Embed Size (px)
Citation preview

INTRODUCTION
Dramatic changes in embryonic development occur when thefunction of one of the clustered homeotic genes is lacking orwhen one of the genes is active in the wrong place (reviewedby McGinnis and Krumlauf, 1992). In flies, worms, and mice,each of the genes is normally transcribed in a specific regionof the animal along the anterior-posterior body axis. Eight
Drosophila homeotic genes are located in two clusters calledthe Antennapedia and bithorax complexes and are believed tohave originally been part of a single linked group. In mice andhumans, a single cluster has been duplicated twice and somegenes have apparently been lost, leading to four clusters calledthe HOX complexes with a total of 38 genes (reviewed byMavilio, 1993). Nematodes appear to have a single corre-sponding cluster (Wang et al., 1993). The parallels in geneorganization and function in vertebrates, insects, and wormsstrongly supports the view of the clusters as homologous(Duboule and Dolle, 1989; Graham et al., 1989).
Homeotic proteins determine cell fates and the organizationof appropriate structures by acting as transcription factors.Their powerful influence on developmental events is thereforethought to be due to regulation of arrays of downstream ‘target’genes, although only a few such genes have been identified(reviewed by Andrew and Scott, 1992; Botas, 1993; Morata,1993). Targets for Antp include connectin, apterous, spaltmajor, and teashirt. Targets for Ubx include apterous,
connectin, decapentaplegic, Distal-less, scabrous, teashirt,and
β3 tubulin. abd-A regulates many of the same genesregulated by Ubx, and also activates wingless. Some homeotictarget genes encode transcription factors; others encode struc-tural proteins, cell-surface molecules, and growth factors.
The homeotic proteins have in common a 61 amino acidDNA-binding domain called the homeodomain (reviewed byScott et al., 1989; Gehring et al., 1990). Each homeotic proteinconfers a particular pathway of development on the embryo,even when misexpressed in parts of the animal where the geneis normally repressed (reviewed by Hayashi and Scott, 1990).Using chimeric proteins, several investigators have shown thatthe homeodomain is crucial for determining the activity ofhomeotic proteins (Kuziora and McGinnis, 1989; Gibson et al.,1990; Lin and McGinnis, 1992; Chan and Mann, 1993; Zenget al., 1993). This is particularly striking because the home-odomains of many of these proteins are highly similar insequence. To understand better how homeodomain proteinsaffect transcription and, in turn, morphogenesis, it is necessaryto define DNA sequences that are bound and regulated by theseproteins in vivo.
DNA binding of homeotic proteins in vitro has beenexamined using random oligonucleotide selection andfragments of DNA from genes likely to be genuine targets(Beachy et al., 1988; Müller et al., 1989; Affolter et al., 1990;Ekker et al., 1991; Regulski et al., 1991; Capovilla et al., 1994;Zeng et al., 1994). Only slight differences in DNA binding
3605Development 120, 3605-3619 (1995)Printed in Great Britain © The Company of Biologists Limited 1994
The clustered homeotic genes encode transcription factorsthat regulate pattern formation in all animals, conferringcell fates by coordinating the activities of downstream‘target’ genes. In the
Drosophila midgut, the Ultrabithorax(Ubx) protein activates and the abdominalA (abd-A) proteinrepresses transcription of the decapentaplegic (dpp) gene,which encodes a secreted signalling protein of the TGFβclass. We have identified an 813 bp dpp enhancer which iscapable of driving expression of a lacZ gene in a correctpattern in the embryonic midgut. The enhancer is activatedectopically in the visceral mesoderm by ubiquitousexpression of Ubx or Antennapedia but not by Sex combsreduced protein. Ectopic expression of abd-A represses theenhancer. Deletion analysis reveals regions required forrepression and activation. A 419 bp subfragment of the 813
bp fragment also drives reporter gene expression in anappropriate pattern, albeit more weakly. Evolutionarysequence conservation suggests other factors work withhomeotic proteins to regulate dpp. A candidate cofactor, theextradenticle protein, binds to the dpp enhancer in closeproximity to homeotic protein binding sites. Mutation ofeither this site or another conserved motif compromisesenhancer function. A 45 bp fragment of DNA from withinthe enhancer correctly responds to both UBX and ABD-Ain a largely tissue-specific manner, thus representing thesmallest in vivo homeotic response element (HOMRE)identified to date.
Key words: homeotic, homeodomain, decapentaplegic, TGFβ,midgut development, Drosophila
SUMMARY
Regulation of a decapentaplegic midgut enhancer by homeotic proteins
J. Robert Manak, Laura D. Mathies and Matthew P. Scott*
Departments of Developmental Biology and Genetics, Howard Hughes Medical Institute, Stanford University School of Medicine,Stanford, California 94305-5427, USA
*Author for correspondence

3606
specificity have been observed for different homeotic proteinsin vitro (e.g. Capovilla et al., 1994), although these differencesmay be significant in vivo. The best defined homeotic responseelement known to be active in a proper pattern in vivo is theDeformed autoregulatory element, which has been defined asa 120 bp element containing a Deformed binding site and a siteof action of other undefined factors (Zeng et al., 1994). Bothsites are required for enhancer function. In other cases as well,DNA binding proteins may interact with the homeotic proteinsto facilitate their action, perhaps by providing additional speci-ficity in binding site selection. The best candidate for such acofactor is the product of the extradenticle (exd ) gene, a genewhich affects the outcome of homeotic protein functionwithout altering where the homeotic genes are expressed(Peifer and Wieschaus, 1990). exd encodes a homeodomainprotein closely related to the mammalian proto-oncogene Pbx1(Rauskolb et al., 1993) and its family members. Recently, tworesearch groups have provided evidence that UBX and EXD
bind cooperatively to DNA (Chan et al., 1994; van Dijk andMurre, 1994). One group suggests that these proteins collabo-rate in activating the dpp midgut enhancer (Chan et al., 1994).
The regulation of Drosophila midgut development has beenuseful for identifying targets of homeotic genes. Four of theclustered homeotic genes, Sex combs-reduced (Scr), Antenna-pedia(Antp), Ubx, and abd-A, plus exd, are expressed in themidgut mesoderm. In the mesoderm, the homeotic genes areexpressed in non-overlapping anterior-posterior domains (Fig.1). The homeotic genes are required for the formation of draw-string-like constrictions in three parts of the midgut and for theoutgrowth of pockets called caeca at the anterior of the midgut,thus dividing the midgut into compartments (Tremml andBienz, 1989; Reuter and Scott, 1990). These morphologicalchanges may be visible manifestations of additional differen-tiation events that functionally distinguish midgut cells alongthe anterior-posterior axis.
The center constriction forms at the junction between theUbx and abd-A expression domains. Ubx activates transcrip-tion of decapentaplegic (dpp), which encodes a TGFβhomolog, while in the adjacent more posterior cells abd-Aactivates wingless (wg), which is a member of the Wnt classof signalling molecules (Fig. 1; Immerglück et al., 1990;Reuter et al., 1990; Hursh et al., 1993). UBX activation of dppappears to be direct, based on compensatory mutations con-structed in a dpp enhancer and in the regulating protein(Capovilla et al, 1994); it is unclear whether ABD-A directlyrepresses dpp by binding to its regulatory regions. Both dppand wg proteins move into the underlying endoderm wherethey are required for the proper expression and localization ofthe homeotic gene labial (Immerglück et al., 1990; Reuter etal., 1990; Tremml and Bienz, 1992). The center constrictionfails to form in the absence of either dpp or wg. Therefore, inaddition to being regulated by homeotic proteins, these targetsare clearly implicated in carrying out morphogenetic functions.
We have identified a small part of the dpp gene capable ofdirecting expression of lacZ in the midgut cells where dpp isexpressed. We report our analysis of this enhancer element.
MATERIALS AND METHODS
Construction of dpp-lacZ and bacterial expression genefusions A 3.1 kb KpnI/EcoRI fragment of plasmid pCS7276 (from J. Masucciand F.M. Hoffmann) was subcloned into pTZ19U (USBiochemical),which produced pTZCS K/R. From this plasmid, a BamHI/EcoRIfragment (813 bp; this fragment was sequenced by the Sanger dideoxymethod), a KpnI/EcoRI fragment (3.1 kb), a SphI/BamHI fragment(2.0 kb), a SacII/KpnI fragment (1.0 kb), and a BamHI/SacII fragment(1.2 kb), were individually subcloned into the P-element transforma-tion vector pC4PLZ. pC4PLZ (K. Wharton and S. Crews, unpublisheddata) contains a basal P-element promoter fused to the lacZ gene andis marked with the white gene. To make the XhoI/PstI fragment fusedto lacZ, pTZ CS K/R was digested with XhoI and blunted with DNApolymerase I, Klenow fragment. Phosphorylated PstI linkers(GCTGCAGC) were ligated onto the blunt ends. The 0.4 kp PstIfragment was subcloned in both orientations into pTZ18U cut withPstI. One of the resulting plasmids, pTZ18 P/X-, was cut with HindIII(in the pTZ polylinker) and blunted with Klenow. BamHI linkers(CGGATCCG) were ligated onto the blunt end and the plasmid wasdigested with BamHI. This produced the 413 bp fragment which was
J. R. Manak, L. D. Mathies and M. P. Scott
wgdpp
abd-AUbx
Mesoderm
Endodermdpp
(protein)
lab
dpp
PS7 PS8
Ubx
wg(protein)
Fig. 1. Diagram of regulatory relationships between Ubx, abd-A,dpp, and wg. Ubx is expressed in PS7 of the visceral mesoderm, asdetermined using a segmentation gene promoter fused to lacZ as amarker (Bienz and Tremml, 1988). abd-A is expressed in theparasegment 8 region of the mesoderm just posterior to the Ubxdomain and in more posterior parts of the mesoderm. The centralconstriction of the midgut forms at the junction betweenparasegments 7 and 8 after the expression of all the genes discussedhere is established. dpp expression is coincident with Ubx and isactivated by it, while wg is activated in parasegment 8 by abd-A(Immerglück et al., 1990; Reuter et al., 1990). wg is also dependentupon dpp for expression in PS8 (Immerglück et al., 1990). dpp isnecessary to maintain Ubx expression in PS7 (Immerglück et al.,1990; Panganiban et al., 1990b). dpp and wg proteins move into theunderlying endoderm (Panganiban et al., 1990a; Reuter et al., 1990);no dpp or wg transcripts are detectable in the endoderm. Therefore, asegmented mesoderm causes local differentiation of an initiallyunsegmented endoderm. One indicator of the differentiation is theexpression and localization of the homeotic gene labial (lab) inresponse to the dpp and wg signals (Immerglück et al., 1990; Reuteret al., 1990). abd-A is a repressor of Ubx transcription inparasegment 8 and more posterior regions (Bienz and Tremml,1988). If Ubx is expressed with a heat-inducible promoter in theposterior regions it is refractory to abd-A repression and the UBXaccumulates to high levels. However Ubx is prevented fromactivating its target dpp unless abd-A function is removed (Reuter etal., 1990). Ubx, abd-A, dpp, and wg are all required for the formationof a proper central constriction of the midgut.

3607dpp regulation by homeotic genes
subcloned in both orientations into the vector pC4PLZ. The 261 bpfragment was produced by digesting pTZ18 P/X- with BssH2 andXhoI, blunting and religating. The resulting construct was digestedwith BamHI and the 261 bp fragment was subcloned in the forwardorientation into pC4PLZ. pC4PLZ2X45mer was produced by ligationof annealed and phosphorylated oligos (GATCCAATTGCA-GCGCGCATTCAAATTTATTACTAATTGGGTGTGAATTG andGATCCAATTCACACCCAATTAGTAATAAATTTGAATGCGC-GCTGCAATTG) into the BamHI site of pC4PLZ. A double insertwas selected and sequenced. The resulting plasmid contains two directrepeats of the 45mer in reverse orientation with relation to the tran-scription start site. The delta DR construct was made as follows: the0.4 kb PstI fragment from pTZ18 P/X- was subcloned into the PstIsite of pBS II S/K+ so that the the 5’ end of the enhancer fragmentwas situated toward the KpnI end of the polylinker. The polylinkerEcoRI site was destroyed by digestion with EcoRI, blunting withKlenow and religation. The plasmid was digested with SphI, whichcuts in the direct repeat, and the SphI site was destroyed by chew-back blunting with T4 DNA polymerase. Three EcoRI linkers weresubcloned onto the blunt ends. The sequence thus readsGCTG(GGAATTCC)3CTGC from nucleotide 185 in Fig. 9. A 0.4 kbKpnI/BamHI fragment from this plasmid was subcloned into pC4PLZin the forward orientation.
Plasmid ptd48-3, which contains exd cDNA (Rauskolb et al.,1993), was digested with SphI and the 1 kb exd fragment, encodingamino acids 176 to 376, was ligated into pUC19 digested with SphI.The resulting plasmid pUCexd Sph+ was digested with PstI andHindIII (sites within the pUC polylinker; PstI site at 5′ end of exdfragment) and the 1 kb fragment was ligated into the bacterialexpression vector pMal c2 (New England Biolabs) digested with PstIand HindIII. This construct (pMal c2 exd Sph) encodes a 26×103 MrEXD polypeptide (which includes part of the N terminus, the home-odomain and all of the C terminus) fused to a 43×103 Mr maltosebinding protein (MBP).
Mutation of the EXD in vitro binding site was performed on boththe 261 and 419 bp enhancer fragments with the U.S.E. mutagenesiskit (Pharmacia) exactly as described in the accompanying protocol.The sequence of the oligonucleotide used to introduce the mutationwas CGAAATGGGTGCTAAGCTTTAGGCCTTTGATCTGC. Themutation created a HindIII site, which was used for screeningpurposes, and converts the sequence ATCAATTA to AAAGCTTA(see Fig. 9).
Library screeningA Drosophila virilis genomic phage library constructed in lambdaEMBL3 by John Tamkun was screened at moderate stringency(hybridization at 65°C in 4× SSPE, 1% sodium dodecyl sulfate, 0.5%non-fat dry milk; washes at 65°C in 2× SSPE, 0.2% sodium dodecylsulfate, 0.05% sodium phosphate) using an 813 bp BamHI to EcoRIfragment isolated from plasmid pTZCS K/R (see below). PstI/HindIIIand PstI/EcoRI fragments of the phage clones that hybridized to theprobe were subcloned into pUC19 to produce pUCvirP/H andpUCvirP/R. The inserts in both plasmids were fully sequenced on bothstrands.
Fly stocks and P-element injectionsAll P-element vector constructs were co-injected with p∆2-3 trans-posase DNA as helper (D. Rio, unpublished data) into Df(l)w1118
embryos as previously described (Rubin and Spradling, 1982). At leastthree independent transformant lines of each construct were generated.
Immunohistochemistry and antibody sourcesImmunostaining was as described in Mathies et al. (1994). Antiserato β-gal protein were generated in rats and rabbits. The rabbit antibodywas affinity purified. The antibody to dpp protein was the kind gift ofF. M. Hoffmann.
Heat shock induction of homeotic proteinsVirgin females from a stock containing the 813 bp enhancer con-structs homozygous on the third chromosome were crossed to malescontaining heat shock inducible homeotic protein constructs (Mannand Hogness, 1990; Zeng et al., 1993). Embryos were collected for10 hours from this transient cross, then induced by heat shock. Theembryos were transferred to 37°C three times for 30 minutes with arecovery at RT for 30 minutes between heat pulses. The embryos werethen aged for 2.5 hours, fixed, and stained for β-gal expression asdescribed in Mathies et al. (1994).
Expression and purification of proteinsDNA-affinity purified UBX protein was provided by B. Johnson andM. Krasnow (Johnson and Krasnow, 1990). ABD-A protein wasprepared as described in Appel and Sakonju, 1993 using plasmidpNB40 abd-A 65 UAC except that a Pharmacia Sephadex column wasused in the FPLC purification. ABD-A protein was further purified byDNA-affinity column chromatography as described (Kadonaga,1991). Briefly, two oligonucleotides of the sequenceGATCCA(TTA)11TTG and GATCCA(ATA)11ATG were annealedand ligated to form concatamers which were then attached to CNBr-activated sepharose CL-2B beads (Pharmacia). ABD-A proteinextract was applied to the DNA affinity column and the ABD-Aprotein was subsequently eluted with a series of concentrations of KClranging from 0.2 M to 1 M. Based on SDS-polyacrylamide gel elec-trophoresis followed by Coomasie Brilliant Blue staining, we estimatethe final protein preparation is greater than 90% pure.
Expression and purification of the 69×103 Mr MBP/EXD fusionprotein from pMal c2 exd Sph was performed exactly as described inthe Protein Fusion and Purification System protocol from NewEngland Biolabs, but using the bacterial strain DH5a. Bacterialcultures harboring the expression plasmid were induced with IPTG.The fusion protein was purified on an amylose column and eluted withmaltose. Similar DNAse I footprint results were obtained with theMBP/EXD fusion protein as well as with protein cleaved with FactorXa, which separates the MBP domain from the EXD polypeptide.Based on staining SDS-polyacrylamide gels with Coomasie BrilliantBlue, we estimate that the eluted fusion protein is greater than 90%pure.
DNase I footprint assaysDNase I protection assays were performed as described by Hoey etal. (1988) except that 0.02 to 1µg of poly dI/dC was used as non-specific competitor DNA for each reaction. In addition, 1µg of bovineserum albumin was added to each reaction to stabilize protein. TheDNA fragments were electrophoresed in 6% polyacrylamide/7 M ureagels. DNA fragments were labelled with [α32P]dCTP and/or dATPand AMV reverse transcriptase. G/A sequencing lanes were preparedas described by Maxam and Gilbert, (1980).
RESULTS
Identification of a dpp midgut enhancerdpp is transcribed in three regions of the midgut visceralmesoderm, overlying where the gastric caeca (GC) form, justanterior to where the central constriction forms (parasegment7, PS7) and, nearly undetectably, close to the posterior con-striction site (St. Johnston and Gelbart, 1987; Fig. 3A).Previous work (Hursh et al., 1993; Masucci and Hoffmann,1993) led to the identification of a midgut expression controlregion within map coordinates 71.3-75.9, several kilobasesupstream of the dpp promoters (Fig. 2). Fragments from thisregion were joined to a short relatively inactive promoter fusedto lacZ and introduced into flies using a P element vector. At

3608
least three independent lines were established for eachfragment, and the pattern of expression was observed with anti-bodies against β-galactosidase (β-gal) protein.
Three fragments (3.1 kb, 813 bp, 419 bp) containing the 419bp sequence drive appropriate midgut visceral mesodermexpression (Fig. 2), although expression from the 419 bpconstruct is somewhat reduced compared with the larger two.Three fragments upstream of the 3.1 kb did not producedetectable β-gal protein. We shall hereby refer to the 813 or419 bp constructs as dpp813 and dpp419.Expression of dpp813 and dpp419 is shown in Fig.3B,C. The precise correspondence of the endoge-nous dpp pattern and the transgene expression isdemonstrated by a double label experiment fordpp813 (Fig. 3D). Both enhancer-lacZ transgenesare expressed more strongly in the PS7 domainthan in the gastric caeca domain, while the endoge-nous gene is detected at roughly equal levels inboth domains (compare Fig. 3A with 3B,C). Bothdpp813 and dpp419 were used to examine regula-tion by homeotic proteins.
Regulation of the enhancer element byhomeotic proteinsLike dpp itself (Reuter et al., 1990), dpp813 isectopically activated in the visceral mesoderm byubiquitous expression of Ubx (Fig. 4A,B). Also,like the dpp gene, the ectopic activation is onlydetected in visceral mesoderm cells that do notcontain ABD-A. Ubiquitous expression of abd-Acauses a decrease in the expression of the dpp-lacZconstruct in the visceral mesoderm (Fig. 4E,F).Ubiquitously expressed Scr, which does notnormally affect dpp expression in the midgut,produced only a very slight ectopic induction justanterior to PS7 in some embryos (data not shown).In contrast ubiquitous Antp ectopically induces thedpp enhancer through most of the anterior midgutin a patchy pattern, albeit much less strongly thanUbx (compare Fig. 4B with 4D). Expression ofdpp419 in Ubx and abd-A mutant embryos is con-sistent with the results obtained with dpp813 in theexperiments above (data not shown).
Isolation of the corresponding DNAfragment from Drosophila virilisComparisons of enhancer sequences from D.melanogaster and D. virilis have been useful foridentifying important control elements (Kassis etal., 1986; Fortini and Rubin, 1990; Maier et al.,1990). The two fly species are thought to have hada common ancestor about 60 million years ago(Beverley and Wilson, 1984). A single-copy bandwas detected upon probing restriction enzyme-digested genomic D. virilis DNA (data not shown),and a corresponding fragment was obtained inphage clones from a genomic D. virilis library. TheD. melanogaster and D. virilis fragments weresequenced. The two sequences were aligned bycomputer and eye to optimize the number ofmatches.
Fourteen regions of dramatic sequence conservation standout (Fig. 5A,B). About 0.5 kb of the D. virilis sequence is notrepresented in the D. melanogaster sequence. All the patchesof sequence similarity are colinear and identically oriented, andthe spacing between them reasonably well preserved,excepting the 0.5 kb insert. Because conserved sequences fromregions 5′ and 3′ of the 419 bp fragment are present, it is likelythat the region of D. virilis DNA studied contains all thesequence relevant to the dpp419 expression pattern.
J. R. Manak, L. D. Mathies and M. P. Scott
lacZTATA
lacZ
lacZ
lacZ
lacZ
lacZ −−
−−− −
TATA
TATA
TATA
1.2 kb
1.0 kb
2.0 kbReverse
Forward
Forward
+++
+++TATA
TATA
PS7 GC
C.
419 bp
3.1 kb
0.8 kb
pC4PLZ
Encodes nuclearß-gal protein
1 2 3 4 5 6 7 8 9 10 11 12 kb
P-lacZwhite 3' P end5' P endpUC
dpp fragments insertedinto polylinker
Polylinker
Orientationtested:
Forward
Reverse
Both
71.3 75.9Kpn I Sph I Sac II BamHI
EcoRIPst I Xho I
dpppromoter
A.
B.
Sal 1
lacZTATA261 bp ++
BssH2Pst I
Forward
Fig. 2. The decapentaplegic midgut control region. The general region required forexpression of dpp in the embryonic midgut was previously defined (Hursh et al.,1993; Masucci and Hoffmann, 1993). There are several different dpp promoters;the one closest to midgut regulatory elements is located at position 78 (Masucciand Hoffmann, 1993), about 3 kb from the position of the 419 bp enhancer shownin the present study to control midgut expression. Several different fragments (A)from the region were tested for their ability to direct lacZ expression in the midgut.The fragments shown in B were tested in the vector shown in C. The orientationstested are indicated; ‘forward’ indicates that the 5′ to 3′ orientation of the fragmentrelative to the transgene promoter was the same as found in the dpp gene, while‘reverse’ indicates the fragment was flipped end for end. PS7 stands for visceralmesoderm expression near the central constriction; GC stands for the expression ofdpp near the gastric caeca. The pC4PLZ vector contains a P element promoterdriving expression of a β-gal protein containing a nuclear targeting signal(Materials and Methods).

3609dpp regulation by homeotic genes
Locations of binding sites for UBX and ABD-AproteinsThe 419 bp fragment is sufficient to direct proper midgutexpression, so we tested whether homeotic proteins can bindthe fragment. DNase I footprint experiments on this fragmentwere done using bacterially produced full-length UBX andABD-A proteins purified to near homogeneity (Fig. 6A-D).Both proteins bind particularly well to a cluster of sites (sitesF-I, Figs 6 and 9) located within about 100 bp of DNA.Another region (site E) binds both proteins, but with consid-erably lower affinity. UBX protein binds better to the assortedenhancer sites than ABD-A: roughly four times as much ABD-A is required to achieve the extent of protection provided byUBX. However, in contrast to the conclusions reached byCapovilla et al. (1994), we do not observe obvious differencesin affinity of UBX versus ABD-A with respect to differentbinding sites. Moreover, we observe ABD-A binding to site B(site 4a as described in Capovilla et al., 1994). Binding sitesE,F and I, which correspond to sites 3, 2, and 1, respectivelyin Capovilla et al. (1994), are located in highly conservedblocks of sequence, and each contains at least one TAATmotif, a sequence known to be a common motif in homeo-domain binding sites (Laughon, 1991). Binding sites A and B,
which correspond to sites 4b and 4a (Capovilla et al., 1994)are located in a region of poor homology. Neither site A norB has a TAAT motif.
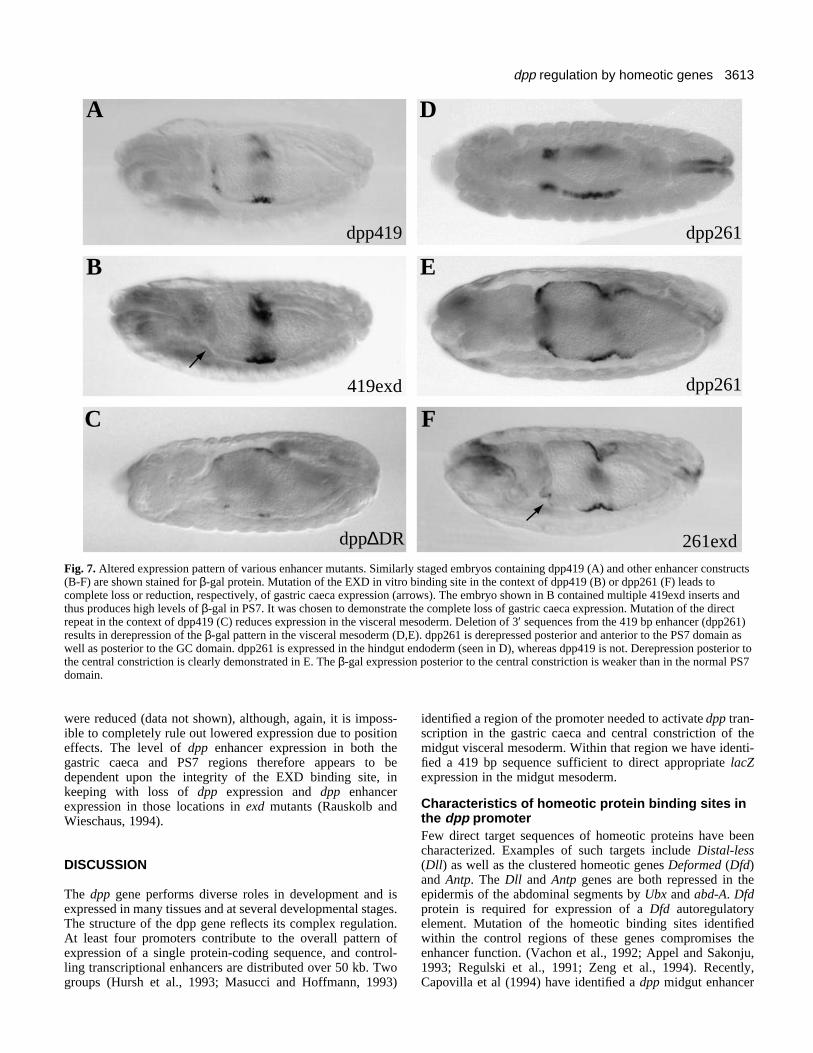
Partial deletion of the enhancer causes ectopicexpressionThe 3′ region of the 419 bp fragment was removed to producethe 261 bp construct (dpp261; Figures 2 and 9), deletinghomology blocks 11 and 12, including one of the strongestbinding sites for homeotic proteins. dpp261 embryos showstriking derepression throughout the midgut mesoderm, sug-gesting that abd-A and other factors can no longer repressexpression in the posterior and anterior midgut (Fig. 7).Bringing the 261 bp enhancer closer to the promoter region ofthe lacZ vector (see Materials and Methods) could cause analtered expression pattern, but the dramatic changes inexpression suggest the deletion, rather than position, is respon-sible. Expression remains highest in the central and anteriorparts of the mesoderm, with strong ectopic expression imme-diately posterior to the normal gastric caeca domain. Strongexpression is also observed in cells where abd-A is expressed,in a gradient tapering to a low level at the posterior. While thetissue specificity is preserved in the midgut, in the hindgut
Fig. 3. Expression patterns of midgut enhancer fragments. The β-gal expression pattern of the 419 bp and 813 bp enhancers correspondsprecisely to the pattern of dpp protein in the midgut visceral mesoderm. dpp expression in a stage 14 wild-type embryo is detected in themidgut cells anterior to the central constriction (ps7) and in the cells that will overly the gastric caeca (gc) shown here in A. The 813 bpfragment (B) and the 419 bp fragment (C) drive β-gal expression in the same two regions of the midgut mesoderm. Both enhancers activatereporter gene expression at higher levels in PS7 than in the GC domain. The enhancer expression domains are precisely the same as theendogenous gene as demonstrated by double label experiments using a confocal microscope (D : dpp protein in red, β-gal protein in green).Regions of overlap in the visceral mesoderm appear yellow in the image (the strong yellow signal in the upper right is yolk auto fluorescence).Separate images of β-gal (E) and dpp (F) protein in PS7 of the same embryo are shown at higher magnification beneath the double labelledimage. gc, gastric caeca; ps7, parasegment 7.

3610
derepression is observed in the endoderm (Fig. 7D). The strongbinding site deleted in constructing dpp261, Site I, maytherefore be a target for repression by ABD-A.
A direct repeat sequence is required for enhancer-driven high-level expressionThe conservation of a direct repeat sequence located aroundnucleotide 192 suggested that this sequence might be importantfor enhancer function. We compared five lines harboring amutation in this sequence in the context of dpp419 (dpp∆DR;see Materials and Methods) with five lines harboring dpp419.The dpp∆DR lines showed reduced expression in the visceralmesoderm (in both gastric caeca and PS7) compared with thedpp419 lines (compare Fig. 7A with C). However, we cannotrule out the possibility that the decreased expression is due tothe insertion site of the constructs.
A 45 bp fragment of the dpp midgut enhancerresponds to homeotic genesSite I and/or its surroundings are required for repression in theanterior and posterior visceral mesoderm. Site F is the otherstrong homeodomain protein binding site. Based on the cross-species homology data, an oligonucleotide was synthesizedcontaining 45 bp centered around site F (Fig. 8). Two copiesof the sequence were built into the lacZ vector, producing the
construct dpp45. Expression, especially early, is mainlylocated within PS7 of the visceral mesoderm (Fig. 8A). Somepatchy ectopic expression is observed anteriorly, as well asaround the third constriction at later stages of development.Expression of the dpp 45mer in PS7 is narrower than thatobserved for either dpp419 or dpp813 (compare expressionindicated by closed arrow in Fig. 8A with PS7 expression inFig. 3C). Although tissue specificity is generally maintained,some expression is observed in glial cells as well as transversemuscles of abdominal segments 1 to 3 (open arrows in Fig.8A). In embryos that contain ubiquitous UBX driven by a heatshock promoter, expression from dpp45 is observed within aswell as anterior to PS7 (Fig. 8B). No expression is observedposterior to PS7. This is exactly the behavior of dpp813 andthe dpp gene itself, and suggests that the 45mer responds toboth UBX and ABD-A.
Locations of binding sites for the EXD proteinPrevious studies suggest that the exd gene product might workin conjunction with a subset of the homeotic proteins,including Ubx and abd-A, to regulate homeotic target genes(Peifer et al., 1991; Rauskolb et al., 1993). dpp expression inPS7 is greatly reduced in exd mutant embryos (Rauskolb andWieschaus, 1994), suggesting that Ubx and exd might collab-orate to regulate dpp in PS7. An EXD fusion protein binds to
J. R. Manak, L. D. Mathies and M. P. Scott
A
C D
F
B
E
WT
WT
WT
HS-Ubx
HS-Antp
HS-abd-AFig. 4. Ectopic induction of the dpp enhancer by homeotic proteins. The 813 bp dpp enhancer is activated by ectopic induction of UBX orANTP, and repressed by ectopic ABD-A. The wild-type expression pattern of the dpp enhancer is seen in A, C, and E at the same stage as theheat shock-induced embryos in B, D, and F. Induction of UBX results in ectopic expression of the enhancer in the midgut visceral mesodermcells anterior to the central constriction (B). The enhancer is also induced in the same cells by ubiquitous production of ANTP (shown in D),but at lower levels than is seen with UBX. Ectopic expression of ABD-A represses the enhancer in the PS7 domain (F).

3611dpp regulation by homeotic genes
1 2 3 4 5 6 7 8 9 10 11 12 13 14
813
1413(including
insert)
68/8
5
28/3
8
28/3
9
47/5
410
/12
37/3
9
58/7
0
26/3
319
/19
29/4
4
50/6
5
32/7
1
34/3
7
35/7
7
DM
DV
493 bp insert
AA Fig. 5. Comparison of the sequences of the D. melanogaster and D.virilis dpp enhancers. The sequences of the 813 bp D. melanogastermidgut enhancer and the 1413 bp D. virilis fragment weredetermined and compared. Dot matrix computer comparisons andfurther refinements by eye were used to identify similar sequences.Conserved regions are interspersed with blocks of very similarsequence. A summary of the positions of the similar blocks (A) iscorrelated by numbers with the specific sequences shown in B. Theratios in the parentheses above homology block numbers indicateexact identity matches over total number of nucleotides within thehomology blocks of D. melanogaster. The assignments of numbersto conserved blocks are made only to facilitate discussion of thesequences. In most cases the divisions between the numbered blocksare placed where differences in spacing between conserved
sequences occur in thetwo species. The largebrackets indicate theextent of the 261 and 419bp enhancers. These twosmaller enhancerfragments have the same5′ end and different 3′ends. The box at thebottom contains theblock of D. virilissequence which belongsat the insertion siteshown. This is the onlymajor difference inspacing or orientationbetween the two species.The orientation shown is5′ to 3′ with respect tothe dpp gene.
B

3612
the 419 bp fragment (Figs 6D and 8, EXD) overlapping the Ebinding site recognized by UBX and ABD-A. The centralsequence in this site (ATCAATTAG) is almost identical to theconsensus binding site of the exd human homolog PBX1(ATCAATCA[A]) determined by in vitro analyses (van Dijket al., 1993; LeBrun and Cleary, 1994).
Enhancer function is affected in flies that harbor a mutationof this sequence in the context of the 419 bp (419exd) or 261
bp (261exd) enhancers (see Fig. 7B and F, respectively).Gastric caeca expression is abolished in the 419exd flies andis greatly reduced in 261exd flies (compare loss of gastriccaeca expression as indicated by an arrow in Fig. 7B and Fwith Fig. 7A and E, respectively). To test whether expressionin PS7 is affected, expression levels in five lines of 419exd flieswere compared with those in five lines of dpp419 flies. Overalllevels of expression in PS7 of flies bearing the mutant enhancer
J. R. Manak, L. D. Mathies and M. P. Scott
Xho IPst I Sph IStu I156 190 4191
IV
I
II
III
V
419 bp dpp enhancer
VI
**
*
*
**
AA
B C D
Fig. 6. DNase I ‘footprint’ experiments with UBX, ABD-A, and EXD proteins.Proteins were expressed in bacteria and partially purified. The pattern of DNase Icleavage sites in the presence of control carrier protein was compared toincreasing amounts of homeotic protein also in the presence of carrier protein.Several different fragments from the 419 bp enhancer were used (A); each waslabeled with 32P at the position shown by an asterisk. Photographs ofrepresentative autoradiograms are shown in B, C, and D, with the fragmentnumbers corresponding to the fragments numbered in A. UBX and ABD-Agenerally bind the same sites. The positions of the protected sequences are shownin Fig. 9. Bubbles next to the footprints indicate protected regions. Very weaklyprotected sequences such as in lanes 9 and 10 below site D, left panel, are notmarked as binding sites. G/A; Maxam-Gilbert (Maxam and Gilbert, 1980)sequencing reaction. 2 µl of UBX is approximately 30 ng of protein; 2 µl ofABD-A is approximately 120 ng; 10 µl EXD fusion protein is approximately 7.5µg. All experiments were done with 1 µg of poly dI/dC non-specific competitorDNA except for the EXD binding experiment, where 0.02 µg was used. In theexperiment with less competitor, UBX binding looks similar to UBX binding inthe presence of 1 µg competitor (compare lanes 2 of the experiments shown in D).
3613dpp regulation by homeotic genes
were reduced (data not shown), although, again, it is imposs-ible to completely rule out lowered expression due to positioneffects. The level of dpp enhancer expression in both thegastric caeca and PS7 regions therefore appears to bedependent upon the integrity of the EXD binding site, inkeeping with loss of dpp expression and dpp enhancerexpression in those locations in exd mutants (Rauskolb andWieschaus, 1994).
DISCUSSION
The dpp gene performs diverse roles in development and isexpressed in many tissues and at several developmental stages.The structure of the dpp gene reflects its complex regulation.At least four promoters contribute to the overall pattern ofexpression of a single protein-coding sequence, and control-ling transcriptional enhancers are distributed over 50 kb. Twogroups (Hursh et al., 1993; Masucci and Hoffmann, 1993)
identified a region of the promoter needed to activate dpp tran-scription in the gastric caeca and central constriction of themidgut visceral mesoderm. Within that region we have identi-fied a 419 bp sequence sufficient to direct appropriate lacZexpression in the midgut mesoderm.
Characteristics of homeotic protein binding sites inthe dpp promoterFew direct target sequences of homeotic proteins have beencharacterized. Examples of such targets include Distal-less(Dll) as well as the clustered homeotic genes Deformed (Dfd)and Antp. The Dll and Antp genes are both repressed in theepidermis of the abdominal segments by Ubx and abd-A. Dfdprotein is required for expression of a Dfd autoregulatoryelement. Mutation of the homeotic binding sites identifiedwithin the control regions of these genes compromises theenhancer function. (Vachon et al., 1992; Appel and Sakonju,1993; Regulski et al., 1991; Zeng et al., 1994). Recently,Capovilla et al (1994) have identified a dpp midgut enhancer
A
B
C
D
E
F
dpp419 dpp261
419exd
261exd
dpp261
dpp∆DR
Fig. 7. Altered expression pattern of various enhancer mutants. Similarly staged embryos containing dpp419 (A) and other enhancer constructs(B-F) are shown stained for β-gal protein. Mutation of the EXD in vitro binding site in the context of dpp419 (B) or dpp261 (F) leads tocomplete loss or reduction, respectively, of gastric caeca expression (arrows). The embryo shown in B contained multiple 419exd inserts andthus produces high levels of β-gal in PS7. It was chosen to demonstrate the complete loss of gastric caeca expression. Mutation of the directrepeat in the context of dpp419 (C) reduces expression in the visceral mesoderm. Deletion of 3′ sequences from the 419 bp enhancer (dpp261)results in derepression of the β-gal pattern in the visceral mesoderm (D,E). dpp261 is derepressed posterior and anterior to the PS7 domain aswell as posterior to the GC domain. dpp261 is expressed in the hindgut endoderm (seen in D), whereas dpp419 is not. Derepression posterior tothe central constriction is clearly demonstrated in E. The β-gal expression posterior to the central constriction is weaker than in the normal PS7domain.

3614
that overlaps with the enhancer described in this report. Theauthors expressed a mutant form of UBX in flies that harboran altered form of the dpp enhancer fused to a lacZ reportergene. The mutant UBX is capable of recognizing the alteredenhancer in vitro. Based on the observed expression pattern oflacZ in the midgut, they argued for a direct interaction betweenUBX and the enhancer. Although this strategy has been usedin a few cases (e.g. Schier and Gehring, 1992), and can providestrong evidence for direct interaction, the altered UBX proteincould recognize the altered site even if there is normally anintermediary between UBX and the dpp enhancer. A strongercase for direct interaction would come from altering one of thecomponents, e.g. the protein sequence, and screening for com-pensatory mutations in the enhancer. In this way an interme-diary could be identified.
Most homeotic protein binding sites studied to date,including those identified in the Dll and Antp enhancers,contain TAAT sequences (reviewed by Laughon, 1991).TAAT serves as a core part of the recognition sequence in thecrystal structures of the engrailed and MATα2 homeodomainsbound to DNA (Kissinger et al., 1990; Wolberger et al., 1991).TAAT alone does not define a homeotic protein binding site,as strong preferences for two nucleotides just 3′ to the TAAT
have been found. The favored nucleotides 3′ to the TAAT forthe UBX homeodomain include GG, TA, AG, and TG in orderof decreasing preference (Ekker et al., 1991), and CC for thequite different bicoid homeodomain (Driever and Nüsslein-Volhard, 1989). The binding sites for UBX and ABD-A in thedpp enhancer (Table 1) are largely consistent with these invitro binding studies. The two nucleotides 3′ to the TAAT ineach dpp binding site are GG or TG, while the block 5 site thatis not bound in vitro contains the sequence TAATCG. Onehigh affinity site (I) contains the TAATGG motif whereasanother (F) contains the motif TAATTG. The medium affinitysite E also contains the sequence TAATTG, so this motif aloneis clearly not sufficient for high affinity DNA binding in vitro.
The sequence conservation between the D. melanogasterand D. virilis dpp midgut enhancers suggests that other factorsnecessary for enhancer function may bind adjacent to thehomeotic proteins. The regions of sequence conservation canextend over 30 bp in length, the conserved blocks generallyexceeding the extent of the DNaseI footprints of the home-odomain proteins. Each homeodomain binding site is flankedby conserved sequence which does not contain a TAAT and isnot bound by the homeotic proteins. We have shown that onesuch element, the direct repeat motif located between sites E
J. R. Manak, L. D. Mathies and M. P. Scott
A B
dpp45
WT HS-Ubx
10/11/129DR85/6/7
UBX
ABD-A andAnteriorRepressor(s)
+ +
C
1 419
dpp261
dpp419
VM Activator
+ ?
ABD-AGC Activator EXD?
dpp45
dpp45
Fig. 8. Expression pattern of dpp45. The dpp45 transgene is expressed in PS7 in a largely tissue-specific manner. A shows dpp45 expression inPS7 (closed arrow) as well as staining in transverse muscles of abdominal segments 1 to 3 (open arrows). B shows expanded dpp45 expressionin the presence of ubiquitously expressed UBX. The embryo in B was slightly enlarged photographically to facilitate comparison of panels Aand B. The model presented in C summarizes our current knowledge of regulation through dpp419. In the model ABD-A represses dpp419through at least two sites located within blocks 9 and 11, based on the loss of responsiveness of dpp261 to ABD-A and the sensitivity of dpp45to it. Other anterior repressors act through blocks 10 and 11. UBX activates dpp419 through at least block 9 and perhaps through other blocks.A general visceral mesoderm activator binds the direct repeat. Either a general or gastric caeca-specific activator, which may or may not beEXD, acts through block 8. DR, direct repeat. Numbers below boxes indicate homology blocks. Numbers at ends indicate enhancercoordinates. Labels to the left indicate various constructs that were tested in this study. Brackets indicate boundaries of constructs.

3615dpp regulation by homeotic genes
and F (indicated in Fig. 9), does not bind either UBX or ABD-A, and yet is required for high-level expression driven by theenhancer. Identification of the proteins that bind this and otherelements will lead to a better understanding of how homeotic-responsive enhancers function. Not every conserved elementneed serve a function in determining the specificity of actionof homeotic proteins. In its place within the array of enhancersin the dpp gene, the 419 bp element may serve multiple rolesin addition to its role in visceral mesoderm expression. Someconserved elements may serve no regulatory function.
Specificity of homeotic protein actionANTP and UBX both activate the dpp enhancer in the anteriormidgut mesoderm, while SCR has no effect. Even thoughANTP does not normally activate the dpp enhancer, perhapsthe high level in the heat shock experiment causes it to havethis abnormal activity. It may, for example, bind toUBX/ABD-A binding sites from which it is normallyexcluded. ANTP and UBX differ by 7 amino acids in thehomeodomain, while ANTP and SCR differ by only fouramino acids near the N-terminal part of the homeodomain.These differences are, however, crucial to the proteins’ specificactions (Kuziora and McGinnis, 1989; Gibson et al., 1990;Chan and Mann, 1993; Zeng et al., 1993). The distinct responseto ANTP and UBX versus SCR suggests that binding of SCRin vivo may be ineffective. The binding affinity and/or speci-ficity of ANTP and UBX could be increased by interactionwith other proteins. One candidate is the product of the exdgene, which is required for dpp expression in PS7 and interactsgenetically with Antp and Ubx, but not Scr. Alternatively SCRmay bind to the dpp enhancer but have no effect once bound.In the cuticle SCR is blocked from regulating denticle pat-terning by UBX and ABD-A, but in the induction of salivaryglands the homeotic gene teashirt is the relevant limitinginfluence (Andrew et al., 1994). The influences limiting SCRfunction in the mesoderm have not been identified.
Repression versus activationRemoval of homology blocks 11 and 12 causes a dramaticderepression of lacZ expression, especially posteriorly.However, Capovilla et al. (1994) report that site-directedmutation of the TAAT region of block 11 (their site 1) has onlya moderate effect on expression. It is unlikely that the removalof block 12 has any effect on expression, because the 303 bp
fragment they have characterized does not contain block 12 yetdrives normal expression. Therefore it is likely that removal ofblock 11 in its entirety causes the dramatic derepression. Thesite-directed mutation of homology block 11 may have beeninsufficient to fully compromise its function.
The derepression observed with dpp261 suggests that acti-vation in the visceral mesoderm is the default state, with avariety of repressors restricting expression. This view is furthersupported by residual dpp expression seen in mutants com-pletely lacking Ubx function (Reuter et al., 1990). Ubx seemsnecessary to augment dpp expression, not to initiate it. Theregulator(s) which activates dpp in PS7 in the absence of Ubxmay also facilitate the activation of dpp by UBX.
ABD-A represses dpp while UBX and ANTP activate dpp,but the in vitro DNA binding data do not explain the differentactivities of the UBX versus ABD-A proteins. The enhancer isbound by UBX and ABD-A proteins at the same sites in vitro,with our preparation of UBX protein binding more stronglythan ABD-A. How then does binding of the proteins producedifferent actions? The pattern of expression of dpp261 suggeststhat this deletion construct is no longer efficiently repressed byabd-A in the posterior, or by different factors in the anteriormidgut and hindgut. The single strong homeodomain bindingsite which distinguishes dpp419 from dpp261 appears to be amajor site of repression acted upon by a variety of proteins,including ABD-A (Fig. 7D,E). dpp261 is expressed morestrongly in PS7 than in PS8 and therefore is sufficient torespond to UBX. However, the homeotic binding site removedto create dpp261 might also be used by UBX in vivo. Similarly,the residual repression of dpp261 in the posterior midgut (thederepression of dpp in abd-A mutants exceeds the derepressionseen with dpp261) could be due to weak effects of abd-A onremaining binding sites.
The difference between repression and activation could bedetermined by the interaction of UBX and ABD-A proteinswith distinct cofactors. Ubiquitous cofactors could differen-tially interact with either UBX or ABD-A, or a cofactor proteincapable of interacting with only one homeotic protein may bemade only in certain cells. A necessary activator protein mightbe absent from PS8, preventing ABD-A from activating dpp,or the factor might be present in PS8 but unable to act withABD-A. Ubiquitous expression of ABD-A prevents dppexpression rather than permitting it, so ABD-A appears unableto cooperate with a PS7 factor to activate dpp. It is also possiblethat UBX or ABD-A could exert their effects by modulatingregulation by a third as yet unidentified regulator. Finally,UBX and ABD-A could interact with the same cofactor buthave different effects once bound.
45 bp of the dpp midgut enhancer defines ahomeotic response element (HOMRE)We have identified an element 45 bp in length that responds toboth UBX and ABD-A homeotic proteins. The response ofonly 45 bp of dpp enhancer to these proteins, with tissue speci-ficity largely preserved, suggests that most factors necessaryfor promoting appropriate visceral mesoderm expression canrecognize 45 bp, or less, of DNA. It is now possible to dissectbase by base this homeotic response element to help under-stand how homeotic protein specificity is achieved. There aretwo TAAT motifs in dpp45. Both ABD-A and UBX may bind
Table 1. Comparison of homeodomain binding sitesequences with TAAT motifs within the 813 bp fragment
Site I (site 1)
AAAACTTAATGGCCACCATAACSite F (site 2) TATTACTAATTGGGTGTGAATTSite E (site 3) GGGTGCTAATTGATAGGCCTTTSite F-rev AATTAGTAATAAATTTGAATGCSite E-3′ GCAGCATAATCGAAATGGGTGCBlock 3 TGACAGTAATGCGACCATTACGBlock 4 CGGTGCTAATGATGATTTTGATBlock14-5′ CGAACTTAATTGCAATTGACTTBlock 14-3′ CTGTGTTAATTCGAATTC
Sites E,F, and I are bound by both UBX and ABD-A. Blocks 3, 4, 14-5′and 14-3′ (which correlate with blocks shown in Fig. 5B) are TAAT motifs inconserved blocks outside of the 419 bp fragment which were not tested forhomeotic protein binding. Numbers in parentheses correspond to TAATregions of in vitro homeotic binding sites as described in Capovilla et al.(1994).

3616
to the same site, or the two proteins may each have a preferredsite of action.
Why is dpp261, which contains the 45bp of dpp45 sequence,posteriorly derepressed while the dpp45 construct is not?Perhaps the ratio between a general visceralmesoderm activator and homeotic repressor(ABD-A) determines whether expression doesor does not occur posteriorly. In the case ofdpp261, one of the few high affinity ABD-Abinding sites has been removed and activationpredominates. In the case of the dpp45construct, ABD-A need only overcome thegeneral activation provided by one or fewvisceral mesoderm activators, assuming thatfew proteins can bind 45 bp of DNA, so repres-sion predominates. TGCATGCA-like motifs,which occur abundantly within the 419 basesof DNA (Fig. 9) and could bind a visceralmesoderm activator, are plentiful within the261 bp fragment but occur only once in dpp45.
The results reported here and by Capovilla etal. (1994) support the existence of tissue-specificinformation for dpp expression tightly linked tohomeotic protein responsiveness. The low levelresidual expression of dpp in PS7 in Ubx nullmutants (Reuter et al., 1990) may be due to theactivators whose existence we infer. Ubx activityin PS7 raises the level of expression, whileunknown repressors in the anterior and abd-A inthe posterior lower dpp expression. In an evolu-tionary context, this refinement of a tissue-specific expression pattern into a more elaboratespatial pattern may reflect acquisition ofHOMRE’s by the dpp gene.
It is interesting to note that dpp45 does notcontain an in vitro EXD binding site nor doesit contain a sequence that resembles theATCAATCA motif (see below). Although it ispossible that EXD does interact with dpp45, itis also possible that other as yet unidentifiedfactors are interacting with UBX and ABD-A.Whether such factors also impart tissue speci-ficity remains to be determined.
Binding by extradenticle proteinexd has long been the best candidate to encodea homeotic protein cofactor. Careful analysesby Peifer and Wieschaus (1990) demonstratedthat although homeotic transformations occurin exd mutants, there is no detectable changein the expression patterns of the clustered HOXclass homeotic genes. The findings are in clearcontrast to the effects of repressors and activa-tors of the Polycomb (Paro, 1993) and trithoraxgroups (Kennison, 1993), where changes inhomeotic gene expression account for thehomeotic transformations seen in the mutants.Peifer and Wieschaus (1990) further substanti-ated the role of exd in homeotic proteinfunction by showing how exd alters the effectsof ectopically expressed homeotic proteins.
They concluded that exd is likely to encode either a geneinvolved in carrying out the orders given by homeotic proteins,a target gene, or a cofactor that helps to control the specificityof target gene recognition or regulation.
J. R. Manak, L. D. Mathies and M. P. Scott
8
5
11
Sequences identical in D. melanogaster and D. virilis
Dyad symmetric direct repeats EXD
G
12
10
A
EXD
E
C D
7
6
419
1 (=262 in Fig. 5)
Underlined: TGCA repeats
9
Boxed: TAAT and ATTA sequences
60
110
142
199
386
(= 680 in Fig.5)
UBX and ABD-A DNase I footprintsStrongMediumWeak
271
329
EXD DNase I footprint
B
F
I
=Interrupted homology
250
H
45mer
8mer repeats
CTGGGAGCCAATTTGATTTGTGGCCCACTCGAG.........................
TCCCCTACCCCCGATCTTTCCTTGCCCCATGCCCAGATTTCAATTCGATTCCCGGAT.
ATTCGGCTCAACAGTTATGGTGGCCATTAAGTTTTATCGATGGCGCTACAGCTCCCGA
TGCGCGCTGCAATTTGCCGCC....................................
TGCAGCACTTGCACTGCCTGCAATTCACACCCAATTAGTAATAAATTTGAA.......
CCGCAGATCAAAGGCCTATCAATTAGCACCCATTTCGATTATGCTGCATGCTGCATA
GTTGTTTTACAATCGCTGTAAATAAATAGGAG..........................
GATTTCCCCATGCCCATTTGGCCGTGCAATGTTTGTTTTATGCACGATCC........
CTGCAGACCCACGAAGACCCGGCGATTACGTGGAGTACTACCCATTTGGCTTCCCATTTC
Fig. 9. Location of binding sites for the UBX, ABD-A, and EXD proteins on the D.melanogaster enhancer. The locations of binding sites for homeotic proteins on the 419bp midgut enhancer are shown in comparison to the pattern of evolutionarily conservedsequences. The sequence is shown in its 5′ to 3′ configuration as it is arranged withrespect to the dpp promoter in the chromosome. The enhancer is numbered from 1 to419, with position 1 corresponding to position 262 in Fig. 5B. Gray shading indicatessequences conserved between D. melanogaster and D. virilis., with the boxed numberscorresponding to the block numbers in Fig. 5. The 261 bp version of the enhancer hasthe same 5′ end as the 419 bp, but ends at the small bracket within conserved block 10.In addition to the 256 bp indicated, the method of building the fragment left anadditional ‘TCGAG’ at the 3′ end for a total of 261 bp. The large brackets demarcatethe 45 bp fragment capable of responding to UBX and ABD-A. Notable features of thesequence are identified by boxes (TAAT sequences which form the core of manyhomeodomain binding sites), underlining (TGCA repeats which are surprisinglyabundant in the sequence), arrows (a direct repeat with dyad symmetrical coreelements), or by horizontal brackets (TGCATGCA-like motifs). The positions of strong,medium, and weak binding sites for UBX and ABD-A proteins are indicated by barsbelow the sequence. The EXD binding site is indicated. The vertical bars indicate D.melanogaster sequence blocks that are conserved but interrupted in D. virilis.

3617dpp regulation by homeotic genes
The exd protein is a homeodomain protein most closelyrelated to the yeast MAT product a1 (Rauskolb et al., 1993),a cofactor for the MATα2 homeodomain protein in repressinghaploid-specific target genes (Johnson, 1992). Although theUBX homeodomain does not look particularly similar toMATα2, the similarity of EXD to MATa1 makes its role as acofactor plausible. Rauskolb and Wieschaus (1994) testedtarget genes known to be regulated by homeotic proteins,including dpp in the midgut mesoderm, to see whether theirexpression patterns are dependent on exd, and found that theyare. Thus the homeotic gene expression patterns are preservedin exd mutants but the target gene expression patterns are not.They found a requirement for EXD in activating dppexpression in PS7.
EXD is closely related to a mammalian protein, PBX1,implicated in human leukemias by chromosome rearrange-ments causing a chimeric PBX1/E2A protein to be produced(Kamps et al., 1990; Nourse et al., 1990). PBX1 and EXD areextremely similar within the homeodomain, and quite similarthroughout the rest of the proteins (Flegel et al., 1993;Rauskolb et al., 1993). A cDNA encoding the hybrid proteinintroduced into mice causes lymphomas (Dedera et al., 1993).One possible interpretation of these observations is that thehomeodomain or other parts of PBX1 targets the chimericprotein to genes normally regulated by PBX1. Because thehybrid protein causes loss of growth control in blood cells,normal targets of PBX may include growth regulating genes.Whatever the exact mechanism of leukemogenesis, the highdegree of similarity between PBX and EXD suggests PBX mayinteract with HOX complex products. HOX genes are activeduring hematopoiesis (e.g. Magli et al., 1991) and PBX maybe active there as well.
exd is a good candidate for a direct regulator of the dppenhancer. The EXD binding site in the dpp enhancer containsa motif (ATCAATTAG) strikingly similar to the PBX1consensus sequence (ATCAATCA[A]) identified by twogroups of workers (van Djik et al., 1993; LeBrun and Cleary,1994). This motif is absolutely conserved in both fly speciesexamined, and EXD protein recognizes this site in vitro.
The apparent reduction of expression in both the gastriccaeca and in PS7 of the visceral mesoderm upon mutation ofthe EXD binding site is consistent with reduction of dpp anddpp enhancer/lacZ expression from both gastric caeca and PS7in exd mutants (Rauskolb and Wieschaus, 1994). However,loss of expression in the gastric caeca is clearly more dramaticthan loss in PS7. This could be due to additional HOMREs inthe 419 bp enhancer that can act in the absence of an EXD sitein PS7 but not in the caeca. Thus, targeting an EXD siteinvolved in both pathways of regulation might have a greatereffect on gastric caeca expression. Two alternative hypothesesare also possible. First, the site we have mutated could be acontrol element that may or may not bind EXD in vivo but isexclusively required for gastric caeca expression. Second, thesite could be a control element that is not bound by EXD invivo but is nonetheless necessary for both gastric caeca andPS7 expression.
Our EXD binding results differ from those of Chan et al.(1994). We have not observed EXD binding to the regionwhich they call dpp80, even though we are able to observe aclear footprint at the ATCAATTA motif. The dpp80 region ofthe enhancer is relatively poorly conserved between fly
species, save for the motif GTTTGTTTTAT (Fig. 9).Moreover, the cooperative binding observed by Chan et al.(1994) involved a several hundred- fold molar excess of EXDover UBX. The results of van Dijk and Murre (1994) show thata region of EXD amino terminal to the homeodomain isabsolutely required for cooperative binding with UBX. Thisdomain is absent from the EXD polypeptide used by Chan etal. (1994). Thus, a number of differences in experimentaldesign may explain the different findings reported in the threepapers.
We especially want to thank James Masucci, F. Michael Hoffmann,Cordelia Rauskolb, and Eric Wieschaus for communicating resultsprior to their publication. George Chen and Chen-sen Wu providedvaluable help with library screens, injections and other experiments.We thank Drs Brad Johnson, Mark Krasnow, Bruce Appel, and ShigeSakonju for gifts of protein and advice on purification, Dr VolkerHartenstein for helping identify muscle cells, Drs Ernesto Sanchez-Herrero and Gines Morata for the heat shock-inducible abd-A flies,the Cell Sciences Imaging Facility for assistance with confocalimaging, and the Indiana University Drosophila Stock Center forproviding many fly stocks. We thank Bruce Appel, Michelle Lamka,Shige Sakonju, James Masucci, F. Michael Hoffmann, CordeliaRauskolb, John Tamkun, and Eric Wieschaus for various plasmids,libraries, and antibodies. Finally, we thank the anonymous reviewersfor careful reviews and thoughtful suggestions for improvements.This work was supported by N.I.H. grant no. HD18163. M. P. S. isan investigator of the Howard Hughes Medical Institute.
REFERENCES
Affolter, M., A. Percival-Smith, M. Müller, W. Leupin, and W. J. Gehring(1990). DNA binding properties of the purified Antennapedia homeodomain.Proc. Nat. Acad. Sci. USA 87, 4093-4097.
Andrew, D. J., M. A. Horner, M. G. Petitt, S. M. Smolik and Scott, M. P.(1994). Setting limits on homeotic gene function: Restraint of Sex combsreduced activity by teashirt and other homeotic genes. EMBO J. in press.
Andrew, D. J. and M. P. Scott (1992). Downstream of the homeotic genes.New Biologist 4, 5-15.
Appel, B. and S. Sakonju (1993). Cell-type-specific mechanisms oftranscriptional repression by the homeotic gene products UBX and ABD-Ain Drosophila embryos. EMBO J. 12, 1099-1109.
Beachy, P. A., M. A. Krasnow, E. R. Gavis, and D. S. Hogness (1988). AnUltrabithorax protein binds sequences near its own and the Antennapedia P1promoters. Cell 55, 1069-1081.
Beverley, S. M. and A. C. Wilson (1984). Molecular Evolution in Drosphilaand the higher Diptera. II. A time scale for fly evolution. J. Mol. Evol. 21, 1-13.
Bienz, M. and G. Tremml (1988). Domain of Ultrabithorax expression inDrosophila visceral mesoderm from autoregulation and exclusion. Nature333, 576-578.
Botas, J. (1993). Control of morphogenesis and differentiation by HOM/Hoxgenes. Curr. Opin. Cell Biol. 5, 1015-1022.
Capovilla, M., M. Brandt, and J. Botas. (1994). Direct regulation ofdecapentaplegic by Ultrabithorax and its role in Drosophila midgutmorphogenesis. Cell 76, 461-475.
Chan, S-K., L. Jaffe, M. Capovilla, J. Botas and R.S. Mann (1994). TheDNA binding specificity of Ultrabithorax is modulated by cooperativeinteractions with Extradenticle, another homeoprotein. Cell 78, 603-615.
Chan, S. K. and R. S. Mann (1993). The segment identity functions ofUltrabithorax are contained within its homeo domain and carboxy-terminalsequences. Genes Dev 7, 796-811.
Dedera, D. A., E. K. Waller, D. P. LeBrun, A. Sen-Majumdar, M. A.Stevens, G. S. Barsh, and M. L. Cleary (1993). Chimeric homeobox geneE2A-PBX1 induces proliferation, apoptosis, and malignant lymphomas intransgenic mice. Cell 74, 833-843.
Driever, W. and C. Nüsslein-Volhard (1989). The bicoid protein is a positiveregulator of hunchback transcription in the early Drosophila embryo. Nature337, 138-143.

3618
Duboule, D. and P. Dolle (1989). The structural and functional organization ofthe murine HOX gene family resembles that of Drosophila homeotic genes.EMBO J. 8, 1497-1505.
Ekker, S. C., K. E. Young, D. P. Von Kessler, and P. A. Beachy (1991).Optimal DNA sequence recognition by the Ultrabithorax homeodomain ofDrosophila. EMBO J. 10, 1179-1186.
Flegel, W. A., A. W. Singson, J. S. Margolis, A. G. Bang, J. W. Posakony,and C. Murre (1993). Dpbx, a new homeobox gene closely related to thehuman proto-oncogene pbx1. Molecular structure and expression. Mech.Dev. 41, 155-161.
Fortini, M. E. and G. M. Rubin (1990). Analysis of cis-acting requirements ofthe Rh3 and Rh4 genes reveals a bipartite organization to rhodopsinpromoters in Drosophila melanogaster. Genes Dev. 4, 444-63.
Gehring, W. J., M. Müller, M. Affolter, S. A. Percival, M. Billeter, Y. Q.Qian, G. Otting, and K. Wüthrich (1990). The structure of thehomeodomain and its functional implications. Trends Genet. 6, 323-329.
Gibson, G., A. Schier, P. LeMotte, and W. J. Gehring (1990). Thespecificities of Sex combs reduced and Antennapedia are defined by a distinctportion of each protein that includes the homeodomain. Cell 62, 1087-1103.
Graham, A., N. Papalopulu, and R. Krumlauf (1989). The murine andDrosophila homeobox gene complexes have common features oforganization and expression. Cell 57, 367-378.
Hayashi, S. and M. P. Scott (1990). What determines the specificity of actionof Drosophila homeodomain proteins? Cell 63, 883-894.
Hoey, T., R. Warrior, J. Manak, and M. Levine (1988). DNA-bindingactivities of the Drosophila melanogaster even skipped protein are mediatedby its homeodomain and influenced by protein context. Mol. Cell. Biol. 8,4598-4607.
Hursh, D. A., R. W. Padgett, and W. M. Gelbart (1993). Cross regulation ofdecapentaplegic and Ultrabithorax transcription in the embryonic visceralmesoderm of Drosophila. Development 117, 1211-1222.
Immerglück, K., P. A. Lawrence, and M. Bienz (1990). Induction acrossgerm layers in Drosophila mediated by a genetic cascade. Cell 62, 261-268.
Johnson, A. (1992). A combinatorial regulatory circuit in budding yeast. InTranscriptional Regulation, 2, (ed. McKnight, S. L. and K. R. Yamamoto) pp.975-1006. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Johnson, F. B. and M. A. Krasnow (1990). Stimulation of transcription by anUltrabithorax protein in vitro. Genes Dev. 4, 1044-1052.
Kadonaga, J. T. (1991). Purification of sequence-specific binding proteins byDNA affinity chromatography. In Methods in Enzymology 208, pp. 10-23.San Diego: Academic Press, Inc.
Kamps, M. P., C. Murre, X.-h. Sun, and D. Baltimore (1990). A newhomeobox gene contributes the DNA binding domain of the t(1;19)translocation protein in pre-B ALL. Cell 60, 547-555.
Kassis, J. A., S. J. Poole, D. K. Wright, and P. H. O’Farrell (1986). Sequenceconservation in the protein coding and intron regions of the engrailedtranscription unit. EMBO J 5, 3583-3589.
Kennison, J. A. (1993). Transcriptional activation of Drosophila homeoticgenes from distant regulatory elements. Trends Genet 9, 75-79.
Kissinger, C. R., B. Liu, B. E. Martin, T. B. Kornberg, and C. O. Pabo(1990). Crystal structure of an engrailed homeodomain-DNA complex at2.8 Å resolution: A framework for understanding homeodomain-DNAinteractions. Cell 63, 579-590.
Kuziora, M. A. and W. McGinnis (1989). A homeodomain substitutionchanges the regulatory specificity of the Deformed protein in Drosophilaembryo. Cell 59, 563-571.
Laughon, A. (1991). DNA binding specificity of homeodomains. Biochemistry30, 11357-11367.
LeBrun, D. P. and M. L. Cleary (1994). Fusion with E2A alters thetranscriptional properties of the homeodomain protein Pbx1 in t(1;19)leukemias. Oncogene 9, 1641-1647.
Lin, L. and W. McGinnis (1992). Mapping functional specificity in the Dfdand Ubx homeo domains. Genes Dev 6, 1071-1081.
Magli, M. C., P. Barba, A. Celetti, G. De Vita, C. Cillo, and E. Boncinelli(1991). Coordinate regulation of HOX genes in human hematopoietic cells.Proc. Natl. Acad. Sci. USA 88, 6348-52.
Maier, D., A. Preiss, and J. R. Powell (1990). Regulation of the segmentationgene fushi tarazu has been functionally conserved in Drosophila. EMBO J. 9,3957-3966.
Mann, R. S. and D. S. Hogness (1990). Functional dissection of Ultrabithoraxproteins in D. melanogaster. Cell 60, 597-610.
Masucci, J. D. and F. M. Hoffmann (1993). Identification of two regions fromthe Drosophila decapentaplegic gene required for embryonic midgutdevelopment and larval viability. Dev. Biol. 159, 276-287.
Mathies, L.D., S. Kerridge, and M.P. Scott (1994). Roles of the teashirt genein Drosophila midgut morphogenesis: Secreted proteins mediate thecombinatorial action of homeotic genes. Development (in press).
Mavilio, F. (1993). Regulation of vertebrate homeobox-containing genes bymorphogens. Eur. J. Biochem. 212, 273-288.
Maxam, A. M. and W. Gilbert (1980). Sequencing end-labeled DNA withbase-specific chemical cleavages. Methods Enzymol. 65, 499-560.
McGinnis, W. and R. Krumlauf (1992). Homeobox genes and axialpatterning. Cell 68, 283-302.
Morata, G. (1993). Homeotic genes of Drosophila. Curr. Opin. Genet. Dev. 3,606-614.
Müller, J., F. Thüringer, M. Biggin, B. Zust, and M. Bienz (1989).Coordinate action of a proximal homeoprotein binding site and a distalsequence confers the Ultrabithorax expression pattern in the visceralmesoderm. EMBO J. 8, 4143-4151.
Nourse, J., J. D. Mellentin, N. Galili, J. Wilkinson, E. Stanbridge, S. D.Smith, and M. L. Cleary (1990). Chromosomal translocation t(1;19) resultsin the synthesis of a homeobox fusion mRNA that codes for a potentialchimeric transcription factor. Cell 60, 535-545.
Panganiban, G. E. F., K. E. Rashka, M. D. Neitzel, and F. M. Hoffmann(1990a). Biochemical characterization of the Drosophila dpp protein, amember of the transforming growth factor-β family of growth factors. Mol.Cell Biol. 10, 2669-2677.
Panganiban, G. E. F., R. Reuter, M. P. Scott, and F. M. Hoffmann (1990b).A Drosophila growth factor homolog, decapentaplegic, regulates homeoticgene expression within and across germ layers during midgutmorphogenesis. Development 110, 1041-1050.
Paro, R. (1993). Mechanisms of heritable gene repression during developmentof Drosophila. Curr. Opin. Cell Biol. 5, 999-1005.
Peifer, M., C. Rauskolb, M. Williams, B. Riggleman, and E. Wieschaus(1991). The segment polarity gene armadillo interacts with the winglesssignaling pathway in both embryonic and adult pattern formation.Development 111, 1029-1043.
Peifer, M. and E. Wieschaus (1990). Mutations in the Drosophila geneextradenticle affect the way specific homeo domain proteins regulatesegmental identity. Genes Dev 4, 1209-1223.
Rauskolb, C., M. Peifer, and E. Wieschaus (1993). extradenticle, a regulatorof homeotic gene activity, is a homolog of the homeobox-containing humanproto-oncogene pbx1. Cell 74, 1011-1112.
Rauskolb, C. and E. Wieschaus (1994). Coordinate regulation of downstreamgenes by extradenticle and the homeotic selector proteins. EMBO J. 13,3561-3569.
Regulski, M., S. Dessain, N. McGinnis, and W. McGinnis (1991). High-affinity binding sites for the Deformed protein are required for the function ofan autoregulatory enhancer of the Deformed gene. Genes Dev 5, 278-286.
Reuter, R., G. E. F. Panganiban, F. M. Hoffmann, and M. P. Scott (1990).Homeotic genes regulate the spatial expression of putative growth factors inthe visceral mesoderm of Drosophila embryos. Development 110, 1031-1040.
Reuter, R. and M. P. Scott (1990). Expression and functions of the homoeoticgenes Antennapedia and Sex combs reduced in the embryonic midgut ofDrosophila. Development 109, 289-303.
Rubin, G. M. and A. C. Spradling (1982). Genetic transformation ofDrosophila with transposable element vectors. Science 218, 348-353.
Schier, A. F. and W. J. Gehring (1992). Direct homeodomain-DNAinteraction in the autoregulation of the fushi tarazu gene. Nature 356, 804-807.
Scott, M. P., J. W. Tamkun, and G. W. Hartzell III (1989). The structure andfunction of the homeodomain. BBA Rev. Cancer 989, 25-48.
St. Johnston, R. D. and W. M. Gelbart (1987). Decapentaplegic transcriptsare localized along the dorsal-ventral axis of the Drosophila embryo. EMBOJ. 6, 2785-91.
Tremml, G. and M. Bienz (1989). Homeotic gene expression in the visceralmesoderm of Drosophila embryos. EMBO J. 8, 2677-2685.
Tremml, G. and M. Bienz (1992). Induction of labial expression in theDrosophila endoderm: Response elements for dpp signalling and forautoregulation. Development 116, 447-456.
Vachon, G., B. Cohen, C. Pfeifle, M. E. McGuffin, J. Botas, and S. M.Cohen (1992). Homeotic genes of the bithorax complex repress limbdevelopment in the abdomen of the Drosophila embryo through the targetgene Distal-less. Cell 71, 437-450.
van Dijk, M.A. and C. Murre (1994). extradenticle raises the DNA bindingspecificity of homeotic selector gene products. Cell 78, 617-624.
van Dijk, M. A., P. M. Voorhoeve, and C. Murre (1993). Pbx1 is converted
J. R. Manak, L. D. Mathies and M. P. Scott

3619dpp regulation by homeotic genes
into a transcriptional activator upon acquiring the N-terminal region of E2Ain pre-B-cell acute lymphoblastoid leukemia. Proc Natl Acad Sci USA 90,6061-6065.
Wang, B. B., M. M. Müller-Immergluck, J. Austin, N. T. Robinson, A.Chisholm, and C. Kenyon (1993). A homeotic gene cluster patterns theanteroposterior body axis of C. elegans. Cell 74, 29-42.
Wolberger, C., A. K. Vershon, B. Liu, A. D. Johnson, and C. O. Pabo(1991). Crystal structure of a MATa2 homeodomain-operator complexsuggests a general model for homeodomain-DNA interactions. Cell 67, 517-528.
Zeng, W., D. J. Andrew, L. D. Mathies, M. A. Horner, and M. P. Scott(1993). Ectopic expression and function of the Antp and Scr homeotic genes:The N terminus of the homeodomain is critical to functional specificity.Development 118, 339-352.
Zeng, C., J. Pinsonneault, G. Gellon, N. McGinnis, and W. McGinnis(1994). Deformed protein binding sites and cofactor binding sites arerequired for the function of a small segment-specific regulatory element inDrosophila embryos. EMBO J. 13, 2362-2377.
(Accepted 23 September 1994)





























