developing stand of Rubus ideaus. Can. J. Bot. 60:2697-2703.
Zak, D.R., and K.S. Pregitzer. 1990. Spatial and temporal variability ofnitrogen cycling in northern Lower Michigan. For. Sci. 36:367-380.
Zak, D.R., K.S. Pregitzer, and G.E. Host. 1986. Landscape variationin nitrogen mineralization and nitrification. Can. J. For. Res.16:1258-1263.





























