Embed Size (px)
Citation preview

Research ArticleIron as the Key Modulator of Hepcidin Expression in ErythroidAntibody-Mediated Hypoplasia
J C Fernandes12 P Garrido1 S Ribeiro23 P Rocha-Pereira24 E Bronze-da-Rocha23
L Belo23 E Costa23 F Reis1 and A Santos-Silva23
1Laboratory of Pharmacology and Experimental Therapeutics IBILI Faculty of Medicine University of Coimbra3000-548 Coimbra Portugal2Institute for Molecular and Cellular Biology University of Porto 4150-180 Porto Portugal3Department of Biological Sciences Faculty of Pharmacy University of Porto Rua de Jorge Viterbo Ferreira No 2284050-313 Porto Portugal4Research Centre in Health Sciences University of Beira Interior 6201-001 Covilha Portugal
Correspondence should be addressed to A Santos-Silva assilvaffuppt
Received 23 June 2014 Accepted 12 October 2014 Published 18 December 2014
Academic Editor Beatrice Charreau
Copyright copy 2014 J C Fernandes et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Erythroid hypoplasia (EH) is a rare complication associated with recombinant human erythropoietin (rHuEPO) therapies due todevelopment of anti-rHuEPO antibodies however the underlying mechanisms remain poorly clarified Our aim was to manage arat model of antibody-mediated EH induced by rHuEPO and study the impact on iron metabolism and erythropoiesis Wistarrats treated during 9 weeks with a high rHuEPO dose (200 IU) developed EH as shown by anemia reduced erythroblastsreticulocytopenia and plasmatic anti-rHuEPO antibodies Serum iron was increased and associated with mRNA overexpressionof hepatic hepcidin and other iron regulatory mediators and downregulation of matriptase-2 overexpression of divalent metaltransporter 1 and ferroportin was observed in duodenum and liver Decreased EPO expression was observed in kidney and liverwhile EPO receptor was overexpressed in liver Endogenous EPO levels were normal suggesting that anti-rHuEPO antibodiesblunted EPO function Our results suggest that anti-rHuEPO antibodies inhibit erythropoiesis causing anemia This leads to aserum iron increase which seems to stimulate hepcidin expression despite no evidence of inflammation thus suggesting iron asthe key modulator of hepcidin synthesisThese findings might contribute to improving new therapeutic strategies against rHuEPOresistance andor development of antibody-mediated EH in patients under rHuEPO therapy
1 Introduction
Erythropoietin (EPO) a 304 kDa glycoprotein is a keyhormone in the regulation of erythropoiesis supportingproliferation survival and terminal differentiation of ery-throid progenitor cells in the bone marrow [1 2] Theseeffects are mediated through the interaction of EPO with itsspecific transmembrane receptormdasherythropoietin receptor(EPOR) [3] Acquired EPO deficiency has been associatedwith chronic kidney disease (CKD) and with chronic inflam-matory diseases Treatment with recombinant human ery-thropoietin (rHuEPO) achieves correction of these types ofanemia however 5ndash10 of CKD patients develop resistanceto the erythropoietic stimuli of rHuEPO Several factors have
been proposed to play a role in the development of thisresistance but the etiology of the impaired erythropoiesisleading to worsening of the anemia or even to pure redcell aplasia (PRCA) is still unknown [4] Casadevall et al(1996) reported the presence of anti-EPO antibodies in apatient with transient PRCA which functionally blockedthe interaction between EPO and EPOR thus resulting inimpaired erythropoiesis [5] Hara et al (2008) also reportedthat serum from a patient with PRCA inhibited EPO-dependent cell proliferation suggesting that the anemia wasmediated by anti-EPO antibodies [6] In several other casesantibodies have been described in PRCA patientsrsquo sera thatwere selectively cytotoxic for marrow erythroid cells or weredirected against EPO [7ndash9]
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 421304 10 pageshttpdxdoiorg1011552014421304
2 BioMed Research International
The current therapeutic use of rHuEPO to correct anemiain CKD patients has been associated with some cases ofPRCA due to the development of cross-reactive anti-EPOantibodies [10 11] Although this complication is very rarethe mechanisms underlying the break in immune toleranceto rHuEPO remain poorly clarified The development ofPRCA should be suspected in patients who have been underrHuEPO therapy for more than 3 months and develop asudden severe unexplained fall in hemoglobin levels andreticulocytopenia and a marked decrease in bone marrowerythroblasts despite continuing with rHuEPO treatment[12] The diagnosis is confirmed by low serum EPO levelsand detection of anti-EPO antibodies [13] The productionof anti-EPO antibodies and the inhibition of EPO-dependenterythroid cells are important mechanisms that might leadto erythroid hypoplasia (EH)PRCA The reduction in func-tional EPO affects the proliferative state of bone marrowerythroid cells which will trigger several changes in ironmetabolism such as in serum iron transferrin and ferritinlevels among others [14ndash16]
Considering the scarce data in literature about howfacing a PRCA condition mediated by anti-EPO antibodiesthe depression of marrow erythroid activity affects ironmetabolism our goal was to assess in male Wistar rats theeffects of this condition on iron metabolism (iron absorptionand iron traffic and storage) as well as on erythropoiesisFor that purpose we performed hematological and biochem-ical studies to evaluate the erythropoietic and iron statusand measured the expression of several mRNA-encodingerythropoiesis and iron metabolism regulating proteins onduodenum liver and kidney tissues
2 Material and Methods
21 Animals and Experimental Protocol Male Wistar rats(Charles River Lab Inc Barcelona Spain) weighing 200ndash250 g weremaintained in an air-conditioned room subjectedto 12 h darklight cycles and given standard laboratory ratdiet (IPM-R20 Letica Barcelona Spain)without iron supple-mentation and free access to tap water Animal experimentswere conducted according to the European CommunitiesCouncil Directives on Animal Care These procedures werein accordance with the Association for Pharmacology andExperimental Therapeutics and were approved by the Insti-tutional Ethics Committee of the Faculty of Medicine fromthe University of Coimbra approval ID FMUC0413 Therats were divided into 3 groups (8 rats each) control treatedwith saline solution 50 IU rHuEPO a therapeutic dose of50 IUKg bodyweight (bw)week of epoetin beta (Recormon-Roche Pharmaceuticals) treatment 200 IU rHuEPO treatedwith 200 IUKg bwweek of rHuEPO a high dose usuallyused on rHuEPO resistant patients All the animals havecompleted the 9-week protocol (Figure 1)
22 Sample Collection and Preparation
221 Blood At the beginning of the experiments (1198790) and
at 3 (1198791) 6 (119879
2) and 9 (119879
3) weeks after the first rHuEpo
3 6 9Time (weeks)
0
administrationAnimals were
sacrificed
Blood sample were collected
Day 1
(50 and 200 IUkgweekthree timesweek sc)
Starting rHuEPO
T0 T1 T2 T3
Figure 1 Experimental protocol
dose the rats were subjected to intraperitoneal anesthesiawith a 2mgkg bwof a 2 1 (v v) 50mgmLketamine (KetalarParke-Davis Lab Pfizer Lda Seixal Portugal) solution in25 chlorpromazine (Largactil Rhone-Poulenc Rorer LabVitoria Amadora Portugal) Blood samples were immedi-ately collected by venipuncture from the jugular vein intovacutainer tubes without anticoagulant (to obtain serum)and with EDTA (to obtain whole blood and plasma) forhematological and biochemical studies 3mL blood sampleswere collected at 119879
0 1198791 and 119879
2to minimize interference
with erythropoiesis mechanism at the end of protocol (1198793)
a 10mL sample was collected in order to perform all thebiochemical and hematological assays
222 Tissues At the end of the protocol blood was col-lected and afterwards the rats were sacrificed by cervicaldislocation duodenum liver and kidneys were immediatelyremoved placed in ice-cold Krebs-Henseleit buffer andcarefully cleaned A bone marrow aspirate from the femurwas also performed
23 Biochemical and Hematological Assays Serum creati-nine and urea concentrations were used as renal functionindexes through automatic validatedmethods and equipment(Hitachi 717 analyser) Red blood cell (RBC) hematocritand hemoglobin (Hb) concentration were assessed in wholeblood EDTA by using an automatic Coulter Counter (Beck-man Coulter Inc CA USA) reticulocyte (RET) countwas performed by microscopic counting on blood smearsafter vital staining with New Methylene Blue (ReticulocyteStain Sigma-Aldrich St Louis MO USA) Serum levelsof erythropoietin were evaluated by rat specific ELISA kit(MyBioSource USA) Serum iron concentration was deter-mined using a colorimetric method (Iron Randox Labo-ratories Ltd UK) whereas serum ferritin and transferrinwere measured by immunoturbidimetry (Laboratories LtdUK) Quantification of total bilirubin was performed by acolorimetric test (diazotized sulfanilic acid reaction RocheDiagnostics) circulating levels of glucose and uric acid weredetermined by routine automated technology (ABX Diag-nostics) Serum levels of interleukin-6 (IL-6) interferon-120574(IFN-120574) transforming growth factor (TGF-1205731) and vascularendothelial growth factor (VEGF) were measured by rat-specific Quantikine ELISA kits from RampD Systems (Min-neapolis USA) High-sensitive C-reactive protein (hsCRP)
BioMed Research International 3
was determined by using a rat-specific ELISA kit from AlphaDiagnostics International (San Antonio USA)
24 Detection of Anti-EPO Antibodies The detection of anti-EPO antibodies was carried out by ELISA according to Urraet al 1997 using rHuEPO (Recormon Roche Pharmaceuti-cals) as antigen and as secondary antibody goat anti-rat IgGconjugated with horseradish peroxidase (Sigma 100 ngmLfor 1 h at room temperature) [17] The substrate tetramethyl-benzidine (TMB) (Sigma) was added and the reaction wasstopped by the addition of sulphuric acid 125molL Theoptical density at 450 nm (OD450) was determined with anautomatic plate reader
25 Gene Expression Analysis In order to isolate total RNA02 g of liver duodenum and kidney samples from each ratwas immersed in RNA later (Ambion Austin USA) uponcollection and stored at 4∘C for 24 h afterwards sampleswere frozen at minus80∘C Subsequently tissue samples weighing50 plusmn 10mg were homogenized in a total volume of 1mLTRI Reagent using a homogenizer and total RNA was iso-lated according to manufacturer instructions (Sigma SintraPortugal) To ensure inactivation of contaminating RNAsesmetal objects and glassware were cleaned with detergentimmersed in RNAse-free water (02 diethyl pyrocarbonate)for 2 h and finally heated at 120∘C for 1 h RNA integrity (RINRNA Integrity Number) was analyzed using 6000 Nano kitinAgilent 2100 bioanalyzer (Agilent TechnologiesWalbronnGermany) and 2100 expert software followingmanufacturerinstructions The yield from isolation was from 05 to 15 120583gRIN values were 78ndash90 and purity (A260A280) was 18ndash20The concentrations of the RNA preparations were confirmedwith NanoDrop1000 (ThermoScientific Wilmington DEUSA) Possible contaminating remnants of genomic DNAwere eliminated by treating these preparations with deoxyri-bonuclease I (amplification grade) prior to RT-qPCR ampli-fication Reverse transcription and relative quantification ofgene expression were performed as previously described [18]Real-time qPCR reactions were performed for the followinggenes EPO EPOR transferrin receptor 2 (TfR2) hepcidin(Hamp) ferroportin (SLC40A1) Hemojuvelin (HJV) trans-ferrin (TF) hemochromatosis (Hfe) divalent metal trans-porter 1 (DMT1) transferrin receptor 1 (TfR1) matriptase-2 (TMPRSS6) and bone morphogenic protein 6 (BMP6)which were normalized in relation to the expression of beta-actin (Actb) and 18S ribosomal subunit (18S) as well as tothe mean of the control group Primer sequences are listed inTable 1 Results were analyzed with SDS 21 software (AppliedBiosystems Foster City CA USA) and relative quantificationwas calculated using the 2minusΔΔCt method [19] In liver tissue westudied the EPO EPORTfR2Hamp SLC40A1HJVTFHfeBMP6 and TMPRSS6 gene expression in duodenum tissuethe gene expressions ofDMT1 and SLC40A1were studied andin the kidney we evaluated the expression of EPO gene
26 Data Analysis For statistical analysis we used theStatistical Package for Social Sciences (SPSS) version 220Results are presented as mean plusmn standard deviation Multiple
Table 1 List of primer sequences (F forward R reverse)
Gene Primer sequences
EPO F 51015840-AGGGTCACGAAGCCATGAAG-31015840
R 51015840-GAT TTC GGC TGT TGC CAG TG-31015840
EPOR F 51015840-GCG ACT TGG ACC CTC TCA TC-31015840
R 51015840-AGT TAC CCT TGT GGG TGG TG-31015840
Hamp F 51015840-GAA GGC AAG ATG GCA CTA AGC-31015840
R 51015840-CAG AGC CGT AGT CTG TCT CG-31015840
TfR2 F 51015840-CAA GCT TCG CCC AGA AGG TA-31015840
R 51015840-CGT GTA AGG GTC CCC AGT TC-31015840
SLC40A1 F 51015840-CAG GCT TAG GGT CTA CTG CG-31015840
R 51015840-CCG AAA GAC CCC AAA GGA CA-31015840
HJV F 51015840-GCC TAC TTC CAA TCC TGC GT-31015840
R 51015840-GGT CAA GAA GAC TCG GGC AT-31015840
TF F 51015840-GGC ATC AGA CTC CAG CAT CA-31015840
R 51015840-GCA GGC CCA TAG GGA TGT T-31015840
Hfe F 51015840-CTG GAT CAG CCT CTC ACT GC-31015840
R 51015840-GTC ACC CAT GGT TCC TCC TG-31015840
DMT1 F 51015840-CAA CTC TAC CCT GGC TGT GG-31015840
R 51015840-GTC ATG GTG GAG CTC TGT CC-31015840
TfR1 F 51015840-GCT CGT GGA GAC TAC TTC CG-31015840
R 51015840-GCC CCA GAA GAT GTG TCG G-31015840
TMPRSS6 F 51015840-CCG AAT ATG AGG TGG ACC CG-31015840
R 51015840-GGT TCA CGT AGC TGT AGC GG-31015840
BMP6 F 51015840-GCT GCC AAC TAT TGT GAC GG-31015840
R 51015840-GGT TTG GGG ACG TAC TCG G-31015840
18S F 51015840-CCA CTA AAG GGC ATC CTG GG-31015840
R 51015840-CAT TGA GAG CAA TGC CAG CC-31015840
Actb F 51015840-GAG ATT ACT GCC CTG GCT CC-31015840
R 51015840-CGG ACT CAT CGT ACT CCT GC-31015840
comparisons between groups were performed by one-wayANOVA supplemented with Tukeyrsquos HSD post hoc testFor single comparisons we used the Mann-Whitney 119880 testSignificance was accepted for a 119875minor than 005
3 Results
The changes in RBC count Hb concentration hematocritand the number of reticulocytes for the three groups arepresented in Figure 2 During the 9-week experimentalprotocol the 50 IU rHuEPO group when compared tothe control group showed similar values for RBC countHb concentration hematocrit and reticulocytes though asignificant increase in reticulocyte count was observed at 119879
2
and 1198793 Concerning the rats treated with 200 IU rHuEPO in
the first 3 weeks we found a significant increase (gt30 thebasal value) in hemoglobin levels RBC count hematocritand reticulocyte count as compared to control and 50 IUrHuEPO groups after this period the hemoglobin concen-tration hematocrit and RBC count as well as the number ofreticulocytes decreased reaching significantly lower values at1198792 as compared to 119879
1 this trend towards decreasing values
was maintained till the end of the experiment (9 weeks)
4 BioMed Research International
Control
3 6 90
200
400
600
800
Time (weeks)
3 6 95
6
7
8
9
10
11
3 6 920
30
40
50
60
Hem
atoc
rit (
)
10
12
14
16
18
20
Hem
oglo
bin
(gd
L)
0
RBC
(1012L
)
0Time (weeks) Time (weeks)
3 6 90 0Time (weeks)
Retic
uloc
ytes
(109L
)
50 IU + rHuEPO200 IU + rHuEPO Control
50 IU + rHuEPO200 IU + rHuEPO
yensect
120575cd
120575
lowast
sect
120575cd
lowast
sect
yensect
120575cd 120575cd
120575
120575
lowast
sect yen
a a
b
Figure 2 Erythrocyte data and reticulocyte count during the follow-up period of 9 weeks under rHuEPO treatment Results are expressedas mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO c119875 lt 0001 versus control d119875 lt 0001 versus 50 IU rHuEPO120575119875 lt 0005 1198790 versus 1198791
lowast119875 lt 005 1198791 versus 1198792
sect119875 lt 005 1198792 versus 1198793
At the end of the protocol serum erythropoietin levelswere similar among the three groups (Table 2) No changeswere observed in glucose creatinine urea uric acid andbilirubin levels for both rHuEPO groups along the entireprotocol when compared to the control The inflammatorymarker IL-6 presented a significant decrease in the 200 IUrHuEPO group when compared with the control and 50 IUrHuEPO groups hsCRP IFN-120574 TGF-1205731 and VEGF showedno significant differences between the three studied groupsConcerning iron metabolism we found that serum iron wassignificantly higher for the 200 IU rHuEPO group whencompared with the control and 50 IU rHuEPO groups nosignificant differences were found in ferritin and transferrinserum levels between the three studied groups (Table 2)
Serum samples from all animals were also analyzed foranti-rHuEPO antibodies at 119879
1 1198792 and 119879
3 however only
at 1198793we detected the presence of anti-rHuEPO antibodies
in circulation These were detected in 5 rats under 50 IUrHuEPO treatment as well as in 5 rats treated with 200 IUrHuEPO (625 for both groups) the antibody titer wasfourfold higher in the group under 200 IU rHuEPO the latterpresented a titer of 1 16 and the group under the lowerrHuEPO dose presented a titer of 1 4 Within each rHuEPOgroup no other significant differences were found betweenthe rats with or without anti-rHuEPO antibodies
Bone marrow examination of rats treated with 200 IUrHuEPO showed a proportion of red-cell precursors signif-icantly lower than those without rHuEPO treatment Themyeloid erythroid ratio was 21 1 for the control groupwhile for the 200 IU rHuEPO group the ratio was 68 1 the50 IU rHuEPO group presented a myeloid erythroid ratiosimilar to that found for the control (25 1)
Major changes were observed in the gene expressionof iron regulatory proteins and in erythropoietic regulatoryproteins in liver tissue (Figure 3) particularly in the 200 IUrHuEPO group Indeed the 50 IU rHuEPO group showedsignificant overexpression of HJV and SLC40A1 genes ascompared to control while the 200 IU rHuEPO group pre-sented significant overexpression of Hamp Hfe HJV EPORSLC40A1 Tf TfR2 and BMP6 genes as compared to controland 50 IU rHuEPO group EPO gene was significantly down-regulated in both groups and TMPRSS6 gene expressionwas significantly downregulated only in the 200 IU rHuEPOgroup as compared to the control and 50 IU rHuEPOgroups
The evaluation of the expression of DMT1 and SLC40A1genes in duodenum showed that only the 200 IU rHuEPOgroup presented a significant upregulation for both genes(Figure 4) as compared to the control Furthermore asobserved in liver the expression of EPO in kidney tissue
BioMed Research International 5
Table 2 Hematological and biochemical data at the end of protocol (9 weeks)
Parameters GroupsControl 50 IU rHuEPO 200 IU rHuEPO
EPO (mIUmL) 187 plusmn 025 165 plusmn 023 180 plusmn 021Iron (120583gdL) 18363 plusmn 4158 18863 plusmn 3527 25514 plusmn 11228ab
Ferritin (ngdL) 16513 plusmn 1919 16850 plusmn 4295 16257 plusmn 3981Transferrin (mgmL) 9775 plusmn 868 9938 plusmn 659 9614 plusmn 968Glucose (mgdL) 16088 plusmn 2904 17462 plusmn 2021 18771 plusmn 3743Creatinine (mgdL) 040 plusmn 005 037 plusmn 003 035 plusmn 006Urea (mgdL) 4510 plusmn 192 4505 plusmn 356 4274 plusmn 510Uric acid (mgdL) 094 plusmn 046 070 plusmn 013 103 plusmn 052Bilirubin (ngmL) 0043 plusmn 0016 0054 plusmn 0013 0064 plusmn 0014IL-6 (pgmL) 13810 plusmn 534 13998 plusmn 612 13016 plusmn 506ab
hsCRP (120583gmL) 22931 plusmn 2455 23749 plusmn 2892 24946 plusmn 3349IFN-120574 (pgmL) 2444 plusmn 2307 2145 plusmn 680 2082 plusmn 1013TGF-1205731 (ngmL) 8021 plusmn 499 8003 plusmn 1722 7810 plusmn 555VEGF (pgmL) 536 plusmn 426 584 plusmn 385 604 plusmn 325Results are expressed as mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO IL-6 interleukin-6 hsCRP high-sensitive C-reactive proteinIFN-120574 interferon-120574 TGF-120573 transforming growth factor beta1 VEGF vascular endothelial growth factor
presented a trend towards downregulation in both rHuEPOgroups when compared to the control (Figure 5)
4 Discussion
Antibody-mediated EHPRCA is a rare pathological com-plication associated with the use of recombinant humanerythropoietins and development of anti-EPO antibodiesHowever the use of EPO biosimilars has increased thenumber of cases in the last years [11 20 21] Clinically thatcondition not only abrogates the effect of the rHuEPO butmay also neutralize endogenous EPO leading to severe EHand transfusion-dependent anemia
In this work we developed an animal model of antibody-mediated EH by long-term treatment with a high dose ofrHuEPO (200 IUKg bwweek during 9 weeks) inWistar ratsIn order to confirm that we were in fact in the presence ofan animal model of this hematological disorder we assessedthe parameters typically used as diagnostic criteria forEHPRCA which include normocytic anemia with suddenonset normocellular bone marrow with a selective reduc-tion in red blood cell precursors beyond proerythroblastsreticulocytopenia normal or slightly decreased leucocytecounts and increased serum iron [22] We found that thegroup treated with 200 IUKg bwweek of rHuEPO along 9weeks fits all these criteria and the majority (65) of therats presented detectable anti-rHuEPO antibodies suggest-ing that we achieved an antibody-mediated EH The groupunder 50 IUKg bwweek rHuEPO treatment presented onlyslight changes as compared to control however most ofthe rats (65) presented anti-rHuEPO antibodies at a lowertiter (four times less) suggesting that a longer experimentalprotocol could also lead to the changes associated with EH
Indeed Casadevall (2005) reported that the detection ofanti-erythropoietin antibodies in circulation is not immedi-ately associated with PRCA [23] Moreover no significant
changes were observed in hsCRP IL-6 and IFN-120574 forboth rHuEPO treated groups showing that no inflamma-tory changes could interfere with erythropoiesis and ironmetabolism
Our data suggest that the development of EH was dueto the antibodies directed to rHuEPO that inhibited theerythropoietic stimuli of both rHuEPO and endogenousEPO Indeed in spite of the normal endogenous EPO levelsthat were similar to those of the control group the 200 IUrHuEPO group developed anemia suggesting that the anti-rHuEPO antibodies also neutralized the action of endoge-nous EPO It has been described that the presence of rHuEPOin circulation may increase severalfold the expression ofEPOR [4] Actually we found an overexpression of EPORin the liver of the 200 IU rHuEPO group The decreasedEPO expression observed in kidney and liver tissues forboth rHuEPO groups seems to be in accordance with aphysiological response to the increased circulating levels ofrHuEPO used in the treatment However we should notexclude the hypothesis raised by Piron et al (2001) thatthis anemia could be due to an inappropriate erythropoieticstimulation or to intrinsic failure of erythroid cells to respondto that stimulus [24]
Anemia usually triggers erythropoiesis by increasingEPO production through the hypoxia-inducible factor (HIF)pathway by mobilizing iron from the iron storage pooland by increasing iron absorption in order to face theincreased iron needs for erythropoiesis [25] Consideringthat in antibody-mediated EH the erythropoietic stimulifail due to the inhibition of rHuEPO by anti-rHuEPOantibodies the serum iron should increase as its absorptionandmobilizationwould be triggered by the anemic conditionand hepcidin should be repressed to favour iron absorptionand mobilization Actually we found a significant increasein serum iron in the 200 IU rHuEPO group It is knownthat increasing serum iron induces hepcidin synthesis by
6 BioMed Research International
Hamp
00
05
10
15
20
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SEPO
00
02
04
06
08
10Hfe
00
05
10
15
20
25
HJV
00
05
10
15
20
25EPOR
00
10
20
30SLC40A1
00
10
20
30
TF TfR2
00
20
40
60TMPRSS6
00
05
10
15
20
BMP6
00
10
20
30
lowastlowast
lowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowast
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SRe
lativ
e mRN
A le
vels
18S
Relat
ive m
RNA
leve
ls18
S
00
05
10
15
20
25
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 3 RelativemRNA expression of erythropoietin and iron regulatory proteins in the liver at the end of the protocol (9 weeks) 18S rRNAwas used as reference gene Results are expressed as mean plusmn SD lowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowast119875 lt 0001 versus control group 119875 lt 005119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
a complex pathway Indeed hepcidin encoded by the Hampgene is believed to play a key regulatory role in ironabsorption It controls plasma iron concentration and tissuedistribution of iron by inhibiting intestinal iron absorptioniron recycling by macrophages and iron mobilization from
stores [26] Hepcidin acts by inhibiting cellular iron efflux inhepatocytes enterocytes and macrophages through bindingto ferroportin inducing its degradation [27] Hepatic Hampexpression is regulated by a cohort of proteins includingHfe TfR2HJV BMP6matriptase-2 and transferrin [28ndash30]
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
The current therapeutic use of rHuEPO to correct anemiain CKD patients has been associated with some cases ofPRCA due to the development of cross-reactive anti-EPOantibodies [10 11] Although this complication is very rarethe mechanisms underlying the break in immune toleranceto rHuEPO remain poorly clarified The development ofPRCA should be suspected in patients who have been underrHuEPO therapy for more than 3 months and develop asudden severe unexplained fall in hemoglobin levels andreticulocytopenia and a marked decrease in bone marrowerythroblasts despite continuing with rHuEPO treatment[12] The diagnosis is confirmed by low serum EPO levelsand detection of anti-EPO antibodies [13] The productionof anti-EPO antibodies and the inhibition of EPO-dependenterythroid cells are important mechanisms that might leadto erythroid hypoplasia (EH)PRCA The reduction in func-tional EPO affects the proliferative state of bone marrowerythroid cells which will trigger several changes in ironmetabolism such as in serum iron transferrin and ferritinlevels among others [14ndash16]
Considering the scarce data in literature about howfacing a PRCA condition mediated by anti-EPO antibodiesthe depression of marrow erythroid activity affects ironmetabolism our goal was to assess in male Wistar rats theeffects of this condition on iron metabolism (iron absorptionand iron traffic and storage) as well as on erythropoiesisFor that purpose we performed hematological and biochem-ical studies to evaluate the erythropoietic and iron statusand measured the expression of several mRNA-encodingerythropoiesis and iron metabolism regulating proteins onduodenum liver and kidney tissues
2 Material and Methods
21 Animals and Experimental Protocol Male Wistar rats(Charles River Lab Inc Barcelona Spain) weighing 200ndash250 g weremaintained in an air-conditioned room subjectedto 12 h darklight cycles and given standard laboratory ratdiet (IPM-R20 Letica Barcelona Spain)without iron supple-mentation and free access to tap water Animal experimentswere conducted according to the European CommunitiesCouncil Directives on Animal Care These procedures werein accordance with the Association for Pharmacology andExperimental Therapeutics and were approved by the Insti-tutional Ethics Committee of the Faculty of Medicine fromthe University of Coimbra approval ID FMUC0413 Therats were divided into 3 groups (8 rats each) control treatedwith saline solution 50 IU rHuEPO a therapeutic dose of50 IUKg bodyweight (bw)week of epoetin beta (Recormon-Roche Pharmaceuticals) treatment 200 IU rHuEPO treatedwith 200 IUKg bwweek of rHuEPO a high dose usuallyused on rHuEPO resistant patients All the animals havecompleted the 9-week protocol (Figure 1)
22 Sample Collection and Preparation
221 Blood At the beginning of the experiments (1198790) and
at 3 (1198791) 6 (119879
2) and 9 (119879
3) weeks after the first rHuEpo
3 6 9Time (weeks)
0
administrationAnimals were
sacrificed
Blood sample were collected
Day 1
(50 and 200 IUkgweekthree timesweek sc)
Starting rHuEPO
T0 T1 T2 T3
Figure 1 Experimental protocol
dose the rats were subjected to intraperitoneal anesthesiawith a 2mgkg bwof a 2 1 (v v) 50mgmLketamine (KetalarParke-Davis Lab Pfizer Lda Seixal Portugal) solution in25 chlorpromazine (Largactil Rhone-Poulenc Rorer LabVitoria Amadora Portugal) Blood samples were immedi-ately collected by venipuncture from the jugular vein intovacutainer tubes without anticoagulant (to obtain serum)and with EDTA (to obtain whole blood and plasma) forhematological and biochemical studies 3mL blood sampleswere collected at 119879
0 1198791 and 119879
2to minimize interference
with erythropoiesis mechanism at the end of protocol (1198793)
a 10mL sample was collected in order to perform all thebiochemical and hematological assays
222 Tissues At the end of the protocol blood was col-lected and afterwards the rats were sacrificed by cervicaldislocation duodenum liver and kidneys were immediatelyremoved placed in ice-cold Krebs-Henseleit buffer andcarefully cleaned A bone marrow aspirate from the femurwas also performed
23 Biochemical and Hematological Assays Serum creati-nine and urea concentrations were used as renal functionindexes through automatic validatedmethods and equipment(Hitachi 717 analyser) Red blood cell (RBC) hematocritand hemoglobin (Hb) concentration were assessed in wholeblood EDTA by using an automatic Coulter Counter (Beck-man Coulter Inc CA USA) reticulocyte (RET) countwas performed by microscopic counting on blood smearsafter vital staining with New Methylene Blue (ReticulocyteStain Sigma-Aldrich St Louis MO USA) Serum levelsof erythropoietin were evaluated by rat specific ELISA kit(MyBioSource USA) Serum iron concentration was deter-mined using a colorimetric method (Iron Randox Labo-ratories Ltd UK) whereas serum ferritin and transferrinwere measured by immunoturbidimetry (Laboratories LtdUK) Quantification of total bilirubin was performed by acolorimetric test (diazotized sulfanilic acid reaction RocheDiagnostics) circulating levels of glucose and uric acid weredetermined by routine automated technology (ABX Diag-nostics) Serum levels of interleukin-6 (IL-6) interferon-120574(IFN-120574) transforming growth factor (TGF-1205731) and vascularendothelial growth factor (VEGF) were measured by rat-specific Quantikine ELISA kits from RampD Systems (Min-neapolis USA) High-sensitive C-reactive protein (hsCRP)
BioMed Research International 3
was determined by using a rat-specific ELISA kit from AlphaDiagnostics International (San Antonio USA)
24 Detection of Anti-EPO Antibodies The detection of anti-EPO antibodies was carried out by ELISA according to Urraet al 1997 using rHuEPO (Recormon Roche Pharmaceuti-cals) as antigen and as secondary antibody goat anti-rat IgGconjugated with horseradish peroxidase (Sigma 100 ngmLfor 1 h at room temperature) [17] The substrate tetramethyl-benzidine (TMB) (Sigma) was added and the reaction wasstopped by the addition of sulphuric acid 125molL Theoptical density at 450 nm (OD450) was determined with anautomatic plate reader
25 Gene Expression Analysis In order to isolate total RNA02 g of liver duodenum and kidney samples from each ratwas immersed in RNA later (Ambion Austin USA) uponcollection and stored at 4∘C for 24 h afterwards sampleswere frozen at minus80∘C Subsequently tissue samples weighing50 plusmn 10mg were homogenized in a total volume of 1mLTRI Reagent using a homogenizer and total RNA was iso-lated according to manufacturer instructions (Sigma SintraPortugal) To ensure inactivation of contaminating RNAsesmetal objects and glassware were cleaned with detergentimmersed in RNAse-free water (02 diethyl pyrocarbonate)for 2 h and finally heated at 120∘C for 1 h RNA integrity (RINRNA Integrity Number) was analyzed using 6000 Nano kitinAgilent 2100 bioanalyzer (Agilent TechnologiesWalbronnGermany) and 2100 expert software followingmanufacturerinstructions The yield from isolation was from 05 to 15 120583gRIN values were 78ndash90 and purity (A260A280) was 18ndash20The concentrations of the RNA preparations were confirmedwith NanoDrop1000 (ThermoScientific Wilmington DEUSA) Possible contaminating remnants of genomic DNAwere eliminated by treating these preparations with deoxyri-bonuclease I (amplification grade) prior to RT-qPCR ampli-fication Reverse transcription and relative quantification ofgene expression were performed as previously described [18]Real-time qPCR reactions were performed for the followinggenes EPO EPOR transferrin receptor 2 (TfR2) hepcidin(Hamp) ferroportin (SLC40A1) Hemojuvelin (HJV) trans-ferrin (TF) hemochromatosis (Hfe) divalent metal trans-porter 1 (DMT1) transferrin receptor 1 (TfR1) matriptase-2 (TMPRSS6) and bone morphogenic protein 6 (BMP6)which were normalized in relation to the expression of beta-actin (Actb) and 18S ribosomal subunit (18S) as well as tothe mean of the control group Primer sequences are listed inTable 1 Results were analyzed with SDS 21 software (AppliedBiosystems Foster City CA USA) and relative quantificationwas calculated using the 2minusΔΔCt method [19] In liver tissue westudied the EPO EPORTfR2Hamp SLC40A1HJVTFHfeBMP6 and TMPRSS6 gene expression in duodenum tissuethe gene expressions ofDMT1 and SLC40A1were studied andin the kidney we evaluated the expression of EPO gene
26 Data Analysis For statistical analysis we used theStatistical Package for Social Sciences (SPSS) version 220Results are presented as mean plusmn standard deviation Multiple
Table 1 List of primer sequences (F forward R reverse)
Gene Primer sequences
EPO F 51015840-AGGGTCACGAAGCCATGAAG-31015840
R 51015840-GAT TTC GGC TGT TGC CAG TG-31015840
EPOR F 51015840-GCG ACT TGG ACC CTC TCA TC-31015840
R 51015840-AGT TAC CCT TGT GGG TGG TG-31015840
Hamp F 51015840-GAA GGC AAG ATG GCA CTA AGC-31015840
R 51015840-CAG AGC CGT AGT CTG TCT CG-31015840
TfR2 F 51015840-CAA GCT TCG CCC AGA AGG TA-31015840
R 51015840-CGT GTA AGG GTC CCC AGT TC-31015840
SLC40A1 F 51015840-CAG GCT TAG GGT CTA CTG CG-31015840
R 51015840-CCG AAA GAC CCC AAA GGA CA-31015840
HJV F 51015840-GCC TAC TTC CAA TCC TGC GT-31015840
R 51015840-GGT CAA GAA GAC TCG GGC AT-31015840
TF F 51015840-GGC ATC AGA CTC CAG CAT CA-31015840
R 51015840-GCA GGC CCA TAG GGA TGT T-31015840
Hfe F 51015840-CTG GAT CAG CCT CTC ACT GC-31015840
R 51015840-GTC ACC CAT GGT TCC TCC TG-31015840
DMT1 F 51015840-CAA CTC TAC CCT GGC TGT GG-31015840
R 51015840-GTC ATG GTG GAG CTC TGT CC-31015840
TfR1 F 51015840-GCT CGT GGA GAC TAC TTC CG-31015840
R 51015840-GCC CCA GAA GAT GTG TCG G-31015840
TMPRSS6 F 51015840-CCG AAT ATG AGG TGG ACC CG-31015840
R 51015840-GGT TCA CGT AGC TGT AGC GG-31015840
BMP6 F 51015840-GCT GCC AAC TAT TGT GAC GG-31015840
R 51015840-GGT TTG GGG ACG TAC TCG G-31015840
18S F 51015840-CCA CTA AAG GGC ATC CTG GG-31015840
R 51015840-CAT TGA GAG CAA TGC CAG CC-31015840
Actb F 51015840-GAG ATT ACT GCC CTG GCT CC-31015840
R 51015840-CGG ACT CAT CGT ACT CCT GC-31015840
comparisons between groups were performed by one-wayANOVA supplemented with Tukeyrsquos HSD post hoc testFor single comparisons we used the Mann-Whitney 119880 testSignificance was accepted for a 119875minor than 005
3 Results
The changes in RBC count Hb concentration hematocritand the number of reticulocytes for the three groups arepresented in Figure 2 During the 9-week experimentalprotocol the 50 IU rHuEPO group when compared tothe control group showed similar values for RBC countHb concentration hematocrit and reticulocytes though asignificant increase in reticulocyte count was observed at 119879
2
and 1198793 Concerning the rats treated with 200 IU rHuEPO in
the first 3 weeks we found a significant increase (gt30 thebasal value) in hemoglobin levels RBC count hematocritand reticulocyte count as compared to control and 50 IUrHuEPO groups after this period the hemoglobin concen-tration hematocrit and RBC count as well as the number ofreticulocytes decreased reaching significantly lower values at1198792 as compared to 119879
1 this trend towards decreasing values
was maintained till the end of the experiment (9 weeks)
4 BioMed Research International
Control
3 6 90
200
400
600
800
Time (weeks)
3 6 95
6
7
8
9
10
11
3 6 920
30
40
50
60
Hem
atoc
rit (
)
10
12
14
16
18
20
Hem
oglo
bin
(gd
L)
0
RBC
(1012L
)
0Time (weeks) Time (weeks)
3 6 90 0Time (weeks)
Retic
uloc
ytes
(109L
)
50 IU + rHuEPO200 IU + rHuEPO Control
50 IU + rHuEPO200 IU + rHuEPO
yensect
120575cd
120575
lowast
sect
120575cd
lowast
sect
yensect
120575cd 120575cd
120575
120575
lowast
sect yen
a a
b
Figure 2 Erythrocyte data and reticulocyte count during the follow-up period of 9 weeks under rHuEPO treatment Results are expressedas mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO c119875 lt 0001 versus control d119875 lt 0001 versus 50 IU rHuEPO120575119875 lt 0005 1198790 versus 1198791
lowast119875 lt 005 1198791 versus 1198792
sect119875 lt 005 1198792 versus 1198793
At the end of the protocol serum erythropoietin levelswere similar among the three groups (Table 2) No changeswere observed in glucose creatinine urea uric acid andbilirubin levels for both rHuEPO groups along the entireprotocol when compared to the control The inflammatorymarker IL-6 presented a significant decrease in the 200 IUrHuEPO group when compared with the control and 50 IUrHuEPO groups hsCRP IFN-120574 TGF-1205731 and VEGF showedno significant differences between the three studied groupsConcerning iron metabolism we found that serum iron wassignificantly higher for the 200 IU rHuEPO group whencompared with the control and 50 IU rHuEPO groups nosignificant differences were found in ferritin and transferrinserum levels between the three studied groups (Table 2)
Serum samples from all animals were also analyzed foranti-rHuEPO antibodies at 119879
1 1198792 and 119879
3 however only
at 1198793we detected the presence of anti-rHuEPO antibodies
in circulation These were detected in 5 rats under 50 IUrHuEPO treatment as well as in 5 rats treated with 200 IUrHuEPO (625 for both groups) the antibody titer wasfourfold higher in the group under 200 IU rHuEPO the latterpresented a titer of 1 16 and the group under the lowerrHuEPO dose presented a titer of 1 4 Within each rHuEPOgroup no other significant differences were found betweenthe rats with or without anti-rHuEPO antibodies
Bone marrow examination of rats treated with 200 IUrHuEPO showed a proportion of red-cell precursors signif-icantly lower than those without rHuEPO treatment Themyeloid erythroid ratio was 21 1 for the control groupwhile for the 200 IU rHuEPO group the ratio was 68 1 the50 IU rHuEPO group presented a myeloid erythroid ratiosimilar to that found for the control (25 1)
Major changes were observed in the gene expressionof iron regulatory proteins and in erythropoietic regulatoryproteins in liver tissue (Figure 3) particularly in the 200 IUrHuEPO group Indeed the 50 IU rHuEPO group showedsignificant overexpression of HJV and SLC40A1 genes ascompared to control while the 200 IU rHuEPO group pre-sented significant overexpression of Hamp Hfe HJV EPORSLC40A1 Tf TfR2 and BMP6 genes as compared to controland 50 IU rHuEPO group EPO gene was significantly down-regulated in both groups and TMPRSS6 gene expressionwas significantly downregulated only in the 200 IU rHuEPOgroup as compared to the control and 50 IU rHuEPOgroups
The evaluation of the expression of DMT1 and SLC40A1genes in duodenum showed that only the 200 IU rHuEPOgroup presented a significant upregulation for both genes(Figure 4) as compared to the control Furthermore asobserved in liver the expression of EPO in kidney tissue
BioMed Research International 5
Table 2 Hematological and biochemical data at the end of protocol (9 weeks)
Parameters GroupsControl 50 IU rHuEPO 200 IU rHuEPO
EPO (mIUmL) 187 plusmn 025 165 plusmn 023 180 plusmn 021Iron (120583gdL) 18363 plusmn 4158 18863 plusmn 3527 25514 plusmn 11228ab
Ferritin (ngdL) 16513 plusmn 1919 16850 plusmn 4295 16257 plusmn 3981Transferrin (mgmL) 9775 plusmn 868 9938 plusmn 659 9614 plusmn 968Glucose (mgdL) 16088 plusmn 2904 17462 plusmn 2021 18771 plusmn 3743Creatinine (mgdL) 040 plusmn 005 037 plusmn 003 035 plusmn 006Urea (mgdL) 4510 plusmn 192 4505 plusmn 356 4274 plusmn 510Uric acid (mgdL) 094 plusmn 046 070 plusmn 013 103 plusmn 052Bilirubin (ngmL) 0043 plusmn 0016 0054 plusmn 0013 0064 plusmn 0014IL-6 (pgmL) 13810 plusmn 534 13998 plusmn 612 13016 plusmn 506ab
hsCRP (120583gmL) 22931 plusmn 2455 23749 plusmn 2892 24946 plusmn 3349IFN-120574 (pgmL) 2444 plusmn 2307 2145 plusmn 680 2082 plusmn 1013TGF-1205731 (ngmL) 8021 plusmn 499 8003 plusmn 1722 7810 plusmn 555VEGF (pgmL) 536 plusmn 426 584 plusmn 385 604 plusmn 325Results are expressed as mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO IL-6 interleukin-6 hsCRP high-sensitive C-reactive proteinIFN-120574 interferon-120574 TGF-120573 transforming growth factor beta1 VEGF vascular endothelial growth factor
presented a trend towards downregulation in both rHuEPOgroups when compared to the control (Figure 5)
4 Discussion
Antibody-mediated EHPRCA is a rare pathological com-plication associated with the use of recombinant humanerythropoietins and development of anti-EPO antibodiesHowever the use of EPO biosimilars has increased thenumber of cases in the last years [11 20 21] Clinically thatcondition not only abrogates the effect of the rHuEPO butmay also neutralize endogenous EPO leading to severe EHand transfusion-dependent anemia
In this work we developed an animal model of antibody-mediated EH by long-term treatment with a high dose ofrHuEPO (200 IUKg bwweek during 9 weeks) inWistar ratsIn order to confirm that we were in fact in the presence ofan animal model of this hematological disorder we assessedthe parameters typically used as diagnostic criteria forEHPRCA which include normocytic anemia with suddenonset normocellular bone marrow with a selective reduc-tion in red blood cell precursors beyond proerythroblastsreticulocytopenia normal or slightly decreased leucocytecounts and increased serum iron [22] We found that thegroup treated with 200 IUKg bwweek of rHuEPO along 9weeks fits all these criteria and the majority (65) of therats presented detectable anti-rHuEPO antibodies suggest-ing that we achieved an antibody-mediated EH The groupunder 50 IUKg bwweek rHuEPO treatment presented onlyslight changes as compared to control however most ofthe rats (65) presented anti-rHuEPO antibodies at a lowertiter (four times less) suggesting that a longer experimentalprotocol could also lead to the changes associated with EH
Indeed Casadevall (2005) reported that the detection ofanti-erythropoietin antibodies in circulation is not immedi-ately associated with PRCA [23] Moreover no significant
changes were observed in hsCRP IL-6 and IFN-120574 forboth rHuEPO treated groups showing that no inflamma-tory changes could interfere with erythropoiesis and ironmetabolism
Our data suggest that the development of EH was dueto the antibodies directed to rHuEPO that inhibited theerythropoietic stimuli of both rHuEPO and endogenousEPO Indeed in spite of the normal endogenous EPO levelsthat were similar to those of the control group the 200 IUrHuEPO group developed anemia suggesting that the anti-rHuEPO antibodies also neutralized the action of endoge-nous EPO It has been described that the presence of rHuEPOin circulation may increase severalfold the expression ofEPOR [4] Actually we found an overexpression of EPORin the liver of the 200 IU rHuEPO group The decreasedEPO expression observed in kidney and liver tissues forboth rHuEPO groups seems to be in accordance with aphysiological response to the increased circulating levels ofrHuEPO used in the treatment However we should notexclude the hypothesis raised by Piron et al (2001) thatthis anemia could be due to an inappropriate erythropoieticstimulation or to intrinsic failure of erythroid cells to respondto that stimulus [24]
Anemia usually triggers erythropoiesis by increasingEPO production through the hypoxia-inducible factor (HIF)pathway by mobilizing iron from the iron storage pooland by increasing iron absorption in order to face theincreased iron needs for erythropoiesis [25] Consideringthat in antibody-mediated EH the erythropoietic stimulifail due to the inhibition of rHuEPO by anti-rHuEPOantibodies the serum iron should increase as its absorptionandmobilizationwould be triggered by the anemic conditionand hepcidin should be repressed to favour iron absorptionand mobilization Actually we found a significant increasein serum iron in the 200 IU rHuEPO group It is knownthat increasing serum iron induces hepcidin synthesis by
6 BioMed Research International
Hamp
00
05
10
15
20
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SEPO
00
02
04
06
08
10Hfe
00
05
10
15
20
25
HJV
00
05
10
15
20
25EPOR
00
10
20
30SLC40A1
00
10
20
30
TF TfR2
00
20
40
60TMPRSS6
00
05
10
15
20
BMP6
00
10
20
30
lowastlowast
lowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowast
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SRe
lativ
e mRN
A le
vels
18S
Relat
ive m
RNA
leve
ls18
S
00
05
10
15
20
25
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 3 RelativemRNA expression of erythropoietin and iron regulatory proteins in the liver at the end of the protocol (9 weeks) 18S rRNAwas used as reference gene Results are expressed as mean plusmn SD lowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowast119875 lt 0001 versus control group 119875 lt 005119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
a complex pathway Indeed hepcidin encoded by the Hampgene is believed to play a key regulatory role in ironabsorption It controls plasma iron concentration and tissuedistribution of iron by inhibiting intestinal iron absorptioniron recycling by macrophages and iron mobilization from
stores [26] Hepcidin acts by inhibiting cellular iron efflux inhepatocytes enterocytes and macrophages through bindingto ferroportin inducing its degradation [27] Hepatic Hampexpression is regulated by a cohort of proteins includingHfe TfR2HJV BMP6matriptase-2 and transferrin [28ndash30]
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 3
was determined by using a rat-specific ELISA kit from AlphaDiagnostics International (San Antonio USA)
24 Detection of Anti-EPO Antibodies The detection of anti-EPO antibodies was carried out by ELISA according to Urraet al 1997 using rHuEPO (Recormon Roche Pharmaceuti-cals) as antigen and as secondary antibody goat anti-rat IgGconjugated with horseradish peroxidase (Sigma 100 ngmLfor 1 h at room temperature) [17] The substrate tetramethyl-benzidine (TMB) (Sigma) was added and the reaction wasstopped by the addition of sulphuric acid 125molL Theoptical density at 450 nm (OD450) was determined with anautomatic plate reader
25 Gene Expression Analysis In order to isolate total RNA02 g of liver duodenum and kidney samples from each ratwas immersed in RNA later (Ambion Austin USA) uponcollection and stored at 4∘C for 24 h afterwards sampleswere frozen at minus80∘C Subsequently tissue samples weighing50 plusmn 10mg were homogenized in a total volume of 1mLTRI Reagent using a homogenizer and total RNA was iso-lated according to manufacturer instructions (Sigma SintraPortugal) To ensure inactivation of contaminating RNAsesmetal objects and glassware were cleaned with detergentimmersed in RNAse-free water (02 diethyl pyrocarbonate)for 2 h and finally heated at 120∘C for 1 h RNA integrity (RINRNA Integrity Number) was analyzed using 6000 Nano kitinAgilent 2100 bioanalyzer (Agilent TechnologiesWalbronnGermany) and 2100 expert software followingmanufacturerinstructions The yield from isolation was from 05 to 15 120583gRIN values were 78ndash90 and purity (A260A280) was 18ndash20The concentrations of the RNA preparations were confirmedwith NanoDrop1000 (ThermoScientific Wilmington DEUSA) Possible contaminating remnants of genomic DNAwere eliminated by treating these preparations with deoxyri-bonuclease I (amplification grade) prior to RT-qPCR ampli-fication Reverse transcription and relative quantification ofgene expression were performed as previously described [18]Real-time qPCR reactions were performed for the followinggenes EPO EPOR transferrin receptor 2 (TfR2) hepcidin(Hamp) ferroportin (SLC40A1) Hemojuvelin (HJV) trans-ferrin (TF) hemochromatosis (Hfe) divalent metal trans-porter 1 (DMT1) transferrin receptor 1 (TfR1) matriptase-2 (TMPRSS6) and bone morphogenic protein 6 (BMP6)which were normalized in relation to the expression of beta-actin (Actb) and 18S ribosomal subunit (18S) as well as tothe mean of the control group Primer sequences are listed inTable 1 Results were analyzed with SDS 21 software (AppliedBiosystems Foster City CA USA) and relative quantificationwas calculated using the 2minusΔΔCt method [19] In liver tissue westudied the EPO EPORTfR2Hamp SLC40A1HJVTFHfeBMP6 and TMPRSS6 gene expression in duodenum tissuethe gene expressions ofDMT1 and SLC40A1were studied andin the kidney we evaluated the expression of EPO gene
26 Data Analysis For statistical analysis we used theStatistical Package for Social Sciences (SPSS) version 220Results are presented as mean plusmn standard deviation Multiple
Table 1 List of primer sequences (F forward R reverse)
Gene Primer sequences
EPO F 51015840-AGGGTCACGAAGCCATGAAG-31015840
R 51015840-GAT TTC GGC TGT TGC CAG TG-31015840
EPOR F 51015840-GCG ACT TGG ACC CTC TCA TC-31015840
R 51015840-AGT TAC CCT TGT GGG TGG TG-31015840
Hamp F 51015840-GAA GGC AAG ATG GCA CTA AGC-31015840
R 51015840-CAG AGC CGT AGT CTG TCT CG-31015840
TfR2 F 51015840-CAA GCT TCG CCC AGA AGG TA-31015840
R 51015840-CGT GTA AGG GTC CCC AGT TC-31015840
SLC40A1 F 51015840-CAG GCT TAG GGT CTA CTG CG-31015840
R 51015840-CCG AAA GAC CCC AAA GGA CA-31015840
HJV F 51015840-GCC TAC TTC CAA TCC TGC GT-31015840
R 51015840-GGT CAA GAA GAC TCG GGC AT-31015840
TF F 51015840-GGC ATC AGA CTC CAG CAT CA-31015840
R 51015840-GCA GGC CCA TAG GGA TGT T-31015840
Hfe F 51015840-CTG GAT CAG CCT CTC ACT GC-31015840
R 51015840-GTC ACC CAT GGT TCC TCC TG-31015840
DMT1 F 51015840-CAA CTC TAC CCT GGC TGT GG-31015840
R 51015840-GTC ATG GTG GAG CTC TGT CC-31015840
TfR1 F 51015840-GCT CGT GGA GAC TAC TTC CG-31015840
R 51015840-GCC CCA GAA GAT GTG TCG G-31015840
TMPRSS6 F 51015840-CCG AAT ATG AGG TGG ACC CG-31015840
R 51015840-GGT TCA CGT AGC TGT AGC GG-31015840
BMP6 F 51015840-GCT GCC AAC TAT TGT GAC GG-31015840
R 51015840-GGT TTG GGG ACG TAC TCG G-31015840
18S F 51015840-CCA CTA AAG GGC ATC CTG GG-31015840
R 51015840-CAT TGA GAG CAA TGC CAG CC-31015840
Actb F 51015840-GAG ATT ACT GCC CTG GCT CC-31015840
R 51015840-CGG ACT CAT CGT ACT CCT GC-31015840
comparisons between groups were performed by one-wayANOVA supplemented with Tukeyrsquos HSD post hoc testFor single comparisons we used the Mann-Whitney 119880 testSignificance was accepted for a 119875minor than 005
3 Results
The changes in RBC count Hb concentration hematocritand the number of reticulocytes for the three groups arepresented in Figure 2 During the 9-week experimentalprotocol the 50 IU rHuEPO group when compared tothe control group showed similar values for RBC countHb concentration hematocrit and reticulocytes though asignificant increase in reticulocyte count was observed at 119879
2
and 1198793 Concerning the rats treated with 200 IU rHuEPO in
the first 3 weeks we found a significant increase (gt30 thebasal value) in hemoglobin levels RBC count hematocritand reticulocyte count as compared to control and 50 IUrHuEPO groups after this period the hemoglobin concen-tration hematocrit and RBC count as well as the number ofreticulocytes decreased reaching significantly lower values at1198792 as compared to 119879
1 this trend towards decreasing values
was maintained till the end of the experiment (9 weeks)
4 BioMed Research International
Control
3 6 90
200
400
600
800
Time (weeks)
3 6 95
6
7
8
9
10
11
3 6 920
30
40
50
60
Hem
atoc
rit (
)
10
12
14
16
18
20
Hem
oglo
bin
(gd
L)
0
RBC
(1012L
)
0Time (weeks) Time (weeks)
3 6 90 0Time (weeks)
Retic
uloc
ytes
(109L
)
50 IU + rHuEPO200 IU + rHuEPO Control
50 IU + rHuEPO200 IU + rHuEPO
yensect
120575cd
120575
lowast
sect
120575cd
lowast
sect
yensect
120575cd 120575cd
120575
120575
lowast
sect yen
a a
b
Figure 2 Erythrocyte data and reticulocyte count during the follow-up period of 9 weeks under rHuEPO treatment Results are expressedas mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO c119875 lt 0001 versus control d119875 lt 0001 versus 50 IU rHuEPO120575119875 lt 0005 1198790 versus 1198791
lowast119875 lt 005 1198791 versus 1198792
sect119875 lt 005 1198792 versus 1198793
At the end of the protocol serum erythropoietin levelswere similar among the three groups (Table 2) No changeswere observed in glucose creatinine urea uric acid andbilirubin levels for both rHuEPO groups along the entireprotocol when compared to the control The inflammatorymarker IL-6 presented a significant decrease in the 200 IUrHuEPO group when compared with the control and 50 IUrHuEPO groups hsCRP IFN-120574 TGF-1205731 and VEGF showedno significant differences between the three studied groupsConcerning iron metabolism we found that serum iron wassignificantly higher for the 200 IU rHuEPO group whencompared with the control and 50 IU rHuEPO groups nosignificant differences were found in ferritin and transferrinserum levels between the three studied groups (Table 2)
Serum samples from all animals were also analyzed foranti-rHuEPO antibodies at 119879
1 1198792 and 119879
3 however only
at 1198793we detected the presence of anti-rHuEPO antibodies
in circulation These were detected in 5 rats under 50 IUrHuEPO treatment as well as in 5 rats treated with 200 IUrHuEPO (625 for both groups) the antibody titer wasfourfold higher in the group under 200 IU rHuEPO the latterpresented a titer of 1 16 and the group under the lowerrHuEPO dose presented a titer of 1 4 Within each rHuEPOgroup no other significant differences were found betweenthe rats with or without anti-rHuEPO antibodies
Bone marrow examination of rats treated with 200 IUrHuEPO showed a proportion of red-cell precursors signif-icantly lower than those without rHuEPO treatment Themyeloid erythroid ratio was 21 1 for the control groupwhile for the 200 IU rHuEPO group the ratio was 68 1 the50 IU rHuEPO group presented a myeloid erythroid ratiosimilar to that found for the control (25 1)
Major changes were observed in the gene expressionof iron regulatory proteins and in erythropoietic regulatoryproteins in liver tissue (Figure 3) particularly in the 200 IUrHuEPO group Indeed the 50 IU rHuEPO group showedsignificant overexpression of HJV and SLC40A1 genes ascompared to control while the 200 IU rHuEPO group pre-sented significant overexpression of Hamp Hfe HJV EPORSLC40A1 Tf TfR2 and BMP6 genes as compared to controland 50 IU rHuEPO group EPO gene was significantly down-regulated in both groups and TMPRSS6 gene expressionwas significantly downregulated only in the 200 IU rHuEPOgroup as compared to the control and 50 IU rHuEPOgroups
The evaluation of the expression of DMT1 and SLC40A1genes in duodenum showed that only the 200 IU rHuEPOgroup presented a significant upregulation for both genes(Figure 4) as compared to the control Furthermore asobserved in liver the expression of EPO in kidney tissue
BioMed Research International 5
Table 2 Hematological and biochemical data at the end of protocol (9 weeks)
Parameters GroupsControl 50 IU rHuEPO 200 IU rHuEPO
EPO (mIUmL) 187 plusmn 025 165 plusmn 023 180 plusmn 021Iron (120583gdL) 18363 plusmn 4158 18863 plusmn 3527 25514 plusmn 11228ab
Ferritin (ngdL) 16513 plusmn 1919 16850 plusmn 4295 16257 plusmn 3981Transferrin (mgmL) 9775 plusmn 868 9938 plusmn 659 9614 plusmn 968Glucose (mgdL) 16088 plusmn 2904 17462 plusmn 2021 18771 plusmn 3743Creatinine (mgdL) 040 plusmn 005 037 plusmn 003 035 plusmn 006Urea (mgdL) 4510 plusmn 192 4505 plusmn 356 4274 plusmn 510Uric acid (mgdL) 094 plusmn 046 070 plusmn 013 103 plusmn 052Bilirubin (ngmL) 0043 plusmn 0016 0054 plusmn 0013 0064 plusmn 0014IL-6 (pgmL) 13810 plusmn 534 13998 plusmn 612 13016 plusmn 506ab
hsCRP (120583gmL) 22931 plusmn 2455 23749 plusmn 2892 24946 plusmn 3349IFN-120574 (pgmL) 2444 plusmn 2307 2145 plusmn 680 2082 plusmn 1013TGF-1205731 (ngmL) 8021 plusmn 499 8003 plusmn 1722 7810 plusmn 555VEGF (pgmL) 536 plusmn 426 584 plusmn 385 604 plusmn 325Results are expressed as mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO IL-6 interleukin-6 hsCRP high-sensitive C-reactive proteinIFN-120574 interferon-120574 TGF-120573 transforming growth factor beta1 VEGF vascular endothelial growth factor
presented a trend towards downregulation in both rHuEPOgroups when compared to the control (Figure 5)
4 Discussion
Antibody-mediated EHPRCA is a rare pathological com-plication associated with the use of recombinant humanerythropoietins and development of anti-EPO antibodiesHowever the use of EPO biosimilars has increased thenumber of cases in the last years [11 20 21] Clinically thatcondition not only abrogates the effect of the rHuEPO butmay also neutralize endogenous EPO leading to severe EHand transfusion-dependent anemia
In this work we developed an animal model of antibody-mediated EH by long-term treatment with a high dose ofrHuEPO (200 IUKg bwweek during 9 weeks) inWistar ratsIn order to confirm that we were in fact in the presence ofan animal model of this hematological disorder we assessedthe parameters typically used as diagnostic criteria forEHPRCA which include normocytic anemia with suddenonset normocellular bone marrow with a selective reduc-tion in red blood cell precursors beyond proerythroblastsreticulocytopenia normal or slightly decreased leucocytecounts and increased serum iron [22] We found that thegroup treated with 200 IUKg bwweek of rHuEPO along 9weeks fits all these criteria and the majority (65) of therats presented detectable anti-rHuEPO antibodies suggest-ing that we achieved an antibody-mediated EH The groupunder 50 IUKg bwweek rHuEPO treatment presented onlyslight changes as compared to control however most ofthe rats (65) presented anti-rHuEPO antibodies at a lowertiter (four times less) suggesting that a longer experimentalprotocol could also lead to the changes associated with EH
Indeed Casadevall (2005) reported that the detection ofanti-erythropoietin antibodies in circulation is not immedi-ately associated with PRCA [23] Moreover no significant
changes were observed in hsCRP IL-6 and IFN-120574 forboth rHuEPO treated groups showing that no inflamma-tory changes could interfere with erythropoiesis and ironmetabolism
Our data suggest that the development of EH was dueto the antibodies directed to rHuEPO that inhibited theerythropoietic stimuli of both rHuEPO and endogenousEPO Indeed in spite of the normal endogenous EPO levelsthat were similar to those of the control group the 200 IUrHuEPO group developed anemia suggesting that the anti-rHuEPO antibodies also neutralized the action of endoge-nous EPO It has been described that the presence of rHuEPOin circulation may increase severalfold the expression ofEPOR [4] Actually we found an overexpression of EPORin the liver of the 200 IU rHuEPO group The decreasedEPO expression observed in kidney and liver tissues forboth rHuEPO groups seems to be in accordance with aphysiological response to the increased circulating levels ofrHuEPO used in the treatment However we should notexclude the hypothesis raised by Piron et al (2001) thatthis anemia could be due to an inappropriate erythropoieticstimulation or to intrinsic failure of erythroid cells to respondto that stimulus [24]
Anemia usually triggers erythropoiesis by increasingEPO production through the hypoxia-inducible factor (HIF)pathway by mobilizing iron from the iron storage pooland by increasing iron absorption in order to face theincreased iron needs for erythropoiesis [25] Consideringthat in antibody-mediated EH the erythropoietic stimulifail due to the inhibition of rHuEPO by anti-rHuEPOantibodies the serum iron should increase as its absorptionandmobilizationwould be triggered by the anemic conditionand hepcidin should be repressed to favour iron absorptionand mobilization Actually we found a significant increasein serum iron in the 200 IU rHuEPO group It is knownthat increasing serum iron induces hepcidin synthesis by
6 BioMed Research International
Hamp
00
05
10
15
20
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SEPO
00
02
04
06
08
10Hfe
00
05
10
15
20
25
HJV
00
05
10
15
20
25EPOR
00
10
20
30SLC40A1
00
10
20
30
TF TfR2
00
20
40
60TMPRSS6
00
05
10
15
20
BMP6
00
10
20
30
lowastlowast
lowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowast
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SRe
lativ
e mRN
A le
vels
18S
Relat
ive m
RNA
leve
ls18
S
00
05
10
15
20
25
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 3 RelativemRNA expression of erythropoietin and iron regulatory proteins in the liver at the end of the protocol (9 weeks) 18S rRNAwas used as reference gene Results are expressed as mean plusmn SD lowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowast119875 lt 0001 versus control group 119875 lt 005119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
a complex pathway Indeed hepcidin encoded by the Hampgene is believed to play a key regulatory role in ironabsorption It controls plasma iron concentration and tissuedistribution of iron by inhibiting intestinal iron absorptioniron recycling by macrophages and iron mobilization from
stores [26] Hepcidin acts by inhibiting cellular iron efflux inhepatocytes enterocytes and macrophages through bindingto ferroportin inducing its degradation [27] Hepatic Hampexpression is regulated by a cohort of proteins includingHfe TfR2HJV BMP6matriptase-2 and transferrin [28ndash30]
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
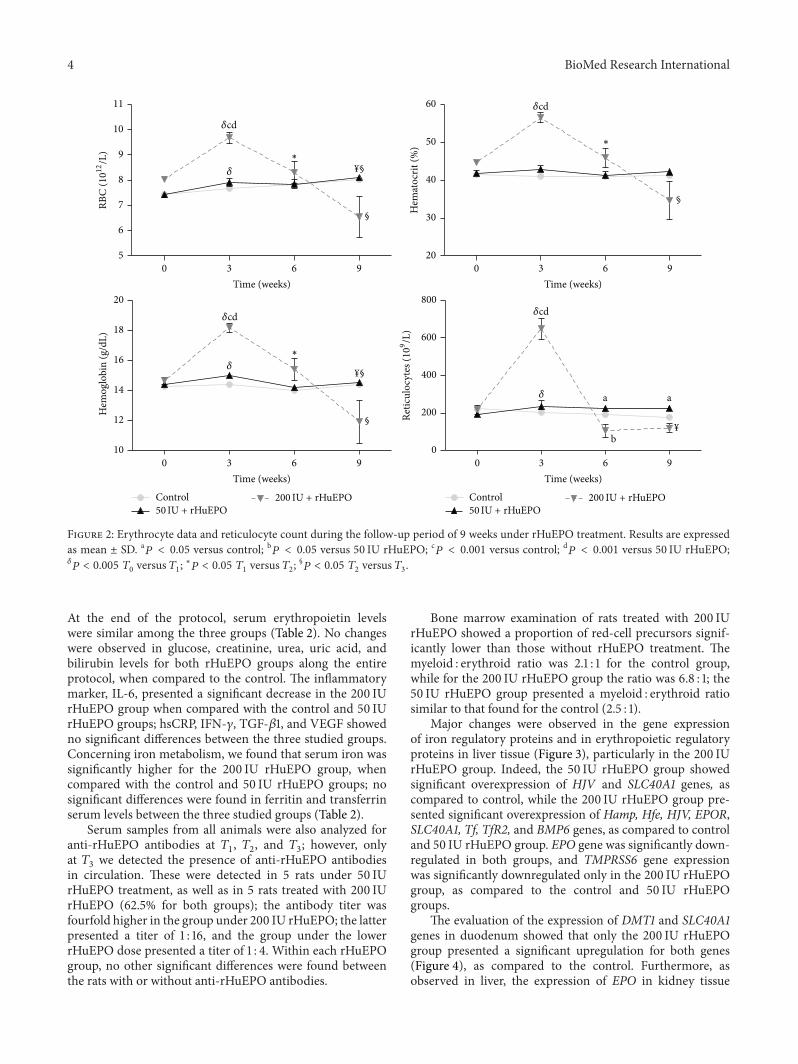
4 BioMed Research International
Control
3 6 90
200
400
600
800
Time (weeks)
3 6 95
6
7
8
9
10
11
3 6 920
30
40
50
60
Hem
atoc
rit (
)
10
12
14
16
18
20
Hem
oglo
bin
(gd
L)
0
RBC
(1012L
)
0Time (weeks) Time (weeks)
3 6 90 0Time (weeks)
Retic
uloc
ytes
(109L
)
50 IU + rHuEPO200 IU + rHuEPO Control
50 IU + rHuEPO200 IU + rHuEPO
yensect
120575cd
120575
lowast
sect
120575cd
lowast
sect
yensect
120575cd 120575cd
120575
120575
lowast
sect yen
a a
b
Figure 2 Erythrocyte data and reticulocyte count during the follow-up period of 9 weeks under rHuEPO treatment Results are expressedas mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO c119875 lt 0001 versus control d119875 lt 0001 versus 50 IU rHuEPO120575119875 lt 0005 1198790 versus 1198791
lowast119875 lt 005 1198791 versus 1198792
sect119875 lt 005 1198792 versus 1198793
At the end of the protocol serum erythropoietin levelswere similar among the three groups (Table 2) No changeswere observed in glucose creatinine urea uric acid andbilirubin levels for both rHuEPO groups along the entireprotocol when compared to the control The inflammatorymarker IL-6 presented a significant decrease in the 200 IUrHuEPO group when compared with the control and 50 IUrHuEPO groups hsCRP IFN-120574 TGF-1205731 and VEGF showedno significant differences between the three studied groupsConcerning iron metabolism we found that serum iron wassignificantly higher for the 200 IU rHuEPO group whencompared with the control and 50 IU rHuEPO groups nosignificant differences were found in ferritin and transferrinserum levels between the three studied groups (Table 2)
Serum samples from all animals were also analyzed foranti-rHuEPO antibodies at 119879
1 1198792 and 119879
3 however only
at 1198793we detected the presence of anti-rHuEPO antibodies
in circulation These were detected in 5 rats under 50 IUrHuEPO treatment as well as in 5 rats treated with 200 IUrHuEPO (625 for both groups) the antibody titer wasfourfold higher in the group under 200 IU rHuEPO the latterpresented a titer of 1 16 and the group under the lowerrHuEPO dose presented a titer of 1 4 Within each rHuEPOgroup no other significant differences were found betweenthe rats with or without anti-rHuEPO antibodies
Bone marrow examination of rats treated with 200 IUrHuEPO showed a proportion of red-cell precursors signif-icantly lower than those without rHuEPO treatment Themyeloid erythroid ratio was 21 1 for the control groupwhile for the 200 IU rHuEPO group the ratio was 68 1 the50 IU rHuEPO group presented a myeloid erythroid ratiosimilar to that found for the control (25 1)
Major changes were observed in the gene expressionof iron regulatory proteins and in erythropoietic regulatoryproteins in liver tissue (Figure 3) particularly in the 200 IUrHuEPO group Indeed the 50 IU rHuEPO group showedsignificant overexpression of HJV and SLC40A1 genes ascompared to control while the 200 IU rHuEPO group pre-sented significant overexpression of Hamp Hfe HJV EPORSLC40A1 Tf TfR2 and BMP6 genes as compared to controland 50 IU rHuEPO group EPO gene was significantly down-regulated in both groups and TMPRSS6 gene expressionwas significantly downregulated only in the 200 IU rHuEPOgroup as compared to the control and 50 IU rHuEPOgroups
The evaluation of the expression of DMT1 and SLC40A1genes in duodenum showed that only the 200 IU rHuEPOgroup presented a significant upregulation for both genes(Figure 4) as compared to the control Furthermore asobserved in liver the expression of EPO in kidney tissue
BioMed Research International 5
Table 2 Hematological and biochemical data at the end of protocol (9 weeks)
Parameters GroupsControl 50 IU rHuEPO 200 IU rHuEPO
EPO (mIUmL) 187 plusmn 025 165 plusmn 023 180 plusmn 021Iron (120583gdL) 18363 plusmn 4158 18863 plusmn 3527 25514 plusmn 11228ab
Ferritin (ngdL) 16513 plusmn 1919 16850 plusmn 4295 16257 plusmn 3981Transferrin (mgmL) 9775 plusmn 868 9938 plusmn 659 9614 plusmn 968Glucose (mgdL) 16088 plusmn 2904 17462 plusmn 2021 18771 plusmn 3743Creatinine (mgdL) 040 plusmn 005 037 plusmn 003 035 plusmn 006Urea (mgdL) 4510 plusmn 192 4505 plusmn 356 4274 plusmn 510Uric acid (mgdL) 094 plusmn 046 070 plusmn 013 103 plusmn 052Bilirubin (ngmL) 0043 plusmn 0016 0054 plusmn 0013 0064 plusmn 0014IL-6 (pgmL) 13810 plusmn 534 13998 plusmn 612 13016 plusmn 506ab
hsCRP (120583gmL) 22931 plusmn 2455 23749 plusmn 2892 24946 plusmn 3349IFN-120574 (pgmL) 2444 plusmn 2307 2145 plusmn 680 2082 plusmn 1013TGF-1205731 (ngmL) 8021 plusmn 499 8003 plusmn 1722 7810 plusmn 555VEGF (pgmL) 536 plusmn 426 584 plusmn 385 604 plusmn 325Results are expressed as mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO IL-6 interleukin-6 hsCRP high-sensitive C-reactive proteinIFN-120574 interferon-120574 TGF-120573 transforming growth factor beta1 VEGF vascular endothelial growth factor
presented a trend towards downregulation in both rHuEPOgroups when compared to the control (Figure 5)
4 Discussion
Antibody-mediated EHPRCA is a rare pathological com-plication associated with the use of recombinant humanerythropoietins and development of anti-EPO antibodiesHowever the use of EPO biosimilars has increased thenumber of cases in the last years [11 20 21] Clinically thatcondition not only abrogates the effect of the rHuEPO butmay also neutralize endogenous EPO leading to severe EHand transfusion-dependent anemia
In this work we developed an animal model of antibody-mediated EH by long-term treatment with a high dose ofrHuEPO (200 IUKg bwweek during 9 weeks) inWistar ratsIn order to confirm that we were in fact in the presence ofan animal model of this hematological disorder we assessedthe parameters typically used as diagnostic criteria forEHPRCA which include normocytic anemia with suddenonset normocellular bone marrow with a selective reduc-tion in red blood cell precursors beyond proerythroblastsreticulocytopenia normal or slightly decreased leucocytecounts and increased serum iron [22] We found that thegroup treated with 200 IUKg bwweek of rHuEPO along 9weeks fits all these criteria and the majority (65) of therats presented detectable anti-rHuEPO antibodies suggest-ing that we achieved an antibody-mediated EH The groupunder 50 IUKg bwweek rHuEPO treatment presented onlyslight changes as compared to control however most ofthe rats (65) presented anti-rHuEPO antibodies at a lowertiter (four times less) suggesting that a longer experimentalprotocol could also lead to the changes associated with EH
Indeed Casadevall (2005) reported that the detection ofanti-erythropoietin antibodies in circulation is not immedi-ately associated with PRCA [23] Moreover no significant
changes were observed in hsCRP IL-6 and IFN-120574 forboth rHuEPO treated groups showing that no inflamma-tory changes could interfere with erythropoiesis and ironmetabolism
Our data suggest that the development of EH was dueto the antibodies directed to rHuEPO that inhibited theerythropoietic stimuli of both rHuEPO and endogenousEPO Indeed in spite of the normal endogenous EPO levelsthat were similar to those of the control group the 200 IUrHuEPO group developed anemia suggesting that the anti-rHuEPO antibodies also neutralized the action of endoge-nous EPO It has been described that the presence of rHuEPOin circulation may increase severalfold the expression ofEPOR [4] Actually we found an overexpression of EPORin the liver of the 200 IU rHuEPO group The decreasedEPO expression observed in kidney and liver tissues forboth rHuEPO groups seems to be in accordance with aphysiological response to the increased circulating levels ofrHuEPO used in the treatment However we should notexclude the hypothesis raised by Piron et al (2001) thatthis anemia could be due to an inappropriate erythropoieticstimulation or to intrinsic failure of erythroid cells to respondto that stimulus [24]
Anemia usually triggers erythropoiesis by increasingEPO production through the hypoxia-inducible factor (HIF)pathway by mobilizing iron from the iron storage pooland by increasing iron absorption in order to face theincreased iron needs for erythropoiesis [25] Consideringthat in antibody-mediated EH the erythropoietic stimulifail due to the inhibition of rHuEPO by anti-rHuEPOantibodies the serum iron should increase as its absorptionandmobilizationwould be triggered by the anemic conditionand hepcidin should be repressed to favour iron absorptionand mobilization Actually we found a significant increasein serum iron in the 200 IU rHuEPO group It is knownthat increasing serum iron induces hepcidin synthesis by
6 BioMed Research International
Hamp
00
05
10
15
20
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SEPO
00
02
04
06
08
10Hfe
00
05
10
15
20
25
HJV
00
05
10
15
20
25EPOR
00
10
20
30SLC40A1
00
10
20
30
TF TfR2
00
20
40
60TMPRSS6
00
05
10
15
20
BMP6
00
10
20
30
lowastlowast
lowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowast
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SRe
lativ
e mRN
A le
vels
18S
Relat
ive m
RNA
leve
ls18
S
00
05
10
15
20
25
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 3 RelativemRNA expression of erythropoietin and iron regulatory proteins in the liver at the end of the protocol (9 weeks) 18S rRNAwas used as reference gene Results are expressed as mean plusmn SD lowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowast119875 lt 0001 versus control group 119875 lt 005119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
a complex pathway Indeed hepcidin encoded by the Hampgene is believed to play a key regulatory role in ironabsorption It controls plasma iron concentration and tissuedistribution of iron by inhibiting intestinal iron absorptioniron recycling by macrophages and iron mobilization from
stores [26] Hepcidin acts by inhibiting cellular iron efflux inhepatocytes enterocytes and macrophages through bindingto ferroportin inducing its degradation [27] Hepatic Hampexpression is regulated by a cohort of proteins includingHfe TfR2HJV BMP6matriptase-2 and transferrin [28ndash30]
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 5
Table 2 Hematological and biochemical data at the end of protocol (9 weeks)
Parameters GroupsControl 50 IU rHuEPO 200 IU rHuEPO
EPO (mIUmL) 187 plusmn 025 165 plusmn 023 180 plusmn 021Iron (120583gdL) 18363 plusmn 4158 18863 plusmn 3527 25514 plusmn 11228ab
Ferritin (ngdL) 16513 plusmn 1919 16850 plusmn 4295 16257 plusmn 3981Transferrin (mgmL) 9775 plusmn 868 9938 plusmn 659 9614 plusmn 968Glucose (mgdL) 16088 plusmn 2904 17462 plusmn 2021 18771 plusmn 3743Creatinine (mgdL) 040 plusmn 005 037 plusmn 003 035 plusmn 006Urea (mgdL) 4510 plusmn 192 4505 plusmn 356 4274 plusmn 510Uric acid (mgdL) 094 plusmn 046 070 plusmn 013 103 plusmn 052Bilirubin (ngmL) 0043 plusmn 0016 0054 plusmn 0013 0064 plusmn 0014IL-6 (pgmL) 13810 plusmn 534 13998 plusmn 612 13016 plusmn 506ab
hsCRP (120583gmL) 22931 plusmn 2455 23749 plusmn 2892 24946 plusmn 3349IFN-120574 (pgmL) 2444 plusmn 2307 2145 plusmn 680 2082 plusmn 1013TGF-1205731 (ngmL) 8021 plusmn 499 8003 plusmn 1722 7810 plusmn 555VEGF (pgmL) 536 plusmn 426 584 plusmn 385 604 plusmn 325Results are expressed as mean plusmn SD a119875 lt 005 versus control b119875 lt 005 versus 50 IU rHuEPO IL-6 interleukin-6 hsCRP high-sensitive C-reactive proteinIFN-120574 interferon-120574 TGF-120573 transforming growth factor beta1 VEGF vascular endothelial growth factor
presented a trend towards downregulation in both rHuEPOgroups when compared to the control (Figure 5)
4 Discussion
Antibody-mediated EHPRCA is a rare pathological com-plication associated with the use of recombinant humanerythropoietins and development of anti-EPO antibodiesHowever the use of EPO biosimilars has increased thenumber of cases in the last years [11 20 21] Clinically thatcondition not only abrogates the effect of the rHuEPO butmay also neutralize endogenous EPO leading to severe EHand transfusion-dependent anemia
In this work we developed an animal model of antibody-mediated EH by long-term treatment with a high dose ofrHuEPO (200 IUKg bwweek during 9 weeks) inWistar ratsIn order to confirm that we were in fact in the presence ofan animal model of this hematological disorder we assessedthe parameters typically used as diagnostic criteria forEHPRCA which include normocytic anemia with suddenonset normocellular bone marrow with a selective reduc-tion in red blood cell precursors beyond proerythroblastsreticulocytopenia normal or slightly decreased leucocytecounts and increased serum iron [22] We found that thegroup treated with 200 IUKg bwweek of rHuEPO along 9weeks fits all these criteria and the majority (65) of therats presented detectable anti-rHuEPO antibodies suggest-ing that we achieved an antibody-mediated EH The groupunder 50 IUKg bwweek rHuEPO treatment presented onlyslight changes as compared to control however most ofthe rats (65) presented anti-rHuEPO antibodies at a lowertiter (four times less) suggesting that a longer experimentalprotocol could also lead to the changes associated with EH
Indeed Casadevall (2005) reported that the detection ofanti-erythropoietin antibodies in circulation is not immedi-ately associated with PRCA [23] Moreover no significant
changes were observed in hsCRP IL-6 and IFN-120574 forboth rHuEPO treated groups showing that no inflamma-tory changes could interfere with erythropoiesis and ironmetabolism
Our data suggest that the development of EH was dueto the antibodies directed to rHuEPO that inhibited theerythropoietic stimuli of both rHuEPO and endogenousEPO Indeed in spite of the normal endogenous EPO levelsthat were similar to those of the control group the 200 IUrHuEPO group developed anemia suggesting that the anti-rHuEPO antibodies also neutralized the action of endoge-nous EPO It has been described that the presence of rHuEPOin circulation may increase severalfold the expression ofEPOR [4] Actually we found an overexpression of EPORin the liver of the 200 IU rHuEPO group The decreasedEPO expression observed in kidney and liver tissues forboth rHuEPO groups seems to be in accordance with aphysiological response to the increased circulating levels ofrHuEPO used in the treatment However we should notexclude the hypothesis raised by Piron et al (2001) thatthis anemia could be due to an inappropriate erythropoieticstimulation or to intrinsic failure of erythroid cells to respondto that stimulus [24]
Anemia usually triggers erythropoiesis by increasingEPO production through the hypoxia-inducible factor (HIF)pathway by mobilizing iron from the iron storage pooland by increasing iron absorption in order to face theincreased iron needs for erythropoiesis [25] Consideringthat in antibody-mediated EH the erythropoietic stimulifail due to the inhibition of rHuEPO by anti-rHuEPOantibodies the serum iron should increase as its absorptionandmobilizationwould be triggered by the anemic conditionand hepcidin should be repressed to favour iron absorptionand mobilization Actually we found a significant increasein serum iron in the 200 IU rHuEPO group It is knownthat increasing serum iron induces hepcidin synthesis by
6 BioMed Research International
Hamp
00
05
10
15
20
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SEPO
00
02
04
06
08
10Hfe
00
05
10
15
20
25
HJV
00
05
10
15
20
25EPOR
00
10
20
30SLC40A1
00
10
20
30
TF TfR2
00
20
40
60TMPRSS6
00
05
10
15
20
BMP6
00
10
20
30
lowastlowast
lowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowast
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SRe
lativ
e mRN
A le
vels
18S
Relat
ive m
RNA
leve
ls18
S
00
05
10
15
20
25
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 3 RelativemRNA expression of erythropoietin and iron regulatory proteins in the liver at the end of the protocol (9 weeks) 18S rRNAwas used as reference gene Results are expressed as mean plusmn SD lowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowast119875 lt 0001 versus control group 119875 lt 005119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
a complex pathway Indeed hepcidin encoded by the Hampgene is believed to play a key regulatory role in ironabsorption It controls plasma iron concentration and tissuedistribution of iron by inhibiting intestinal iron absorptioniron recycling by macrophages and iron mobilization from
stores [26] Hepcidin acts by inhibiting cellular iron efflux inhepatocytes enterocytes and macrophages through bindingto ferroportin inducing its degradation [27] Hepatic Hampexpression is regulated by a cohort of proteins includingHfe TfR2HJV BMP6matriptase-2 and transferrin [28ndash30]
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
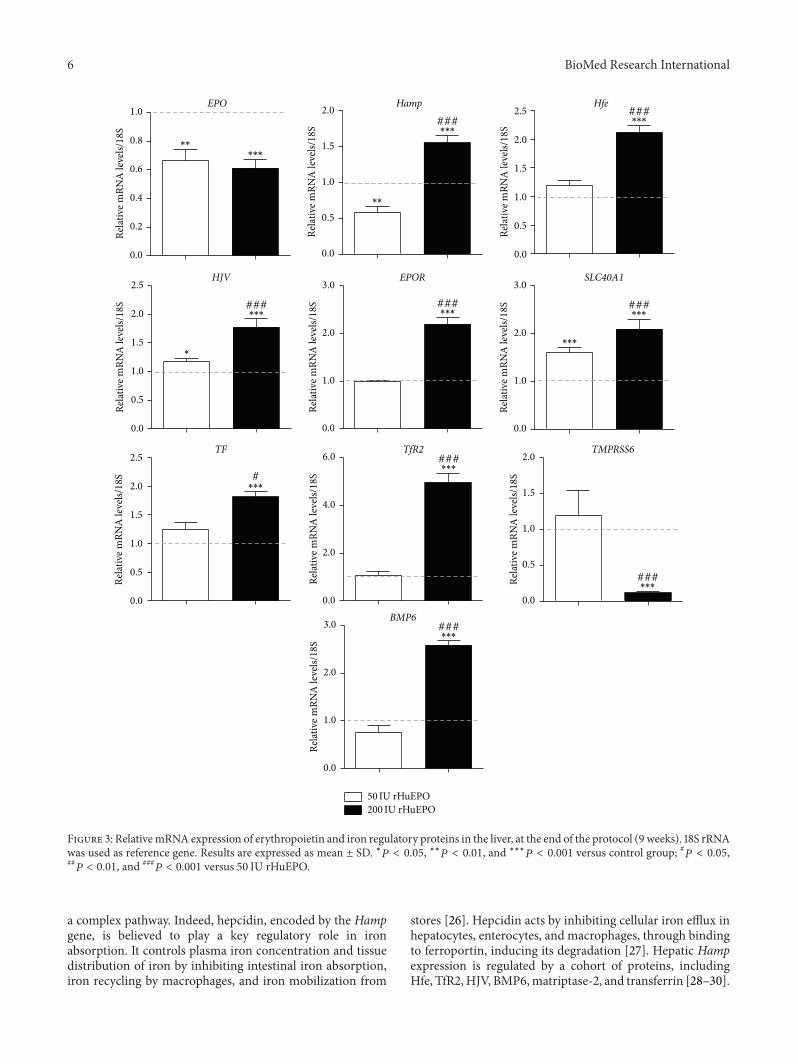
6 BioMed Research International
Hamp
00
05
10
15
20
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SEPO
00
02
04
06
08
10Hfe
00
05
10
15
20
25
HJV
00
05
10
15
20
25EPOR
00
10
20
30SLC40A1
00
10
20
30
TF TfR2
00
20
40
60TMPRSS6
00
05
10
15
20
BMP6
00
10
20
30
lowastlowast
lowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowast
Relat
ive m
RNA
leve
ls18
S
Relat
ive m
RNA
leve
ls18
SRe
lativ
e mRN
A le
vels
18S
Relat
ive m
RNA
leve
ls18
S
00
05
10
15
20
25
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 3 RelativemRNA expression of erythropoietin and iron regulatory proteins in the liver at the end of the protocol (9 weeks) 18S rRNAwas used as reference gene Results are expressed as mean plusmn SD lowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowast119875 lt 0001 versus control group 119875 lt 005119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
a complex pathway Indeed hepcidin encoded by the Hampgene is believed to play a key regulatory role in ironabsorption It controls plasma iron concentration and tissuedistribution of iron by inhibiting intestinal iron absorptioniron recycling by macrophages and iron mobilization from
stores [26] Hepcidin acts by inhibiting cellular iron efflux inhepatocytes enterocytes and macrophages through bindingto ferroportin inducing its degradation [27] Hepatic Hampexpression is regulated by a cohort of proteins includingHfe TfR2HJV BMP6matriptase-2 and transferrin [28ndash30]
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
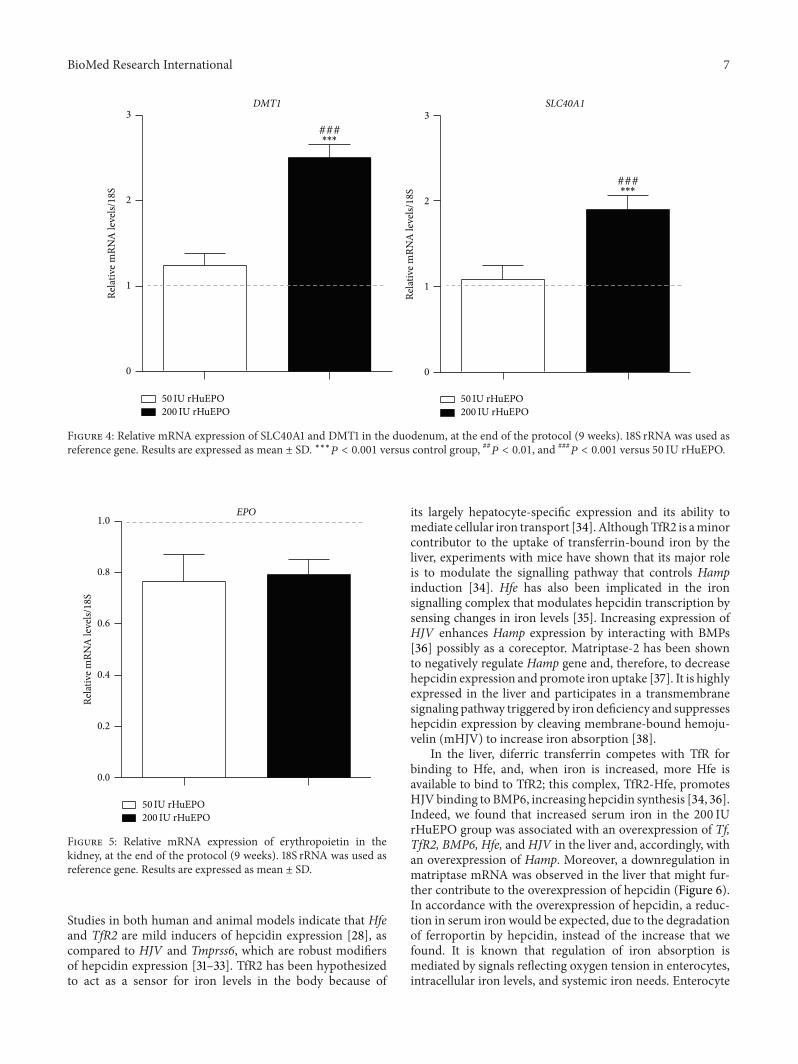
BioMed Research International 7
DMT1
0
1
2
3SLC40A1
50 IU rHuEPO200 IU rHuEPO
Relat
ive m
RNA
leve
ls18
S
0
1
2
3
Relat
ive m
RNA
leve
ls18
S
lowastlowastlowast
lowastlowastlowast
50 IU rHuEPO200 IU rHuEPO
Figure 4 Relative mRNA expression of SLC40A1 and DMT1 in the duodenum at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD lowastlowastlowast119875 lt 0001 versus control group 119875 lt 001 and
119875 lt 0001 versus 50 IU rHuEPO
EPO
00
02
04
06
08
10
Relat
ive m
RNA
leve
ls18
S
50 IU rHuEPO200 IU rHuEPO
Figure 5 Relative mRNA expression of erythropoietin in thekidney at the end of the protocol (9 weeks) 18S rRNA was used asreference gene Results are expressed as mean plusmn SD
Studies in both human and animal models indicate that Hfeand TfR2 are mild inducers of hepcidin expression [28] ascompared to HJV and Tmprss6 which are robust modifiersof hepcidin expression [31ndash33] TfR2 has been hypothesizedto act as a sensor for iron levels in the body because of
its largely hepatocyte-specific expression and its ability tomediate cellular iron transport [34] AlthoughTfR2 is aminorcontributor to the uptake of transferrin-bound iron by theliver experiments with mice have shown that its major roleis to modulate the signalling pathway that controls Hampinduction [34] Hfe has also been implicated in the ironsignalling complex that modulates hepcidin transcription bysensing changes in iron levels [35] Increasing expression ofHJV enhances Hamp expression by interacting with BMPs[36] possibly as a coreceptor Matriptase-2 has been shownto negatively regulate Hamp gene and therefore to decreasehepcidin expression and promote iron uptake [37] It is highlyexpressed in the liver and participates in a transmembranesignaling pathway triggered by iron deficiency and suppresseshepcidin expression by cleaving membrane-bound hemoju-velin (mHJV) to increase iron absorption [38]
In the liver diferric transferrin competes with TfR forbinding to Hfe and when iron is increased more Hfe isavailable to bind to TfR2 this complex TfR2-Hfe promotesHJV binding to BMP6 increasing hepcidin synthesis [34 36]Indeed we found that increased serum iron in the 200 IUrHuEPO group was associated with an overexpression of TfTfR2 BMP6 Hfe andHJV in the liver and accordingly withan overexpression of Hamp Moreover a downregulation inmatriptase mRNA was observed in the liver that might fur-ther contribute to the overexpression of hepcidin (Figure 6)In accordance with the overexpression of hepcidin a reduc-tion in serum iron would be expected due to the degradationof ferroportin by hepcidin instead of the increase that wefound It is known that regulation of iron absorption ismediated by signals reflecting oxygen tension in enterocytesintracellular iron levels and systemic iron needs Enterocyte
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
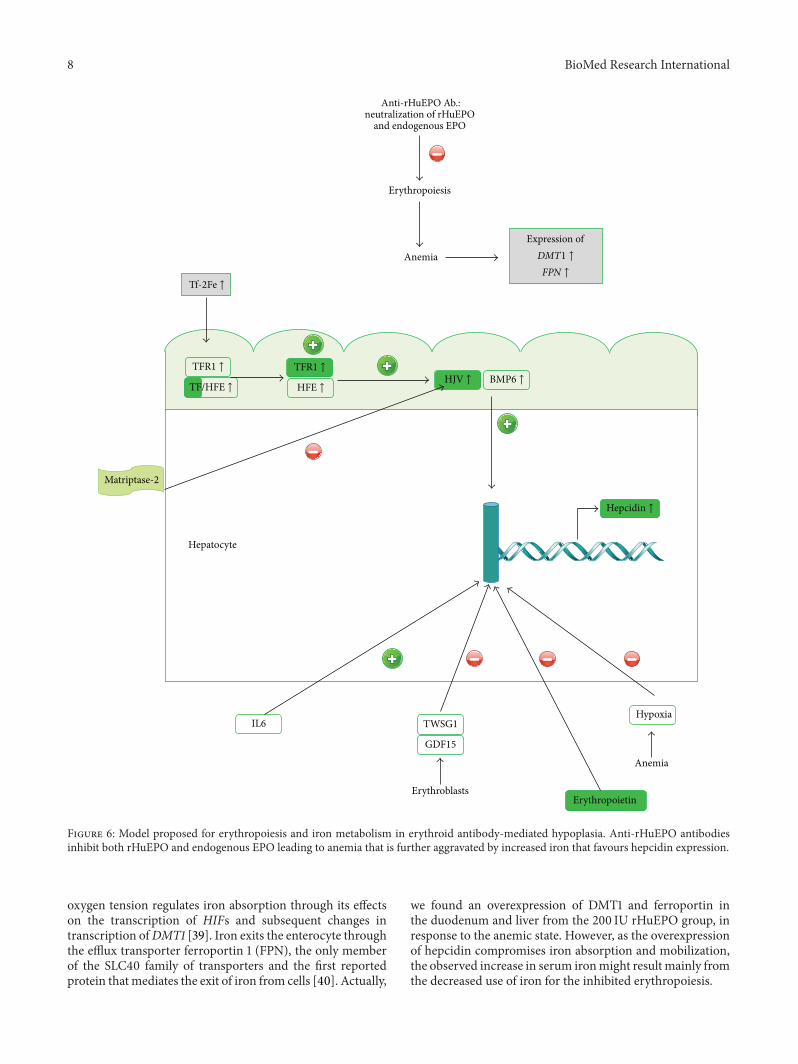
8 BioMed Research International
TWSG1
GDF15
Hypoxia
Erythropoietin
IL6
Matriptase-2
Erythroblasts
Hepatocyte
Anemia
Erythropoiesis
Anemia
Hepcidin uarr
TFR1 uarr
TFHFE uarr
TFR1 uarr
HFE uarrHJV uarr BMP6 uarr
Tf-2Fe uarr
Expression ofDMT1 uarrFPN uarr
Anti-rHuEPO Abneutralization of rHuEPO
and endogenous EPO
Figure 6 Model proposed for erythropoiesis and iron metabolism in erythroid antibody-mediated hypoplasia Anti-rHuEPO antibodiesinhibit both rHuEPO and endogenous EPO leading to anemia that is further aggravated by increased iron that favours hepcidin expression
oxygen tension regulates iron absorption through its effectson the transcription of HIFs and subsequent changes intranscription ofDMT1 [39] Iron exits the enterocyte throughthe efflux transporter ferroportin 1 (FPN) the only memberof the SLC40 family of transporters and the first reportedprotein thatmediates the exit of iron from cells [40] Actually
we found an overexpression of DMT1 and ferroportin inthe duodenum and liver from the 200 IU rHuEPO group inresponse to the anemic state However as the overexpressionof hepcidin compromises iron absorption and mobilizationthe observed increase in serum ironmight result mainly fromthe decreased use of iron for the inhibited erythropoiesis
BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 9
Hypoxia EPO as well as twisted gastrulation protein 1(TWSG1) and growth differentiation factor 15 (GDF15) [41]both produced by erythroblasts is known to downregulatehepcidin synthesis Considering the inhibition of erythro-poiesis through the anti-rHuEPO antibodies the productionof TWSG1 and GDF15 might be reduced and therefore mighbe unable to downregulate hepcidin synthesis We also foundthat rHuEPO treatment did not alter renal function andglucose levels and was not associated with an inflammatoryprocess when compared to the control
Considering the four functionally hepcidin regulatorypathways described [42ndash45] erythropoiesis iron status oxy-gen tension and inflammation we found that in our modelerythropoiesis was blunted leading to anemia iron wasincreased and therewas no inflammation It seems thereforethat the increased iron concentration is the key modulatorof hepcidin synthesis in this type of erythroid hypoplasia Infact it has been recently reported that the reduction of serumiron by iron chelation therapy promoted an improvement inerythropoiesis in PRCA though themechanismwhereby thisis achieved is still unclear [46]
In conclusion our data suggest that in the caseof erythroid antibody-mediated hypoplasiaPRCA inducedby a high rHuEPO dose (200 IU) as erythropoiesis isblunted through anti-rHuEPO antibodies iron concentra-tion becomes the key modulator for hepcidin synthesiswhich will probably contribute to further aggravation ofthe anemia These findings might be important to improvenew therapeutic strategies against rHuEPO resistance andordevelopment of antibody-mediated EHPRCA in patientsunder rHuEPO therapy
Conflict of Interests
The authors declare no conflict of interests
Authorsrsquo Contribution
J C Fernandes and P Garrido have contributed equally tothis paper
Acknowledgments
This study was supported by Fundacao para a Cienciae Tecnologia (FCT PTDCSAU-TOX1142532009 andPestCSAU32822013) and cofinanced by COMPETE FCThas also contributed with a postdoctoral Grant attributed toJoao Fernandes (SFRHBPD819682011) and PhD degreeGrants attributed to PatrıciaGarrido (SFRHBD610202009)and Sandra Ribeiro (SFRHBD798752011)
References
[1] E Moore and R Bellomo ldquoErythropoietin (EPO) in acutekidney injuryrdquo Annals of Intensive Care vol 1 no 1 pp 1ndash102011
[2] D Choi M Kim and J Park ldquoErythropoietin physico- andbiochemical analysisrdquo Journal of Chromatography B BiomedicalApplications vol 687 no 1 pp 189ndash199 1996
[3] P Sathyanarayana A Dev J Fang et al ldquoEPO receptor circuitsfor primary erythroblast survivalrdquo Blood vol 111 no 11 pp5390ndash5399 2008
[4] D W Johnson C A Pollock and I C Macdougall ldquoErythro-poiesis-stimulating agent hyporesponsivenessrdquoNephrology vol12 no 4 pp 321ndash330 2007
[5] N Casadevall E Dupuy P Molho-Sabatier G Tobelem BVaret and PMayeux ldquoAutoantibodies against erythropoietin ina patient with pure red-cell aplasiardquoTheNew England Journal ofMedicine vol 334 no 10 pp 630ndash633 1996
[6] A Hara T Wada S Kitajima et al ldquoCombined pure redcell aplasia and autoimmune hemolytic anemia in systemiclupus erythematosus with anti-erythropoietin autoantibodiesrdquoAmerican Journal of Hematology vol 83 no 9 pp 750ndash7522008
[7] B Viron C-A Dupuy A Kolta and N Casadevall ldquoSuccessfulre-challenge with darbepoetin in a patient with rHu-EPO-induced pure red cell aplasia refractory to immunosuppressivedrugsrdquo Nephrology Dialysis Transplantation vol 23 no 7 pp2416ndash2418 2008
[8] N Casadevall ldquoPure red cell aplasia and anti-erythropoietinantibodies in patients treated with epoetinrdquoNephrology DialysisTransplantation vol 18 supplement 8 pp viii37ndashviii41 2003
[9] S-K Shin S J Moon S K Ha et al ldquoImmunogenicity ofrecombinant human erythropoietin in Korea a two-year cross-sectional studyrdquo Biologicals vol 40 no 4 pp 254ndash261 2012
[10] S K Shin S P Pack J-G Oh et al ldquoAnti-erythropoietinand anti-thrombopoietin antibodies induced after adminis-tration of recombinant human erythropoietinrdquo InternationalImmunopharmacology vol 11 no 12 pp 2237ndash2241 2011
[11] K-U Eckardt and N Casadevall ldquoPure red-cell aplasia due toanti-erythropoietin antibodiesrdquoNephrology Dialysis Transplan-tation vol 18 no 5 pp 865ndash869 2003
[12] N Casadevall J Nataf B Viron et al ldquoPure red-cell aplasia andantierythropoietin antibodies in patients treated with recombi-nant erythropoietinrdquoTheNew England Journal of Medicine vol346 no 7 pp 469ndash475 2002
[13] P Fisch R Handgretinger and H-E Schaefer ldquoPure red cellaplasiardquo British Journal of Haematology vol 111 no 4 pp 1010ndash1022 2000
[14] B Moulouel D Houamel C Delaby et al ldquoHepcidin regulatesintrarenal iron handling at the distal nephronrdquo Kidney Interna-tional vol 84 no 4 pp 756ndash766 2013
[15] S K Srai B Chung J Marks et al ldquoErythropoietin regulatesintestinal iron absorption in a rat model of chronic renalfailurerdquo Kidney International vol 78 no 7 pp 660ndash667 2010
[16] J B Wish ldquoAssessing iron status beyond serum ferritin andtransferrin saturationrdquo Clinical Journal of the American Societyof Nephrology CJASN vol 1 pp S4ndashS8 2006
[17] J M Urra M de la Torre R Alcazar and R Peces ldquoRapidmethod for detection of anti-recombinant human erythropoi-etin antibodies as a new form of erythropoietin resistancerdquoClinical Chemistry vol 43 no 5 pp 848ndash849 1997
[18] M Teixeira P Rodrigues-Santos P Garrido et al ldquoCardiacantiapoptotic and proproliferative effect of recombinant humanerythropoietin in amoderate stage of chronic renal failure in theratrdquo Journal of Pharmacy and Bioallied Sciences vol 4 no 1 pp76ndash83 2012
[19] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 BioMed Research International
[20] J M McKoy R E Stonecash D Cournoyer et al ldquoEpoetin-associated pure red cell aplasia past present and futureconsiderationsrdquo Transfusion vol 48 no 8 pp 1754ndash1762 2008
[21] A V Kharagjitsingh J C Korevaar J P Vandenbroucke etal ldquoIncidence of recombinant erythropoietin (EPO) hypore-sponse EPO-associated antibodies and pure red cell aplasia indialysis patientsrdquo Kidney International vol 68 no 3 pp 1215ndash1222 2005
[22] S-H Bi L-T Cheng W Chen and T Wang ldquoCharacteristicchanges in anti-erythropoietin antibodies in a peritoneal dial-ysis patient who developed pure red cell aplasiardquo Hong KongJournal of Nephrology vol 8 no 2 pp 71ndash74 2006
[23] N Casadevall ldquoWhat is antibody-mediated pure red cell aplasia(PRCA)rdquo Nephrology Dialysis Transplantation vol 20 supple-ment 4 pp iv3ndashiv8 2005
[24] M Piron M Loo A Gothot F Tassin G Fillet and Y BeguinldquoCessation of intensive treatment with recombinant humanerythropoietin is followed by secondary anemiardquo Blood vol 97no 2 pp 442ndash448 2001
[25] V H Haase ldquoHypoxic regulation of erythropoiesis and ironmetabolismrdquoThe American Journal of PhysiologymdashRenal Phys-iology vol 299 no 1 pp F1ndashF13 2010
[26] T Ganz ldquoHepcidin and iron regulation 10 years laterrdquo Bloodvol 117 no 17 pp 4425ndash4433 2011
[27] G C Preza R Pinon T Ganz and E Nemeth ldquoCellularcatabolism of the iron-regulatory peptide hormone hepcidinrdquoPLoS ONE vol 8 no 3 Article ID e58934 2013
[28] A-S Zhang ldquoControl of systemic iron homeostasis by thehemojuvelin-hepcidin axisrdquo Advances in Nutrition vol 1 no 1pp 38ndash45 2010
[29] K E Finberg ldquoUnraveling mechanisms regulating systemiciron homeostasisrdquo Hematology the Education Program of theAmerican Society of Hematology vol 2011 no 1 pp 532ndash5372011
[30] E Canavesi C Alfieri S Pelusi and L Valenti ldquoHepcidin andHFE protein Iron metabolism as a target for the anemia ofchronic kidney diseaserdquoWorld Journal of Nephrology vol 1 no6 pp 166ndash176 2012
[31] Y Cui Q Wu and Y Zhou ldquoIron-refractory iron deficiencyanemia newmolecular mechanismsrdquoKidney International vol76 no 11 pp 1137ndash1141 2009
[32] T Ganz and E Nemeth ldquoThe hepcidin-ferroportin system asa therapeutic target in anemias and iron overload disordersrdquoHematologythe Education Program of the American Society ofHematology vol 2011 pp 538ndash542 2011
[33] A Nai A Pagani L Silvestri et al ldquoTMPRSS6 rs855791modulates hepcidin transcription in vitro and serum hepcidinlevels in normal individualsrdquo Blood vol 118 no 16 pp 4459ndash4462 2011
[34] E L Mackenzie K Iwasaki and Y Tsuji ldquoIntracellular irontransport and storage from molecular mechanisms to healthimplicationsrdquo Antioxidants amp Redox Signaling vol 10 no 6 pp997ndash1030 2008
[35] S Pelucchi R Mariani P Trombini et al ldquoExpression ofhepcidin and other iron-related genes in type 3 hemochro-matosis due to a novel mutation in transferrin receptor-2rdquoHaematologica vol 94 no 2 pp 276ndash279 2009
[36] A-S Zhang F Yang J Wang H Tsukamoto and C A EnnsldquoHemojuvelin-neogenin interaction is required for bone mor-phogenic protein-4-induced hepcidin expressionrdquo The Journalof Biological Chemistry vol 284 no 34 pp 22580ndash22589 2009
[37] L Silvestri A Pagani A Nai I de Domenico J Kaplan andC Camaschella ldquoThe serine protease matriptase-2 (TMPRSS6)inhibits hepcidin activation by cleaving membrane hemoju-velinrdquo Cell Metabolism vol 8 no 6 pp 502ndash511 2008
[38] A Piperno R Mariani P Trombini and D Girelli ldquoHepcidinmodulation in human diseases from research to clinicrdquo WorldJournal of Gastroenterology vol 15 no 5 pp 538ndash551 2009
[39] R E Fleming and P Ponka ldquoIron overload in human diseaserdquoThe New England Journal of Medicine vol 366 no 4 pp 348ndash359 2012
[40] A T McKie P Marciani A Rolfs et al ldquoA novel duodenaliron-regulated transporter IREG1 implicated in the basolateraltransfer of iron to the circulationrdquo Molecular Cell vol 5 no 2pp 299ndash309 2000
[41] T Tanno P Porayette O Sripichai et al ldquoIdentification ofTWSG1 as a second novel erythroid regulator of hepcidinexpression in murine and human cellsrdquo Blood vol 114 no 1pp 181ndash186 2009
[42] E Fung and E Nemeth ldquoManipulation of the hepcidin pathwayfor therapeutic purposesrdquo Haematologica vol 98 no 11 pp1667ndash1676 2013
[43] Q Liu O Davidoff K Niss and V H Haase ldquoHypoxia-inducible factor regulates hepcidin via erythropoietin-inducederythropoiesisrdquo Journal of Clinical Investigation vol 122 no 12pp 4635ndash4644 2012
[44] N K N Shanmugam and B J Cherayil ldquoSerum-induced up-regulation of hepcidin expression involves the bone morpho-genetic protein signaling pathwayrdquo Biochemical and BiophysicalResearch Communications vol 441 no 2 pp 383ndash386 2013
[45] YHamada andM Fukagawa ldquoIs hepcidin the star player in ironmetabolism in chronic kidney diseaserdquo Kidney Internationalvol 75 no 9 pp 873ndash874 2009
[46] M Kojima S Machida A Sato et al ldquoDeferasirox treatmentimproved hematopoiesis and led to complete remission in apatient with pure red cell aplasiardquo International Journal ofHematology vol 98 no 6 pp 719ndash722 2013
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





























