Embed Size (px)
Citation preview

Čedomil LucuCenter for Marine ResearchInstitute Ruđer BoškovićRovinj, Istria, Croatia
PHYSIOLOGICAL ADAPTIVE MECHANISMS OF ORGANISMS IN MARINE AND BRACKISH WATER ENVIRONMENT.
Na+ K+ ATPase AND CARBONIC ANHYDRASE
Adriatic Sea 36 -39 pptBrackish waters < 30 pptFreshwater < 0.5 ppt

European lobster Homarus gammarus (Linnaeus, 1758.)

Table 1.
Osmolarity and ion concentrations of the haemolymph of intermoult lobsters
Homarus gammarus acclimated to 38 ppt SW and 20 ppt DSW. Medium
concentration in which lobsters were acclimated are indicated. Each point is
given as the mean ± SD. In abbreviations are numbers of tested individuals.
OSMOLARITY Na Cl
(mOsmol/l) (mmol/ l)
SEAWATER
1126±58 (6) 528±53 (6) 532±21 (5)
HAEMOLYMPH
1168±68 (6) 538±19 (6) 575±20 (5)
DILUTE SEAWATER
548.3±17.7 (6) 274±13 (6) 306±14 (6)
HAEMOLYMPH
701±30 (6) 334±6 (6) 334±16 (6)

e
i
vt
k
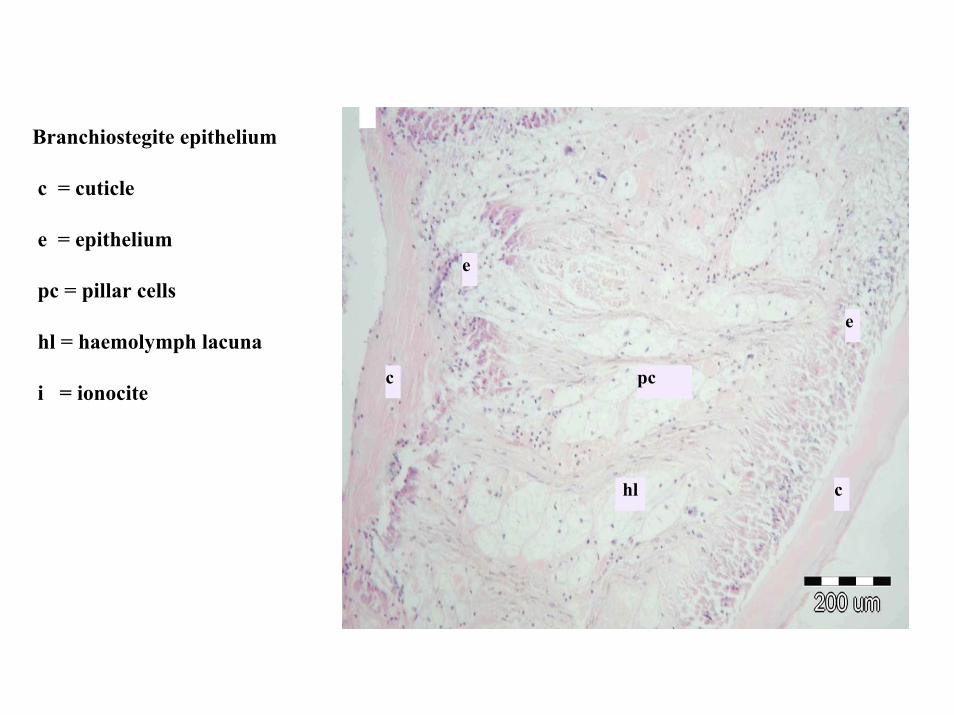
e
c
c
pc
hl
e
Branchiostegite epithelium
c = cuticle
e = epithelium
pc = pillar cells
hl = haemolymph lacuna
i = ionocite

DILUTE SEAWATER ACCLIMATION
Morphological and ultrastructural changesIn the branchial cavity of homarid lobsters tissues i.e. branchiostegite and epipoditeincrease in tisssue thickness, basolateral infoldings, apical microvilli, increase in numbers of mitochondria, microvesicular bodies.
Biochemical and physiologica changesIncreased oxydation rates of glucose and amino acids –36% of total CO2 released contribution of alanine oxidation (Pressley and Graves, 1983)
increase of cytochrome c oxidase (Welcomme and Devos, 1988)
Increased oxygen consumption rate in the gills in vitro (Lucu and Pavičić, 1995)
Olfactory function sustained (Gleeson et al.,1997)
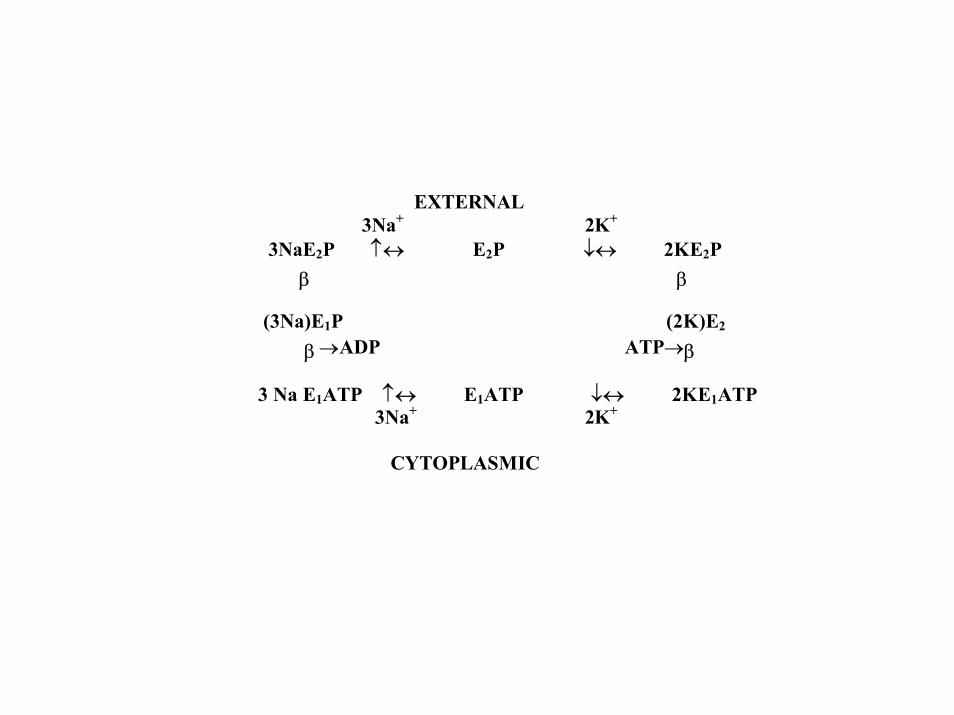
EXTERNAL 3Na+ 2K+ 3NaE2P ↑↔ E2P ↓↔ 2KE2P β β (3Na)E1P (2K)E2 β→ADP ATP→β 3 Na E1ATP ↑↔ E1ATP ↓↔ 2KE1ATP 3Na+ 2K+ CYTOPLASMIC

0
20
40
60
80
100
% O
F IN
HIB
ITIO
N
-5 -4 -3 -2-log Ouabain (mol/l)
0.0
0.2
0.4
0.6
0.8
( u
mol
Rb/
h/m
g pr
otei
n)
980 mosmol/l
650 mosmol/l
**
**
**
µ
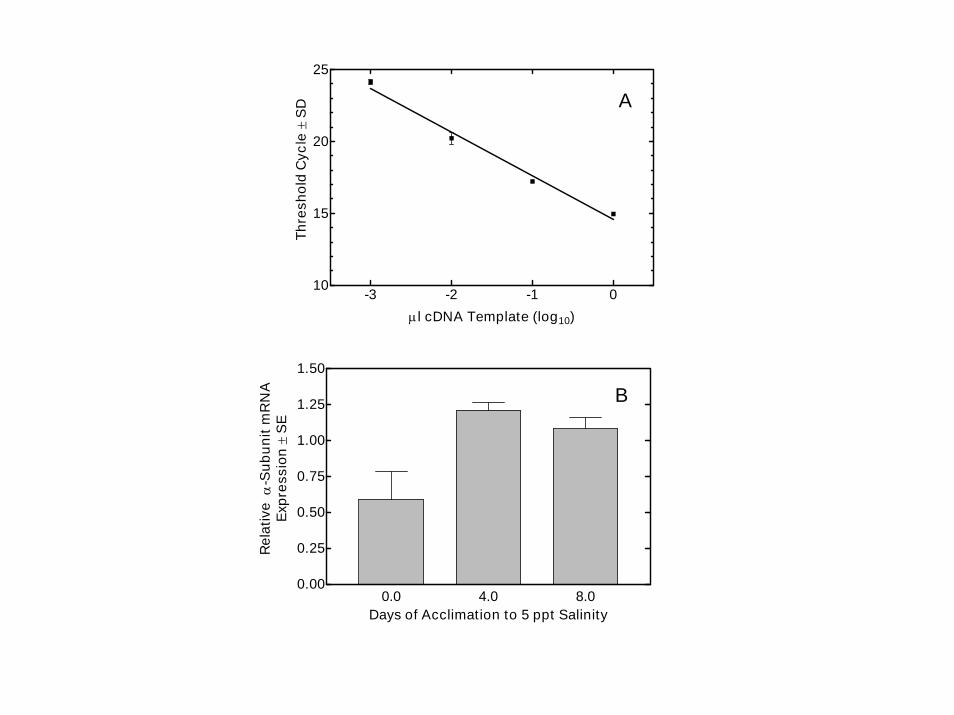
-3 -2 -1 010
15
20
25
A
µl cDNA Template (log10)Th
resh
old
Cyc
le±
SD
0.0 4.0 8.00.00
0.25
0.50
0.75
1.00
1.25
1.50
Days of Acclimation to 5 ppt Salinity
Rel
ativ
eα
-Sub
unit
mR
NA
Expr
essi
on±
SE
B

Callinectes : MADTGRTDSYRHATDRNIPDDNRTVKGDPKSKKKNVKGKRKGEKEKDMDNLKQELELDEHKVPIEELFQRLSVNPDTGLTQAEARRRLERDGPNALTPPK : 100Homarus : MADTGRTDSYRHATDRTIPDDNRTVKGDPKSTKKMPKAKAK-KGEKDMDNLKRELELDEHKVPIEELFQRLSVNPDSGLTQAEALRRTERDGPNALTPPK : 99Artemia1 : ------------------------------------MAKGKQKKGKDLNELKKELDIDFHKIPIEECYQRLGSNPETGLTNAQARSNIERDGPNCLTPPK : 64Gallus3 : -----------------------------MGDKGEKESPKKGKGKRDLDDLKKEVAMTEHKMSIEEVCRKYNTDCVQGLTHSKAQEILARDGPNALTPPP : 71Gallus2 : ---------------------MDGREYSPAATTSENGGGRRKQKEKELDELKKEVNLDDHKLSLDELGRKYQVDLSRGLSNARAAEVLAQDGPNALTPPP : 79Gallus1 : ------------------MGKGAGRDKYEPTATSEHGTKKKKAKERDMDELKKEISMDDHKLSLDELHRKYGTDLSRGLTTARAAEILARDGPNTLTPPP : 82Artemia2 : ---------------------------------------MGKKQGKQLSDLKKELELDQHKIPLEELCRRLGTNTETGLTSSQAKSHLEKYGPNALTPPR : 61 Callinectes : QTPEWVKFCKNLFGGFSLLLWIGAILCFIAYSIEAAS-EEEPNNDNLYLGIVLTAVVIITGIFSYYQESKSSRIMESFKNLVPQYAIVIREGEKLNVQAE : 199Homarus : QTPEWVKFCKNLFGGFSLLLWIGAILCFIAYSIEAVS-EEEPNNDNLYLGIVLTAVVIITGVFSYYQENKSSRIMESFKNMVPQYAIVVREGEKQNVQAE : 198Artemia1 : TTPEWIKFCKNLFGGFALLLWTGAILCFLAYGIEASSGNEDMLKDNLYLGIVLATVVIVTGIFSYYQENKSSRIMDSFKNLVPQYALALREGQRVTLKAE : 164Gallus3 : TTPEWVKFCRQLFGGFSILLWIGAILCFLAYGIQAGT-EDEPSNDNLYLGIVLAAVVIITGCFSYYQEAKSSKIMESFKNMVPQQALVIREGEKMQLNAE : 170Gallus2 : TTPEWVKFCRQLFGGFSILLWIGAILCFLAYGIQAAM-EDEPSNDNLYLGVVLAAVVIVTGCFSYYQEAKSSKIMDSFKNMVPQQALVIREGEKIQINAE : 178Gallus1 : TTPEWVKFCRQLFGGFSLLLWIGSLLCFLAYGITSVM-EGEPNSDNLYLGVVLAAVVIITGCFSYYQEAKSSKIMESFKNMVPQQALVVRNGEKMSINAE : 181Artemia2 : TTPEWIKFCKQLFGGFQMLLWIGSILCFIAYTMEKYK-NPDVLGDNLYLGLALLFVVIMTGCFAYYQDHNASKIMDSFKNLMPQFAFVIRDGKKIQLKAE : 160 Callinectes : ELCIGDIIDVKFGDRIPADMRVIEARGFKVDNSSLTGESEPQSRSPEFTSENPLETKNLAFFSTNAVEGTCKGIVINIGDNTVMGRIAGLASGLETGETP : 299Homarus : ELCIGDIIEVKFGDRIPADVRVIEARGFKVDNSSLTGESEPQSRSPEFTSENPLETKNLAFFSTNAVEGTARGIVINIGDNTVMGRIAGLASGLETGETP : 298Artemia1 : ELTMGDIVEVKFGDRVPADLRVLEARSFKVDNSSLTGESEPQARSPEFTNDNPLETKNLAFFSTNAVEGTMRGIVIGIGDNTVMGRIAGLASGLDTGETP : 264Gallus3 : EVVVGDLVEVKGGDRVPADLRIISAHGCKVDNSSLTGESEPQTRSPDCTHDNPLETRNITFFSTNCVEGTARGVVIATGDRTVMGRIATLASGLEVGKTP : 270Gallus2 : NVVVGDLVEVKGGDRVPADMRIISSHGCKVDNSSLTGESEPQTRSPEFTHENPLETRNICFFSTNCVEGTARGIVISTGDRTVMGRIASLASGLEVGRTP : 278Gallus1 : GVVVGDLVEVKGGDRIPADLRIISAHGCKVDNSSLTGESEPQTRSPDFSNENPLETRNIAFFSTNCVEGTAVGIVISTGDRTVMGRIASLASGLEGGKTP : 281Artemia2 : EVTVGDLVEVKFGDRIPADIRITSCQSMKVDNSSLTGESEPQSRSTECTNDNPLETKNLAFFFTNTLEGTGRGIVINVGDDSVMGRIACLASSLDSGKTP : 260 Callinectes : IAKEISHFIHIITGVAVFLGVTFFVIAFIMGYHWLDAVVFLIGI-IVANVPEGLLATVTVCLTLTAKRMAAKNCLVKNLEAVETLGSTSTICSDKTGTLT : 398Homarus : IAKEISHFIHIITGVAVFLGVTFFVIAFILGYHWLDAVVSLIGYHCKPNVPEGLLATVTVCLTLTAKRMAAKNCLVKNLEAVETLGSTSTICSDKTGTLT : 398Artemia1 : IAKEIAHFIHIITGVAVFLGVTFFIIAFVLGYHWLDAVVFLIGI-IVANVPEGLLATVTVCLTLTAKRMASKNCLVKNLEAVETLGSTSTICSDKTGTLT : 363Gallus3 : IAVEIEHFIQLITGVAVFLGISFFVLSLILGYTWLEAVIFLIGI-IVANVPEGLLATVTVCLTLTAKRMARKNCLVKNLEAVETLGSTSTICSDKTGTLT : 369Gallus2 : IAMEIEHFIRLITGVAVFLGLSFFILSLILGYTWLEAVIFLIGI-IVANVPEGLLATVTVCLTLTAKRMARKNCLVKNLEAVETLGSTSTICSDKTGTLT : 377Gallus1 : IAMEIEHFIHLITGVAVFLGVSFFILSLILEYTWLEAVIFLIGI-IVANVPEGLLATVTVCLTLTAKRMARKNCLVKNLEAVGTLGSTSTICSDKTGTLT : 380Artemia2 : IAREIEHFIHIITAMAVSLAAVFAVISFLYGYTWLEAAIFMIGI-IVAKVPEGLLATVTVCLTLTAKRMAKKNCLVRNLEAVETLGSTSTICSDKTGTLT : 359 Callinectes : QNRMTVAHMWFDNTIIEADTSEDQSGCQYDKTSDGWKALSRIAALCNRAEFKTGQEDVPILKREVNGDASEAALLKCVELAIGDVRGWRSRNKKVCEIPF : 498Homarus : QNRMTVAHMWFDNTIIEADTSEDQSGCQYDKTSEGWKTLSRIAALCNRAEFKTGQEEVAILKREVNGDASEAALLKCVELAVGDVKGWRARNKKVCEVPF : 498Artemia1 : QNRMTVAHMWFDGTITEADTTEDQSGAQFDKSSAGWKALVKIAALCSRAEFKPNQSTTPILKREVTGDASEAAILKCVELTTGETEAIRKRNKKICEIPF : 463Gallus3 : QNRMTVAHMWFDNQIHEADTTEDQSGTSFDKSSATWVALSHIAGLCNRAVFKGGQENVPILKRDVAGDASESALLKCIELSSGSVKVMRERNKKVAEIPF : 469Gallus2 : QNRMTVAHMWFDNQIHEADTTEDQSGATFDKRSPTWAALSRIAGLCNRAVFKPGQENISISKRDTAGDASESALLKCIQLSCGSVKKMRDKNPKVTEIPF : 477Gallus1 : QNRMTVAHMWFDNQIHEADTTENQSGASFDKSSATWLALSRIAGLCNRAVFQANQENVPILKRAVAGDASESALLKCIELCCGSVKEMRERYPKVVEIPF : 480Artemia2 : QNRMTVAHMWFDQKIVTADTTENQSGNQLYRGSKGFPELIRVASLCSRAEFKTEHAHLPVLKRDVNGDASEAAILKFAEMSTGSVMNIRSKQKKVSEIPF : 459 Callinectes : NSTNKYQVSIHETQDKNDLRYLLVMKGAPERILERCSTIFMNGEEKPLDEEMKESFNNAYLELGGLGERVLGFCDYVLPSDKYPLGYPFDADAVNFPVHG : 598Homarus : NSTNKYQVSIHDTEDKNDPRYLLVMKGAPERILERCSTIYINGEEKPLDEELKESFNNAYLELGGLGERVLGFCDYILPSDKYPLGYPFDSDNPNFPVHG : 598Artemia1 : NSANKFQVSIHENEDKSDGRYLLVMKGAPERILERCSTIFMNGKEIDMTEELKEAFNNAYMELGGLGERVLGFCDYLLPLDKYPHGFAFNADDANFPLTG : 563Gallus3 : NSTNKYQLSIHETEDPNDNRYLLVMKGAPERILDRCSTILLQGKEQPLDEEMKEAFQNAYLELGGLGERVLGFCHFYLPEEQYPKGFAFDCDDVNFATDN : 569Gallus2 : NSTNKYQLSIHEREEDPQG-HILVMKGAPERILERCSRILLQGQEVPLDEEMKEAFQNAYLELGGLGERVLGFCHLYLPPDKFPRGFRFDADEVNFPTSD : 576Gallus1 : NSTNKYQLSIHKNANAGESRHLLVMKGAPERILDRCDSILIHGKVQPLDEEIKDAFQNAYLELGGLGERVLGFCHLALPDDQFPEGFQFDTDEVNFPVEK : 580Artemia2 : NSANKYQVSVHEREDKSG--YFLVMKGAPERILERCSTILIDGTEIPLDNHMKECFNNAYMELGGMGERVLGFCDFELPSDQYPRGYVFDADEPNFPISG : 557 Callinectes : LRFVGLMSMIDPPRAAVPDAVAKCRSAGIKVIMVTGDHPITAKAIAKSVGIISEGNETVEDIAQRLNIPIKEVDPREAKAAVVHGSELRDMTSEQLDDVL : 698Homarus : LRFVGLMSMIDPPRAAVPDAVAKCRSAGIKVIMVTGDHPITAKAIAKSVGIISEGNETVEDIAQRLNIPIKEVDPREAKAAVVHGSELRDMTSEQLDDIL : 698Artemia1 : LRFAGLMSMIDPPRAAVPDAVAKCRSAGIKVIMVTGDHPITAKAIAKSVGIISEGNETVEDIAARLNIPVSEVNPRDAKAAVVHGGELRDITPDALDEIL : 663Gallus3 : LCFVGLMSMIDPPRAAVPDAVGKCRSAGIKVIMVTGDHPITAKAIAKGVGIISEGNETVEDIAARLNIPVSQVNPRDAKACVIHGTDLKDMSSEQIDEIL : 669Gallus2 : LCFVGLMSMIDPPRAAVPDAVGKCRSAGIKVIMVTGDHPITAKAIAKGVGIISEGNETVEDIAARLNIPVSQVNPREAKACVVHGSDLKDMTAEQLDEIL : 676Gallus1 : LCFVGLMSMIDPPRAAVPDAVGKCRSAGIKVIMVTGDHPITAKAIAKGVGIISDGNETVEDIAARLNIPVSQVNPRDAKACVVHGSDLKDMTSEQLDDIL : 680Artemia2 : LRFVGLMSMIDPPRAAVPDAVSKCRSAGIKVIMVTGDHPITAKAIARQVGIISEGHETVDDIAARLNIPVSEVNPRSAQAAVIHGNDLKDMNSDQLDDIL : 657 Callinectes : IHHTEIVFARTSPQQKLIIVEGCQRMGAIVAVTGDGVNDSPALKKADIGVAMGIAGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNLKKSIAYTLTS : 798Homarus : IHHTEIVFARTSPQQKLIIVEGCQRMGAIVAVTGDGVNDSPALKKADIGVAMGIAGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNLKKSIAYTLTS : 798Artemia1 : RHHPEIVFARTSPQQKLIIVEGCQRQGAIVAVTGDGVNDSPALKKADIGVAMGIAGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNLKKSIVYTLTS : 763Gallus3 : QNHTEIVFARTSPQQKLIIVEGCQRQGAIVAVTGDGVNDSPALKKADIGVAMGIRGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNLKKSIAYTLTS : 769Gallus2 : RNHTEIVFARTSPQQKLIIVEGCQRQGAIVAVTGDGVNDSPALKKADIGIAMGIAGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNLKKSIAYTLTS : 776Gallus1 : LHHTEIVFARTSPQQKLIIVEGCQRQGAIVAVTGDGVNDSPALKKADIGVAMGIAGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNLKKSIAYTLTS : 780Artemia2 : RHYREIVFARTSPQQKLIIVEGVQRQGEFVAVTGDGVNDSPALKKADIGVAMGIAGSDVSKQAADMILLDDNFASIVTGVEEGRLIFDNIKKSIAYTLTS : 757 Callinectes : NIPEISPFLFFMIASVPLPLGTVTILCIDLGTDMVPAISLAYEEAESDIMKRQPRNPFTDKLVNERLISMAYGQIGMIQALAGFYVYFVIMAENGFLPPV : 898Homarus : NIPEISPFLFFMIASVPLPLGTVTILCIDLGTDMVPAISLAYEEAESDIMKRQPRNPFTDKLVNERLISMAYGQIGMIQALAGFFVYFVIMAENGFLPPK : 898Artemia1 : NIPEISPFLLFILFDIPLPLGTVTILCIDLGTDMVPAISLAYEEAESDIMKRRPRNPVTDKLVNERLISLAYGQIGMIQASAGFFVYFVIMAECGFLPWD : 863Gallus3 : NIPEITPFLLFIMANIPLPLGTITILCIDLGTDMVPAISLAYEAAESDIMKRQPRNPRSDKLVNERLISMAYGQIGMIQALGGFFSYFVILAENGFLPSC : 869Gallus2 : NIPEITPFLLFIIANIPLPLGTVTILCIDLGTDMVPAISLAYEAAESDIMKRQPRNPRTDKLVNERLISMAYGQIGMIQALGGFFTYFVILAENGFLPAR : 876Gallus1 : NIPEITPFLIFIIANIPLPLGTCTILCIDLGTDMVPAISLAYEQAESDIMKRQPRNPKTDKLVNERLISMAYGQIGMIQALGGFFTYFVIMAENGFLPSG : 880Artemia2 : KIPELSPFLMYILFDLPLAIGTVTILCIDLGTDVVPAISMAYEGPEAD--PRKPRDPVKEKLVNERLISMAYGQIGVMQAFGGFFTYFVIMGECGFLPNR : 855 Callinectes : LFGIREQWDSKAINDLEDYYGQEWTYHDRKILEYTCHTAFFVAIVVVQWADLIICKTRRNSILHQGMKNMVLNFGLCFETTLAAFLSYTPGMDKGLRMYP : 998Homarus : LFGIREKWDSKAINDLEDHYGQEWTYYDRKVLEYTCHTAFFVSIVIVQWADLIVCKTRRNSIVHQGMKNMVLNFGLCFETTLAAFLSYTPGMDKGLRMYP : 998Artemia1 : LFGLRKHWDSRAVNDLTDSYGQEWTYDARKQLESSCHTAYFVSIVIVQWADLIISKTRRNSVFQQGMRNNILNFALVFETCLAAFLSYTPGMDKGLRMYP : 963Gallus3 : LVGIRLSWDDRTINDLEDSYGQQWTYEQRKVVEFTCHTAFFVSIVVVQWADLIICKTRRNSVFQQGMKNKILIFGLFEETALAAFLSYCPGMDVALRMYP : 969Gallus2 : LLGVRLAWDDRSTNDLEDSYGQEWTYEQRKVVEFTCHTAFFASIVVVQWADLIICKTRRNSVFQQGMKNKILIFGLLEETALAAFLSYCPGMGVALRMYP : 976Gallus1 : LVGIRLQWDDRWINDVEDSYGQQWTFEQRKIVEFTCHTAFFVSIVVVQWADLIICKTRRNSVFQQGMKNKILIFGLFEETALAAFLSYCPGMDVALRMYP : 980Artemia2 : LFGLRKWWESKAYNDLTDSYGQEWTWDARKQLEYTCHTAFFISIVIVQWTDLIICKTRRLSLFQQGMKNGTLNFALVFETCVAAFLSYTPGMDKGLRMYP : 955 Callinectes : LKFYWWLPPLPFSLLIFVYDECRRFVLRRNPGGWVEMETYY : 1039Homarus : LKFYWWLPALPFSLLIFVYDECRRFILRRNPGGWMEMETYY : 1039Artemia1 : LKINWWFPALPFSFLIFVYDEARKFILRRNPGGWVEQETYY : 1004Gallus3 : LKPSWWFCAFPYSFLIFVYDEIRKLILRRNPGGWVEKETYY : 1010Gallus2 : LKVTWWFCAFPYSLLIFAYDEVRKLILRRYPGGWVEKETYY : 1017Gallus1 : LKPTWWFCAFPYSLLIFLYDEIRKLIIRRNPGGWVERETYY : 1021Artemia2 : LKIWWWFPPMPFSLLILVYDECRKFLMRRNPGGFLERETYY : 996
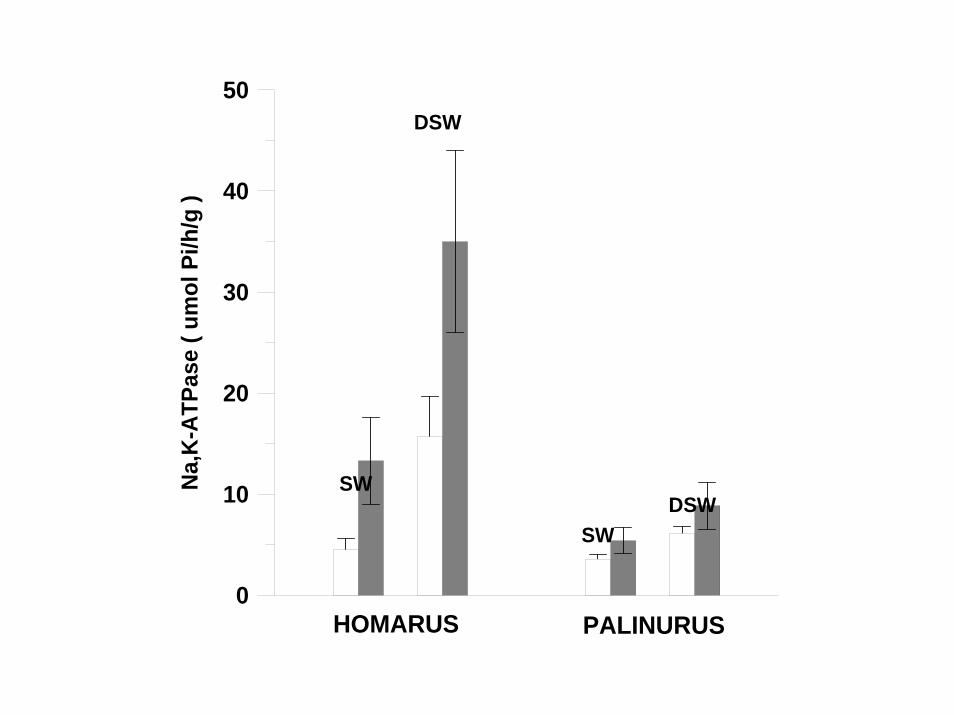
0
10
20
30
40
50
Na,
K-A
TPas
e ( u
mol
Pi/h
/g )
HOMARUS PALINURUS
SW
DSW
SWDSW
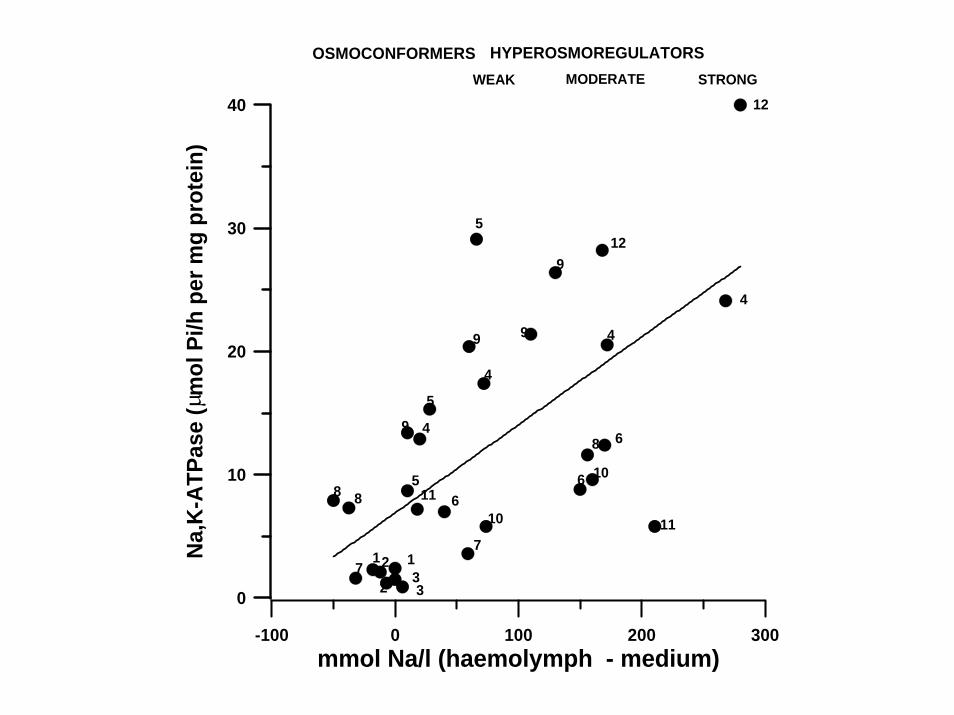
-100 0 100 200 300mmol Na/l (haemolymph - medium)
0
10
20
30
40
Na,
K-A
TPas
e (
mol
Pi/h
per
mg
prot
ein)
9
9
118 8
8
7
6
6
5
5
5
4
4
10
133
71
2
2
6
4
4
HYPEROSMOREGULATORSWEAK STRONG
OSMOCONFORMERSMODERATE
10
9
9
11
12
12
µ
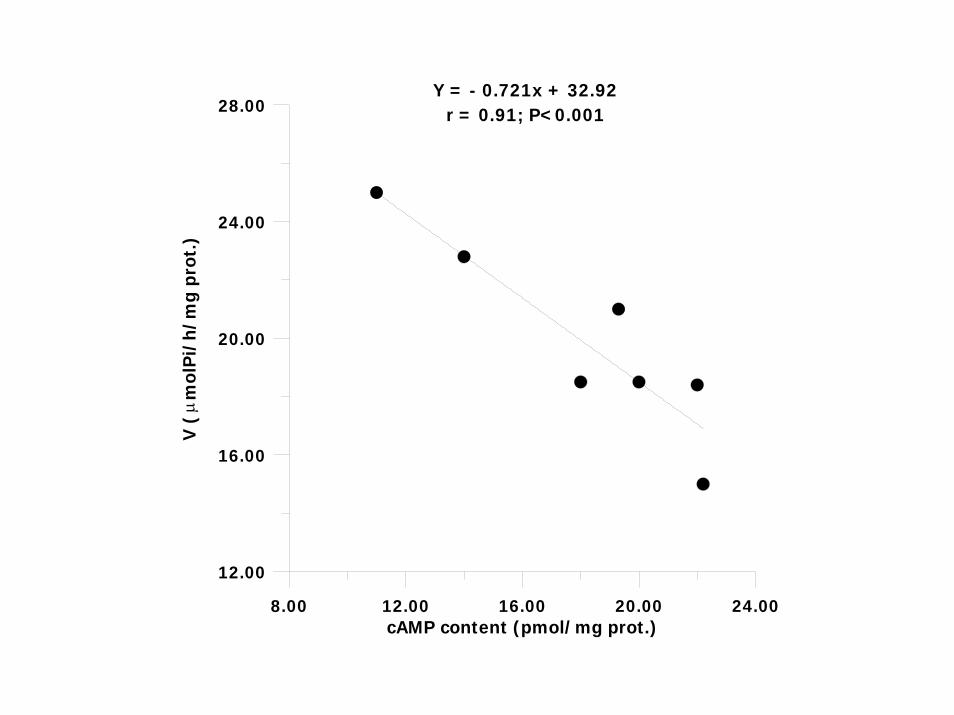
8.00 12.00 16.00 20.00 24.00cAMP content (pmol/mg prot.)
12.00
16.00
20.00
24.00
28.00
V (
m
olP
i/h/
mg
prot
.)µ
Y = - 0.721x + 32.92r = 0.91; P<0.001

Short-circuit current ( ISC) and conductance in epipodite preparation from lobster Homarus gammarus acclimated in seawater (980 mosmol/l; ISC in SW saline) and dilute seawater (600 mosmol/l; ISC in DSW).
ISC CONDUCTANCE (µA cm-2 ) (mS cm-2 )
SW acclimated -138.2±43.2 (7) 94.5±28.3 (7)
DSW acclimated -341.3±76.3 (14)** 56.5±12.1 (12)*
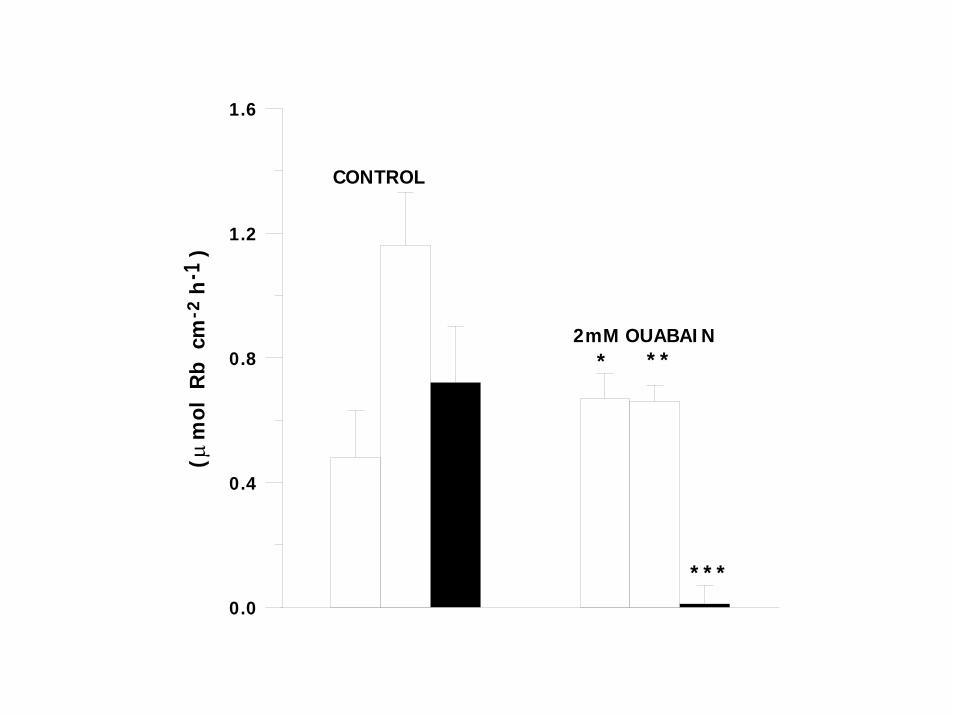
0.0
0.4
0.8
1.2
1.6
( m
ol R
b c
m
h
)CONTROL
2mM OUABAIN
µ-2
-1
* **
***
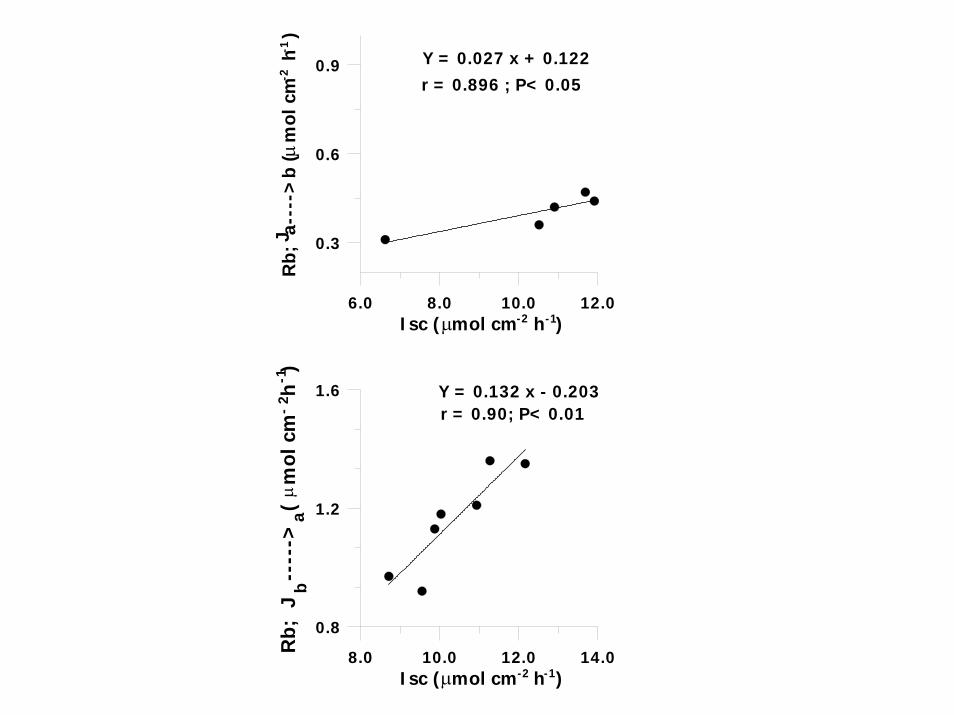
8.0 10.0 12.0 14.0 Isc ( mol cm h )
0.8
1.2
1.6
Rb;
J
----
-> (
m
ol c
m
h )
6.0 8.0 10.0 12.0 Isc ( mol cm h )
0.3
0.6
0.9
Rb;
a--
-->
b (
mol
cm
h
)µ
-1-2
µ -2 -1
µ -1-2
Jb
aµ
- 2
-1
Y = 0.027 x + 0.122r = 0.896 ; P< 0.05
Y = 0.132 x - 0.203r = 0.90; P< 0.01
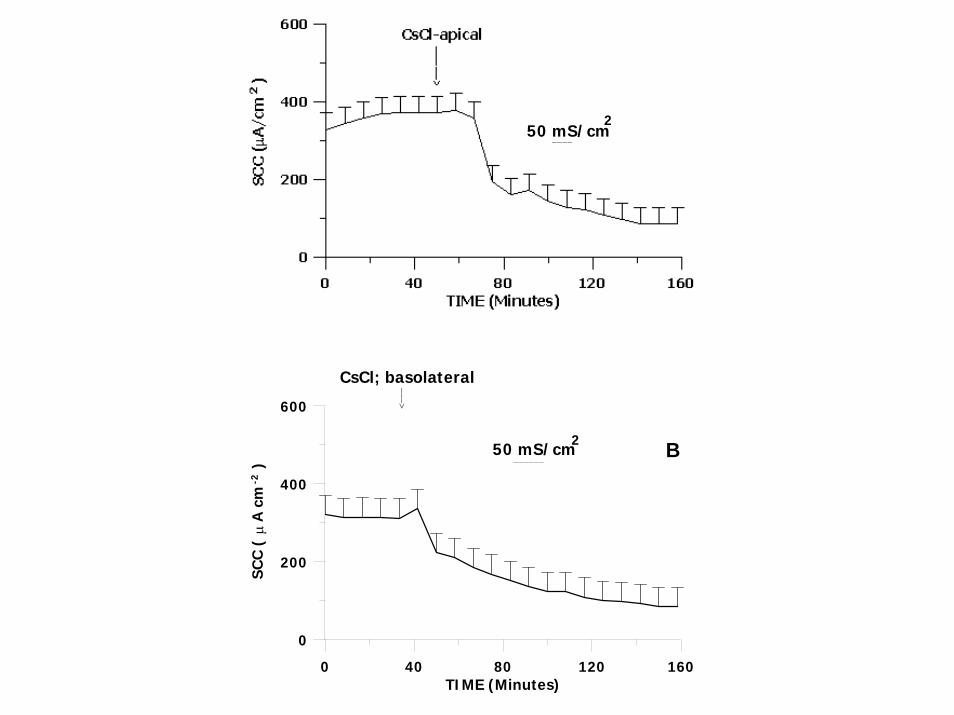
0 40 80 120 160TIME (Minutes)
0
200
400
600
SC
C (
A
cm
)
-2
CsCl; basolateral<−−−
A
B
µ
50 mS/cm2
50 mS/cm____
2
______
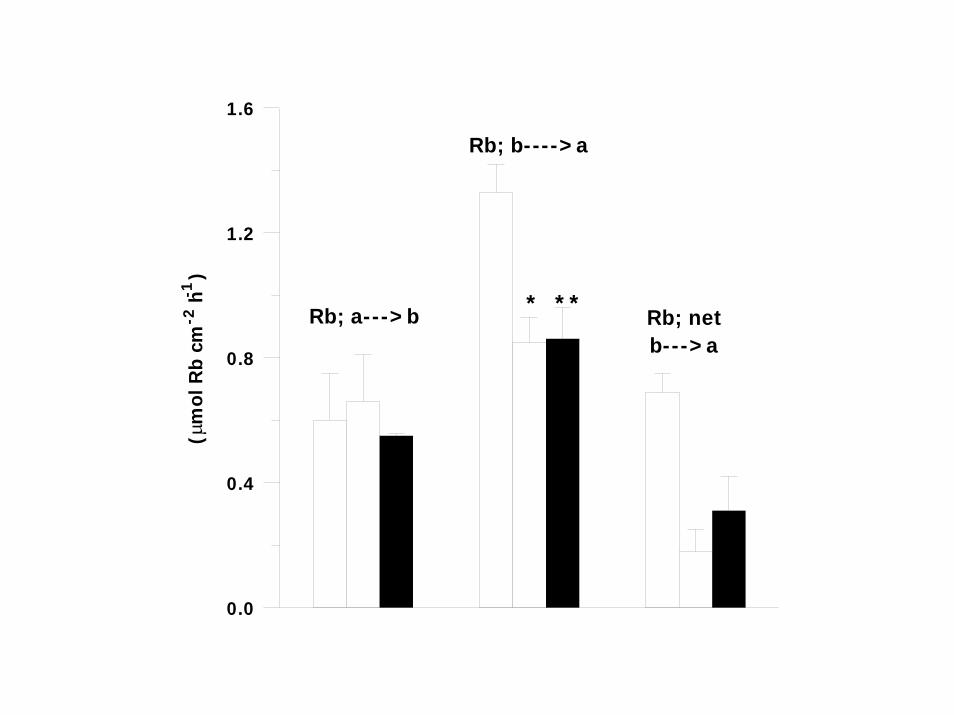
0.0
0.4
0.8
1.2
1.6
( m
ol R
b cm
h
)
Rb; a--->b
Rb; b---->a
Rb; netb--->a
µ-2
-1 * **
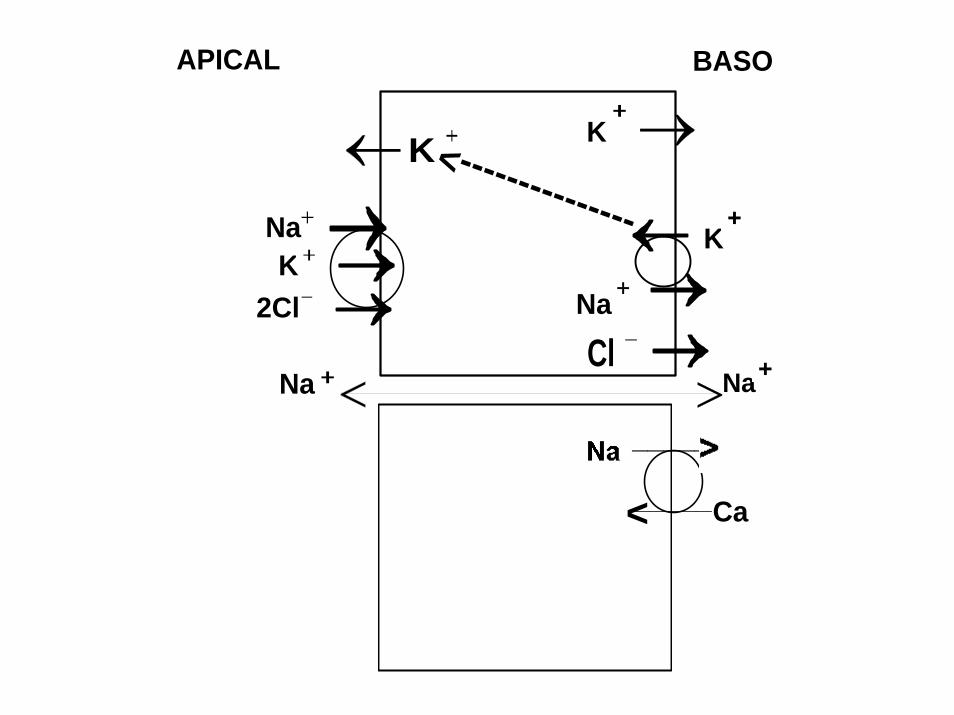
Na
Na Na
K
Na
K
K
K
Cl2Cl
APICAL BASO
--------------
->
_____
_____< Ca

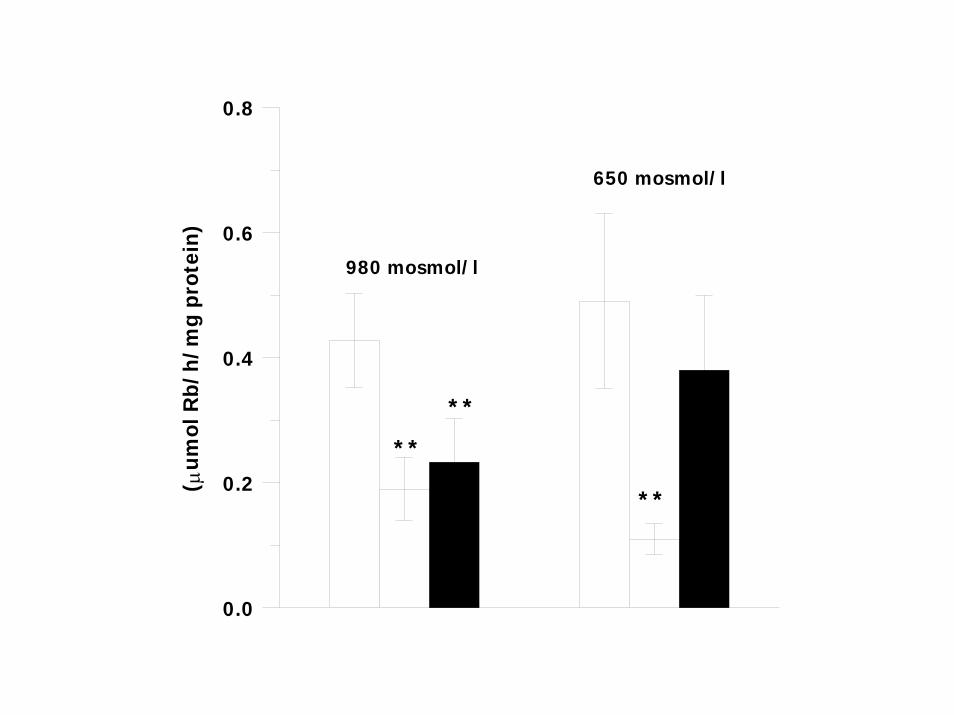
SPECIFIC ACTIVITY OF Na+ ,K+ -ATPase INCREASE IN DILUTE SEAWATER (DSW). INCREASE OF AMOUNT OF Na+,K+-ATPase PROTEIN (WESTERN BLOT)
A NEGATIVE CORRELATION BETWEEN Na,K-ATPase and cAMP IN VIVO ENZYME ACTIVITY REGULATION THROUGH cAMP –DEPENDENT PROTEIN KINASE A
SHORT CIRCUITED EPIPODITE EPITHELIUM OF LOBSTER GENERATE -341 µA/cm2 AND CONDUCTIVITY OF 57 mS/cm2.
UNIDIRECTIONAL K (86Rb) SECRETIVE FLUXES WERE 2.5 –FOLD LARGER THAN INFLUXES
OUABAIN SENSITIVE PORTION OF SECRETIVE K(Rb) EFFLUXES WAS BLOCKED BY POTASSIUM CHANNEL BLOCKER CsCl.
APICALLY LOCATED K CHANNEL IS Na+ , K+ -ATPase REGULATED MODULATING UPREGULATION OF K CONDUCTANCE
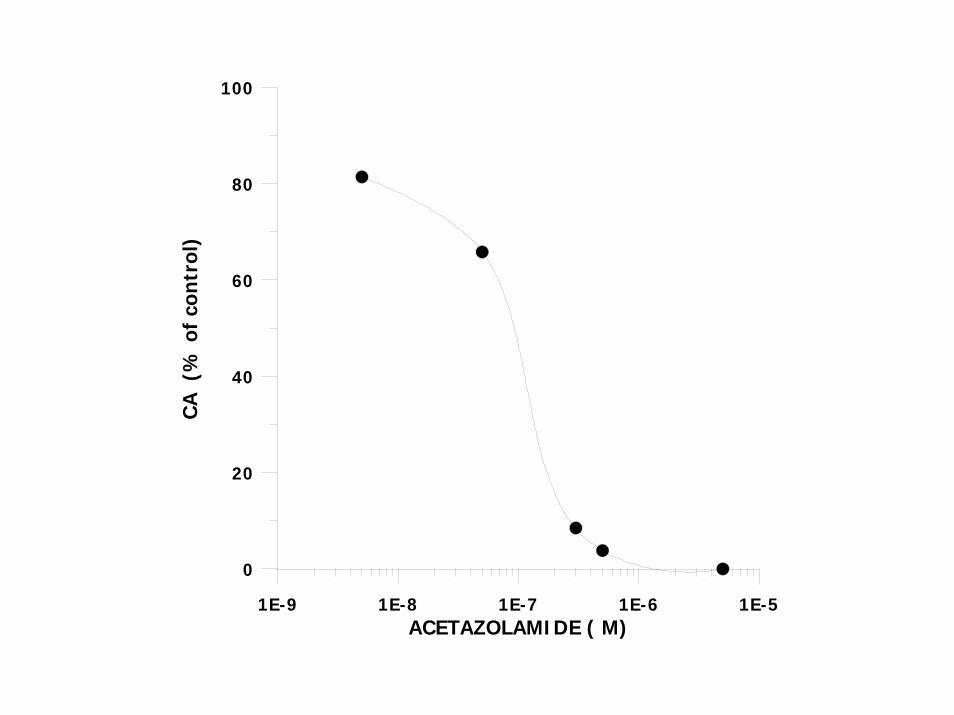
1E-9 1E-8 1E-7 1E-6 1E-5ACETAZOLAMIDE ( M)
0
20
40
60
80
100
CA
(%
of
cont
rol)

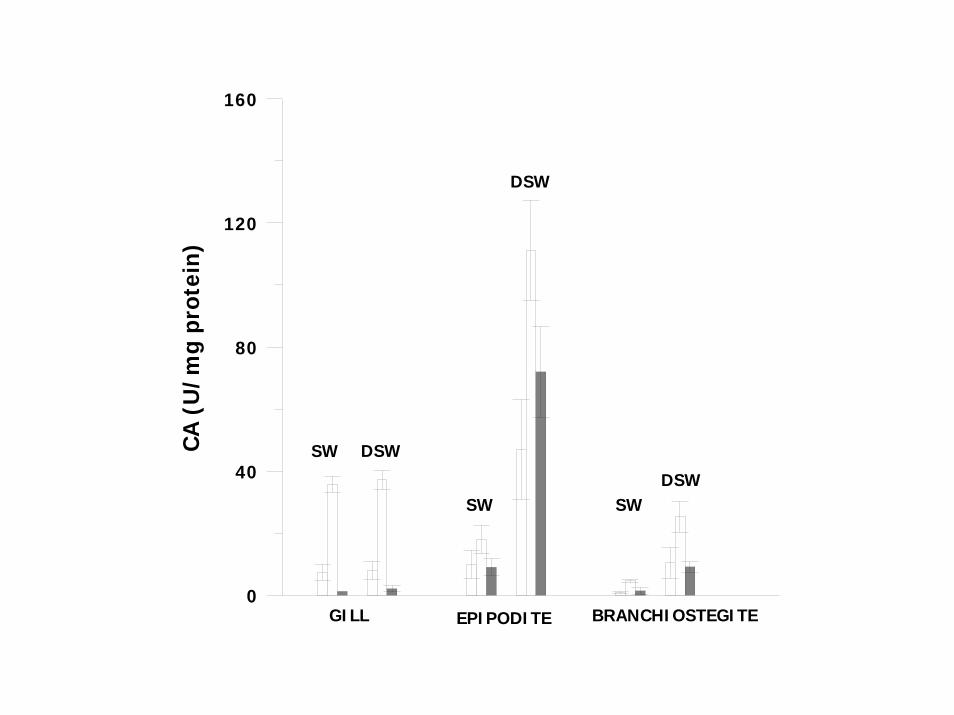
CA (U/mg protein) Purification GILLS Seawater H 7.2±2.6 (6) V 35.8±18.9 (5) 5.2 ± 0.9 (5) C 1.2± 0.3 (3) 0.3 (3)
Dilute seawater
H 8.0±3.0 (8) V 37.2±15.7 (8) 5.1±1.3 (8) C 2.1±0.4 (4)* 0.3±0.1 (4)
EPIPODITES
Sea water H 9.9±4.6 (7) V 18.0±7.9 (4) 1.9±1.4 (4) C 9.0±2.8 (7) 0.9±0.1 (7)
Dilute seawater H 46.9±16.2 (8)*** V 111.1±24.9 (5)*** 2.7±1.2 (4)
C 72.0±14.6 (6)*** 1.1±0.4 (8)
BRANCHIOSTEGITE Seawater H 0.8±0.4 (8)
V 4.6±1.9 (4) 6.1±1.6 (4) C 1.4±1.2 (3) 0.4±0.1 (3)
Dilute seawater
H 10.5±5.0 (11)*** V 27.2±4.5 (7)*** 3.6±0.5 (5)∗ C 9.1±1.9 (3)** 0.6±0.3 (3) Specific activities of carbonic anhydrase (U/mg protein) in homogenates (H),vesicles (V) and cytosolic fraction (C). Purification = ratio of specific activity of partially purified vesicles or cytosolic fraction to that of homogenates (V/H; C/H). *P<0.05; ** P<0.02; ***P<0.001
0
40
80
120
160
CA
(U
/mg
prot
ein)
GILL EPIPODITE BRANCHIOSTEGITE
SW DSW
SW
DSW
SWDSW

N M V C0
40
80
% O
F C
A A
CTI
VIT
Y
N M V C0
40
80
%
OF
CA
AC
TIV
ITY
0
40
80
% O
F C
A A
CTI
VIT
Y
N M V C
BRANCHIOSTEGITE
EPIPODITE
GILL

ACETAZOLAMIDE INHIBITS DOSE DEPENDENTLY CA IN HOMOGENATES OF EPIPODITE (IC50 = 0.12µM; MEMBRANE VESICLES AND CYTOSOL) UPON ADAPTATION TO DSW, CA IN BRANCHIOSTEGITE AND EPIPODITE HOMOGENATES, MEMBRANE FRACTION AND CYTOSOL WAS INCREASED SEVERAL FOLD OVER THESE IN SW FRACTIONS THE MAIN PROPORTION OF CA (EPIPODITE AND BRANCHIOSTEGITE) 60 –76% IN CYTOSOL . MAMBRANE FRACTION CONTAINED 7 – 14 %. REMAINDER IN MITOCHONDRIAL AND NUCLEAR FRACTIONS

Lucu Č.,Wendelaar Bonga.S. and Flik,G. Potassium secretion modulated by Na+,K+-ATPase in the epipodite of American lobster Homarus americanus. J. Physiol. (Lond.) in press
Lucu, Č. and Towle, D. Na+,K+-ATPase in Aquatic Crustacea. Comp. Biochem. Physiol. review paper ; in press
Pavičić-Hamer,D., Devescovi,M.,Lucu,Č. 2003. Activation of carbonic anhydrase in branchial cavity tissues of lobsters (Homarus gammarus) by dilute seawater exposure. J. Exp. Mar. Biol. Ecol. 287, 79-92.
Lucu,Č., Devescovi,M.,Skaramuca,B.,Kožul, V. 2000 Gill Na+,K+-ATPase in the spiny lobster Palinurus elephas and other marine osmoconformers. Adaptivness of enzymes from osmoconformity to hyperregulation. J. Exp. Mar. Biol. Ecol. 246, 162-178
Lucu,Č. and Flik, G. 1999. Na+,K+-ATPase and Na/Ca exchange activity in gills of hyperregulating Carcinus maenas. Am. J. Physiol. 45, R490-R499





























