Embed Size (px)
Citation preview

Ti
th
Developmental Biology 211, 323–334 (1999)Article ID dbio.1999.9311, available online at http://www.idealibrary.com on
Overexpression of ptc1 Inhibits Induction of Shharget Genes and Prevents Normal Patterning
n the Neural Tube
Lisa V. Goodrich, David Jung, Kay M. Higgins, and Matthew P. Scott1
Department of Developmental Biology and Department of Genetics, Howard Hughes MedicalInstitute, Stanford University School of Medicine, Stanford, California 94305-5427
Patched (Ptc) is a human tumor suppressor protein and a candidate receptor for Hedgehog (Hh) proteins, which regulategrowth and patterning in embryos. Ptc represses expression of Hh target genes such as Gli1 and ptc1 itself. Localizedsecretion of Hh appears to induce transcription of target genes in specific patterns by binding to Ptc and preventing it fromfunctioning in recipient cells. People who are heterozygous for PTC1 exhibit a range of developmental defects, suggestingthat some genes are inappropriately expressed when there is not enough Ptc protein. To test the idea that a balance betweenHh and Ptc activities is essential for normal development, we overexpressed Ptc in the neural tube. We find that excess Ptcis sufficient to inhibit expression of Gli1 and ptc1, suggesting that Sonic hedgehog (Shh) cannot signal effectively. This leadso partial dorsalization of the neural tube and a wide spectrum of neural defects, ranging from embryonic lethality toydrocephaly. © 1999 Academic Press
Key Words: patched; hedgehog; segment polarity; induction; vertebrate development; signaling; neural development.
INTRODUCTION
The Hedgehog (Hh) pathway is a conserved signalingsystem that plays an important role in cell–cell communi-cation during development and growth. Hh proteins areautocatalytically cleaved ligands (Lee et al., 1994; Bumcrotet al., 1995; Porter et al., 1996a,b) that can bind to thetransmembrane protein Patched (Ptc) (Marigo et al., 1996a;Stone et al., 1996) and induce transcription of target genesin the recipient cell (Hammerschmidt et al., 1997). Invertebrates, Sonic hedgehog (Shh) secreted from axial me-soderm is believed to induce expression of the transcriptionfactor HNF-3b and differentiation of the floor plate in theadjacent neural tube (Echelard et al., 1993; Krauss et al.,1993; Roelink et al., 1994, 1995). Shh and the relatedproteins Indian hedgehog (Ihh) and Desert hedgehog (Dhh)also organize development of the skeleton (Fan and Tessier-Lavigne, 1994; Johnson et al., 1994; Vortkamp et al., 1996),limbs (Riddle et al., 1993), gut (Roberts et al., 1995), andgermline (Bitgood et al., 1996).
Despite the large number of biological activities attrib-
1 To whom correspondence should be addressed. Fax: 415-725-7739. E-mail: [email protected].
0012-1606/99 $30.00Copyright © 1999 by Academic PressAll rights of reproduction in any form reserved.
uted to Hh proteins, little is known about how Hh signalingaffects Ptc function and subsequently causes changes ingene transcription. Ptc has 12 potential transmembranedomains and a proposed sterol-sensing domain (Goodrich etal., 1996; Carstea et al., 1997; Loftus et al., 1997; Johnsonand Scott, 1998). It is related to the Niemann Pick Type Cdisease gene NPC1 (Carstea et al., 1997; Loftus et al., 1997)which regulates cholesterol transport and homeostasis, butno biochemical function other than Hedgehog binding hasbeen ascribed to Ptc.
One clear biological function of Ptc is the regulation ofHh target gene transcription. Ptc represses transcriptionof genes that are induced by Hh (Goodrich et al., 1997).Because Hh and Ptc have opposing activities, Hh proteinis proposed to bind and inactivate the Ptc protein (Marigoet al., 1996a; Stone et al., 1996). Although some Hh targetgenes vary, two are nearly always induced: the zinc fingertranscription factor Gli1 and ptc1 itself. ptc1 and Gli1 areexpressed in cells that receive Hh signals and are inducedby ectopic sources of Hh (Marigo et al., 1996b; Hynes etal., 1997; Lee et al., 1997; Platt et al., 1997; Sasaki et al.,1997). Furthermore, ptc1 expression decays in Shh and
Dhh mutant mice (Bitgood et al., 1996; Chiang et al.,1996). Conversely, in mouse ptc1 mutants, Gli1 and ptc1323

1HidtVtv
wadPd(c(pPsJR1asPdataoc
bnmA1iisePa
aemSntd
324 Goodrich et al.
are ectopically expressed, consistent with the idea thatPtc protein normally represses target gene transcriptionin cells that do not receive the Hh signal (Goodrich et al.,
997). Gli1 (Kinzler et al., 1987; Walterhouse et al., 1993;ui et al., 1994) is a vertebrate homolog of Cubitus
nterruptus (Ci), a transcription factor that appears to beirectly responsible for activation and repression of Hharget genes in flies (Forbes et al., 1993; Hooper, 1994;on Ohlen et al., 1997). Similarly, Gli1 can induce Hh
arget gene expression in mice (Hynes et al., 1997) and initro (Sasaki et al., 1997).Additional insight into biological roles for ptc1 cameith the discovery that germline PTC1 mutations are
ssociated with the human disease basal cell nevus syn-rome (BCNS) (Hahn et al., 1996; Johnson et al., 1996).eople with BCNS are large and suffer from a number ofevelopmental disorders such as polydactyly and bifid ribsGorlin, 1987). In addition, BCNS patients have an in-reased frequency of the skin tumor basal cell carcinomaBCC), the deadly brain tumor medulloblastoma, and otherroliferative disorders such as jaw cysts (Gorlin, 1987).TC1 mutations have also been found in a number ofporadic tumors (Gailani et al., 1996; Hahn et al., 1996;ohnson et al., 1996; Unden et al., 1996; Pietsch et al., 1997;affel et al., 1997; Vorechovsky et al., 1997; Wolter et al.,997; Xie et al., 1997). The developmental defects associ-ted with BCNS are believed to arise from PTC1 haploin-ufficiency while the tumors generally have both copies ofTC1 inactivated. Mice that are heterozygous for ptc1isplay some phenotypes reminiscent of the human diseasend are at increased risk for medulloblastoma and otherumors (Goodrich et al., 1997; Hahn et al., 1998). Inddition, homozygous ptc1 mutant mice die early in devel-pment with severe neural tube defects and an abnormalirculatory system.The defects observed in ptc1 mutants suggest that a
alance between Hh and Ptc activities is necessary forormal development. For instance, in BCNS patients thereay not be enough Ptc to keep Hh target genes off.lternatively, as has been observed in flies (Taylor et al.,993; Chen and Struhl, 1996), Hh ligand may diffuse farthern tissues with less Ptc protein. Also, in vitro experimentsn vertebrates suggest that Shh acts as a dose-dependentignal to induce diverse cell types in the neural tube (Martıt al., 1995a; Roelink et al., 1995; Ericson et al., 1997).erhaps differences in the relative amounts of Shh and Ptcctivities lead to distinct cell fate decisions.To explore the idea that a highly regulated balance in Shh
nd Ptc activities is necessary for normal development, wexamined the effects of Ptc overexpression in transgenicice. Here we show that excess Ptc attenuates induction of
hh target genes and causes abnormal patterning in theeural tube. These results demonstrate that Ptc is sufficient
o oppose Shh signaling and reveal additional roles for Ptc inevelopment and growth.Copyright © 1999 by Academic Press. All right
MATERIALS AND METHODS
Plasmids
449-8. To remove the 59 UTR, a piece of ptc1 was amplifiedfrom plasmid M2-1 (Goodrich et al., 1996) using primers PATG andS3. This product was digested and cloned into the AvrII and XbaIsites of M2-1 to make plasmid 331-2. To remove the 39 UTR, ptc1was amplified from plasmid M9 (Goodrich et al., 1996) usingprimers S5 and PTGA. This product, which ends at the ptc1 stopcodon, was digested and cloned into the NcoI and XbaI sites of M9to create plasmid 264-6. 331-2 was digested with SacI and EcoRV,filled in with T4 DNA polymerase, and then cloned into the EcoRVsite of 264-6, resulting in plasmid 447-1. 447-1 was digested withBsaBI to drop repeated sequences and then reclosed to createplasmid 449-8. This plasmid contains the entire ptc1 coding region,minus both 59 and 39 UTRs and including an optimized Kozakconsensus sequence for the AUG (Kozak, 1991). All PCR-derivedDNA and the cloning junctions were sequenced and found to becorrect.
487-1. A HindIII/XbaI fragment containing the second intronof the nestin gene was released from construct F (Zimmerman etal., 1994) and cloned into pTKb (Clontech). The resulting plasmidcontains a lacZ gene under the control of a thymidine kinase basicpromoter and the nestin enhancer. Plasmid sequences were re-moved by digestion with HindIII and SalI prior to microinjection.
499-6. The lacZ gene was removed from plasmid 487-1 bydigestion with NotI and replaced with a NotI fragment from 449-8that contains the ptc1 coding region. Plasmid sequences wereremoved by digestion with HindIII and SalI prior to microinjection.
Generation of Transgenic Mice
Purified linearized plasmids 487-1 and 499-6 were microinjectedinto fertilized oocytes from B6CBAF2 mice (Hogan et al., 1994). Totest the nestin enhancer, embryos injected with 487-1 were col-lected and stained with X-gal (Bonnerot and Nicolas, 1993). Aspreviously reported, staining was observed throughout the centralnervous system (Zimmerman et al., 1994). Two stable lines weregenerated from oocytes injected with 499-6: Nptc77 and Nptc118.Transgenic animals were genotyped by Southern blot or PCR.Genomic DNA was digested with EcoRI and probed with the 59NsiI fragment of the ptc1 cDNA. Due to the high copy numbers inboth lines, it proved difficult to determine with certainty whetheranimals were heterozygous or homozygous for the transgene; thehigh hybridization signal from the transgene generally obscured theendogenous bands, making it impossible to normalize betweensamples. PCR was performed with primers H56 and S2 under thefollowing conditions: 94°C 59; [94°C 300, 52°C 350, 72°C 900] 3 25cycles; 72°C 109. All transgenic lines were maintained usingheterozygous matings. For embryo collections, noon on the day ofplugging was called E0.5.
In Situ Hybridization
Endogenous ptc1 expression was detected using a 752-bp ribo-probe (M21-39 UTR) corresponding to nucleotides 4435–5187 in the39 UTR. Endogenous and exogenous ptc1 expression was detectedusing riboprobes from plasmids M2-2 and M2-3 (Goodrich et al.,1996). These two probes yielded identical results and will subse-
quently be referred to as ptc1 probes. The Shh probe was generatedfrom Hh-16.1 (Echelard et al., 1993). A 1.7-kb riboprobe encom-s of reproduction in any form reserved.

i(
ds(am
ipgnd
lmhpt
stgWpE
ghiEnetp(tnfl
bEbTodvosnoms
noafaIt
325Excess Ptc Interferes with Neural Patterning
passing most of the zinc finger domain was used to detect Gli1 (Huiet al., 1994). The Pax 3 probe corresponded to a 519-bp PstI/HindIIIfragment from the 39 end of the gene (Goulding et al., 1991).
Digoxygenin in situ hybridization and radioactive in situ hybrid-zation of sections were performed and photographed as describedStorm and Kingsley, 1996; Oro et al., 1997).
Immunohistochemistry
Immunohistochemistry was performed as described (Ericson etal., 1997) using mouse anti-Nkx2.2 (Ericson et al., 1997) and rabbitanti-Pax6 antibodies (MacDonald et al., 1995).
Double-Label in Situ Hybridization andImmunohistochemistry
In situ hybridization was performed as above but with thefollowing modifications. 1:5000 K5 anti-islet antibody (Ericson etal., 1992) was added to the anti-DIG antibody and incubatedovernight at 4°C. After several PBS washes, the sections wereincubated with secondary antibody in 1% normal goat serum.Depending on the secondary antibody, the signal either was di-rectly visualized using diaminobenzidine as a substrate or wasamplified and visualized using the VectaStain ABC system (VectorLaboratories). The slides were then washed in PBS and Buffer B1and transferred to Buffer B3 for visualization as in the original insitu hybridization protocol. Nuclei were stained with DAPI.
Histology
Embryos and postnatal brains were dissected in PBS, fixedovernight in 4% paraformaldehyde, and embedded in paraffinaccording to standard protocols. Six- to 8-mm paraffin sections wereeparaffinized in xylene and rehydrated through a graded alcoholeries before staining with hematoxylin (10 min) and eosin (2 min)Newcomer Supply). Excess stain was removed in water and 0.4%cid water. Sections were dehydrated, cleared in xylene, andounted in Permount (Sigma).
Primers
Primers used included PATG, GCCACCATGGCCTCGGCTG(Kozak sequence underlined); S3, CAAGGGCCACATCAAG; S5,GTTCTGGACGGTGCTG; PTGA, TGAGTTGGAGCTGCTC-CCC; H56, GTGGCTGAGAGCGAAGTTTCAG; and S2, GAAG-CAGTCCAAAGGTG.
RESULTS
Defects Caused by Overexpression of Ptc in theNeural Tube
To gain insight into the antagonistic relationship be-tween Shh and Ptc as well as the role of this pathway inneural development, we overexpressed ptc1 in the develop-ng murine nervous system. The coding region of ptc1 waslaced under the control of an enhancer from the nestinene that drives high-level expression in most central
ervous system progenitors beginning at about embryonicay 9 (E9) (Fig. 1A) (Zimmerman et al., 1994). Two viablest
Copyright © 1999 by Academic Press. All right
ines of mice, Nptc77 and Nptc118, were generated byicroinjection of embryos with this construct. Both lines
ave 10 to 20 copies of the transgene and exhibit the samehenotypes. The lines were maintained by matings ofransgenic animals.
The nestin enhancer drives high-level, persistent expres-ion of ptc1 in cells throughout the neural tube. In wild-ype embryos, ptc1 is expressed in a ventral to dorsalradient in the ventricular zone of the neural tube (Fig. 1B).e confirmed that ptc1 is overexpressed in the nestin
attern in E10.5, E11.5, and P0 animals from both lines. In10.5 and E11.5 transgenic embryos, ptc1 is strongly tran-
scribed throughout the anterior neural tube except for thefloor plate (Fig. 1C). In progressively more posterior regionsof the embryo, ptc1 is overexpressed more highly in theventral neural tube than in the dorsal neural tube (Fig. 1D).As in wild-type embryos, ptc1 is primarily expressed incells of the ventricular zone. In a small number of trans-genic embryos (2/19 examined), ptc1 transcription wasvariegated, causing a punctate staining pattern (Fig. 1E).
The dramatic increase in ptc1 RNA production in trans-enic lines matches the pattern reported for nestin en-ancer activity. Enhancer-driven transcription normallynitiates in the ventral forebrain, midbrain, and hindbrain of9 embryos and expands posteriorly and dorsally over theext 2 days of development (Zimmerman et al., 1994). Thenhancer continues to drive expression in neural progeni-ors throughout development. Similarly, ptc1 expressionersists in the postnatal cerebellum of transgenic animalsdata not shown). In summary, in Nptc animals, after neuralube closure, ptc1 is expressed at high levels throughouteurogenesis and throughout the neural tube, except for theoor plate.Nptc animals exhibited dramatic neural defects as em-
ryos, pups, and adults. Of 95 embryos collected between10.5 and E11.5, 13 (13.7%) had obviously abnormal hind-rains and/or partially open neural tubes (Figs. 2A and 2B).he hindbrain defect seems to be due to an apparentvergrowth of the neural tube in this region (Fig. 1E andata not shown) and a collapse of the thin roof of the fourthentricle (arrowhead, Figs. 2A and 2B). Spina bifida wasbserved to varying degrees ranging from failure to close inmall regions of the posterior neural tube to severe anterioreural tube defects (Fig. 2B). While transgenic embryosccasionally appeared smaller than their wild-type litter-ates, there was no consistent reduction in the area of the
pinal cord and no reproducible delay in development.While a majority of transgenic embryos were grossly
ormal, an increased incidence of perinatal lethality wasbserved in Nptc offspring. In the Nptc118 line, which hadslightly more severe phenotype, the average litter size
rom E10.5 to E11.5 was 8.16 (n 5 6 litters), but only anverage of 4.7 (n 5 7 litters) animals were alive at weaning.n addition, dying pups or carcasses were often observed athe day of birth. Hematoxylin and eosin staining of brain
ections from P0 pups revealed defects in 5/8 (62.5%)ransgenic animals. In the least affected animals, the ven-s of reproduction in any form reserved.
ns
dte(ispio
he coin A
326 Goodrich et al.
tricles were enlarged and the corpus callosum appearedslightly disorganized (Figs. 2C and 2D). One animal, which
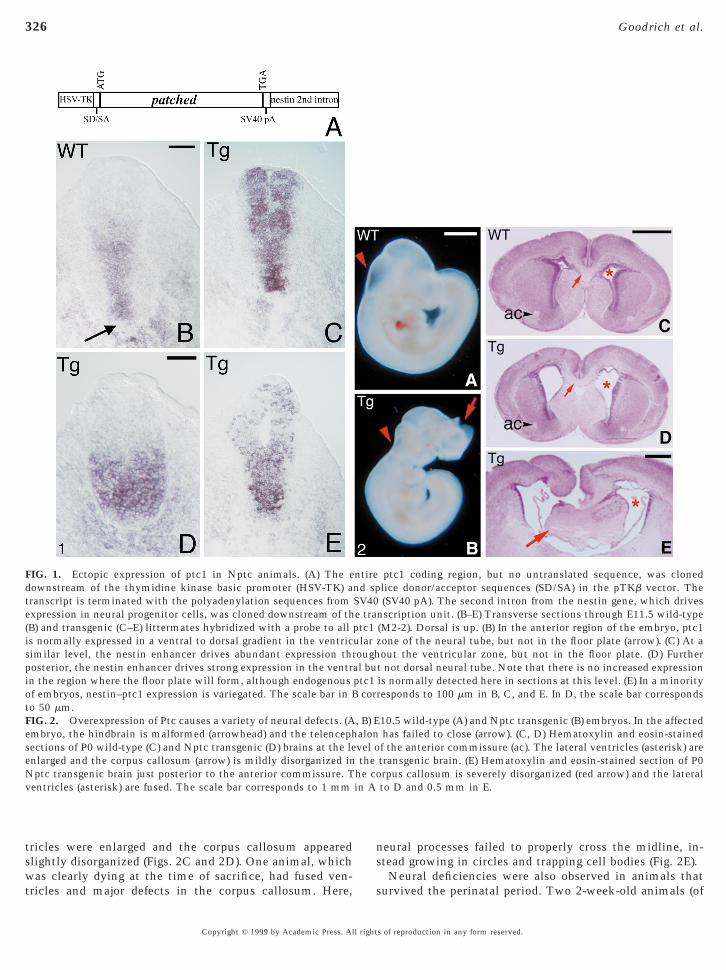
FIG. 1. Ectopic expression of ptc1 in Nptc animals. (A) The eownstream of the thymidine kinase basic promoter (HSV-TK) aranscript is terminated with the polyadenylation sequences fromxpression in neural progenitor cells, was cloned downstream of thB) and transgenic (C–E) littermates hybridized with a probe to alls normally expressed in a ventral to dorsal gradient in the ventricimilar level, the nestin enhancer drives abundant expression throsterior, the nestin enhancer drives strong expression in the ventrn the region where the floor plate will form, although endogenousf embryos, nestin–ptc1 expression is variegated. The scale bar in B
to 50 mm.FIG. 2. Overexpression of Ptc causes a variety of neural defects. (Aembryo, the hindbrain is malformed (arrowhead) and the telencephsections of P0 wild-type (C) and Nptc transgenic (D) brains at the leenlarged and the corpus callosum (arrow) is mildly disorganized inNptc transgenic brain just posterior to the anterior commissure. Tventricles (asterisk) are fused. The scale bar corresponds to 1 mm
was clearly dying at the time of sacrifice, had fused ven-tricles and major defects in the corpus callosum. Here, s
Copyright © 1999 by Academic Press. All right
eural processes failed to properly cross the midline, in-tead growing in circles and trapping cell bodies (Fig. 2E).
ptc1 coding region, but no untranslated sequence, was clonedlice donor/acceptor sequences (SD/SA) in the pTKb vector. The(SV40 pA). The second intron from the nestin gene, which drives
nscription unit. (B–E) Transverse sections through E11.5 wild-type(M2-2). Dorsal is up. (B) In the anterior region of the embryo, ptc1zone of the neural tube, but not in the floor plate (arrow). (C) At aout the ventricular zone, but not in the floor plate. (D) Further
t not dorsal neural tube. Note that there is no increased expressionis normally detected here in sections at this level. (E) In a minorityresponds to 100 mm in B, C, and E. In D, the scale bar corresponds
10.5 wild-type (A) and Nptc transgenic (B) embryos. In the affectedhas failed to close (arrow). (C, D) Hematoxylin and eosin-stainedf the anterior commissure (ac). The lateral ventricles (asterisk) aretransgenic brain. (E) Hematoxylin and eosin-stained section of P0rpus callosum is severely disorganized (red arrow) and the lateralto D and 0.5 mm in E.
ntirend spSV40e traptc1ularough
al buptc1
cor
, B) Ealonvel othe
Neural deficiencies were also observed in animals thaturvived the perinatal period. Two 2-week-old animals (of
s of reproduction in any form reserved.

epppnhPlwtpgtn
tnacdnwolsnLaaht
hbbbedtmn
assbsfptvmi(
eaflFputp
afsavdbrapn
eaoasmwn
327Excess Ptc Interferes with Neural Patterning
55 closely followed) were much smaller than their siblingsand died before weaning. One of these was obviouslyhydrocephalic. Hydrocephaly was also observed in 4 ani-mals after weaning; 3 additional animals were found dead at3 months of age but a cause of death could not be estab-lished due to deterioration of tissues. To better assess theincidence of hydrocephaly, brains from 24 transgenic ani-mals were collected at 6 to 8 weeks of age. Of these, 4(16.6%) had excess fluid and obviously deformed brains.Histological analysis of one of these brains showed exten-sive thinning of the cortex, fused ventricles, and massivenecrosis (data not shown). Hydrocephaly has never beenobserved in wild-type animals of the colony. Other trans-genic animals survive and reproduce without exhibitingobvious phenotypes.
Since some of the problems observed in newborn animalsand hydrocephalic adults may be secondary to earlier devel-opmental abnormalities, subsequent studies focused on theeffects of Ptc overexpression in the embryo.
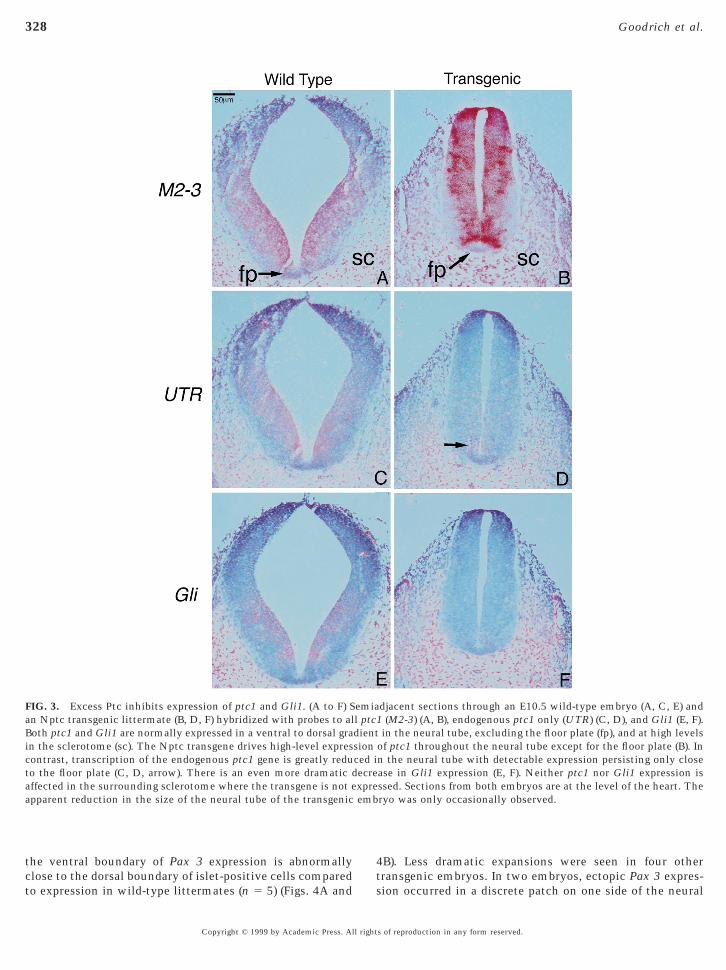
Excess Ptc Decreases Expression of ptc1 and Gli1in the Neural Tube
Hh and Ptc proteins have opposing activities: Hh inducesexpression of target genes that are otherwise kept off by Ptc.An apparently universal target is ptc1 itself. Hh seems tostablish a highly regulated feedback system by directingroduction of its own antagonist. In wild-type neural tubes,tc1 is therefore expressed at high levels close to the floorlate, which produces Shh, but at low levels in the dorsaleural tube (Fig. 1; Goodrich et al., 1996). Paradoxically,igh levels of ptc1 transcription correspond to low levels oftc function, due to the inactivation of Ptc by the Shhigand (Goodrich et al., 1997). Conversely, ptc1 is expressedeakly where the protein is most active. In Nptc animals,
he feedback loop is circumvented because expression oftc1 is under the control of the nestin enhancer. In trans-enic embryos, Ptc is produced in excess relative to Shh. Weested whether this imbalance interferes with the effective-ess of the Shh signal.Two good indicators of Shh signaling are the transcrip-
ional inductions of ptc1 and Gli1. To distinguish endoge-ous ptc1 from ptc1 produced from the transgene, we usedprobe from the ptc1 39 UTR, which is not included in the
onstruct (Fig. 1A). In 5/11 E10.5 embryos examined, en-ogenous ptc1 expression was significantly decreased in theeural tube, while expression in the surrounding mesodermas unaffected (Figs. 3C and 3D). Weaker effects werebserved in five other transgenic embryos (n 5 5 wild-typeittermates for comparison). Gli1 expression was even moreeverely affected (obvious decrease in six of six embryos;5 3 wild-type embryos for comparison) (Figs. 3E and 3F).
oss of ptc1 and Gli1 transcripts was most prominent in thenterior region of the embryo, probably reflecting earlyctivation of the nestin enhancer in the midbrain and
indbrain (Fig. 1; Zimmerman et al., 1994). The neuralubes shown in Fig. 3, for example, are at the level of thent
Copyright © 1999 by Academic Press. All right
eart; much less reduction in ptc1 and Gli1 expression cane seen at the level of the hindlimbs (data not shown). Sinceoth ptc1 and Gli1 are normally highly expressed longefore the stages we examined, these changes in genexpression are unlikely to be caused by any developmentalelay in transgenic embryos. These results suggest that Shharget gene expression is either not effectively induced oraintained in the presence of excess Ptc in the embryonic
eural tube.
Overexpression of Ptc Dorsalizes the Neural Tube
As indicated by ptc1 and Gli1 expression, excess Ptcppears to interfere either directly or indirectly with Shhignaling in the neural tube. Shh has been studied exten-ively as an inducer of floor plate development, but it haseen difficult to study other, later roles in the nervousystem. In Shh mutant embryos, for instance, no floor plateorms so it is impossible to know whether subsequentatterning errors are due to the absence of the floor plate orhe absence of Shh per se (Chiang et al., 1996). Based on initro experiments in chick and marker gene expression inouse, the floor plate and motor neurons appear to be
nduced early in development, before neural tube closureSasaki and Hogan, 1993; Echelard et al., 1993; Yamada etal., 1993). In Nptc embryos, the nestin enhancer initiatesctopic Ptc expression—reducing effective Shh signaling—fter neural tube closure and thus after induction of theoor plate and motor neurons (Zimmerman et al., 1994).urthermore, the transgene is not expressed in the floorlate (Figs. 1C and 3B). Nptc embryos therefore provide anique opportunity to study the role of this pathway afterhe floor plate forms and in intermediate and dorsal cellopulations.Cell culture experiments have shown that graded
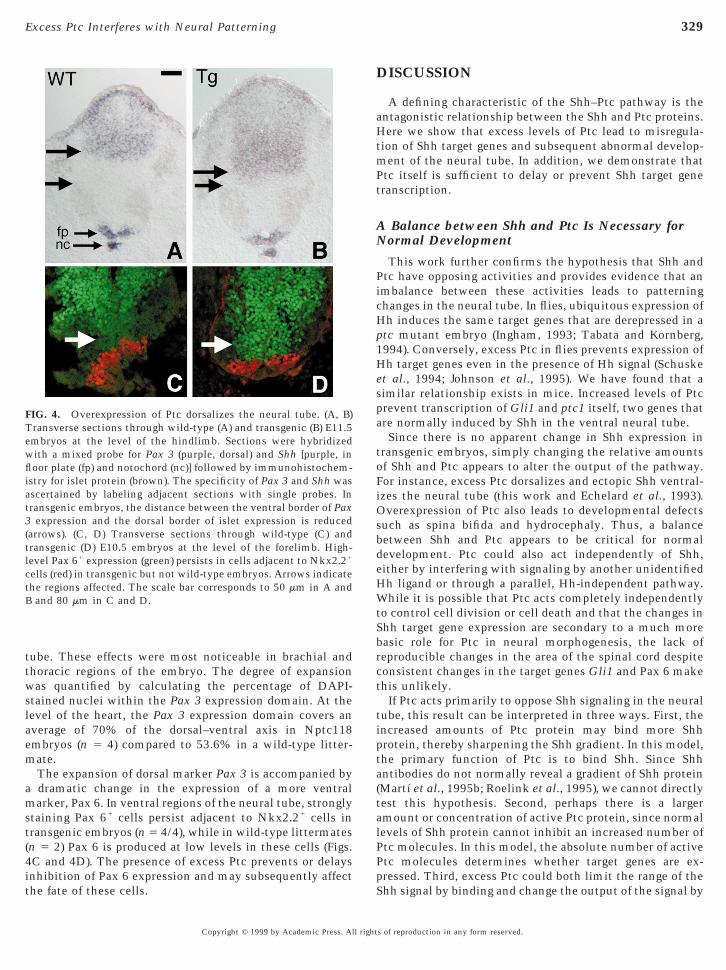
mounts of Shh regulate expression of key transcriptionactors and therefore generate diverse types of neurons atpecific dorsal–ventral levels in the neural tube (Martı etl., 1995a; Roelink et al., 1995; Ericson et al., 1997). Initro, small amounts of Shh protein inhibit expression oforsal marker Pax 3 and intermediate marker Pax 6. Inhi-ition of Pax 6 expression has the indirect effect of relievingepression of Nkx2.2, which is subsequently induced in,nd lateral to, the floor plate. Pax 3, Pax 6, and Nkx2.2ositive progenitors differentiate into specific kinds ofeurons along the dorsal–ventral axis.Changes in the expression of Pax 3 and Pax 6 in Nptc
mbryos reveal a role for the Ptc in regulating cell diversityfter formation of the floor plate. Floor plate developmentccurs normally as assessed by expression of Shh (Figs. 4And 4B) in E10.5–E11.5 transgenic embryos. In addition,taining with an a-islet antibody suggests normal develop-ent of islet-positive motor neuron progenitors, althoughe cannot rule out subtle changes in the motor neuronumber or pool identities (Figs. 4A and 4B). In the dorsal
eural tube, however, there is a marked expansion of Pax 3ranscription in some transgenic embryos (6/15) such thats of reproduction in any form reserved.
4
xpreemb
328 Goodrich et al.
the ventral boundary of Pax 3 expression is abnormally
FIG. 3. Excess Ptc inhibits expression of ptc1 and Gli1. (A to F) San Nptc transgenic littermate (B, D, F) hybridized with probes to alBoth ptc1 and Gli1 are normally expressed in a ventral to dorsal grain the sclerotome (sc). The Nptc transgene drives high-level exprescontrast, transcription of the endogenous ptc1 gene is greatly reduto the floor plate (C, D, arrow). There is an even more dramatic daffected in the surrounding sclerotome where the transgene is not eapparent reduction in the size of the neural tube of the transgenic
close to the dorsal boundary of islet-positive cells comparedto expression in wild-type littermates (n 5 5) (Figs. 4A and
ts
Copyright © 1999 by Academic Press. All right
B). Less dramatic expansions were seen in four other
djacent sections through an E10.5 wild-type embryo (A, C, E) and1 (M2-3) (A, B), endogenous ptc1 only (UTR) (C, D), and Gli1 (E, F).in the neural tube, excluding the floor plate (fp), and at high levels
of ptc1 throughout the neural tube except for the floor plate (B). Inn the neural tube with detectable expression persisting only closease in Gli1 expression (E, F). Neither ptc1 nor Gli1 expression isssed. Sections from both embryos are at the level of the heart. Theryo was only occasionally observed.
emial ptcdientsionced iecre
ransgenic embryos. In two embryos, ectopic Pax 3 expres-ion occurred in a discrete patch on one side of the neural
s of reproduction in any form reserved.
m
ams
4it
fli
329Excess Ptc Interferes with Neural Patterning
tube. These effects were most noticeable in brachial andthoracic regions of the embryo. The degree of expansionwas quantified by calculating the percentage of DAPI-stained nuclei within the Pax 3 expression domain. At thelevel of the heart, the Pax 3 expression domain covers anaverage of 70% of the dorsal–ventral axis in Nptc118embryos (n 5 4) compared to 53.6% in a wild-type litter-
ate.The expansion of dorsal marker Pax 3 is accompanied bydramatic change in the expression of a more ventralarker, Pax 6. In ventral regions of the neural tube, strongly
taining Pax 61 cells persist adjacent to Nkx2.21 cells intransgenic embryos (n 5 4/4), while in wild-type littermates(n 5 2) Pax 6 is produced at low levels in these cells (Figs.C and 4D). The presence of excess Ptc prevents or delays
FIG. 4. Overexpression of Ptc dorsalizes the neural tube. (A, B)Transverse sections through wild-type (A) and transgenic (B) E11.5embryos at the level of the hindlimb. Sections were hybridizedwith a mixed probe for Pax 3 (purple, dorsal) and Shh [purple, in
oor plate (fp) and notochord (nc)] followed by immunohistochem-stry for islet protein (brown). The specificity of Pax 3 and Shh was
ascertained by labeling adjacent sections with single probes. Intransgenic embryos, the distance between the ventral border of Pax3 expression and the dorsal border of islet expression is reduced(arrows). (C, D) Transverse sections through wild-type (C) andtransgenic (D) E10.5 embryos at the level of the forelimb. High-level Pax 61 expression (green) persists in cells adjacent to Nkx2.21
cells (red) in transgenic but not wild-type embryos. Arrows indicatethe regions affected. The scale bar corresponds to 50 mm in A andB and 80 mm in C and D.
nhibition of Pax 6 expression and may subsequently affecthe fate of these cells.
Copyright © 1999 by Academic Press. All right
DISCUSSION
A defining characteristic of the Shh–Ptc pathway is theantagonistic relationship between the Shh and Ptc proteins.Here we show that excess levels of Ptc lead to misregula-tion of Shh target genes and subsequent abnormal develop-ment of the neural tube. In addition, we demonstrate thatPtc itself is sufficient to delay or prevent Shh target genetranscription.
A Balance between Shh and Ptc Is Necessary forNormal Development
This work further confirms the hypothesis that Shh andPtc have opposing activities and provides evidence that animbalance between these activities leads to patterningchanges in the neural tube. In flies, ubiquitous expression ofHh induces the same target genes that are derepressed in aptc mutant embryo (Ingham, 1993; Tabata and Kornberg,1994). Conversely, excess Ptc in flies prevents expression ofHh target genes even in the presence of Hh signal (Schuskeet al., 1994; Johnson et al., 1995). We have found that asimilar relationship exists in mice. Increased levels of Ptcprevent transcription of Gli1 and ptc1 itself, two genes thatare normally induced by Shh in the ventral neural tube.
Since there is no apparent change in Shh expression intransgenic embryos, simply changing the relative amountsof Shh and Ptc appears to alter the output of the pathway.For instance, excess Ptc dorsalizes and ectopic Shh ventral-izes the neural tube (this work and Echelard et al., 1993).Overexpression of Ptc also leads to developmental defectssuch as spina bifida and hydrocephaly. Thus, a balancebetween Shh and Ptc appears to be critical for normaldevelopment. Ptc could also act independently of Shh,either by interfering with signaling by another unidentifiedHh ligand or through a parallel, Hh-independent pathway.While it is possible that Ptc acts completely independentlyto control cell division or cell death and that the changes inShh target gene expression are secondary to a much morebasic role for Ptc in neural morphogenesis, the lack ofreproducible changes in the area of the spinal cord despiteconsistent changes in the target genes Gli1 and Pax 6 makethis unlikely.
If Ptc acts primarily to oppose Shh signaling in the neuraltube, this result can be interpreted in three ways. First, theincreased amounts of Ptc protein may bind more Shhprotein, thereby sharpening the Shh gradient. In this model,the primary function of Ptc is to bind Shh. Since Shhantibodies do not normally reveal a gradient of Shh protein(Martı et al., 1995b; Roelink et al., 1995), we cannot directlytest this hypothesis. Second, perhaps there is a largeramount or concentration of active Ptc protein, since normallevels of Shh protein cannot inhibit an increased number ofPtc molecules. In this model, the absolute number of activePtc molecules determines whether target genes are ex-
pressed. Third, excess Ptc could both limit the range of theShh signal by binding and change the output of the signal bys of reproduction in any form reserved.

1lHmpwt1wscoappr
aae(tmttaaFbazootu
adbtatt
330 Goodrich et al.
actively inhibiting target gene transcription. In favor of thisinterpretation, excess Ptc in the fly imaginal disc causespatterning and growth defects when expressed only in theposterior compartment, where Hh is produced, but thephenotype is more severe when Ptc levels are increasedthroughout the wing disc (Johnson et al., 1995). In the firstcase, Ptc presumably limits the diffusion of Hh into theanterior compartment, where target genes are normallyexpressed, but has no effect on transcription of these genesin the posterior compartment, where they are not normallyexpressed. In the second case, Ptc interferes with Hhdiffusion and in addition actively inhibits transcription oftarget genes in the anterior compartment, thereby causing agreater effect. These two functions of the Ptc protein havebeen experimentally separated in the fly with a mutant Ptcprotein that can bind Hh but cannot efficiently inhibittranscription (Chen and Struhl, 1996).
The Hh Pathway Resists Subtle Changes in theLevel of Ptc Protein
A balance between Shh and Ptc appears to be rigorouslymaintained by the Hh signaling pathway itself. In fact, inboth flies and mice, it has been difficult to interrupt theHh–Ptc feedback loop. Initial reports suggested that over-expression of ptc in flies, while sufficient to rescue a ptcmutant, has no substantial effect on target gene transcrip-tion or the arrangement of cell types in the larval cuticle(Ingham et al., 1991; Sampedro and Guerrero, 1991). Simi-larly, we have found that overexpression of ptc1 in trans-genic mice often has no drastic phenotypic effect.
Two features of the pathway help to explain the negativeresults observed in both flies and mice. First, Ptc protein isquite unstable, with very little protein residing at theplasma membrane (Ingham et al., 1991; Capdevila et al.,994a). Second, Hh is a potent signal and may not beimiting in normal embryos. To overcome the effects of theh signal, Ptc protein must be present in great excess andaintained at that level, even in the presence of rapid
rotein turnover. In flies, a persistently high level of Ptcas accomplished by repeated induction of ptc or by using
he GAL4–UAS system (Schuske et al., 1994; Johnson et al.,995). In the present work, stable and high levels of Ptcere achieved by using the nestin enhancer, which is very
trong and drives enduring expression from neural tubelosure until the end of neurogenesis. Even so, developmentccurred fairly normally in the majority of transgenicnimals, further testament to the intrinsic resilience of thisathway. Since Hh itself ultimately controls the level of Ptcrotein, self-regulation within the pathway may alloweestablishment of a proper balance.
Phenotypic Variation in Nptc Animals
A notable feature of Nptc overexpression is the wide
amount of phenotypic variation within each Nptc line,ranging from early embryonic lethality to no visible effectCopyright © 1999 by Academic Press. All right
t all. Although transcription of the transgene is occasion-lly variegated (Fig. 1E), in general the amount of ptc1xpressed from the transgene is consistent within each linedata not shown). A second possibility is that insertion ofhe transgene interrupted another gene that somehowodifies the effect of excess Ptc. This is also unlikely since
he same range of defects was observed in two independentransgenic lines. Third, since these animals were created on
hybrid background, there may be polymorphisms indditional genes that suppress or enhance the phenotype.inally, some of the variation may be due to differencesetween embryos with one and two copies of the transgenicrray. Due to difficulties in definitively identifying hetero-ygote and homozygote embryos (see Materials and Meth-ds), we cannot comment on dose-dependent effects of Ptcverexpression. It is likely that multiple factors, includinghe Ptc dosage and the genetic background, affect theltimate phenotype.The nature of the Shh–Ptc signaling cascade itself may
lso contribute to the observed variation. During normalevelopment a balance between Shh and Ptc is establishedy a feedback loop that adjusts the amount of Ptc wheneverhe relative amounts of Shh and Ptc are changed. In Nptcnimals, although expression of the transgene is not underhe control of Shh or Ptc, Shh still regulates expression ofhe endogenous ptc1 gene (Fig. 3) and hence can still change
the total amount of Ptc protein. If cells assume distinct cellfates depending on the relative amount of Ptc activity, thenany changes in the final amount of active Ptc protein maylead to very different outcomes. Similar variation has beenobserved in hypomorphic alleles of fly ptc or when ptc isoverexpressed in the wing imaginal disc (Phillips et al.,1990; Capdevila et al., 1994b; Johnson et al., 1995). Further-more, ectopic expression of both Pax 3 and Pax 6,which are expanded in Nptc embryos, causes variablephenotypes with low penetrance (Schedl et al., 1996;Tremblay et al., 1996), suggesting that other signalingsystems may compensate for forced changes in the neuraltube.
Does Shh Act as a Morphogen in the Neural Tube?
Hh is proposed to act as a morphogen to pattern the flydorsal cuticle (Heemskerk and DiNardo, 1994), but it hasbeen difficult to demonstrate a direct role for graded Shhsignaling in the neural tube. If Shh is indeed a morphogen,then it should act in a dose-dependent fashion, the proteinshould be found in the responding cells, and the effectsshould be independent of secondary signaling proteins. Ourresults provide additional evidence for the first two criteria,but do not address the lingering problem of what othersignals might also be acting.
Excess Ptc causes predictable changes in gene expressionthat support the idea of a dose-dependent Shh signal. Invitro, twofold changes in the concentration of Shh regulate
the expression of three transcription factors—Pax 3, Pax 6,and Nkx2.2 (Ericson et al., 1996, 1997). Depending ons of reproduction in any form reserved.

aeevmaathiia
nom
pmammhns
331Excess Ptc Interferes with Neural Patterning
which of these transcription factors are expressed, progeni-tors develop as specific types of motor neurons or interneu-rons at defined locations in the neural tube. Overexpressionof Ptc causes an expansion of Pax 3 RNA and of high-levelPax 6 protein, both of which can be repressed by Shh invitro. Since Shh and Ptc have opposite effects on the sametarget genes, a simple model is that cell fates are deter-mined by the relative amount of Shh and Ptc activity. Thebalance is changed in the presence of excess Ptc, consistentwith the idea that different concentrations of Shh normallylead to different levels of Ptc activity and therefore differenttarget gene expressions. Although Ptc overexpression atleast partially dorsalizes the neural tube, we cannot resolvewhether individual ventral cells assume new fates, whethera population of dorsal progenitors divides and expandsventrally, or whether dorsal progenitors survive in abnor-mally ventral positions. Cells could become confused andexpress both dorsal and ventral markers, but the expansionof the dorsal marker Pax 3 appears to be coordinated with achange in the level of Pax 6 protein in more ventral cells.
The observed changes in Pax 3 and Pax 6 raise newquestions about when and how Shh patterns the neuraltube. In vitro, one of the earliest actions for Shh appears tobe inhibition of dorsal markers, including Pax 3 (Ericson etal., 1996). Subsequently, floor plate and motor neuronmarkers are induced in a dose-dependent fashion. Yet, inthe presence of excess Ptc, we see changes in the expressionof Pax 3 in the absence of changes to floor plate (indicatedby Shh RNA) and motor neurons (indicated by Islet protein).Why are the changes in Pax 3 expression limited to thedorsal neural tube? Shh inhibits Pax 3 expression for anextended period of time, first close to the floor plate andthen further away. Perhaps by the time Ptc levels aresufficient to block Shh signaling, Pax 3 expression ispermanently repressed in the most ventral cells, but not inthe more intermediate cell populations. Given the apparentcontinued need for Shh signaling after the initial inductionof cell fates, increased Ptc levels may interfere with themaintenance, but not the initiation, of Pax 3 repression. Inthis case, Pax 3 is initially turned off by Shh, but then isreexpressed in cells that still require effective Shh signaling.Both possibilities raise the interesting idea that intermedi-ate cell populations are patterned after ventral cell popula-tions or that these cells respond differently to Shh than theventral explant cells used for the in vitro experiments. Theultimate fate of the affected cells remains to be determined.Excess Ptc may delay rather than prevent the expression ofappropriate genes. Subtle changes in motor neurons or othercell populations cannot be assessed without additionalmarkers.
The effects of excess Ptc also provide indirect evidencefor long-range action of Shh. Whether Shh protein actsdirectly on cells outside of the floor plate is difficult todetermine. First, Shh protein, which is tightly associatedwith the cell surface, has not been detected outside of the
floor plate (Martı et al., 1995b; Roelink et al., 1995).However, function-blocking antibodies reveal a require-cc
Copyright © 1999 by Academic Press. All right
ment for Shh in motor neuron induction (Ericson et al.,1996). We have found that overexpression of Ptc inhibitsexpression of Gli1 and ptc1, even far away from the floorplate. Taken together with previous evidence that both Shhand Ptc regulate Pax 3 and Pax 6 and that Ptc opposes Shhactivity (Goodrich et al., 1997), the simplest interpretationof these results is that Shh protein normally reaches thecells that express these target genes. This implies that, invivo, Shh protein has a range of at least as far as the ventralborder of Pax 3. Alternatively, Ptc overexpression mightaffect only the cells closest to the floor plate and somehowinterfere with production of a different signal that indepen-dently regulates expression of Gli1, ptc1, Pax 3, and Pax 6.However, there is no evidence for such a signal. If Shhprotein does in fact travel as far as the ptc1 expressionpattern implies, then it will be important to determine howthe protein gets there and why it is so difficult to detect.
Roles for Shh and Ptc in Neural Tube Patterning
Very little is known about Shh and Ptc activities in thenervous system outside of the developing spinal cord.However, the other phenotypes observed in Nptc animalsmight also be caused by a forced imbalance between Shhand Ptc. For example, the fusion of ventricles and themidline defects observed in some newborn Nptc pups arereminiscent of holoprosencephaly (HPE), a disorder thatarises in people who are heterozygous for Shh (Roessler etl., 1996). HPE-like phenotypes are also caused by ectopicxpression of Pax 3 throughout the neural tube (Tremblayt al., 1996). Indeed, Pax 3 is ectopically expressed in theentral neural tube of Nptc embryos (Fig. 4). While there areany potential causes for the hydrocephaly seen in Nptc
dults, it is interesting to note that mice mutant forpolipoprotein B have a 50% reduction in plasma choles-erol levels and exhibit a number of neural defects includingydrocephaly (Homanics et al., 1995). A cholesterol moietys added to Hh proteins during autoproteolysis and may bemportant for their function (Lee et al., 1994; Bumcrot etl., 1995; Porter et al., 1996a,b). In addition, two teratogens,
cyclopamine and jervine, are sterols and can inhibit induc-tion of Shh target genes in neural explants (Cooper et al.,1998; Incardona et al., 1998). Further analysis of the Nptc
eural phenotypes may reveal Hh-independent roles for Ptcr additional, unexpected roles for Shh or other Hh familyembers.Although the details remain a mystery, the Hh–Ptc
athway clearly has an essential role in human develop-ent and disease. We have found that an imbalance in Shh
nd Ptc leads to cell fate changes and subsequently abnor-al development in the murine nervous system. Similarisregulation of the Hh pathway is associated with two
uman genetic disorders, holoprosencephaly and basal cellevus syndrome, and also with sporadic cancer. Under-tanding how this balance is achieved has important impli-
ations for future treatment of human birth defects andancer.s of reproduction in any form reserved.

H
H
H
H
H
H
H
332 Goodrich et al.
ACKNOWLEDGMENTS
We thank Dr. C. Kaznowski and Dr. L. Milenkovic for assistancewith brain collection and Dr. S. McConnell and Dr. D. Kingsley foradvice and helpful discussions. Antibodies were kindly provided byDr. T. Jessell (Columbia University) and Dr. R. MacDonald (KingsCollege London). L.V.G. was supported by a predoctoral fellowshipfrom the Howard Hughes Medical Institute. M.P.S. is an investi-gator of the Howard Hughes Medical Institute.
REFERENCES
Bitgood, M. J., Shen, L., and McMahon, A. P. (1996). Sertoli cellsignaling by desert hedgehog regulates the male germline. Curr.Biol. 6, 298–304.
Bonnerot, C., and Nicolas, J. (1993). Application of lacZ genefusions to postimplantation development. In “Guide to Tech-niques in Mouse Development” (P. M. Wassarman and M. L.DePamphilis, Eds.). Academic Press, San Diego.
Bumcrot, D. A., Takada, R., and McMahon, A. P. (1995). Proteolyticprocessing yields two secreted forms of Sonic hedgehog. Mol.Cell. Biol. 15, 2294–2303.
Capdevila, J., Estrada, M. P., Sanchez-Herrero, E., and Guerrero, I.(1994b). The Drosophila segment polarity gene patched interactswith decapentaplegic in wing development. EMBO J. 13, 71–82.
Capdevila, J., Pariente, F., Sampedro, J., Alonso, J. L., and Guerrero,I. (1994a). Subcellular localization of the segment polarity pro-tein patched suggests an interaction with the wingless receptioncomplex in Drosophila embryos. Development 120, 987–998.
Carstea, E. D., Morris, J. A., Coleman, K. G., Loftus, S. K., Zhang,D., Cummings, C., Gu, J., Rosenfeld, M. A., Pavan, W. J.,Krizman, D. B., Nagle J., Polymeropoulos, M. H., Sturley, S. L.,Ioannou, Y. A., Higgins, M. E., Comly, M., Cooney, A., Brown,A., Kaneski, C. R., Blanchette-Mackie, E. J., Dwyer, N. K.,Neufeld, E. B., Chang, T. Y., Liscum, L., Strauss, J. F., III, Ohno,K., Zeigler, M., Carmi, R., Sokol, J., Markie, D., O’ Neill, R. R.,van Diggelen, O. P., Elleder, M., Patterson, M. C., Brady, R. O.,Vanier, M. T., Pentchev, P. G., and Tagle, D. A. (1997). Niemann-Pick C1 disease gene: Homology to mediators of cholesterolhomeostasis. Science 277, 228–231.
Chen, Y., and Struhl, G. (1996). Dual roles for patched in seques-tering and transducing hedgehog. Cell 87, 553–563.
Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L.,Westphal, H., and Beachy, P. A. (1996). Cyclopia and defectiveaxial patterning in mice lacking Sonic hedgehog gene function.Nature 383, 407–413.
Cooper, M. K., Porter, J. A., Young, K. E., and Beachy, P. A. (1998).Teratogen-mediated inhibition of target tissue response to Shhsignaling. Science 280, 1603–1607.
Echelard, Y., Epstein, D. J., St-Jacques, B., Shen, L., Mohler, J.,McMahon, J. A., and McMahon, A. P. (1993). Sonic hedgehog, amember of a family of putative secreted signaling molecules, isimplicated in the regulation of CNS polarity. Cell 75, 1417–1430.
Ericson, J., Morton, S., Kawakami, A., Roelink, H., and Jessell,T. M. (1996). Two critical periods of sonic hedgehog signalingrequired for the specification of motor neuron identity. Cell 87,661–673.
Ericson, J., Rashbass, P., Schedl, A., Brenner-Morton, S., Kawakami,A., Heyningen, V. v., Jessell, T. M., and Briscoe, J. (1997). Pax 6
controls progenitor cell identity and neuronal fate in response tograded Shh signaling. Cell 90, 169–180.Copyright © 1999 by Academic Press. All right
Ericson, J., Thor, S., Edlund, T., Jessell, T. M., and Yamada, T.(1992). Early stages of motor neuron differentiation revealed byexpression of homeobox gene Islet-1. Science 256, 1555–1560.
Fan, C., and Tessier-Lavigne, M. (1994). Patterning of mammaliansomites by surface ectoderm and notochord: Evidence for scle-rotome induction by a hedgehog homolog. Cell 79, 1175–1186.
Forbes, A. J., Nakano, Y., Taylor, A. M., and Ingham, P. W. (1993).Genetic analysis of hedgehog signalling in the Drosophila em-bryo. Development Suppl., 115–124.
Gailani, M. R., Ståhle-Backdahl, M., Leffell, D. J., Glynn, M.,Zaphiropoulos, P. G., Pressman, C., Unden, A. B., Dean, M.,Brash, D. E., Bale, A. E., and Toftgård, R. (1996). The role of thehuman homologue of Drosophila patched in sporadic basal cellcarcinomas. Nature Genet. 14, 78–81.
Goodrich, L. V., Johnson, R. L., Milenkovic, L., McMahon, J., andScott, M. P. (1996). Conservation of the hedgehog/patched sig-naling pathway from flies to mice: Induction of a mouse patchedgene by hedgehog. Genes Dev. 10, 301–312.
Goodrich, L. V., Milenkovic, L., Higgins, K. M., and Scott, M. P.(1997). Altered neural cell fates and medulloblastoma in mousepatched mutants. Science 277, 1109–1113.
Gorlin, R. J. (1987). Nevoid basal-cell carcinoma syndrome. Medi-cine 66, 98–109.
Goulding, M. D., Chalepakis, G., Deutsch, U., Erselius, J. R., andGruss, P. (1991). Pax-3, a novel murine DNA binding proteinexpressed during early neurogenesis. EMBO J. 10, 1135–1147.
Hahn, H., Wicking, C., Zaphiropoulous, P. G., Gailani, M. R.,Shanley, S., Chidambaram, A., Vorechovsky, I., Holmberg, E.,Unden, A. B., Gillies, S., Negus, K., Smyth, I., Pressman, C.,Leffell, D. J., Gerrard, B., Goldstein, A. M., Dean, M., Toftgård,R., Chenevix-Trench, G., Wainwright, B., and Bale, A. E. (1996).Mutations of the human homolog of Drosophila patched in thenevoid basal cell carcinoma syndrome. Cell 85, 841–851.ahn, H., Wojnowski, L., Zimmer, A. M., Hall, J., Miller, G., andZimmer, A. (1998). Rhabdomyosarcomas and radiation hypersen-sitivity in a mouse model of Gorlin syndrome. Nature Med. 4,619–622.ammerschmidt, M., Brook, A., and McMahon, A. P. (1997). Theworld according to hedgehog. Trends Genet. 13, 14–21.eemskerk, J., and DiNardo, S. (1994). Drosophila hedgehog acts asa morphogen in cellular patterning. Cell 76, 449–460.ogan, B., Beddington, R., Costantini, F., and Lacy, E. (1994).“Manipulating the Mouse Embryo: A Laboratory Manual.” ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.omanics, G. E., Maeda, N., Traber, M. G., Kayden, H. J., Dehart,D. B., and Sulik, K. K. (1995). Exencephaly and hydrocephaly inmice with targeted modification of the apolipoprotein B (Apob)gene. Teratology 51, 1–10.
Hooper, J. E. (1994). Distinct pathways for autocrine and paracrineWingless signalling in Drosophila embryos. Nature 372, 461–464.ui, C.-C., Slusarski, D., Platt, K. A., Holmgren, R., and Joyner,A. L. (1994). Expression of three mouse homologs of the Drosoph-ila segment polarity gene Cubitus interruptus, Gli, Gli-2, andGli-3, in ectoderm- and mesoderm-derived tissues suggests mul-tiple roles during postimplantation development. Dev. Biol. 162,402–413.ynes, M., Stone, D. M., Dowd, M., Pitts-Meek, S., Goddard, A.,Gurney, A., and Rosenthal, A. (1997). Control of cell pattern in
the neural tube by the zinc finger transcription factor andoncogene Gli-1. Neuron 19, 15–26.s of reproduction in any form reserved.

I
I
J
J
J
J
K
K
K
L
L
L
M
M
M
M
M
O
P
P
P
P
P
R
R
R
R
R
R
S
S
S
333Excess Ptc Interferes with Neural Patterning
Incardona, J. P., Gaffield, W., Kapur, R. P., and Roelink, H. (1998).The teratogenic Veratrum alkaloid cyclopamine inhibits sonichedgehog signal transduction. Development 125, 3553–3562.
ngham, P. W., and Hidalgo, A. (1993). Regulation of winglesstranscription in the Drosophila embryo. Development 117, 283–291.
ngham, P. W., Taylor, A. M., and Nakano, Y. (1991). Role of theDrosophila patched gene in positional signalling. Nature 353,184–187.
ohnson, R. L., Grenier, J. K., and Scott, M. P. (1995). patchedoverexpression alters wing disc size and pattern: Transcriptionaland post-transcriptional effects on hedgehog targets. Develop-ment 121, 4161–4170.
ohnson, R. L., Laufer, E., Riddle, R. D., and Tabin, C. J. (1994).Ectopic expression of sonic hedgehog alters dorsal–ventral pat-terning of somites. Cell 79, 1165–1173.
ohnson, R. L., Rothman, A. L., Xie, J., Goodrich, L. V., Bare, J. W.,Bonifas, J. M., Quinn, A. G., Myers, R. M., Cox, D. R., Epstein, E.,Jr., and Scott, M. P. (1996). Human homolog of patched, acandidate gene for the basal cell nevus syndrome. Science 272,1668–1671.
ohnson, R. L., and Scott, M. P. (1998). Control of cell growth andfate by patched genes. Cold Spring Harb. Symp. Quant. Biol. 62,205–215.inzler, K. W., Bigner, S. H., Bigner, D. D., Trent, J. M., Law, M. L.,O’ Brien, S. J., Wong, A. J., and Vogelstein, B. (1987). Identifica-tion of an amplified, highly expressed gene in a human glioma.Science 236, 70–73.
ozak, M. (1991). An analysis of vertebrate mRNA sequences:Intimations of translational control. J. Cell Biol. 115, 887–903.rauss, S., Concordet, J.-P., and Ingham, P. W. (1993). A function-ally conserved homolog of the Drosophila segment polarity genehh is expressed in tissues with polarizing activity in zebrafishembryos. Cell 75, 1431–1444.
ee, J., Platt, K. A., Censullo, P., and Ruiz i Altaba, A. (1997). Gli1is a target of sonic hedgehog that induces ventral neural tubedevelopment. Development 124, 2537–2552.
ee, J. J., Ekker, S. C., von Kessler, D. P., Porter, J. A., Sun, B. I., andBeachy, P. A. (1994). Autoproteolysis in hedgehog protein bio-genesis. Science 266, 1528–1537.
oftus, S. K., Morris, J. A., Carstea, E. D., Gu, J. Z., Cummings, C.,Brown, A., Ellison, J., Ohno, K., Rosenfeld, M. A., Tagle, D. A.,Pentchev, P. G., and Pavan, W. J. (1997). Murine model ofNiemann-Pick C disease: Mutation in a cholesterol homeostasisgene. Science 277, 232–235.acdonald, R., Barth, K. A., Xu, Q., Holder, H., Mikkola, I., andWilson, S. W. (1995). Midline signaling is required for Pax generegulation and patterning of the eyes. Development 121, 3267–3278.arigo, V., Davey, R. A., Zuo, Y., Cunningham, J. M., and Tabin,C. J. (1996a). Biochemical evidence that patched is the Hedgehogreceptor. Nature 384, 176–179.arigo, V., Johnson, R. L., Vortkamp, A., and Tabin, C. J. (1996b).Sonic hedgehog differentially regulates expression of GLI andGLI3 during limb development. Dev. Biol. 180, 273–283.artı, E., Bumcrot, D. A., Takada, R., and McMahon, A. P. (1995a).Requirement of 19K form of sonic hedgehog for induction ofdistinct ventral cell types in CNS explants. Nature 375, 322–325.artı, E., Takada, R., Bumcrot, D. A., Sasaki, H., and McMahon,A. P. (1995b). Distribution of sonic hedgehog peptides in the
developing chick and mouse embryo. Development 121, 2537–2547.Copyright © 1999 by Academic Press. All right
ro, A. E., Higgins, K. M., Hu, Z., Bonifas, J. M., Epstein, E. H., andScott, M. P. (1997). Basal cell carcinomas in mice overexpressingsonic hedgehog. Science 276, 817–821.
hillips, R. G., Roberts, I. J. H., Ingham, P. W., and Whittle, J. R. S.(1990). The Drosophila segment polarity gene patched is in-volved in a position-signalling mechanism in imaginal discs.Development 110, 105–114.
ietsch, T., Waha, A., Koch, A., Kraus, J., Albrecht, S., Tonn, J.,Sorensen, N., Berthold, F., Henk, B., Schmandt, N., Wolf, H. K.,von Deimling, A., Wainwright, B., Chenevix-Trench, G., Wiest-ler, O. D., and Wicking, C. (1997). Medulloblastomas of thedesmoplastic variant carry mutations of the human homologueof Drosophila patched. Cancer Res. 57, 2085–2088.
latt, K. A., Michaud, J., and Joyner, A. L. (1997). Expression of themouse Gli and ptc genes is adjacent to embryonic sources ofhedgehog signals suggesting a conservation of pathways betweenflies and mice. Mech. Dev. 62, 121–135.
orter, J. A., Ekker, S. C., Park, W. J., von Kessler, D. P., Young,K. E., Chen, C. H., Ma, Y., Woods, A. S., Cotter, R. J., Koonin,E. V., and Beachy, P. A. (1996a). Hedgehog patterning activity:Role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell 86, 21–34.
orter, J. A., Young, K. E., and Beachy, P. A. (1996b). Cholesterolmodification of hedgehog signaling proteins in animal develop-ment. Science 274, 255–259.affel, C., Jenkins, R. B., Frederick, L., Hebrink, D., Alderete, B.,Fults, D. W., and James, C. D. (1997). Sporadic medulloblastomascontain PTCH mutations. Cancer Res. 57, 842–845.
iddle, R. D., Johnson, R. L., Laufer, E., and Tabin, C. (1993). Sonichedgehog mediates the polarizing activity of the ZPA. Cell 75,1401–1416.oberts, D. J., Johnson, R. L., Burke, A. C., Nelson, C. E., Morgan,B. A., and Tabin, C. (1995). Sonic hedgehog is an endodermalsignal inducing BMP-4 and Hox genes during induction andregionalization of the chick hindgut. Development 121, 3163–3174.oelink, H., Augsburger, A., Heemskerk, J., Korzh, V., Norlin, S.,Ruiz i Altaba, A., Tanabe, Y., Placzek, M., Edlund, T., Jessell,T. M., and Dodd, J. (1994). Floor plate and motor neuroninduction by vhh-1, a vertebrate homolog of hedgehog expressedby the notochord. Cell 76, 761–775.
oelink, H., Porter, J. A., Chiang, C., Tanabe, Y., Chang, D. T.,Beachy, P. A., and Jessell, T. M. (1995). Floor plate and motorneuron induction by different concentrations of the amino-terminal cleavage product of sonic hedgehog autoproteolysis.Cell 81, 445–455.
oessler, E., Belloni, E., Gaudenz, K., Jay, P., Berta, P., Scherer,S. W., Tsui, L. C., and Muenke, M. (1996). Mutations in thehuman sonic hedgehog gene cause holoprosencephaly. Nat.Genet. 14, 357–360.
ampedro, J., and Guerrero, I. (1991). Unrestricted expression of theDrosophila gene patched allows a normal segment polarity.Nature 353, 187–190.
asaki, H., and Hogan, B. L. M. (1993). Differential expression ofmultiple fork head related genes during gastrulation and axialpattern formation in the mouse embryo. Development 118,47–59.
asaki, H., Hui, C.-C., Nakafuku, M., and Kondoh, H. (1997). Abinding site for Gli proteins is essential for HNF-3b floor plate
enhancer activity in transgenics and can respond to Shh in vitro.Development 124, 1313–1322.s of reproduction in any form reserved.

S
S
S
T
T
U
V
V
V
W
W
X
Y
Z
334 Goodrich et al.
Schedl, A., Ross, A., Lee, M., Engelkamp, D., Rashbass, P., Heynin-gen, V. v., and Hastie, N. D. (1996). Influence of Pax 6 genedosage of development: Overexpression causes severe eye abnor-malities. Cell 86, 71–82.
chuske, K., Hooper, J. E., and Scott, M. P. (1994). patchedoverexpression causes loss of wingless expression in Drosophilaembryos. Dev. Biol. 164, 300–311.
tone, D. M., Hynes, M., Armanini, M., Swanson, T. A., Gu, Q.,Johnson, R. L., Scott, M. P., Pennica, D., Goddard, A., Phillips,H., Noll, M., Hooper, J. E., de Sauvage, F., and Rosenthal, A.(1996). The tumour-suppressor gene patched encodes a candidatereceptor for sonic hedgehog. Nature 384, 129–134.
torm, E. E., and Kingsley, D. M. (1996). Joint patterning defectscaused by single and double mutations in members of the bonemorphogenetic protein (BMP) family. Development 122, 3969–3979.abata, T., and Kornberg, T. (1994). Hedgehog is a signaling proteinwith a key role in patterning Drosophila imaginal discs. Cell 76,89–102.remblay, P., Pituello, F., and Gruss, P. (1996). Inhibition of floorplate differentiation by Pax 3: Evidence from ectopic expressionin transgenic mice. Development 122, 2555–2567.nden, A. B., Holmberg, E., Lundh-Rozell, B., Ståhle-Backdahl, M.,Zaphiropoulos, P. G., Toftgård, R., and Vorechovsky, I. (1996).Mutations in the human homologue of Drosophila patched(PTCH) in basal cell carcinomas and the Gorlin syndrome:Different in vivo mechanisms of PTCH inactivation. Cancer Res.56, 4562–4565.on Ohlen, T., Lessing, D., Nusse, R., and Hooper, J. E. (1997).Hedgehog signaling regulates transcription through cubitus in-terruptus, a sequence-specific DNA binding protein. Proc. Natl.Acad. Sci. USA 94, 2404–2409.
orechovsky, I., Tingby, O., Hartman, M., Stromberg, B., Nister,M., Collins, V. P., and Toftgård, R. (1997). Somatic mutations inCopyright © 1999 by Academic Press. All right
the human homologue of Drosophila patched in primitive neu-roectodermal tumours. Oncogene 15, 361–366.
ortkamp, A., Lee, K., Lanske, B., Segre, G. V., Kronenberg, H. M.,and Tabin, C. J. (1996). Regulation of rate of cartilage differen-tiation by Indian hedgehog and PTH-related protein. Science 273,613–622.alterhouse, D., Ahmed, M., Slusarski, D., Kalamaras, J., Boucher,D., Holmgren, R., and Iannaccone, P. (1993). Gli, a zinc fingertranscription factor and oncogene, is expressed during normalmouse development. Dev. Dyn. 196, 91–102.olter, M., Reifenberger, J., Sommer, C., Ruzicka, T., and Reifen-berger, G. (1997). Mutations in the human homologue of theDrosophila segment polarity gene patched (PTCH) in sporadicbasal cell carcinomas of the skin and primitive neuroectodermaltumors of the central nervous system. Cancer Res. 57, 2581–2585.ie, J., Johnson, R. L., Zhang, X., Bare, J. W., Waldman, F. M.,Cogen, P. H., Menon, A. G., Warren, R. S., Chen, L. C., Scott,M. P., and Epstein, E., Jr. (1997). Mutations of the PATCHEDgene in several types of sporadic extracutaneous tumors. CancerRes. 57, 2369–2372.
amada, T., Pfaff, S. L., Edlund, T., and Jessell, T. M. (1993).Control of cell pattern in the neural tube: Motor neuron induc-tion by diffusible factors from notochord and floor plate. Cell 73,673–686.immerman, L., Parr, B., Lendahl, U., Cunningham, M., McKay,R., Gavin, B., Mann, J., Vassileva, G., and McMahon, A. (1994).Independent regulatory elements in the nestin gene direct trans-gene expression to neural stem cells or muscle precursors.Neuron 12, 11–24.
Received for publication January 28, 1999
Revised March 30, 1999Accepted April 1, 1999s of reproduction in any form reserved.





























