Embed Size (px)
Citation preview

1

2
PROFORMA – I
PROFORMA FOR SUBMISSION OF PROJECT PROPOSALS ON RESEARCH ANDDEVELOPMENT, PROGRAMME SUPPORT
(To be filled by the applicant)
PART I: GENERAL INFORMATION1. Name of the Institute/University/Organisation submitting the Project Proposal:
BANARAS HINDU UNIVERSITY
2. State: UTTAR PRADESH
3. Status of the Institute: CENTRAL UNIVERSITY(Please see Annexure-I)
4. Name and designation of the Executive Authority of the Institute/University forwarding theapplication: Registrar, Banaras Hindu University
5. Project Title:. Understanding molecular cross talk between Parkin, p53 and JNKmediated Apoptosis in Neurodegenerative Parkinson’s Disease Model of Drosophila
6. Category of the Project (Please tick):.R&D/ Programme Support: Pilot Proposal
7. Specific Area (Please see Annexure - II): Neurosciences & Neurobiology
8. Duration: 2…Years. 0…. Months
9. Total Cost (Rs.) 20,10,400 (Twenty Lakh Ten thousand Four hundred only, excludingover heads)
10. Is the project Single Institutional or Multiple-Institutional (S/M)? : Single
11. If the project is multi-institutional, please furnish the following: NA
12. Scope of application indicating anticipated product and processes
The outcome of the project will help understanding the molecular basis of Parkinson’s disease,
Alzheimer’s disease as well as other Neurodegenerative Diseases particularly in relation to JNK
mediated regulation of apoptosis and /or cell survival signaled by Parkin and p53 candidates.
This would lead to identification of specific and potential therapeutic targets and hence results in
formulating effective therapeutic strategies in future. The study also reveal the connecting link
between Sporadic as well as Genetic pathways of the disease and help understanding common
molecular candidates, which will help designing better drug targets in future. Assessment of p53
role in this study not only benefits Neurodegenerative diseases in particular, but also would
throw light on key regulatory role of these candidates in Cancer Biology in general.

3
13. Project Summary (Not to exceed one page. Please use separate sheet).
Parkinson's disease, a prevalent neurodegenerative disease, is characterized by the reduction of
dopaminergic neurons resulting in the loss of motor control, resting tremor, the formation of
neuronal inclusions and ultimately leading to premature cell death. Parkin, an E3 ubiquitin
ligase, is the most prevalent genetic factor in the onset of Autosomal Recessive Juvenile
Parkinsonism (AR-JP) as mutations in this gene and associated autosomal recessively inherited
genes, namely PINK1 and DJ-1, typically lead to early onset of Parkinsonism. Parkinson’s
disease (PD) typically present in sporadic fashion, but the identification of disease-causing
mutations in PD genes has provided crucial insight into the pathogenesis of this disorder.
However, the molecular mechanisms underlying this neuron loss remain elusive. Mitochondrial
dysfunction caused by oxidative stress and genetic defects have been implicated in the loss of
dopaminergic neurons in Parkinson’s disease. However, the key molecular events that provoke
neurodegeneration still remain poorly understood.
Extensive work has been done to delineate the cellular mechanisms responsible for the
exacerbated cell death of dopaminergic neurons observed in PD. Overall, the oncogene p53 has
been identified as a key effector protein. The clues linking p53 to the etiology of PD and the
evidences that this protein may be at the center of multiple signaling cascades not only altered by
mutations of various proteins responsible for familial cases of PD but also in general sporadic
cases of this devastating disease. Interestingly, activation of c-Jun N-terminal Kinase (JNK) has
also been implicated in neuronal cell death in Parkinson’s disease (PD). Moreover, Drosophila
models for AR-JP, loss of function mutants of parkin, also show dopaminergic neural
degeneration associated with hyper activation of JNK, increased apoptosis along with
mitochondrial defects. However, the molecular mechanism by which Parkin protects cells from
apoptosis remains unclear.
We are interested in understanding the molecular mechanisms that regulate neuronal and non
neuronal cell death processes. In particular, we will focus on the role of Parkin, p53 and JNK
candidates in cell death processes. Our study mainly aimed at understanding Genetic and
Molecular interactions between these candidates at RNA and Protein levels in neuronal and
nonneuronal tissues using tissue specific GAL4 mediated UAS-RNAi constructs, deficiency/over
expressed lines and also gene specific transposable mutant alleles. We are also interested to fish
out novel interactors of Parkin and p53. We propose to examine the exact role of Parkin and p53
in mitochondrial dysfunction through altered fusion and fission dynamics in PD models.

4
PART II: PARTICULARS OF INVESTIGATORS
(One or more co-investigators are preferred in every project. Inclusion of co-investigator(s) ismandatory for investigators retiring before completion of the project)
Principal Investigator:
14. Name:.Dr. S. Srikrishna
Date of Birth: 12-08-1973 Sex (M/F): Male
Designation:.Assistant Professor
Department: Biochemistry
Institute/University:.BANARAS HINDU UNIVERSITY
Address:. Department of Biochemistry,
Faculty of Science,
Banaras Hindu University,
Varanasi, Uttar Pradesh, India
PIN:. 221 005
Telephone: +91-542-2575780; Fax:.+91-542-2368457
E-mail: [email protected]; [email protected]
Number of research projects being handled at present: Nil
Co-Investigator
15. Name: Dr. Surya Pratap Singh
Date of Birth: 23-07-1962 Sex (M/F): Male
Designation: Associate Professor
Department: Biochemistry Department
Institute/University: Banaras Hindu University
Address: Department of Biochemistry, Faculty of Science, Banaras Hindu University,Varanasi PIN: 221 005
Telephone: +91-542-2570027 Fax: 91-0542-2368174 E-mail: [email protected] of Research projects being handled at present: Nil
Note : Use separate page, if more investigators are involved

5
PART III : TECHNICAL DETAILS OF PROJECT(Under the following heads on separate sheets)
16. Introduction (not to exceed 2 pages or 1000 words)
16.1 Origin of the proposal
The main question underlying this proposal is to understand the molecular link betweensporadic and genetic forms of the Parkinson’s disease (PD). How exactly the twopathways differ in their disease pathology? The end result of the Neuronal cell death inboth the cases is apparently due to loss of mitochondrial function. In sporadic cases it ismainly due to increased ROS production while in familial cases it is through JNKmediated apoptosis due to alterations of genetic factors, like Parkin, PINK1, DJ and p53,etc. But, how these processes are leading to cell death is not well understood? Recentevidences suggest the involvement of p53 in JNK mediated apoptosis in Parkinson’sdisease. How exactly these candidates interact with each other in both sporadic andfamilial form s of the Parkinson’s disease is the main focus of this proposal. Besideschemically induced Parkinson’s disease, we will use neuronal tissue specific UAS-RNAifly constructs (P {TRiP.JF01200} attP2 constructs, Transgenic RNAi Project, Ni et al,2008) specific to parkin, p53 and JNK pathway genes and drive them with specificGAL4 lines to generate familial forms of the Parkinson’s disease.
Parkinson’s disease, a progressive neurodegenerative movement disorder, is the secondmost common neurodegenerative disease after Alzheimer disease. Even though PD islargely sporadic disorder, about 15% of cases are of inherited in nature, and many ofthese familial forms share common features with sporadic form of PD. Althoughpotential candidate genes having role in PD were identified, the underlying mechanisminvolving these genes in PD is still not clearly understood due to which proper andeffective cure of the disease is far from reality. The exact mechanisms of orchestratedneuronal cell death, apoptosis, in PD are unknown, but recent studies suggest anantagonistic role between p53 and Parkin in Neuronal cell death or survival processes.But the genetic control of these molecules and the molecular interactors are not wellunderstood in this disease context. We will examine molecular cross talk between theseinteresting candidates in the present project.
16.2 (a) Rationale of the study supported by cited literature (b) Hypothesis (c) Keyquestions.
(a) Rationale of the study supported by cited literature:
Parkinson’s disease being the most frequent neurodegenerative movement disordermay be considered as multi-system disorder that severely damages predisposed celltypes in restricted regions of the nervous system in human (Meco et al, 2008). Overthe past decade, studies reveal that mutations in genes encoding alpha synuclein andLeucine Rich Repeat kinase-2 (LRRK-2) mediate autosomal dominant forms of PD,while genes encoding parkin, DJ-1 and PTEN induced kinase-1 (PINK-1) mediateautosomal recessive forms (Wood-Kaczmar et al, 2006; Venderova et al, 2009).Although, Familial form of PD accounts for 5-15% of all cases due to mutations inaforementioned genes, studies done have reveal the involvement of these genes inSporadic forms of PD (Wood-Kaczmar et al, 2006).

6
The combination of Mitochondrial dysfunction, oxidative stress, sexual dysfunctionand aberrant protein inclusions are implicated in PD pathogenesis (Meco et al, 2008;Banerjee et al, 2009; Saini et al, 2010). In both familial and sporadic form,mitochondrial impairment involving electron transport chain, impaired calciumbuffering, and anomalies in mitochondrial morphology and dynamics occurs (Park etal, 2009). Whether mitochondrial dysfunctioning is cause or effect of PDpathogenesis, is not clearly understood (Banerjee et al, 2009). PD instead of being auniform disease, it comprises a numerous familiar and spontaneous motor disorderscaused by different genetic and environmental factors initiating sequence of eventsconverging in a common pathological condition, involving dopaminergic neuron lossin an age dependent manner (Botella et al, 2009). It’s worth mentioning that to datethere is no cure for PD, and not even the etiopathology of PD is resolved (Banerjee etal, 2009).
The recent identification of genes involved in PD has enthused the hunt forunderstanding the mechanisms behind familial PD pathology, and this would alsohelp in mechanisms leading to sporadic PD pathology. Since past few years, severalstudies have been done with a focus on identifying functions of aforementioned geneproducts and many possible pathogenic mechanisms, including protein misfolding,aberrant protein accumulation, oxidative stress, mitochondrial dysfunctioning andcaspase activation (reviewed in Da Costa and Checler, 2011). In an attempt toidentify the etiology of PD, promising clues have been attained from studies ofanimal models for PD, including mice, rats, flies and worms. It’s worth mentioningthat use of animal models for human disease is of immense significance as itrepresents a very valuable prospect not only to test working hypotheses but also toidentify new information that could be of great significance in terms of diagnosis,drug design and therapeutics for such diseases like PD. The use of Drosophila as amodel organism to study PD presents a number of advantages including small numberof neuronal and glial cells in central nervous system as compared to vertebrates, sharesimilar neurotransmitter systems like GABA, glutamate, Dopamine, etc., conservedgenes and signal transduction pathways between human and flies. Moreover, most ofthe genes involve in PD in humans have counterparts in Drosophila (reviewed inSoriano and Paricio, 2011).
Parkin gene encode Ubiquitin E3 ligase, which is highly conserved betweenvertebrates and invertebrates (Botella et al, 2009), apparently modulate mitochondrialmorphogenesis during spermiogenesis (Riparbelli and Callaini, 2007) and it enhancesmitochondrial biogenesis in proliferating cells through transcription and replication ofmitochondrial DNA (Kuroda et al, 2006), mitochondrial remodeling (fission-fusion)and Mitophagy (Narendra et al, 2008), transcription regulator (Da Costa et al, 2009),and also have been implicated to have a role in fertility (Riparbelli and Callaini,2007). Interestingly, Parkin has been shown to negatively regulate JNK pathway indopaminergic neurons in Drosophila (Cha et al, 2005; reviewed in Soriano andParicio, 2011). It also suppresses c-Jun N-terminal Kinase induced cell death viatranscriptional regulation in Drosophila and AR-JP (Autosomal Recessive JuvenileParkinson’s) models of drosophila show dopaminergic neural degeneration associatedwith hyper activation of JNK, increased apoptosis and Mitochondrial defects (Hwanget al, 2010). Hence it is interesting to see the role of p53 in UAS-parkin, parkinRNAi
and Parkin status in UAS-p53, p53RNAi genetic backgrounds as it is not clear howParkin, p53 regulate JNK activity and Mitochondrial dysfunction in neuronal cells.

7
(b) Hypothesis
Sporadic and familial forms of the PD leading to cell death, which is mainly due toaltered mitochondrial fission/fusion dynamics. How these sequential alterations arecaused by these forms of the disease is least understood and different laboratoriesacross the world focused on mitochondrial dysfunction during PD pathogenesis.Interestingly, recent studies shed light on JNK mediated cell death pathways. But howexactly this pathway involved in selective neuronal cell death process is not clear.However, it is important to note that the p53 is emerging as key candidate interactorof the Parkin, which is required for the cell survival. Interestingly, loss of Parkinleads to upregulation of p53 also pointing towards the key role played by these twocandidates in PD. Hence, thorough understanding of the interactive role played thesetwo candidates in relation to JNK pathway offer significant cues to identify specifictherapeutic targets in future.
(c) Key questions
1. How Sporadic and Familial forms of PD differ in their mechanism of pathogenesis?Are there any common mechanisms linking these two pathways?
2. How JNK pathway regulates the survival or death decisions in Neuronal cells?
3. Are p53 and Parkin directly/indirectly interacting with each other to regulate the JNKmediated cell survival/death processes?
4. How Parkin and p53 affect the fission and fusion dynamics of mitochondria inDopaminergic neurons as well as in other tissues?
5. What are the common interactors of p53 and Parkin in PD cells?
16.5 Current status of research and development in the subject (both international and nationalstatus)
International status:
PD is a debilitating neurodegenerative disease with no cure and unknown etiology. It wasreported that the number of individuals with Parkinson’s disease in 15 countries (France,Spain, Germany, the United Kingdom, China, India, Indonesia, the United States, Brazil,Pakistan, Bangladesh, Nigeria, Japan, Russia and Italy) will grow from 4.1 to 8.7 millionby the year 2030, with an estimated growth of almost double by next generation indeveloping countries of Asia (Dorsey et al, 2005).
Over the past few years’ studies have been done using animal models to identify theunderlying mechanisms involved in PD using gene specific mutation(s) in Parkin,PINK1, DJ-1, LRRK2, and α-synuclein. Parkin gene mutations serve as the mostcommon cause of juvenile onset PD along with delayed onset cases (Lucking et al, 2000& Farrer et al, 2001). Parkin gene product is an ubiquitin E3 ligase whose function is to

8
mediate polyubiquitination in order to remove aggregation of prone substrates forproteasomal degradation, loss of parkin E3 ligase function has been shown to result inaccumulation of toxic substrates leading to autosomal form of PD (Dawson, 2006). Morerecent studies suggest the role of parkin in mediating monoubiquitination in addition toparkin role in proteasomal machinery (Moore, 2006). Such role is known to mediatefunctions involved in protein trafficking, transcriptional regulation and neuroprotectivesignalling (Fallon et al, 2006; Mukhopadhyay and Reizman, 2007). Various studies donein recent past suggest possible potential role of parkin in mitochondrial membranedepolarization, through proteasomal activity (Yoshii et al, 2011). Although, Parkin isexpressed ubiquitously, studies using sub-cellular fractionation demonstrate associationof parkin with outer mitochondrial membrane (Stichel et al, 2000 & Darios et al, 2003),suggesting a potential role in mitochondrial functioning. Over expression of parkinresulted in transcription and replication of mitochondrial DNA (mtDNA) in proliferatingcells, while knockdown of parkin inhibited this function (Banerjee et al, 2009).Transcription of mtDNA in proliferating cells is consequence of functional association ofParkin with mitochondrial transcription factor A (TFAM), thus suggesting role of Parkinin mitochondrial transcription and replication (Banerjee et al, 2009). Gene knockout ofparkin in flies and mouse show striking mitochondrial defects (Pesah et al, 2004;Palacino et al, 2004), involving swollen mitochondria, fragmented cristae, decrease in theabundance of protein subunits of the mitochondrial electron transport chain complex I &IV , decrease in ATP production (Pesah et al, 2004; Palacino et al, 2004; Yong-Kee et al,2011). Similarly, loss of Parkin function in Drosophila also shows dopaminergicneurodegeneration, reduced lifespan, enhanced flight muscle degeneration and malesterility (Whitworth et al., 2006; Moore et al, 2008). Earlier studies show that the Parkinplays an antioxidative role in maintaining mitochondria (Whitworth et al, 2005; Yang etal, 2007 & Trinh et al, 2008). The activation of JNK signaling pathway in dopaminergicneurons of parkin mutant flies results in shrinkage of dopaminergic neurons withdecreased tyrosine hydroxylase level, while parkin with normal functioning negativelyregulates JNK signaling pathway (Cha et al, 2005), suggesting possible and importantrole of Parkin in down regulation of JNK mediated apoptosis. JNK role in apoptosis iswell defined while exact mechanism involved seems to be either stimulus based or tissuespecific. Sequential regulation of JNK activation is critical to cellular responses.Interestingly, early transient JNK activation promotes cell survival while prolonged JNKactivation mediates apoptosis (Weston et al, 2007). Drosophila model for AR-JP withparkin gene mutation shows Dopaminergic neuron loss associated with hyperactivationof JNK with increased apoptosis. In contrast, Drosophila Parkin was shown to suppressJNK signaling pathway in developing tissues via transcriptional regulation of Basket(Hwang et al, 2010). The eye degeneration phenotype with apoptosis in eye imaginal discinduced by Hemipterous (Hep), a Drosophila JNK kinase, is also suppressed by ectopicexpression of parkin. On the other hand over expression of p53 positively regulatesHemipterous to activate apoptosis. Interestingly, one of the recent studies implicatedParkin as a transcriptional regulator as it’s endogenous over expression resulted inphysical interaction of Parkin with p53 promoter to down regulate p53 expression asevidenced by decreased levels of p53 mRNA (reviewed by Da Costa et al, 2010). Theseexperimental evidences strongly suggest molecular cross talk between Parkin, p53 andJNK to regulate apoptosis and cell survival. The objectives of the present project,therefore, aimed at understanding the mechanism of Parkin, p53 and JNK mediatedapoptosis in Neuronal and non-neuronal cells, as molecular mechanisms behind this deathpathway remains unclear till date.

9
National status:
There is growing number of research groups working on vertebrate models ofParkinson’s disease (PD), though there is a lack of uniform, large scale, nationwideepidemiological data on the incidence or prevalence of PD in India. Earlier studies onepidemiology and treatment of Parkinson’s disease in India have reported PD prevalencebut epidemiological and clinical data appears to be insufficient and crude (Singhal et al,2003). However, Dr. B. K. Thelma’s group at Department of Genetics, University ofDelhi is systematically working on Mutation frequency of parkin in Familial andSporadic Parkinson’s disease. Studies carried out by this group have shown parkin genemutation frequency of 8.5% among Indian patients with Parkinson’s disease (Chaudharyet al, 2006). Dr. Jharna Ray’s group at “S.N. Pradhan centre for Neuroscience” inUniversity of Kolkata is working in an area of molecular pathogenesis in Parkinson’sdisease, where they are trying to identify mutations in parkin gene in Indian parients.Among Eastern Indian population studied, 7.24% cases of Parkinson are associated withmutations in parkin gene (Biswas et al, 2006). Profound deficit in dopamine content, lowglutathione and elevated lipid peroxidation in dopaminergic neurons results in oxidativestress, mitochondrial dysfunction and apoptosis. Dr. Pravat K Mandal atNeurospectroscopy/ Neuroimaging laboratory, NBRC (National Brain Research Centre)is actively engaged in Brain imaging and Neurochemical analysis of Parkinson’s andother Neurodegenerative diseases using in vivo Magnetic Resonance Spectroscopy(Mandal, 2011). Dr. Bimal Nehru’s lab at Department of Biophysics at Panjab Universityis focused on rotenone induced induced oxidative stress in an animal models ofParkinson’s disease. Studies on rodents shown that chronic administration of rotenoneresulted in significant oxidative damage in the midbrain region in contrary to otherregions of the brain. Centrophenoxine was used as potential anti oxidant therapeutics andit diminished most of the changes brought about due to rotenone usage (Verma andNehru, 2009). Dr. Akshay Anand’s research group at Department of Neurology,PGIMER, Chandigarh, working on Genetic screening of PARK2 mutations andreduced Parkin expression conferring risk for Parkinsonism in North West India(Vinish et al, 2009). While Dr. Sasanka Chakrabarti at Institute of Post-graduate Medicaleducation Research, Kolkata and Dr. Birija S Patro at BARC, Mumbi are focused atmitochondrial Dysfunction and Pathogenesis of Parkinson’s Disease. They have shownthat predominant involvement of quinone oxidation products in cytotoxic effects ofdopamine and the mitochondrial impairment in Parkinson’s Pathogenesis (Jana et al,2011). Further, Dr. Bharat’s group at NIMHANS, Bangalore, is pursuingNeuroprotection against Nitrosative stress and damaged mitochondrial membranepotential and their implications for Parkinson’s disease (Mythri et al, 2010).Mitochondria seem to be central key target involved in PD etiopathology. The studyfocusing the oxidative stress implemented due to absence or non functional parkin geneproduct and its interactor (s) will perhaps provide a better insight towards its role in PDetiopathology in the context of mitochondrial dysfunction. While majority are working onvertebrate models, very few laboratories are working on Invertebrate models ofParkinson’s disease in India. Drosophila offers an excellent Invertebrate Genetic ModelSystem to study Neurological disorders as detailed in international status above and alsodue to the fact that ~ 70% of its genome is homologous to Humans. Moreover, Humanorthologues for parkin and its interactor genes are present in drosophila. Initial studiesdone in our lab on parkin mutant flies have shown reduced longevity, diminished flightability, infertility both in male as well as in female when compared with their wild typecounterparts (Saurabh Srivastava and Srikrishna S, Unpublished results). The GAL4 and

10
UAS-RNAi based down regulation of parkin resulted in Neuronal cell death leading toapoptotic eye phenotypes in parkinRNAi flies (Saurabh Srivastava and Srikrishna S,unpublished results). Further studies with different genetic combinations will help inunderstanding mechanisms involved in disease progression and to workout theNeuroprotective mechanisms in future.
References:
Banerjee R, Starkov AA, Beal M.F, Thomas B (2009) Mitochondrial dysfunction in thelimelight of Parkinson’s disease pathogenesis. Biochemica et Biophysica Acta, 1792, 651-663.
Biswas A, Gupta A, Naiya T, Das G, Neogi R, Datta S, Mukherjee S, Das SK, Ray K, Ray J(2006) Molecular pathogenesis of Parkinson’s disease: Identification of mutations in theParkin gene in Indian patients. Parkinsonism and Related Disorders. 12, 420–426.
Botella JA, Bayersdorfer F, Gmeiner F, and Schneuwly S (2009) Modelling Parkinson’sdisease in Drosophila.Neurmol Med, 11, 268-280,.
Cha GH, Kim S, Park J et al (2005) “Parkin negatively regulates JNK pathway in thedopaminergic neurons of Drosophila,” Proceedings of the National Academy of Sciences ofthe United States of America, 102, 10345–10350.
Cha GH, Kim S, Park J, Lee E, kim M, Lee SB., Kim JM., Chung J, and Cho KS (2005)Parkin negatively regulates JNk pathway in the dopaminergic neurons of Drosophila. Proc.Natl. Acad. Sci.USA, 102, 10345-10350.
Chaudhary S, Behari M, Dihana M, Swaminath PV, Govindappa ST, Jayaram S, Goyal V,Maitrae A, Muthane UB, Juyal RC, and Thelmaa BK (2006) Parkin mutations in familialand sporadic Parkinson’s disease among Indians. Parkinsonism and Related Disorders. 12,239–245.
Da Costa CA, and Checler F (2010) Apoptosis in Parkinson's disease: Is p53 the missinglink between genetic and sporadic Parkinsonism? Cellular Signalling, 23, 963-968.
Da costa CS, Sunyach C, Giaime E, West A, Corti O, Brice A, Safe S, Abou-Sleiman PM,Wood NM, Takahashi H, Goldberg MS, Shen J, and Checler F (2009) Transcriptionalrepression of p53 by parkin and impairment by mutations associated with autosomalrecessive juvenile Parkinson’s disease. Nat Cell Biol, 11, 1370-1375.
Darios F, Corti O, Lucking CB, Hampe C, Muriel MP, Abbas N, Gu WJ, Hirsch EC, RooneyT, Ruberg M, and Brice A (2003) Parkin prevents mitochondrial swelling and cytochrome crelease in mitochondria dependent cell death. Human Molecular Genetics, 12, 517-526.
Dawson TM (2006) Parkin and defective ubiquitination in Parkinson’s disease. J. Neural.Transm. Suppl, 209-213.
Dorsey ER, Constantinescu R, Thompson JP, Biglan KM, and Tanner CM (2007) Projectednumber of people with Parkinson disease in the most populous nations, 2005 through 2030.Neurology, 68, 384-386.

11
Fallon L, Belanger CM, Corera AT, Kontogiannea M, Regan-Klaoisz E, Moreau F,Voortman J, Haber M, Rouleau G, Thorarinsdottir T, Brice A, VanBergen En HenegouwenPM., and Fon EA (2006) Aregulated interaction with UIM protein Eps15 implicates parkin inEGF receptor trafficking and PI)3(K-Akt signalling. Nat. Cell Biol, 8, 834-842.
Farrer M, Chan P, Chen R, Tan L, Lincoln S, Hernandez D, Forno L, Gwinnhardy K,Petrucelli L, Hussey J, Singleton A, Tanner C, Hardy J, and Longston JW (2001) Lewybodies and Parkinsonism in families with parkin mutations. Ann.Neurol, 50, 293-300.
Hwang S, Kim D, Choi G et al (2010) Parkin Suppresses c-Jun N-terminal Kinase inducedcell death via transcriptional regulation in Drosophila. Molecules and Cells, 29, 575-580.
Hwang S, Kim D, Choi G, Woo An S, Hong YK, Suh YS, Lee MJ, and Cho KS (2010)Parkin suppresses c-Jun N-Terminal kinase-induced cell death via Transcriptional Regulationin Drosophila.Mol. Cells, 29, 575-580.
Jana S, Sinha M, Chanda D, Roy T, Banerjee K, Munshi S, Patro BS and Chakrabarti S(2011) Mitochondrial dysfunction mediated by quinone oxidation products of dopamine:Implications in dopamine cytotoxicity and pathogenesis of Parkinson's disease. Biochimica etBiophysica Acta, 1812, 663–673.
Kuroda Y, Mitsui T, Kunishige M, Shono M, Akaike M, Azuma H, and Matsumoto T(2006) Parkin enhances mitochondrial biogenesis in proliferating cells. Human MolecularGenetics, 15, 883-895.
Lucking CB, Durr A, Bonifati V, Vaughan J, DeMichele G, Gasser T, Harhangi BS, MecoG, Denefle P, Wood NW, Agid Y, and Brice A (2000) Association between early-onsetParkinson’s disease and mutations in the parkin gene. N.Engl. J. Med, 342, 1560-1567.
Mandal PK (2011) In vivo proton magnetic resonance spectroscopic signal processing forthe absolute quantitation of brain metabolites Eur J Radiol. doi:10.1016/j.ejrad.2011.03.07
Meco G, Rubino A, Caravona N and Valente M (2008) Sexual dysfunction in Parkinson’sdisease. Parkinsonism and Related Disorder, 14, 451-456.
Moore DJ and Dawson TM (2008) Value of genetic models in understanding the cause andmechanisms of Parkinson’s disease. Curr.Neurol.Neurosci.Rep, 8, 288-296.
Moore DJ (2006) Parkin: a multifaceted ubiquitin ligase. Biochem.Soc.Trans, 34, 749-753.
Mukhopadhyay D and Riezman H (2007) Proteasome-independent functions of ubiquitin inendocytosis and signalling. Science, 315, 201-205.
Mythri RB, Harish G, Dubey SK, Misra K, and Srinivas Bharath MM (2010) Glutamoyldiester of the dietary polyphenol curcumin offer improved protection against peroxynitrite-mediated nitrosative stress and damage of brain mitochondria in vitro: implications forParkinson’s disease. Mol Cell Biochem, 347, 135-143.Narendra D, Tanaka A, Suen DF and Youle RJ (2008) Parkin is recruited selectively toimpaired mitochondria and promotes their autophagy. J. Cell Biol, 183, 795 – 803.

12
Ni, JQ, Markstein M, Binari R, Pfeiffer B, Liu LP, Villalta C, Booker M, Perkins L,Perrimon N (2008) Vector and parameters for targeted transgenic RNA interference inDrosophila melanogaster. Nat. Methods 5(1): 49-51.
Palacino JJ, Sagi D, Goldberg MS, Krauss S, Motz C, Wacker M, Klose J, and Shen J(2004) Mitochondrial dysfunction and oxidative damage in parkin deficient mice.J.Biol.Chem, 279, 18614-18622.
Park J, Kim Y and Chung J (2009) Mitochondrial dysfunction and parkinson’s diseasegenes: insights from Drosophila. Disease Models & Mechanisms, 2, 336-340.
Pesah Y, Pham T, Burgess H, Middlebrooks B, Verstreken P, Zhou Y, Harding M, Bellen H,and Mardon G (2004) Drosophila Parkin mutants have decreased mass and cell size andincreased sensitivity to oxygen radical stress. Development, 131, 2183-2194.
Riparbelli MG and Callaini G, (2007) “The Drosophila parkin homologue is required fornormal mitochondrial dynamics during spermiogenesis,” Developmental Biology, vol. 303,108–120.
Riparbelli MG, and Callaini G (2007) The Drosophila parkin homologue is required fornormal mitochondrial dynamics during spermiogenesis. Dev. Bio, 303, 108-120.
Saini N, Oelhafen S, Hua H, Georgiev O, Schaffner W, and Bueler H (2010) Extended lifespan of Drosophila parkin mutants through sequestration of redox-active metals andenhancement of anti-oxidative pathways. Neurobiol. Dis, 2155, 1-11.
Singhal B, Lalkaka J and Sankhlab C (2003) Epidemiology and treatment of Parkinson’sdisease in India. Parkinsonism and Related Disorders. 9, S105-S109.
Soriano VM and Paricio N (2011) Drosophila models of Parkinson’s disease: discoveringrelevant pathways and novel therapeutic strategies. Parkinsons Dis, Article ID 520640, DOI:10.4061/2011/520640
Stichel CC, Augustin M, Kuhn K, Zhu X.R., Engels P, Ullmer C, and Lubbert H (2000)Parkin expression in the adult mouse brain. Eur. J. Neurosci, 12, 4181-4194.
Trinh K, Moore K, Wes PD, Muchowski PJ, Dey J, Andrews L and Pallanck LJ (2008)Induction of the phase II detoxification pathway suppresses neuron loss in Drosophilamodels of Parkinson’s disease. J. Neurosci, 28, 465-472.
Venderova K, Kabbach G, Abdel-Messih E et al (2009) Leucine rich repeat kinase-2interacts with Parkin, DJ-1 and PINK-1 in Drosophila melanogaster model of Parkinson’sdisease. Human Molecular Genetics, 18, 4390-4404.Verma R and Nehru B (2009) Effect of centrophenoxine against rotenone-induced oxidativestress in an animal model of Parkinson’s disease. Neurochemistry International, 55, 369–375.
Vinish M, Prabhakar S, Khullar M, Verma I and Anand A (2009) Genetic screeningreveals high frequency of PARK2 mutations and reduced Parkin expression conferringrisk for Parkinsonism in North West India. J Neurol Neurosurg Psychiatry. 81, 166-170.

13
Weston CR and Davis RJ (2007) The JNK signal transduction pathway. Current Opinion inCell Biology, 19, 142–149.
Whitworth AJ, Theodore DA, Greene JC, Benes H, Wes PD and Pallanck LJ (2005)Increased glutathione s-transferase activity rescues dopaminergic neuron loss in Drosophilamodels of Parkinson’s disease. Proc. Natl. Acad. Sci.USA, 102, 8024-8029.
Withworth AJ, Wes PD, Pallanck LJ (2006) Drosophila models pioneer a new approach todrug discovery for parkinson’s Disease. Drug Discov. Today, 11, 119-126.
Wood-kaczmar A, Gandhi S, and Wood NW (2006) Understanding the molecular causes ofParkinson’s disease. Trends Mol. Med, 12, 521-528.
Yang H, Zhou HY, Li B, Niu GZ and Chen SD (2007) Downregulation of parkin damagesantioxidant defences and enhances proteasome inhibition-induced toxicity in PC 12 cells. J.Neuroimmune Pharmacol, 2, 276-283.
Yong-Kee CJ, Sidorova E, Hanif A, Perera G and Nash JE (2011) MitochondrialDysfunction Precedes Other Sub-CellularAbnormalities in an In Vitro Model Linked withCell Death in Parkinson’s Disease. Neurotox Res, DOI 10.1007/s12640-011-9259-6.
Yoshii SR, Kishi C, Ishihara N, and Mizushima N (2011) Parkin mediates proteasome-dependent protein degradation and rupture of the outer mitochondrial membrane. JBC,M110.209338.
16.6 The relevance and expected outcome of the proposed study
The work is unique for the following reasons:
1) Most of the work done in this area so far is either focused at sporadic or familialseparately and rarely one can find connecting studies between sporadic and familialaspects together. Hence the proposed work can lead to better understanding of theinterrelation between sporadic and familial disease mechanisms, which will help toworkout specific therapeutic targets.
2) JNK signaling is a key regulatory process for cell survival or death. Current literatureshow very few studies related to this signaling processes particularly in the context ofPD and in Neurodegenerative diseases in general. Hence, understanding the exact roleplayed by Parkin and p53 in relation to JNK pathway will through light on key eventsleading to progressive neuronal cell death in Parkinson’s disease. This also helps inscreening and designing suitable therapeutic strategies.
14
16.7 Preliminary work done so far
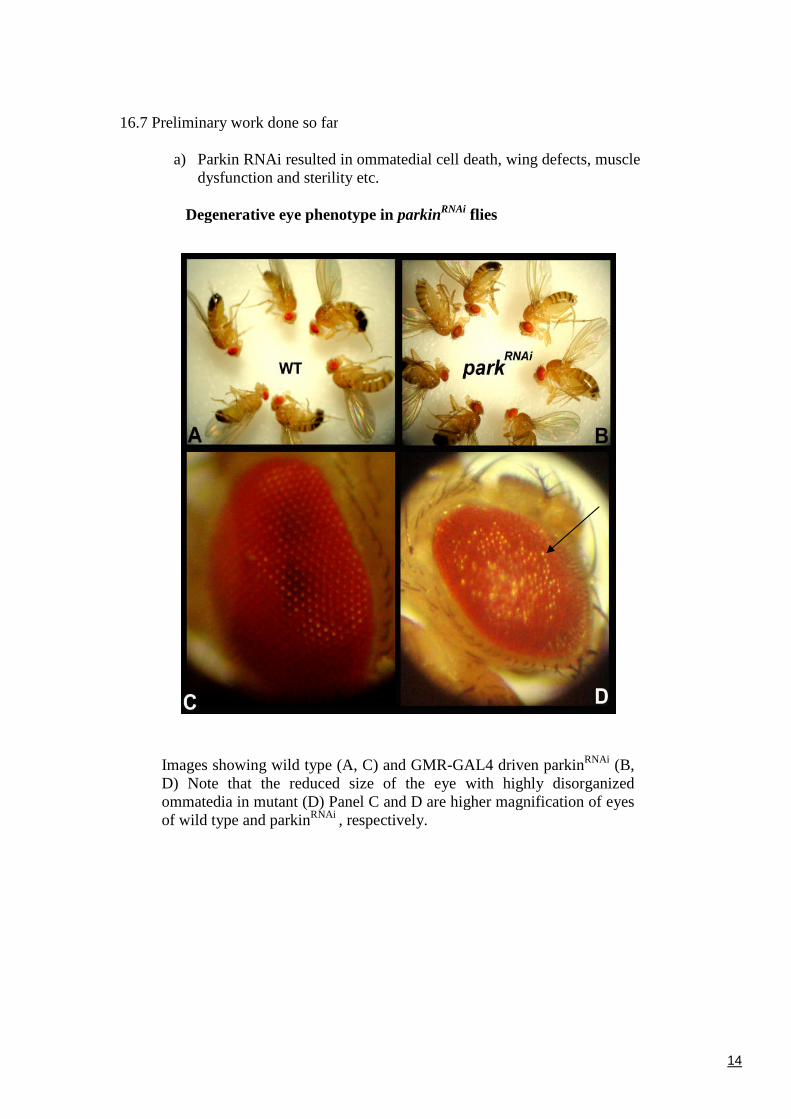
a) Parkin RNAi resulted in ommatedial cell death, wing defects, muscledysfunction and sterility etc.
Degenerative eye phenotype in parkinRNAi flies
Images showing wild type (A, C) and GMR-GAL4 driven parkinRNAi (B,D) Note that the reduced size of the eye with highly disorganizedommatedia in mutant (D) Panel C and D are higher magnification of eyesof wild type and parkinRNAi , respectively.
15
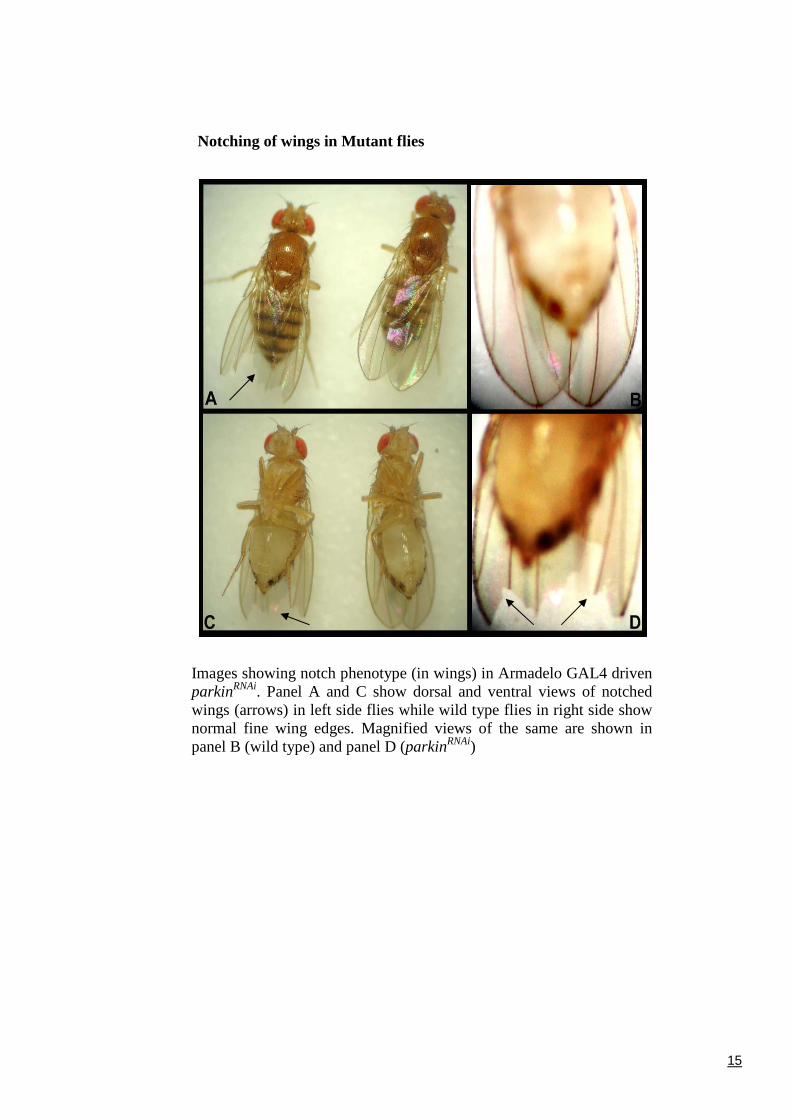
Notching of wings in Mutant flies
Images showing notch phenotype (in wings) in Armadelo GAL4 drivenparkinRNAi. Panel A and C show dorsal and ventral views of notchedwings (arrows) in left side flies while wild type flies in right side shownormal fine wing edges. Magnified views of the same are shown inpanel B (wild type) and panel D (parkinRNAi)
16
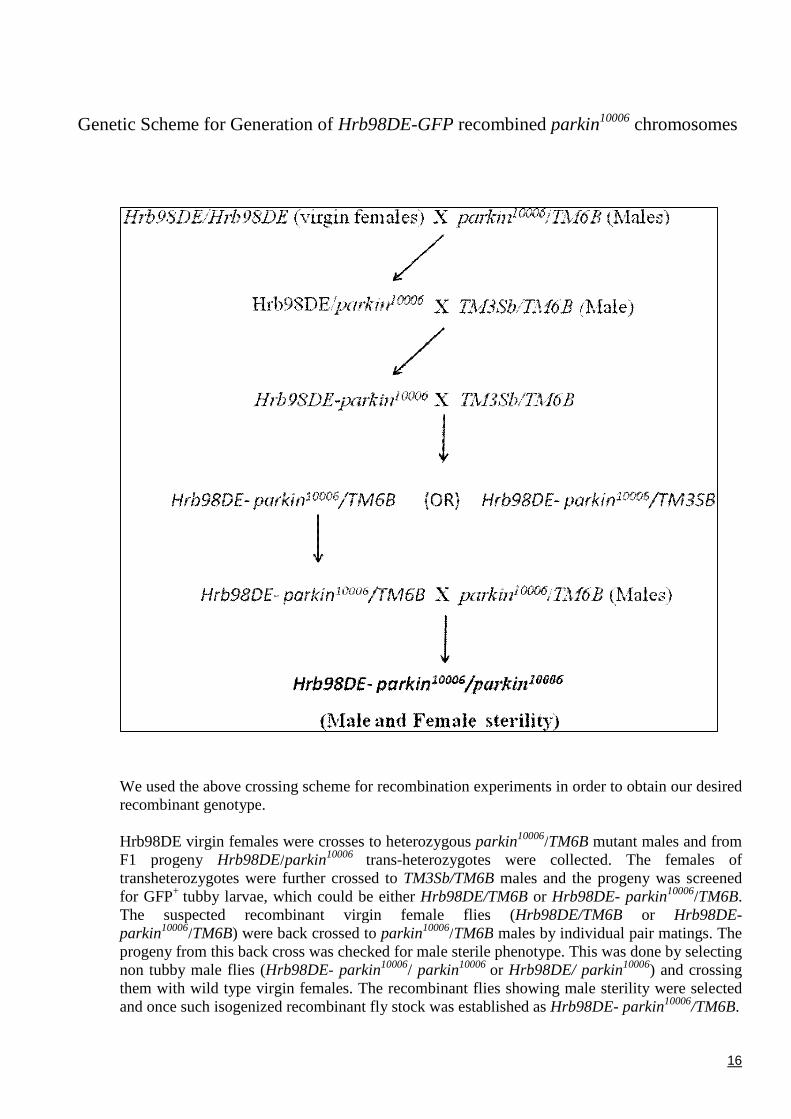
Genetic Scheme for Generation of Hrb98DE-GFP recombined parkin10006 chromosomes
We used the above crossing scheme for recombination experiments in order to obtain our desiredrecombinant genotype.
Hrb98DE virgin females were crosses to heterozygous parkin10006/TM6B mutant males and fromF1 progeny Hrb98DE/parkin10006 trans-heterozygotes were collected. The females oftransheterozygotes were further crossed to TM3Sb/TM6B males and the progeny was screenedfor GFP+ tubby larvae, which could be either Hrb98DE/TM6B or Hrb98DE- parkin10006/TM6B.The suspected recombinant virgin female flies (Hrb98DE/TM6B or Hrb98DE-parkin10006/TM6B) were back crossed to parkin10006/TM6B males by individual pair matings. Theprogeny from this back cross was checked for male sterile phenotype. This was done by selectingnon tubby male flies (Hrb98DE- parkin10006/ parkin10006 or Hrb98DE/ parkin10006) and crossingthem with wild type virgin females. The recombinant flies showing male sterility were selectedand once such isogenized recombinant fly stock was established as Hrb98DE- parkin10006/TM6B.
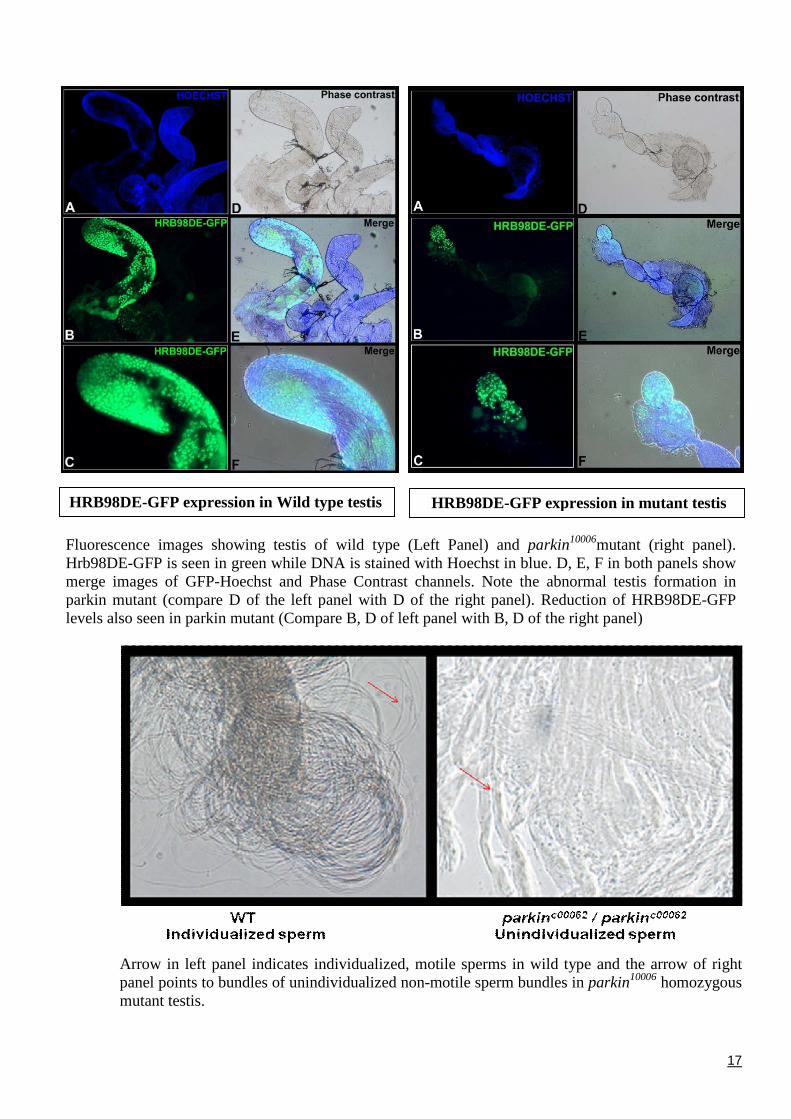
Fluorescence images showing testis of wild type (Hrb98DE-GFP is seen in green while DNA is stainedmerge images of GFP-Hoechst and Phase Contrastparkin mutant (compare D of the left panel with Dlevels also seen in parkin mutant (Compare B, D of lef
Arrow in left panel indicates individualized,panel points to bundles of unindividualized nomutant testis.
HRB98DE-GFP expression in Wild type testis
17
Left Panel) and parkin10006mutant (right panel).with Hoechst in blue. D, E, F in both panels showchannels. Note the abnormal testis formation in
of the right panel). Reduction of HRB98DE-GFPt panel with B, D of the right panel)
motile sperms in wild type and the arrow of rightn-motile sperm bundles in parkin10006 homozygous
HRB98DE-GFP expression in mutant testis
18
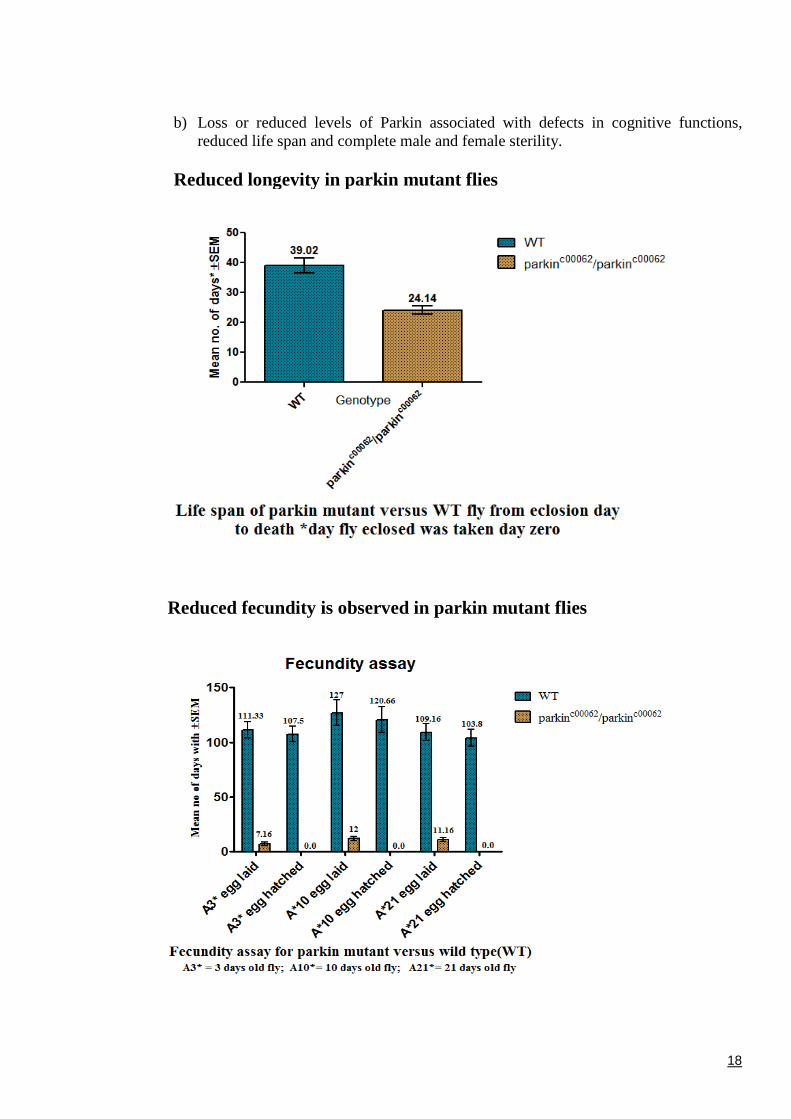
b) Loss or reduced levels of Parkin associated with defects in cognitive functions,reduced life span and complete male and female sterility.
Reduced longevity in parkin mutant flies
Reduced fecundity is observed in parkin mutant flies

19
c) Studies are in progress with JNK related genetic experiments involving both UAS-parkin and UAS-parkinRNAi along with UAS-p53 and UAS-p53RNAi lines.
We have made some progress towards the above objectives and further work is inprogress, which include large scale chromosomal recombinations to bring differentmutant combinations in desired genetic backgrounds. This will be done by bringingUAS-p53 under UAS-parkinRNAi and vice a versa, UAS-parkin with UAS-p53RNAi andUAS-parkin DN under UAS-p53 also in reverse genetic combinations. In all thesecases the JNK pathway candidates will be assessed by RT-PCR, Immunostaining aswell as by western blotting, etc.
Recombination construction work has already begun and also work is progressing inrecombining GFP constructs, of protein trap origin, with desired mutantchromosomes.
17. Specific objectives (should be written in bulleted form, a short paragraph indicating themethods to be followed for achieving the objective and verifiable indicators of progress shouldfollow each specific objective)
To study the spatio-temporal expression and co-localization of Parkin, p53 and JNK inNeuronal versus non-neuronal tissues in Sporadic and Parkinson’s mutants.
To understand the cross-talk between p53, Parkin and JNK during Parkinson’s disease intissue specific knockdown of parkinRNAi, p53RNAi, JNKRNAi and with different geneticcombinations in neuronal versus non-neuronal tissues using GAL4-UAS-RNAiknockdown approaches.
To find out the molecular link between Sporadic and Familial forms of Parkinson’sdisease and to identify the novel common interactors of Parkin and p53 in PD cells.
Screening for various natural and synthetic antioxidant compounds that alleviate JuvenileParkinson’s disease in sporadic as well as parkin mutant flies.
18. Work Plan: should not exceed 3-4 pages (the section can be divided according to the specificaims and under each specific aim, the following should be stated clearly as sub headings)
18.1 Work plan (methodology/experimental design to accomplish the stated aim)
Initial phase of the project involves fly genetics like large scale chromosome recombinations tobring desired mutations (parkin, p53, JNK, Basket, DJ and PINK1, etc) and UAS-RNAi, UAS-GFP, UAS-mCD8::GFP, UAS-mCD8::mCherry carrying chromosomes together in singlechromosome. Once desired chromosomes carrying combinations of mutations are generated theywill be balanced over suitable balancers. These constructs will be used for further molecularstudies.
The over all work plan comprises of In situ expression of desired genes using mRNA specificnon-radioactive (digoxygenin or fluorescent labeled NTPs) riboprobes in whole mount tissues orpartially squashed tissues. Antibody staining and co-localization studies will be done for Parkin,p53 and JNK pathway candidates using anti-primary antibodies under different genetic

20
backgrounds and signal detection will be done using fluorescence conjugated secondaryantibodies (Alexa-conjugates, Cy3, Cy4, FITC, Rhodamine, TRITC, etc). The results obtainedby in situ hybridization, antibody staining, western blotting analysis will be validated withextensive RT-PCR analysis of the candidate genes under various genetic backgrounds. Thisrequires isolation of totals RNA, cDNA preparation, designing gene specific primers andstandardization of PCR, etc. Based on the in situ co-localization and RT-PCR studies, thephysical interaction of the molecules will be assesses by Co-IP (antibody Co-Immunoprecipitations) and western blot analysis, etc. This will be done in different genetic backgrounds(both knockdown and overexpression backgrounds) to validate the true molecular interaction.
To understand the link between sporadic and Genetic/familial forms of the Parkinson’s disease,we will induce PD like situation by various chemicals like Paraquat and assess the geneticfactors like Parkin, p53 and JNK candidates expression. Simultaneously, ROS (Reactive OxygenSpecies) candidates will be assayed by suitable methods in both Sporadic as well as in geneticmutation backgrounds. In this process we will try to identify genetic candidates common for bothSporadic and Familial forms of the disease. Parallelly, we will also go for large scale screeningfor supressors of PD using natural as well as synthetic organic/inorganic compounds thatinterfere with Dopaminergic Neuronal functions. If screening results in any positive candidates,they will be tested further against overexpressed and knockdown/null genetic backgrounds of PDto further validate the specificity of the compounds.
Objective 1: Spatio-temporal expression and co-localization of Parkin, p53 and JNK in PDtissues
We will examine the spatio-temporal in situ expression of Parkin, p53 and JNK pathwaycandidates in neuronal as well as nonneuronal tissues with the help of antibody staining, RNA:RNA in situ hybridization using specific antibodies and riboprobes, respectively. All thesetechniques will be done on wild type and gene specific knockdown/overexpression constructs(UAS-parkin, UAS-parkinRNAi, UAS-parkinDN, UAS-p53, UAS-p53RNAi, UAS-p53DN, UAS-JNK, UAS-JNKRNAi, UAS-JNKDN, UAS-Basket, UAS-BasketRNAi, and UAS-BasketDN) with thehelp of tissue specific GAL4 driver lines. In addition western blotting also will be done to furthervalidate these results.
Objective 2: To understand the cross talk between p53, Parkin and JNK during Parkinson’sdisease
We will examine the expression levels of each candidate through RT-PCR as well as westernblot analysis. The colocalization studies will be carried out using Fluorescence microscopy.Different genetic combinations will be tested for up regulation or down regulation of candidategenes through PCR analysis of desired candidate genes in neuronal versus nonneuronal tissues.Based on the results obtained, a set of Co-Immunoprecipitation (Co-IP) experiments will bedesigned to identify the novel interactors of the Parkin, p53 and JNK in Parkinson’s diseasetissues.
Objective 3: To understand the molecular link between sporadic and familial forms ofParkinson’s disease
Experiments will be designed to understand the link between sporadic and familial forms of theParkinson’s disease. This include analysis of ROS activities in whole organism as well asmitochondrial fractions by various enzymatic and non-enzymatic fluorescence marker based
21
methods to see the status ROS in both sporadic and familial forms. Also we will examine thestatus of Parkin, p53 and JNK candidates in Paraquat induced sporadic PD tissues by RT-PCR aswell as western blotting analysis.
Objective 4: Screening for various antioxidants that alleviate Juvenile Parkinson’s disease
This will be done by feeding flies/larvae with various antioxidants from natural as well assynthetic sources. This will be done in different concentrations in different genetic backgroundsto see if there is any rescue of PD symptoms. We will try to identify positive compounds in thisscreening. This will be followed by studying the molecular mechanism of rescue of PD andmolecular interactors of these compounds. Also we will examine for desired gene expression byRT-PCR, Antibody staining as well as western blotting methods.
18.2 Connectivity of the participating institutions and investigators (In case of multi- institutionalprojects only) NA
18.3 Alternate strategies (if the proposed experimental design or method does not work what isthe alternate strategy)
We are certainly hopeful in getting significant output through this project. In case, if theproposed methods does not work then based on the output of the project we will redesign ourmethodologies by scheming new genetic constructs using advanced tools and techniques, whichwill ultimately enable us to achieve our goals.
19. Timelines: (Please provide quantifiable outputs)
Period of study Achievable targets
6 Months Culturing the fly stocks, making desired genetic recombinants andstandardization of molecular techniques like RT-PCR, In vitrotranscription and RNA: RNA in situ hybridization, combinedantibody co-localization and Co-Immuno Precipitation, etc.
12 Months Expression studies of Parkin, p53 and JNK candidates in desiredgenotypes and their co-localization and isolation of proteininteractors of Parkin, p53, JNK using Immunoprecipitationtechniques.
18 Months Identification of novel interactors and Genetic/ Molecularcharacterization of these novel candidates. Screening of natural andsynthetic antioxidant compounds that rescue Parkinson’s diseasephenotypes.
24 Months Analysis of results, interpretation and compilation of the work doneand submission to the funding agency. Communication of the resultsin peer reviewed journals.
22
20. Name and address of 5 experts in the field
Sr.No. Name Designation Address
1 Dr. D. Kar Chowdhuri Scientist FEmbryo Toxicology Division ITRC,[email protected]
2 Dr. Sreenivasulu Kurukuti AssociateProfessor
Department of Animal SciencesUniversity of Hyderabad, HyderabadTel: +91-40-23134534 (office)[email protected]
3 Dr. Abhay SharmaSeniorScientist Institute of Genomics and Integrative
Biology (IGIB), New Delhi.Ph: +91-11-27666156/7 Ext-166.Fax: [email protected]
4 Dr. Raghuvir Singh Tomar AssistantProfessor
Department of Biology, IISER, BhopalPh: +91-755-4092307Fax: [email protected]
5 Dr. Basant Kumar Patel Assistantprofessor
Department of Biotechnology, IIT,Hyderabad. Ph: +91 (40) 2301 [email protected]
PART IV: BUDGET PARTICULARSBudget (In Rupees)A. Non-Recurring (e.g. equipments, accessories, etc.)
S.No.
Item Year 1 Year 2 Year 3 Total
12
345
PCRPhase contrast Microscope withcamera attachmentBODDeep freezer (-300C)Western Transfer apparatus
3,00,000
1,40,0001,00,000
60,000
Nil Nil 3,00,0003,50,000
1,40,0001,00,000
60,000
Sub-Total (A) = 9, 50,000
23
B. RecurringB.1 Manpower (See guidelines at Annexure-III)
S.No.
PositionNo.
ConsolidatedEmolument
Year 1 Year 2 Year 3 Total
1 Project Fellow 8,000 pm(includingHRA @ 20%)
1,15,200 1,15,200 Nil 2,30,400
Sub-Total (B.1) = 2, 30,400B.2 Consumables
S.No.
Item Quantity Year 1 Year 2 Year 3 Total
1
2
Reagents (Primary &Alexa/CY3-CY4tagged Secondaryantibodies, Primersand other PCRreagents
Glass, Plastic wares &Micro Pipetmans etc.
4,50,000
50,000
1,50,000
50,000
Nil
Nil
6,00,000
1,00,000
Sub-Total (B.2) = 7, 00,000
Other items ConsolidatedEmolument
Year 1 Year 2 Year 3 Total
B.3 Travel 15,000 15,000 15,000 NA 30,000
B.4 Contingency 50,000 50,000 50,000 NA 1,00,000
B.5 Overhead (@20%)(If applicable)
2,50,000 1,52,080 NA 4,02,080
Sub-total of B(B.1+B.2+B.3+B.4+B.5)
14,62,480
Grand Total (A + B) 24,12,480
Note : Please give justification for each head and sub-head separately mentioned in the above table.Financial Year : April - MarchIn case of multi-institutional project, the budget estimate to be given separately for each institution.

24
Justification:
A. Non-Recurring (e.g. equipments, accessories, etc.)
PCR is required to check the expression status of various genes involved in this study.Phase contrast microscope with high resolution camera is essential to capture the imagesof fly specimens especially eye phenotypes and the stained tissues of the larval and adulttissues, etc. We need additional BOD to maintain and culture large scale fly stocks. Theproposed work involves large scale recombination and deficiency screening of hundredsof flies, which require sufficient BOD space. Deep freezer is very much needed to storethe samples of protein, RNA and DNA as well as for maintenance of tissue preparationsbesides storing of variety of molecular grade reagents. We have Western blottingapparatus but are lacking transfer apparatus. Western Transfer apparatus will be used totransfer the separated proteins either through PAGE or Immuno Precipitation (Co-IP)experiments. This will be very much required as we would like to go for ECL blottingafter western analysis.
B. Recurring
B.1 Manpower (See guidelines at Annexure-III)
One Project Fellow is the minimum requirement for the proposed project
B.2 Consumables
Culturing and maintenance of the fly stocks requires polypropylene plastic wares.Isolation of DNA, RNA, and Proteins from desired tissues will need high qualitymolecular biology grade reagents, enzymes, restriction endonucleases, proteaseinhibitors, RNAse and DNAse free chemicals and plastic wares, etc. PVDF membrane isrequired for Protein transfer and immunodetection. ECL (Enhanced ChemiLuminiscence) cassettes and reagents are required for chemiluminiscence signaldetection. PCR and RT-PCR experiments will require PCR grade reagents, eppendorftubes, Primers and other enzymes required for polymerase reaction etc. High precisionmicropipettes, good quality reagent bottles will be purchased from reputed companies.All these items are quite expensive and hence require sufficient amount of budget toprocure in the first year of the project.
B.3 Travel
Travel grant will be utilized for participating in workshops, conferences, attendingseminars and presentations of the project work etc.
B.4 Contingency
It will be utilized to purchase minor routine equipments, maintenance of instruments, dataanalysis at outstation laboratories and to meet expenses of stationary, postage and othermiscellaneous items.
B.5 Overhead: @ 20% as per institution norms.

25
PART V: EXISTING FACILITIES
Resources and additional information
1. Laboratory:
a. Manpower
Currently Two JRFs are working in the Laboratory Two research trainees are working for short term projects
b. Equipments
BOD incubator (small) Table top centrifuge Oven Simple Binocular Microscope
2. Other resources such as clinical material, animal house facility, glass house.Experimental garden, pilot plant facility etc.
Drosophila fly culture lab is available
26
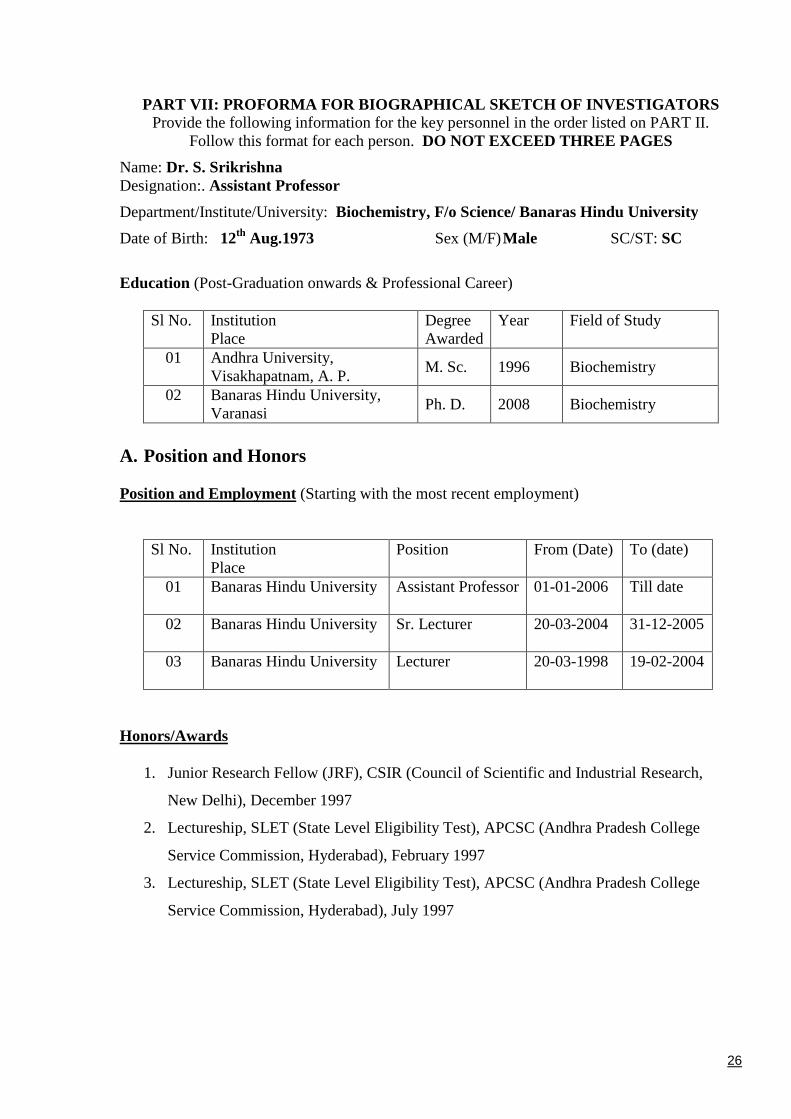
PART VII: PROFORMA FOR BIOGRAPHICAL SKETCH OF INVESTIGATORSProvide the following information for the key personnel in the order listed on PART II.
Follow this format for each person. DO NOT EXCEED THREE PAGES
Name: Dr. S. SrikrishnaDesignation:. Assistant Professor
Department/Institute/University: Biochemistry, F/o Science/ Banaras Hindu University
Date of Birth: 12th Aug.1973 Sex (M/F)Male SC/ST: SC
Education (Post-Graduation onwards & Professional Career)
Sl No. InstitutionPlace
DegreeAwarded
Year Field of Study
01 Andhra University,Visakhapatnam, A. P.
M. Sc. 1996 Biochemistry
02 Banaras Hindu University,Varanasi
Ph. D. 2008 Biochemistry
A. Position and Honors
Position and Employment (Starting with the most recent employment)
Sl No. InstitutionPlace
Position From (Date) To (date)
01 Banaras Hindu University Assistant Professor 01-01-2006 Till date
02 Banaras Hindu University Sr. Lecturer 20-03-2004 31-12-2005
03 Banaras Hindu University Lecturer 20-03-1998 19-02-2004
Honors/Awards
1. Junior Research Fellow (JRF), CSIR (Council of Scientific and Industrial Research,
New Delhi), December 1997
2. Lectureship, SLET (State Level Eligibility Test), APCSC (Andhra Pradesh College
Service Commission, Hyderabad), February 1997
3. Lectureship, SLET (State Level Eligibility Test), APCSC (Andhra Pradesh College
Service Commission, Hyderabad), July 1997
27
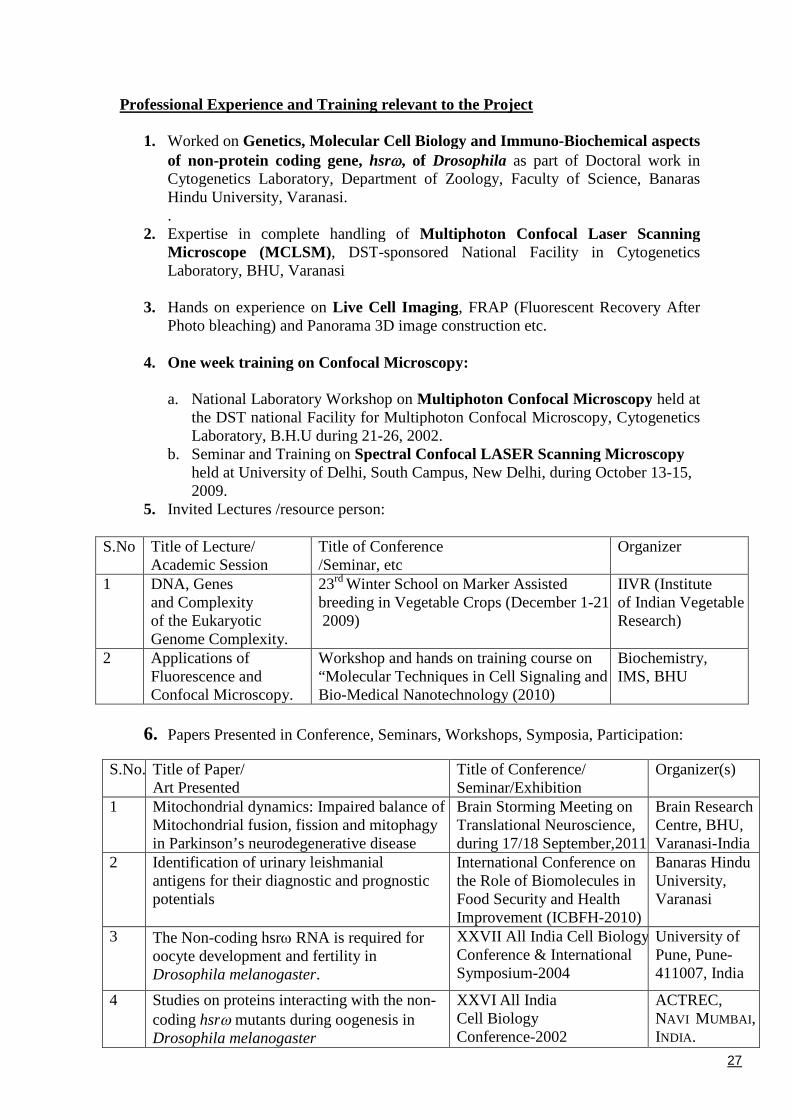
Professional Experience and Training relevant to the Project
1. Worked on Genetics, Molecular Cell Biology and Immuno-Biochemical aspectsof non-protein coding gene, hsr, of Drosophila as part of Doctoral work inCytogenetics Laboratory, Department of Zoology, Faculty of Science, BanarasHindu University, Varanasi..
2. Expertise in complete handling of Multiphoton Confocal Laser ScanningMicroscope (MCLSM), DST-sponsored National Facility in CytogeneticsLaboratory, BHU, Varanasi
3. Hands on experience on Live Cell Imaging, FRAP (Fluorescent Recovery AfterPhoto bleaching) and Panorama 3D image construction etc.
4. One week training on Confocal Microscopy:
a. National Laboratory Workshop on Multiphoton Confocal Microscopy held atthe DST national Facility for Multiphoton Confocal Microscopy, CytogeneticsLaboratory, B.H.U during 21-26, 2002.
b. Seminar and Training on Spectral Confocal LASER Scanning Microscopyheld at University of Delhi, South Campus, New Delhi, during October 13-15,2009.
5. Invited Lectures /resource person:
S.No Title of Lecture/Academic Session
Title of Conference/Seminar, etc
Organizer
1 DNA, Genesand Complexityof the EukaryoticGenome Complexity.
23rd Winter School on Marker Assistedbreeding in Vegetable Crops (December 1-21,2009)
IIVR (Instituteof Indian VegetableResearch)
2 Applications ofFluorescence andConfocal Microscopy.
Workshop and hands on training course on“Molecular Techniques in Cell Signaling andBio-Medical Nanotechnology (2010)
Biochemistry,IMS, BHU
6. Papers Presented in Conference, Seminars, Workshops, Symposia, Participation:
S.No. Title of Paper/Art Presented
Title of Conference/Seminar/Exhibition
Organizer(s)
1 Mitochondrial dynamics: Impaired balance ofMitochondrial fusion, fission and mitophagyin Parkinson’s neurodegenerative disease
Brain Storming Meeting onTranslational Neuroscience,during 17/18 September,2011
Brain ResearchCentre, BHU,Varanasi-India
2 Identification of urinary leishmanialantigens for their diagnostic and prognosticpotentials
International Conference onthe Role of Biomolecules inFood Security and HealthImprovement (ICBFH-2010)
Banaras HinduUniversity,Varanasi
3 The Non-coding hsr RNA is required foroocyte development and fertility inDrosophila melanogaster.
XXVII All India Cell BiologyConference & InternationalSymposium-2004
University ofPune, Pune-411007, India
4 Studies on proteins interacting with the non-coding hsr mutants during oogenesis inDrosophila melanogaster
XXVI All IndiaCell BiologyConference-2002
ACTREC,NAVI MUMBAI,INDIA.
28
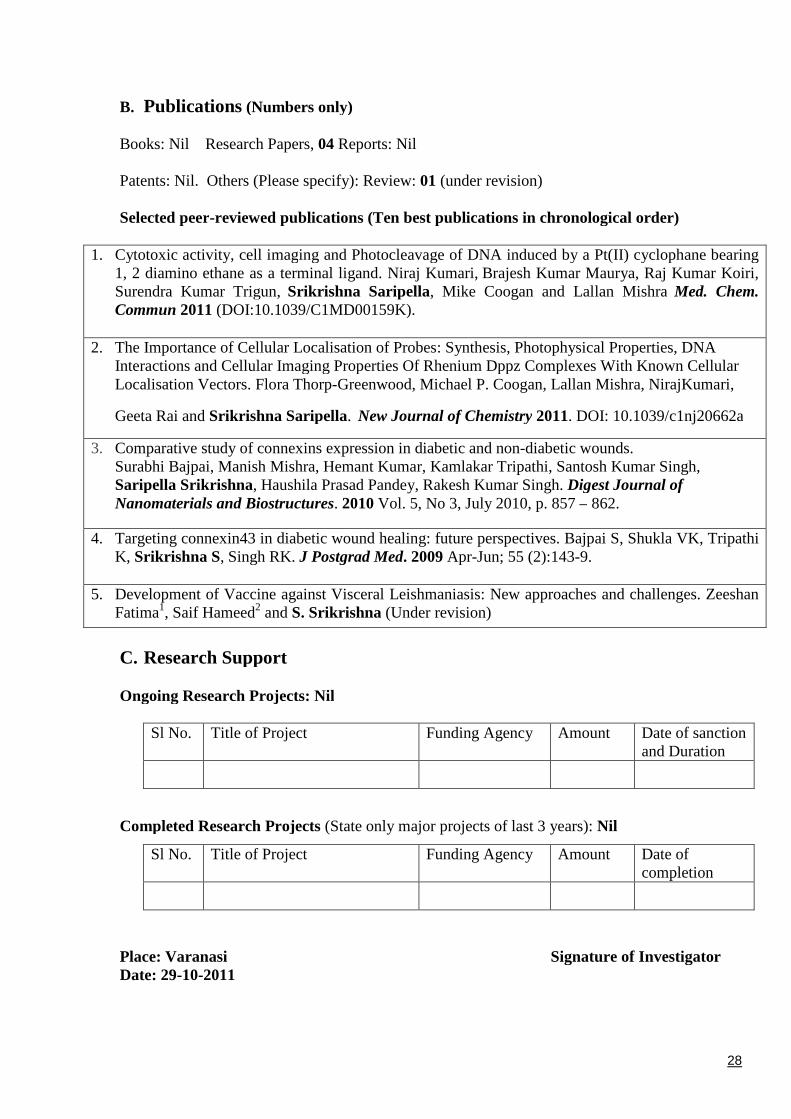
B. Publications (Numbers only)
Books: Nil Research Papers, 04 Reports: Nil
Patents: Nil. Others (Please specify): Review: 01 (under revision)
Selected peer-reviewed publications (Ten best publications in chronological order)
1. Cytotoxic activity, cell imaging and Photocleavage of DNA induced by a Pt(II) cyclophane bearing1, 2 diamino ethane as a terminal ligand. Niraj Kumari, Brajesh Kumar Maurya, Raj Kumar Koiri,Surendra Kumar Trigun, Srikrishna Saripella, Mike Coogan and Lallan Mishra Med. Chem.Commun 2011 (DOI:10.1039/C1MD00159K).
2. The Importance of Cellular Localisation of Probes: Synthesis, Photophysical Properties, DNAInteractions and Cellular Imaging Properties Of Rhenium Dppz Complexes With Known CellularLocalisation Vectors. Flora Thorp-Greenwood, Michael P. Coogan, Lallan Mishra, NirajKumari,
Geeta Rai and Srikrishna Saripella. New Journal of Chemistry 2011. DOI: 10.1039/c1nj20662a
3. Comparative study of connexins expression in diabetic and non-diabetic wounds.Surabhi Bajpai, Manish Mishra, Hemant Kumar, Kamlakar Tripathi, Santosh Kumar Singh,Saripella Srikrishna, Haushila Prasad Pandey, Rakesh Kumar Singh. Digest Journal ofNanomaterials and Biostructures. 2010 Vol. 5, No 3, July 2010, p. 857 – 862.
4. Targeting connexin43 in diabetic wound healing: future perspectives. Bajpai S, Shukla VK, TripathiK, Srikrishna S, Singh RK. J Postgrad Med. 2009 Apr-Jun; 55 (2):143-9.
5. Development of Vaccine against Visceral Leishmaniasis: New approaches and challenges. ZeeshanFatima1, Saif Hameed2 and S. Srikrishna (Under revision)
C. Research Support
Ongoing Research Projects: Nil
Sl No. Title of Project Funding Agency Amount Date of sanctionand Duration
Completed Research Projects (State only major projects of last 3 years): Nil
Sl No. Title of Project Funding Agency Amount Date ofcompletion
Place: Varanasi Signature of InvestigatorDate: 29-10-2011



















![STUDIES OF THE DROSOPHILA BRAIN USING P[GAL4] ENHANCER …theses.gla.ac.uk/75487/1/13832077.pdf · 2019. 11. 19. · STUDIES OF THE DROSOPHILA BRAIN USING P[GAL4] ENHANCER TRAP LINES](https://img.pdfslide.us/doc/110x75/613774870ad5d2067648a163/studies-of-the-drosophila-brain-using-pgal4-enhancer-2019-11-19-studies-of.jpg)









