Embed Size (px)
Citation preview

Neuroscience Vol. 55, No. 4, pp. 953-964, 1993 Printed in Great Britain
0306-4522/93 $6.00 + 0.00 Pergamon Press Ltd
0 1993 IBRO
RAPID CHANGES IN THE FREQUENCY TUNING OF NEURONS IN CAT AUDITORY CORTEX RESULTING
FROM PURE-TONE-INDUCED TEMPORARY THRESHOLD SHIFT
M. B. CALFoRD,*j’ R. RAJAN$ and D. R. F. IRVINE$
$Department of Psychology, Monash University, Clayton, Victoria, Australia
*Vision, Touch and Hearing Research Centre, Department of Physiology and Pha~awlo~, The University of Queensland, Queensland 4072, Australia
A&&act-The response areas (frequency by intensity) of single neurons in primary auditory cortex of anesthetized cats were studied before and after temporary threshold shifts in cochlear sensitivity induced by an intense pure tone. Cochlear temporary threshold shift was monitored through the threshold of the gross auditory nerve compound action potential and in most cases involved a notch-like loss centered at the characteristic frequency of the unit under study. Only two neurons showed changes in response area that mirrored the changes at the auditory periphery. Most neurons (14) showed more complex changes involving both expansion and contraction of response areas. Expansion of response areas was indicated by lower thresholds at some frequencies and by the emergence of ~nsitivity to previously ineffective frequencies. A change was classified as contraction when the response area after the intense-tone exposure was smaller fhan would be expected by applying the profile of the temporary threshold shift to the initial response area. Contraction of both upper (high intensity) and lower boundaries of response areas was found; in the most extreme cases, neurons were totally unresponsive after the intense-tone exposure.
The complexity of effects of temporary threshold shifts on the response areas of cortical neurons is likely to be related to mechanisms that normally determine the frequency response limits of these neurons. The response areas of cortical neurons are more complex than those of auditory nerve fibers, and are thought to reflect the integration of excitatory and inhibitory inputs. The variety of effects observed in this study are consistent with the excitatory and inhibitory components of the response area of a given neuron being differenti~ly affected by the temporary threshold shift.
A brief intense sound can produce a temporary loss in cochlear sensitivity (temporary threshold shift, TTS) that is restricted to a limited range of frequen- cies. The effects of such restricted TTS on frequency selectivity at the cochlea are due to the temporary loss in cochlear mechanical and receptojr sensitivity at the affected frequencies. 9*1Q~29,30 Losses in sensitivity are manifested in auditory nerve (AN) fibers with re- sponse areas (tuning curves) that cover the affected frequencies.
In contrast to the wealth of studies on the effects of TTS on frequency tuning of auditory nerve fibers, only a few studies have examined the effect of TTS on frequency tuning in the central auditory sys- tem33*YJ2*” and there have heen no studies on the effects at the auditory cortex. The excitatory response areas of cortical neurons are complex compared with those of AN fibers,3’,32 although sharpness of tuning close to threshold is not different from the lower pathway.’ One aspect of this complexity is the
tTo whom correspondence should be addressed Abbreuiations: AI, primary auditory cortex; AN, auditory
nerve; BF, best frequency defined by maximal discharge; CF, characteristic frequency; Nl, first wave of the gross auditory nerve potential; SPL, sound pressure level; TTS, temporary threshold shift.
presence of surround inhibition in the response areas of many neurons. The nature of the tuning curves suggests that factors affecting neural sensitivity, such as a hearing loss at the cochlea, may affect the tuning of cortical neurons in a more complicated way than they affect the tuning of AN fibers. Thus, the aim of the present study was to describe the effects of an acute peripheral hearing loss, created by exposure to an intense pure tone, on the excitatory response areas of single neurons in primary auditory cortex (AI). Furthermore, since surround inhibition is a factor likely to shape the tuning propertiks of AI neurons, this aspect of neural response areas was specifically examined in our study.
Another motivation for this study came from the consideration of neuronal plasticity in adult animals. Reorganization of the topographic representation of frequency in cortex has heen reported following long-term lesions of limited regions of the cochlea in guinea-pigs.& Equivalent reorganizations have been reported in visual and somatosensory cortices follow- ing small lesions of the retina2 or digit ~pu~tions,2* respectively. In the somatosensory system, it has been found that, after a small peripheral denervation, receptive fields of affected cortical neurons expand rapidly into the areas surrounding the denervated region.’ In the longer term these expanded fields
953

954 M. B. CALFORD er al,
shrink-j Thus there is an initial unmasking of pre- viously ineffective inputs followed by a form of plasticity which allows some of these inputs to be- come dominant. It is of interest then to compare the short term and long term effects of a hearing loss that is limited in frequency. Whereas the Iong-term response was studied with direct rn~h~~l lesions of the co&lea,” in this study we used intense-tone induced TTS. It was not approp~te to study the immediate effects of a mechanical lesion since the procedure is likely to have an acute desensitizing effect extending over a wide frequency range. Also the use of TTS allows the response areas of individual neurons to be compared before and after the hearing loss.
Two alternative procedures have been used in studies of the effects of such TTS on central auditory neurons. In some studies the response characteristics of neurons in animals with noise-induced hearing fosses were compared to those in normal-hearing animals.““*43 Although this procedure has the advan- tage of allowing data to be collected from a large number of neurons, it has the disadvantage that differences attributable to noise exposure are con- founded with differences between the neuronal samples in the two groups of animals. This is a major limitation where the structure under investigation is characte~~d by considerable heterogeneity of neur- onal response characte~stics, as is the case in AI.92 The alternative procedure, used in other previous studies of the central auditory system2~,33,~.~’ and also in the present study, is to study the responses of an individual neuron before and after the induction of a peripheral bearing loss. ~though this method has the disadvan~g~ of allowing only a single neuron (or, by judicious choice of frequencies tested as detailed below, a small number of neurons) to be studied in each animal, it has the advantage of allowing direct within-cell comparisons. Recovery from the effects of intense pure-tone exposure on the cochlea involves a rapid phase of l-5 min followed by a slow phase in which thresholds change by only a few decibels over a period of hours. In the present study, the post- exposure effects on neurons in AI were examined in the slow recovery phase.
Experiments were carried out on I I adutt cats (Monash University animal house), shown by otoscopic examination to he free of middle- and outer-ear infections. Each cat was anesthetized with sodium ~nto~rbi~ (40 mg/kg, i.p.> and was given an injection of atropine sulphate (0.2 trig, i.m.) to reduce mucous secretions. The trachea and a radial vein were ~nulat~, and anesthesia was rn~n~~~ during surgery and pb~iolo~~l recording by additional 1.v. inleo tions of s&un &ntobarbitat _ aS required [appiox. lS-2Smgfkg ner h). The methods used to rnmoort the animal’s &ad, expose the right tympanic bulla, a~~irn~l~t a round window electrode to monitor @ochtear potentials have been described in detail previo~sly~” The bulla was
vented by a length of fine-bore polyethylene tubing to allow static pressure eq~lization within the middle ear. A portion of the skull was removed to expose the ectosylvian gyri on the left side, and the dura was relIeeted. A modified Davies chamber was mounted on the skull such that its top was approximately parallel to the surface of the middle eetosyl- vian gyms. The chamber was filled with silicone oil.
The cat was ioeated in an electrically shielded, sound- attending room, The stimuhrs delivery system was changed during the course of this study. Detailed accounts of the stimulus generation and delivery system used in the first l&e experiments have been provided eisewhere?3*a Briefly, pure tone stimuli generated by a computer- controlled frequency synthesizer were gated and shaped as tone bursts by a digitimer and electronic switch before delivery through a computer-controlled attenuator, am- plifier and condenser microphone driver. In experiments involving two-tone stimuli (see below), this system was used to generate the variable “test” stimulus, and a second, manually-controlled oscillator, electronic switch, attenuator and amplifier were used to generate the fixed “probe” stimulus. The outputs of the two channels were mixed before delivery to the transducer. In later experiments the tone pulses were generated by a digital stimulus system3’ and transdu~ by a Beyer DT48 h~dphone. In two-tone experiments using this system, the test and probe tones were generated by the two channels of the digital stimulating system and combined in the same manner as in the first series of experiments. In the course of data collection, sound pressure levels (SPLs in dB re 2OpPa) were set under computer control (Nova 4C or PDP 1 l-73) in accordance with calibrations of the sound delivery system. In the first series of experiments this calibration was conducted in siru at the beginning of each experiment using a probe tube attached to a Briiel and Kjaer 4133 microphone with the tip positioned near the tympanic membrane.” In the later experiments calibration of the sound delivery system was made using a closed system coupling of the sound delivery tube to a Briiel and Kjaer 4133 microphone, and the calibration table was common to all experiments in this series. In both series the output of the microphone amplifier (Briiel and Kjaer 2807) was digitized and the attenuators were adjusted to maintain the output of the amplifier within its operating range as the signal was swept across frequency. The attenuation and output voltage of the amplifier at each frequency were used to generate a correction table for use during stimulus presentation, Cochlear potentials, recorded with an electrode an the round window, were amplified, filtered (0.3-10.0 kHz) and displayed on an oscilloscope. Extracellular recordings of cortical neuronal activity were made using glass-insulated tungsten microelectrodes, which were advaneed into the cortex by means of a stepper-motor- driven miniature hydraulic microdrive mounted on the glass top-plate of the Davies chamber. Neural activity was am- plified (x lo), shaped by a graphic equalizer, and dis- played on an oscilloscope and audio-monitor. The action potentials of well-isolated neurons were converted by a Schmitt trigger to rectangular pulses which were sent to the computer. Response histograms and spike-count data were generated on-line, and stimulus and response event times were stored for later analysis.
The audiogram of the first wave of the gross auditory nerve potential (NI), for short tone pulses (40 ms duration; 1 ms rise-fall time; 8 kHz rate) f&m 2 to 4OkHz, was determined with the visual detection threshold methods7 at the start of each experiment to ascertain if it was within normal limits for the eat. Subsequent measures of the audiogram were used to assess the extent of TTS produced by exposure to intense pure tones.

Effects of temporary ~eshold shift on auditory cortex
A I3 0
955
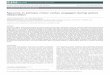
Fig. I. Two examples of the effects of intense pure tone exposure on the Ni potential half an octave above the exposure frequency. Over the first few minutes after presentation of the intense tone there is a rapid recovery of sensitivity. The loss is then maintained within 1 or 2 dB of this level for approximately 1 h;
redeterminations of neural response areas were made during this period.
The lo&ion of mimoelectrode penetrations in AI was established by the conformity of the cbaraeteristic frequency (CF; frequency at which threshold is lowest) of neurons in successive penetrations to the established tonotopi~ty of this field.n*” In the first five ex~~rn~~, the ~~hoId tuning curves of isolated single neurons {unit numbers pre- fixed with 88) were stank manually by varying the frequency and §I% of the stimufus and noting the bound- aries of response areas. These boundaries were later check& and, if necessary, adjusted on the basis of subsequent spike-count data. In later experiments (units prefixed with 89), a manual estimate was made of the CF and threshold of a neuron, and tone bursts were then presented under computer control at various frequency-intensity combi- nations in a matrix, the frequency range of which centered on the neuron’s estimated CF. Measurements were made over as wide an SPL range as possible, to ensure that the upper boundaries of the response areas of neurons with circumscribed response areas were well defined. The entire fr~~~y~n~sity matrix was sampled in ~~Qr~dorn order, with a single tone-pulse presented at each frequency- intensity combination. The number of repetitions of the matrix (usually four or eight, with each repetition in a difI&nt order) was determined by the experimenter. After the bust repetition of tbe matrix, the total munber of spikes falling within a specified count-window was dis- played on a terminal and printed out for each point in the matrix.
Testing the egtcts of intense pure-tone expomres
A brief intense tone at a frequency greater than 3 kHx can induce a TTS affactng a limited range of frequencies with n%tximum etTect at a freqmy half an octave above the stimuIating frequency. t325 Hence, in the present study, the frequency of a tone used to induce ‘ITS was always set at half an octave lower than the CF of the unit under study. Prior to the intense-tone exposure, the first wave of the gross auditory nerve potential (NI) audiogram was ~ete~n~ and the efIect of the exposure was subsequently monitored in the Nf ~01~. Since data on tb cochlear effects of pure-tone exposures in the cat are limited, the intensity and duration of exposure had to be arbitrarily set on the basis of @~perien~e with the cochkar effects of intense sounds in the guinea-pig.” Immediately after exposure, very larae loa& in sensitivity of the Nl potential were found (up io 64 dR), but sensitivity rapidly recoveted (5-20 min) to stabil- ize at a level which varied further by only 1-3 dR over many hours (Fig. 1). if after stabiIixati& the initial exposure had not produced a strfficient loss in the Ni audiogram at CF,
a second exposure was presented. During the period in which losses in the Ni potential threshold at CF were stable the excitatory response area for the unit was reexamined and a full NI audio~am was coIlected. It was not possible to compare neural responses once thmahol~ had returned to normal since this recovery would have exceeded the period of the experiments.
Since the et&t of the TTS, as determined by changes to the Nl a~~og~, was either notch-like around the most affected frequency (e.g. Fig. 43, C) or had a broad effect on higher frequencies (Figs 3B, 4A), it was possible to examine effects on more than one cortical neuron per animal pro- vided that the response area of a subsequently studied neuron was well below the previously a&ted frequencies (see Fig. 2 in which all units come from the same animal). &nplo$ng this strategy we were able, in the best case, to isolate and test four cortical neurons (with pro~ively lower CFs), but in each of six animals only one unit was successfully studied.
Auditory cortical neurons in ~rbitura~“an~theti~ cats have little or no resting discharge, and inhibitory input to these neurons can only be detected in extra&IuIar record- ings by a change in the response to some form of probe stimulus. To avoid the complications introduced at the cochlea by simultaneous presentation of two tones, for example,“*‘* we used a two-tone paradigm of the type used in forward-masking experiments. The effect of a first (“test”) tone of variable frequency and intensity on the rqonse to a tixed second (“probe”) tone was determined. Each tone pulse was 50 ms in duration with a 4-ms rise-fall time, and the interval between test-tone offset and probe-tone onset was 40 or 50 ms. The probe tone was always at the neuron’s CF and at a level 1.5 dB above CF threshold. The test tone was varied over a range of frequencies and intensities extending well beyond the neuron’s excitatory response area. In the tist series of experiments, the two-tone stimuIus was presented 20 times at each point in the fmquency- intensity matrix for the test stimulus. In the second series of experiments, the frequencyintensity matrix was sampled pseudo-randomly in the same fashion as in the determi- nation of the tuning curve, with either four or, more commonly, eight repetitions. In each series, appropriately specified count windows allowed the responses to the test and probe stimuli to be measured separately. Criteria for delineating inhibitory surrounds are discussed in the Results.

956 M. B. CALFORD et al.
1
Frequency (kHz)
0- 4 15 20 25 30
Frequency (kHz)
Fig. 2. (A, B) Examples of neuronal response profiles in which ‘ITS produced a change in response similar to that expected of primary auditory nerve afferents, i.e. AN-like. (C) The single cell in this study for which there was no discernible change in response area as a result of T’fS. The solid lines indicate the limits of the excitatory response area (tuning curve) prior to TTS. Lines with closed circles indicate (in dB) the losses in the gross auditory nerve potential (Nl) produced by the intensstone-induced ‘ITS. The lines with onen circles outline the extent of the excitatory response area after induction of ‘ITS. Thus, to compare pie- and post-l% response area limits compare thesolid lines with no symbols to those with open circles. (A) Unit 89-26-3. CF = 15.7 kHz. sound exoosure Il.2 kHz at 105 dBSPL for 1 min. fB) Unit 89-26-5. CP = 7 kHz, exposure 5 kHz at 105 dBSPL ior 2 min. (C) Unit 89-26-L CF = 24 kHz, exposure 17.1 kHz
at 1lOdBSPL for 1 min. Shaded areas are regions of masking suppression as de&d in Fig. 5.
RESULTS
Effect of peripheral threshold losses on excitatory response areas of cortical neurons
Seventeen units were isolated for enough time to examine possible changes in their excitatory response areas resulting from peripheral threshold losses caused by pure-tone exposures. TTS produced a variety of changes in tuning and no two units showed identical effects. Nevertheless, a simple analysis of the data reveals four broad categories of effect: AN-like, expanded response area, contracted response area, and no response after TTS, and one cell for which there was no discernible change in the response area. Representative examples of these types of effects are illustrated in Figs 2-4. To allow judgment of effects attributable directly to the peripheral TTS, the figures present pre-‘ITS (solid lines) and post-TTS (open circles) excitatory response areas of the AI neurons, together with the relative effect of the intense pure- tone exposure on the Nl audiogram (filled circles) on the same axes. However, in comparing the neural and Nl effects, the limits of the Nl response in reflecting the response of AN fibers tuned to a particular frequency must be recognized. Whereas the Nl threshold at a given frequency is normally determined by the sensitivity of fibers with CF equal to the test frequency, this need not be the case after pure-tone TTS. A loss in threshold of fibers tuned to a restricted frequency range may be compensated for in the Nl potential by the o&CF response of fibers tuned to
adjacent less affected frequencies. Characterization of the peripheral loss in neural sensitivity in terms of Nl loss also does not address the issue of possible variation in the effects of TTS with the threshold of individual AN fibers. Nevertheless, the effect of the exposure on the Nl audiogram provides a rough measure of the extent of the ‘ITS and a means for comparing the effect across the combination of fre- quencies and exposure times used in this study.
For three neurons (17%), the changes in the excit- atory response area resulting from the TTS appeared to reflect peripheral loss as determined by changes in the Nl audiogram (Fig. 2). Since for these neurons the effects of intense sounds are similar to the effects of intense sounds on auditory nerve fibers, primarily
a loss in sensitivity at the tip of the tuning curve, we have termed the effects AN-like. In one case (Fig. 2B), the losses in the Nl audiogram were reflected almost precisely (within the IO-dB resolution of the tuning curve determination) in the loss of sensitivity in the unit’s excitatory response area. This correspondence was not as clear for the example shown in Fig. 2A, where the sharply tuned peak of the original tuning curve was lost after TTS but where there was little change in sensitivity to frequencies outside this area. Fig. 2C presents an example in which the full extent of the loss in the Nl potential is not retlected in the loss in the neural threshold at CF and for which the post-TTS curve is marginally expanded with respect to the initial tuning curve at a number of frequencies. Since the changes are small and are close to the

Effects of temporary threshold shift on auditory cortex 957
6 9 12 15 18 16 20 22 24 26 28 30 Frequency (kHz) Frequency (kHz)
Fig. 3. Examples of response profiles recorded from neurons that showed an expansion of response area as a result of TTS. Symbols as for Fig. 2 with the addition in D of lines with squares which show the change in the unit’s excitatory response area (open squares) and Nl sensitivity (closed squares) following a second exposure to an intense tone. (A) Unit 88-49-3, CF = 6.4 kHz, sound exposure 4.6 kHz at 104 dBSPL for 4 min. (B) Unit 89-24-3, CF = 12 kHz, exposure 8.58 kHz at 110 dBSPL for 1 mm. (C) Unit 88-49-2, CF = 12.2 kHz, exposure 8.7 kHz at 102 dBSPL for 15 min. (D) Unit 88-53-4, CF = 26 kHz, first exposure 18.6 kHz at 105 dBSPL for 5 min, second exposure 18.6 kHz at 110 dBSPL for 10 min. Shaded
areas are regions of masking suppression as defined in Fig. 5.
resolution of the frequency by level matrix used to study this cell, it is not clear that any change has taken place.
A neuron was considered to have shown expansion of its excitatory response area if the post-TTS tuning curve revealed responsiveness at frequencies to which the unit was previously insensitive (at any sound level). Fig. 3 presents TTS-induced changes in the excitatory response areas of four units classified as having expanded response areas. The unit illustrated in Fig. 3A originally had a circumscribed response area, the most sensitive region (6-7.5 kHz) of which was lost after ‘ITS. However, at 5 and 5.5 kHz, the unit became more sensitive and the response area boundaries expanded considerably in both frequency and sound level (upper) dimensions. A similar expan- sion was found at lower frequencies for the unit illustrated in Fig. 3B. However, this unit, which originally had a nonmonotonic response profile at 11 and 12 kHz, displayed a contraction of the response area at higher frequencies. This pattern of a contrac- tion of the response ares at some frequencies, to- gether with a considerable expansion of the response area at other frequencies, was found for one other unit. The TTS produced when studying the unit illustrated in Fig. 3C was greater than intended. This not only resulted in a massive loss of sensitivity at all frequencies to which the cell was originally responsive but also in considerable expansion of the responsive- ness to higher frequencies. The neural response profile illustrated in Fig. 3D displays a minor expan-
sion in the response area at low frequencies and a generalized loss in threshold at other frequencies after an initial small TTS. A larger TTS produced further contraction of the response area over most of its frequency range but also resulted in a region of sensitivity to fairly intense stimulation at originally ineffective low frequencies. This was the most mar- ginal change in the sample of units classified as having shown expansion of their response area.
Neurons classified as having a contracted response area exhibited a post-TTS reduction of response area that exceeded the peripheral losses. The TTS shown in Fig. 4A was broad and increased with frequency. The effect on the tuning of the unit, CF at 15 kHz, was to reduce sensitivity at higher frequencies and also to produce an area nonresponsive to high-inten- sity stimulation at 14 and 15 kHz (cross-hatching). The neural response illustrated in Fig. 4B showed a similar reduction (open circles), involving the appear- ance of an unresponsive area at high intensities and a general shrinking of the total response area follow- ing induction of a notch-like TTS (filled circles). A subsequent TTS resulted in a larger and broader Nl threshold loss and a further contraction of the re- sponse area. In addition to the three units that showed contraction of their response areas following TTS, four units failed to respond after TTS (e.g. Fig. 4C). The fact that spontaneous discharges could still be recorded from these units indicated there had been no change in the recording conditions, but no pure-tone stimuli tested could elicit discharges. No

0 8 10 12 14 16 18 20 22 3 6 9 12 15
1 c Frequency &Hz) s&l Z
g60 40
-20 12 16 20 24 28 32
Flequmcy(k~z)
Fig. 4. Exampies of unit response profiles in which there was a contraction of the excitatory response Ada as a result of 8n intense-tone induced TIS. Symbols as in Figs ! and 2. (A) Unit 89-22-Z CF = 14.7 kI+z, the area marked with diagonal lines indiites a rqion wfrich was unresponsive after induction of ‘ITS. The intense tone (exposure IO.7 kIiZ at I f O dFSPL for 5 ntin) induced a broad k&h f%quency toss and no Nf p&e&al was apparent above 19 kHz. (B) Uait 8&50-I, CF = 9.2 kHz t&t exposure 6.7 kHz at fOS d3SPL for 5 mia, second exposure 6.7 kHz at fO5 dEIsPL for fO min. (C) One of four units in Be sampie for which, after induction of the ITS, there was no r-se to tonat stimuli; unit W-24-2, CF = 24 kflz, exposure 17.1 kHz at I fO dBSPL for 3 min. Shaded areas are regions of masking
suppression as deftned in Fig. 5.
features of the original response areas, or of the ‘ITS produced, otherwise distin~ish the experiments on these units.
Relationship of temporary threshoid shift inducpd changes and bthibitim in the original response ureas
Two character&ties which show aspects of inhibi- tory ~nt~butious to the response properties of corti- cal neurons were examined for correlation with the outcome of TTS: the presence of lateral inhibition and nonmonotonicity of the spike-count versus sound level function. Prior to the induction of TTS, the 17 units were studied with a forward masking paradigm (see Experimental Procedures) to reveal the extent of inhibitory response areas. In Figs 2-4 regions of forward masking suppression are pre- sented as shaded regions. Shading extending beyond the excitatory tuning curve indicates regions of sur- round- or ~ate~~-in~biti~~. The method used to dehneate the inhibitory regions is presented in Fig. 5, where the spike counts to a probe stimulus (15 dB above thresh&d at CF) are plotted as a Freon of the stim&s levei and f&+rency of a preceding test tone (40 ms iu~tirnul~ interval). Suppression within the limits of the excitatory response area cannot, with certainty, be at~but~ to inhibitor mechanisms, since it could be the result of both central and peripheral adaptation processes. How- ever, it is noteworthy that the total suppression of the response to the probe tone found, for most of our sampb, when the test stimulus was within the excit-
atory response area, exceeds the reported 20-40% suppression in auditory nervea and eochlear nucleus’ for comparable inte~~mulus delays. Response sup- pression outside the excitatory response area is not seen in AN fibers and must therefore reflect central inhibitory processes. The examples shown in Fig. 5R-I3 show clear areas of surround inhibition. A complete description of the extent of this inhibitian in temporal and frequency domains was not at- tempted in this study. The method was used simply to test if the TTS-induced changes in the response areas are related to the presence of surround inhi- bition. A criterion of greater than 50% reduction in spike count to the probe tone was used to draw the boundaries of the region of suppression (interpolated where uecessary). Thus the responses summarized in Figs 2A, 4C and SA, for which the region of suppres- sion does not extend beyond the excitatory response area (the single point in Fig. 5A is not considered s~~i~~t)~ are deemed not to have inhibitory sur- rounds No~onotoni~ty in the response pro&e at CF was de&d as a greater than 50% reduction in discharges to stimula~on at or above 80dBSPL compared with the ~~~ discharge (non- monotoni~ty at non-CF frequencies was judged rela- tive to the urns discharge at those f~~~cies).
Of the 17 neurons studied with TTS, 12 showed surround inhibition, In terms of the shape of the discharge rate to sound level function, eight had nonmonotonic profiles, six had monotonic profRes and three showed either profile depending upon the

Effects of temporary threshold shift on auditory cortex 959
frequency of stimulation (mixed). In Table 1 these data are compared to the effect of ITS for each cell studied. Clear relationships between the indicators of inhibitory contributions to the response area and outcome of pure-tone TTS are not obvious. Only three of the units showed no evidence of inhibition in terms of either indicator; one of these was unrespon- sive after TTS and two showed changes which were considered AN-like.
Although there was a variety of change in the overall response area as a result of TTS, the loss in sensitivity at CF reflected the loss in Nl-~tential threshold at the corresponding frequency in the com- bined sample of neurons (Fig. 6A). The strong linear- ity of this relationship, which did not discriminate the groupings of neurons according to the overall effect of TTS, showed a near linear gain (Fig. 6A, re- gression slope = 1.1, correlation r = 0.86, P < 0.05). Of course the four units which failed to respond after TTS could not be included in this analysis. Despite the significant linear relationship there is considerable scatter in the loss in neural threshold for similar Nl threshold losses (e.g. around 15 dB). Fig. 6B shows that for the Nl threshold loss range of 15-20 dB, this scatter appears to be accounted for by variation in the absolute thresholds of the neurons at CF. Units with lower thresholds had progressively larger losses.
A
312433
313*42
24322 4422443
222222221‘24311
z 3 4 5 7 8 9 lo 11 12
DISCUSSION
The major result of this study is that reduced sensitivity of the auditory periphery over a restricted frequency range centered at, or about, the CF of neurons in AI can produce changes to the excitatory response areas of AI neurons that do not always simply reflect the peripheral changes. This change is most often expansion of the response area (seven neurons) but may also be contraction of the response area (three neurons) or total disappearance of the driven response (four neurons). Three neurons showed changes in response area that appeared simi- lar to those reported at the auditory periphery. It is this variety of change resulting from ITS directed at the CF of each unit that is the most important finding of this study, for such variation is not a feature of TTS effects at lower levels of the auditory pathway.
At the cochlea, the basic effect of TTS induced by a pure tone is to decrease the responsiveness of AN fibers tuned to some frequencies while having little effect on fibers tuned to other frequencies. If the response area of AI neurons with a given CF were derived solely from AN fibers with the same CF, the effects of TTS on AI neurons would be expected to be a simple reflection of the changes seen at the auditory nerve. However, a number of studies provide evidence indicating that the response areas of AI neurons (and of central auditory neurons at
D
Fig. 5. Illustration of the excitatory (solid lines) and forward masking determined inhibitory response areas (stippling) for four single units from primary auditory cortex. The spike counts shown represent the response to a SO-ms probe tone at the combination marked by the * when preceded (40 or 50 ms delay) by a ZO-ms test tone at the combinations of frequency and sound level indicated by the position of the spike-count value. In A and B spike counts are for four repetitions of the stimulus while those in C and D represent counts to 20 repetitions. The area in which the teat tone suppresses the respone to the probe tone was defined as the limits in frequency and level where the test tone produced a reduction of greater
than 50% in responses to the probe tone, and is shaded.

960 M. B. CALFORD et al.
subcortical levels) are derived via convergent input from AN fibers tuned to a range of frequencies.8’32’4* Such convergence is proposed for both excitatory and inhibitory inputs. While no clear relationship was found between either of the two indicators of inhibi- tory components of receptive fields which we exam- ined (Table 1) and the effects of TTS, it is clear that there is ample evidence for inhibitory components in the response of the vast majority of cortical neurons, We propose that, in the AI neurons that showed expansion of their excitatory response areas after TTS, there was a greater effect upon the inhibitory inputs to those neurons than upon the excitatory inputs. The opposite effect would account for the contraction or disappearance of the response area of other neurons. Furthermore, the ‘ITS-induced change appears to depend on the excitatory/ inhibitory inputs elicited at each combination of frequency and intensity. Data from two neurons (e.g. Fig. 3B) give emphasis to this point, since expansion of the excitatory response area occurred at some frequencies but contmction of the response area occurred at others. The proposed differential effects of TTS on central excitatory and inhibitory channels is in accord with similar suggestions proposed pre- viously to explain post-TTS changes in response patterns,5’ maximal response rates2”TM or single-tone inhibition of spontaneous activity43 of central audi- tory neurons.
Four neurons failed to respond to tonal stimuli after TTS (e.g. Fig. 4C). Given the induction of, or increased, nonmonotonicity to stimulus level shown for some neurons (Fig. 4A, B), and the losses at low intensities due directly to the TTS seen for all neur- ons, the disappearance of response in these neurons can be viewed as an extreme form of contraction of
the response area and not as a separate category of TTS-induced change. Thus, the increasing contrac- tion of response areas illustrated by the neurons in Fig. 4A-C may be viewed as a continuum,
In this study there was no aspect of the original response profile of a neuron which predicted the pattern of change that resulted from TTS (Table 1). However, all seven neurons showing expansion post- TTS showed at least one of the two ~m~nents of inhibition that we examined, as did six of seven units showing either contraction or disappearance of the response area post-ITS. There was an expectation that neurons with clear excitatory/inhibitory inter- actions, as evidenced by lateral inhibition, would show the greatest changes resulting from TTS. While this was generally the case, there were exceptions, and neurons with lateral inhibition showed both contrac- tion and expansion of response areas. A problem with the two indices of inhibition used is that neither is useful in measuring inhibition in the stimulus regions most affected by pure-tone ITS (namely low sound levels around a neuron’s CF), as nonmonotonicity indicates inhibition at high sound levels, and forward masking confounds central inhibitory effects with peripheral masking and habituation when the mask- ing stimulus is inside the response area. Also, it is not possible to compare the extent of forward masking prior to TTS with that obtained from similar methods after ITS. This is because the boundaries of forward masking vary with the sound level of the probe tone and choice of an appropriate probe level after TTS (either in absolute or threshold relative terms) would be arbitrary.
It was surprising to find that a plot of the loss in ~nsitivity of the Nl potential against neural threshold loss, at CF, showed near unity gain even
Table 1. Relationship of the outcome of pure-tone temporary threshold shift directed at the characteristic frequency of each unit to two indicators of inhibitory components of the response area
Unit number Figure Level function Forward masking lateral inhibition
Effect of T’TS on response area
89-26- 1 2c 89-26-3 2A 89-26-5 2B, 5A 88-45-2 SC 88-53-4 3D 88-49-2 3c 88-49-3 3A 88-46-2 89-24-3 89-25-3 88-50- 1 89-22-2 89-27-6 89-24-2 89-25-2 88-49- 1
1B 3B, 5B
4B, IA 4A
4c
5D
M M
!z M
NM NM NM NM NM
Mixed Mixed Mixed
& NM
Yes No No Yes Yes Yes Yes
No change AN-like AN-like
Expansion Expansion Expansion Exnansion
YeS Expansion Yes Expansion No Expansion YeS Shrinkage Yes Shrinkage Yes Shrinkage No No response No No response Yes No resnonse
89-26-6 NM Yes No resbnse
The spike count to stimulus level function was classed as monotonic (M) or nonmonotonic (NM); some neurons showed either profile dependent upon stimulus frequency. The presew~ or absence of lateral inaction as seen in a forward masking paradigm (see Fig. 5) is indicated. AN-Eke-change that may be expected from an auditory nerve fiber-see text.

Effects of temporary threshold shift on auditory cortex %I
o-l/ I . * a 0 I 0 10 20 30 40 50
Loss in k4;;ensitivity
-I 01 IO 20 30 40 50
Fig. 6. (A) Piot of the increase in unit threshold against loss in sensitivity of the auditory nerve potential @I) at the CF of the unit for sin@e exposures to an intense pure tone. The four units which did not respond after ITS and one unit for which the Nl threshold was not measured exactly at CF are not shown. The dashed line shows a linear regression analysis (slope 5 1.1, y-intercept = -3 dBf which gave a high correlation between the two losses (r = 0.86). (B) The relationship between unit threshold at CF to the loss in this threshold for cases in which the effect of TTS was to reduce
the threshold of the Nl potential by 1520dB.
when the TTS was relatively large (Fig. 6A), and given that the sample was made up from all of the cells that were responsive post-TTS. A prediction of the form of this relationship is not possible since both unitary responses and the Nl potential involve re- cruitment across frequency channels at higher inten- sities. However, for a given cell, such recruitment would be expected to be related to the type of effect that TTS induced in the overall response area (expan- sion, contraction, AN-like). The four units that be- came unresponsive could not be considered in this analysis. The absence of these cells and the finding that losses are affected by the absolute sensitivity of the neurons (Fig. 6B), indicate that the strong linear- ity of this ~lations~p may be somewhat artificial. The effect of threshold is pro~bly a result of across fr~uency~~~el interaction at higher intensities such that frequency-specific losses in the Nl potential need not be directly reflected when neural thresholds
at CF are high. An example is provided in Fig. 4A where the best threshold of the excitatory response was well above the threshold of inhibition, illustrating an interaction of excitation and inhibition in the formation of the response area of this neuron even at threshold at CF. After the production of a large TTS in the Nl response, which had a broad ~~-fluency profile and a loss of 21 dB at CF, the corresponding loss in neural sensitivity was only 10dB. Alterna- tively, for neurons with greatest sensitivity (matching that of the Nl) the relationship between Nl threshold change and neural losses would be expected to have unity gain for small losses (e.g. Fig. 3A).
Comparison of temporary threshold shift induced changes in primary auditory cortex tuning to effects in lower auditory nuclei
The effects in this study can be compared to the effect on tuning seen in the few studies that have examined the effect of a sound-induced loss in coch- lear sensitivity on the response areas of single neurons in brainstem or midbrain nuclei.33”3*51 A common result from these studies is that TTS results in a broadening of frequency selectivity due to loss of the most sensitive regions of tuning curves. In the present analysis we have termed such changes AN-like, and have found that only two of the 17AI units tested with intense sound exposure showed this pattern of change. In the other neurons, the results suggested that TTS had produced differential effects on excit- atory and inhibitory inputs making up the response area. Study of the e&cts of TTS on AN fibers has shown that extensive exposure to intense tones (using higher level or much longer exposures than the present study) can induce hypersensitivity to low frequencies (~4 kHz)> We did not observe such effects, nor could they provide an explanation for any of the cells that showed expansion below CF at higher frequencies (e.g. Fig. 3B, D).
The interaction of inhibitory and excitatory inputs is a common characteristic of the responses of neur- ons in many subcortical auditory nuclei.2~‘6~‘7~‘9~4’~47~52~s~ It is therefore surprising that the effects of TTS on the tuning of neurons in the cochlear nucleus or in the inferior colliculus generally appear to be simple reflec- tions of the effects on AN fihers,33r43 which do not have such interactions. Willott and Lus’ attributed TTS-induced changes in the response patterns of collicular neurons to changes in the balance of excit- atory and inhibitory inputs. Although they did not discuss changes in tuning in any detail, one of their tuning curves (their Fig. 2D) shows expansion of the type we have observed in AI. Differential effects of TTS on excitatory and inhibitor inputs have also been postulated to explain TTS-induced changes in other response characteristics of s&cortical central auditory neurons (namely maximal response rates,24 single-tone inhibition of spontaneous activity43). Salvi et al.” found increases in neural response rates at CF after a pure-tone TTS aimed at

962 M. B. CALFORD ef al
frequencies half an octave above CF. They reasoned that such a TTS would have more effect on presumed inhibitor surrounds than on excitatory com~n~ts of a neuron’s response area. However, complex changes in threshold tuning curves, of the type that we have observed in AI, have generally not been reported. The failure to report such effects at subcor- tical levels may reflect the limited amount of data that has been presented on tuning curves, but it is also possible that complex effects of ITS on tuning are less common at subcortical levels.
Both the basic structure of cortex, and parallels with plasticity attributed to other sensory cortices,” give support for consideration of at least some in- volvement of processing at this level. Incoming thala- mocortical afferents to AI make only excitatory synapses. However, in addition to synapses with the large output pyramidal cells of cortex, the afferents also connect with small, inhibitory interneurons.” A general model that involves the basic elements of cortical circuitry has been presented by Shamma and Symmes4j to account for lateral inhibition seen in awake squirrel monkey auditory cortex, and similar mechanisms have been suggested to operate in so- matosensory” and visual& cortices. Basically, it is thought that the dense intemeuron network provides inhibition which limits effective responses by pyrami- dal cells through suppressing weaker inputs, thus com~n~ting for the wide arbor&&ion of incoming afferents.18*u In this way, the structure of cerebral cortex provides a basis for the interaction of excit- atory and inhibitory contributions to neural response areas.
Relationship to other evidence of plasticity in auditory and sornatosensory cortex
In addition to the present demonstration of rapid changes in response areas, changes in the response of cortical neurons to tonal stimuli have been demon- strated through a conditioning paradigm.49*s0 For sharply tuned neurons in primary auditory cortex, responses to tone pulses at best frequency defined by maximal discharge (BP) are enhanced when a con- ditioning stimulus is matched to BF. When the conditioning stimulus is at other frequencies within the response area, the neural response can be selec- tively enhanced such that the BF itself is altered. Whether this change in BF is associated with a change in threshold sensitivity, and whether response enhancement can change the frequency limits of the response area have not been examined. However, the finding that the response at some frequencies can be enhanced while at other frequencies it is suppressed indicates that this short-term plasticity involves changes in the excitato~-inhibitory balance within a neuron’s response area of the type that appear to underlie some of the effects we have described. Rapid unmasking of large receptive fields, indicating a change in excitatory and inhibitory balance, has also been demonstrated for neurons in somatosensory
cortex following various m~p~ations of their pri- mary afferents. 46*26 When the relationship between the long-term effect of a small peripheral denervation (digit ~pu~tion) and the immediate effect was specifically studied, it was found that the final recep- tive field of a multi-unit cluster in cortex was a subset of the rapidly unmasked field of the same cluster.3 Thus an initial unmasking is followed by some form of plasticity in which there is a shrinking of receptive fields and a re-establishment of topographic orderazg
The results of the present study can be compared to those obtained by Robertson and Irvine@ who made restricted cochlear lesions in adult guinea-pigs and two to three. months later examined the fre- quency organization of the rostra1 auditory field in cortex contralateral to the lesioned cochlea. The regions of cortex deprived of their normal input by the cochlear lesion were found to be wholly or partially occupied by expanded representations of adjacent frequencies, and the neurons in these regions of expanded representation had thresholds at their “new” CFs that were not significantly different from thresholds of neurons with those Cl3 in normal animals. Similar reorganization has been reported in AI of mature cats with restricted cochlear lesions.36
The time-course of reorganization following chronic lesions has not been described, and the relationship of these changes in frequency selectivity to those resulting from immediate changes such as those brought about by TTS remains uncertain. However it would appear that the degree of unmask- ing revealed in the present study is insufficient to account for the effects observed chronically after a restricted cochiear lesion. Less than half of the neur- ons showed expansion of their response areas after TTS, and for these the changes were small compared to the chronic effect. In particular, it is difficult to account for the apparently normal thresholds throughout the expanded representations of frequen- cies corresponding to the edge of the lesions36q40 without invoking some sort of neuronal plasticity beyond the unmasking resulting from a disruption of inhibition suggested by the effects of TTS on those neurons showing expansion of their response area. This contrasts with the comparison of immediate5 and long-term effects3 of small peripheral denerva- tions in the somatosensory system. However, as for the somatosensory system, some form of neuronal plasticity must be postulated to explain the long-term reorganization in auditory cortex following restricted cochlear Iesions. The involvement of subcortical st~ctures in these longer-tee effects in either system has not yet been estabIish~.
The aim of the present study was to describe the effects of an acute peripheral hearing loss (TTS), created by exposure to a loud pure tone, on the excitatory response areas of single neurons in primary

Effects of temporary threshold shift on auditory cortex 963
auditory cortex. A variety of complex effects on the response areas of cortical neurons was found to result from ‘ITS, and for the majority of neurons the effects (expansion or contraction of response areas) are quite different from those described for auditory nerve fibers. This complexity is likely to be related to mechanisms that normally determine the frequency response limits of cortical neurons. It is thought that the frequency tuning of auditory cortex neurons involves integration of excitatory and inhibitory inputs, and the present data are consistent with differential effects of ‘ITS on these components. However, classi~cation of neurons according to the presence of two forms of inhibition, lateral inhibition (judged with a forward masking paradigm) and non- monotonicity of the spike-count vs sound level func-
tion, failed to correspond with the type of change induced by ‘ITS. Although the expansion of the excitatory response area was quite extensive in some ‘neurons, thresholds at these frequencies were high. Thus, these acute effects cannot account for the near-normal thresholds seen in expanded represen- tations of frequencies bordering on those directly affected by chronic cochlear Iesions.40
Acknowledgemenrs-us work was supported by grants from the National Health and Medical Research Council of Australia and a Co~on~th Special Research Centres Award to the Vision, Touch and Hearing Research Centre. We thank Rowan Tweedale, Leah Krubitxer and Tom Yin for their comments on the manuscript, and Bob Wood and John Casseii for their technical assistance.
1.
2.
3.
4.
5.
6.
7.
8.
9.
IO.
Il.
12.
13.
14. 15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
REFERENCES
Boettcher F. A., Saivi R. J. and Saunders S. S. (19%) Recovery from short-term adaptation in single neurons in the cochiear nucleus. Hearing Res. 48, 125-144. Brownell W. E., Manis P. B. and Ritz L. A. (1979) Ipsilateral inhibitory responses in the cat lateral superior olive. Brain Res. 07, 189-193. Caiford M. B. and Tweedale R. (1988) Immediate and chronic changes in responses of somatosensory cortex in adult flying-fox after digit amputation. Naiure 332, 446448. Caiford M. B. and Tweedale R. (1990) Interhemisphe~~ transfer of plasticity in the cerebral cortex. Science 249, 805807. Calford M. 8. and Tweedaie R. (1991) Acute changes in cutaneous receptive fields in primary somatosensory cortex after digit denervation in adult flying fox. J. Neurophysioi. 65, 178-187. Calford M. B. and Tweedale R. (1991) C-fibres provide a source of masking inhibition to primary somatosensory cortex. Proc. R. Sot. Lond. B 243, 269-275. Calford M. B., Webster W. R. and Semple M. N. (1983) Measurement of frequency selectivity of single neurons in the central auditory pathway. Hearing Res. 11, 395-401. Carney L. H. (1990) Sensitivities of ceils in anteroventrai cochlear nucleus of cat to spatiotemporal discharge patterns across primary afferents. J. Neurophysiot. 64, 437-156. Cody A. R. and Johnstone B. M. (1980) Single auditory neuron response during acute acoustic trauma. Hearing Res. 3, 316. Cody A. R. and Russell I. J. (1985) Outer hair ceils in the mammalian cochlea and noise-induced hearing loss. Nature 315, 662-665. Dailos P. and Cheatham M. A. (1976) Compound action potential (AP) tuning curves. J. Acousr. Sot. Am. 59, 591-597. Dallos P. and Cheatham M. A. (1977) Analog of two-tone suppression in whole nerve responses. J. Aeoust. Sot, Am. 62, 1048-1051. Davis H., Morgan C. T., Hawkins J. E. J., Galambos R. and Smith F. W. (1950) Temporary deafness following exposure to loud tones and noise. Acta otolar., Suppl. 88, i-56. Dykes R. W. (1983) Parallel processing of somatosensory information: A theory. Brain Res. Rev. 6, 47-l 15. Dykes R. W. (1990) Acetylcholine and neuronal plasticity in somatosensory cortex. In Brain Chohnergic Systems (eds Steriade M. and Biesold D.), pp. 294-313. Oxford University Press, New York. Ehret G. and Merzenich M. M. (1989) Complex sound analysis (frequency resolution, filtering and spectral integration) by single units of the inferior colliculus of the cat. Brain. Res. Rev. 13, 139-163. Erulkar S. D. (1959) The responses of single units of the inferior colliculus of the cat to acoustic stimulation. Proc. R. Sot. Lond. B 150, 336-35.5. Garraghty P. E. and Sur M. (1990) Morphology of single intracellularly stained axons terminating in area Jb of macaque monkeys. J. camp. Neurol. 294, 583-593. Godfrey D. A., Kiang N. Y.-S. and Norris B. E. (1975) Single unit activity in the posteroventral cochlear nucleus of the cat. J. camp. Neurol. 142, 247-268. Harris D. M. and Dalios P. (1979) Forward masking of auditory nerve fiber responses. J. N~rophys~ol. 42, 1083-l 107. Houser C. R., Vaughn J. E., Hendry S. H. C., Jones E. G. and Peters A. (1984) GABA neurons in the cerebral cortex. In Cerebral Coriex, Volume 2-Functional Properties of Cortical Cells (eds Jones E. G. and Peters A.), pp. 63-89. Plenum Press, New York. Kaas J. H., Krubitzer L. A., Chino Y. M., Langston A. L., Poiley E. H. and Blair N. (19%) Reorganization of retinotopic cortical maps in adult mammals after lesions of the retina. Science M@, 229-231. Landry p. and Deschenes M. (1981) Intracortical arborizations and receptive fields of identified ventrobasal thalamocortical alferents to the primary somatic sensory cortex in the cat. J. camp. Neural. 199, 345-371. Lonsbury-Martin B. L. and Martin G. K. (1981) Effects of moderately intense sound on auditory sensitivity in rhesus monkeys: behavioral and neural observations. J. Neurophysiol. 46, 563-586. Lonsbury-Martin R. L. and Meikie M. B. (1978) Neural correlates of auditory fatigue: frequency-dependent changes in activity of single cochlear nerve fibers. J. ~europhys~o~, 41, 987-1006.

964 M. B. CALFORD et at,
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38. 39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
Merzenicb M. M., Kaas J. H., Wall J. T., Sur M., Nelson R. J. and Fellernan D. J. (1983) Progression of change following median nerve section in tbe cortical representation of the band in areas 3b and I in adult owl and squirrel monkeys. Neuroscience 10, 639-665. Merzenicb M. M., Knight P. L. and Roth G. L. (1975) Representation of cochlea within primary auditory cortex in the cat. J. Neurophysiol. 38, 231-249. Menenich M. M., Nelson R. J., Stryker M. P., Cynader M. S., Scboppmann A. and Zook J. M. (1984) Somatosensory cortical map changes following digit amputation in adult monkeys. J. camp. Neural. 224, 591-605. Patuzzi R., Jobnstone B. M. and Sellick P. M. (1984) The alteration of the vibration of the basilar membrane produced by loud sound. Hearing Res. 13,99-100. Patuzzi R. B., Yates G. K. and Johnstone B. M. (1989) Changes in cocblear micropbonic and neural sensitivity produced by acoustic trauma. Hearing Res. 39, 189-202. Phillips D. P. and Orman S. S. (1984) Responses of single neurons in posterior field of cat auditory cortex to tonaf stimulation. J. N~rophysioI. 51, 147-163. Phihips D. P., Orman S. S., Musicant A. D. and Wilson G. F. (1985) Neurons in the cat’s primary auditory cortex distinguished by their responses to tones and wide-spectrum noise. Hearing Res. 18, 73-86. Popeliir J. and Syka J. (1982) Noise impairment in the guinea pig. II. Changes of single unit responses in the inferior colliculus. Hearing Res. 8, 273-283. Popelar J., Syka J. and Ulehlova L. (1978) Effect of high-intensity sound on cochlear microphonics and activity of inferior colliculus neurons in the guinea pig. Arch. ororhinolar. 221, 115-122. Rajan R. (1988) Effect of electrical stimulation of the crossed obvococblear bundle on temporary threshold shifts in auditory sensitivity. I. Dependence on electrical stimulation parameters. J. Neurophysiol. 60, 549-568. Rajan R., Irvine D. R. F., Calford M. B. and Wise L. 2. (1992) Effect of frequency-specific losses in cochlear neural sensitivity in the processing and representation of frequency in primary auditory cortex. In Noise-Induced Hearing Loss (eds Dancer A. L., Henderson D., Salvi R. J. and Hamernik R. P.), pp. 119-129. Mosby-Year Book, St Louis. Rajan R., Irvine D. R. F. and CasseH .I. (1991) Normative NI audiogram data for tbe ~rbiturate-anaesthetis~ domestic cat. Hearing Rex 53, 153-158. Reale R. A. and Imig T. J. (1980) Tonotopic organi~tion in auditory cortex of the cat. J. camp. Neural. 192,265-291. Rhode W. S. (1976) A digital system for auditory neuropbysiolo~cal research. In Currenr Computer TechnoZogy in N~~o~io~ogy (ed. Brown P.), pp. 543-567. Hemisphere, Washington, D.C. Robertson D. and Irvine D. R. F. (1989) Plasticity of frequency organization in auditory cortex of guinea pigs with partial unilateral deafness. J. camp. Neural. t82, 456-471. Ryan A. and Miller J. (1978) Single unit responses in the inferior colbculus of the awake and performing rhesus monkey. Expi Brain Res. 32, 389407. Salvi R. J. (1976) Central components of the temporary threshold shift. In EJjrects of Noise on Hearing (eds Henderson D., Hamernik P., Dosanjb D. S. and Mills J. H.), pp. 247-262. Raven Press, New York. Salvi R. J., Hamernik R. P. and Henderson D. (1978) Discharge patterns in the cochlear nucleus of the chinchilla following noise induced asvmntotic threshold shift. Exn! Brain Res. 32. 301-320. Salvi R.-J., Powers N. L., Saunders S. S., Boettcher F.-A. and Clock A. E. (1992) Enhancement of evoked response amplitude and single unit activity after noise exposure. In Noise-Induced Hearing Loss (eds Dancer A. L., Henderson D., Safvi R. J. and Hamemik R. P.), pp. 156-174. Mosby-Year Book, St Louis. Shamma S. A. and Symmes D. (1985) Patterns of inhibition in auditory cortical cells in awake souirrel monkevs. Hearing Ref. 19, I-Ii. Silbto A. M. (1977) inhibitory processes underlying the directional specificity of simple, complex and hy~rcomp~ex cells in the cat’s visual cortex. J. Phvsiol., Land. 271. 699-720. van Gishergen J. A. M., Grasbuis J: L., Johannesma P. I. M. and Vendrik A. J. H. (1975) Spectral and temporal characteristics of activation and suppression of units in the eocblear nuclei of tbe anaesthetized cat. Expr Brain Res. 23, 367-386. Volkov I. 0. and Galazjuk A. V. (1991) Formation of spike response to sound tones in cat auditory cortex neurons: interaction of excitatory and inhibitory effects. NeuroscKnce 43, 307-321. Weinherger N. M., Ashe J. H., Metberate R., McKenna T. M., Diamond D. M. and Bakin J. (1990) Retuning auditory cortex by learning: a preliminary model of receptive field plasticity. Concepts Neurosci. 1, 91-132. Weinherger N. M. and Diamond D. M. (1987) Physiological plasticity in auditory cortex: rapid induction by learning. Prog. Neurobiol. 29, l-55. Willott J. F. and Lu S. (1982) Noise-induced bearing loss can alter neural coding and increase excitability in tbe central nervous system. Science 214 1331-1332. Willott J. F. and Urban G. P. (1978) Response properties of neurons in nuclei of the mouse inferior collicuius. J. camp. P?rysioZ. 127, 175-I 84. Wise L. Z. and Irvine D. R. F. (1983) Auditory response properties of neurons in deep layers of cat superior colbcuhrs. J. Neurophysiol. 49, 674-685. Wise L. Z. and Irvine D. R. F. (1985) Topographic organization of interaural intensity difference sensitivity in deep layers of cat superior cohicuhts: impbcations for auditory spatial representation. J. Neurophysiof. 54, 185-211. Young E. D. and Brownell W. E. (1976) Responses to tones and noise of single cells in dorsal cochlear nucteus of unanesthetized cats. J. Neurophysiol. 39, 282-300.
(Accepted 26 February 1993)