Embed Size (px)
Citation preview

(CANCER RESEARCH 50, 2909-2916, May 15, 1990]
Parameters Affecting Tumor-specific Delivery of Anti-CD5 Antibody-Ricin A Chain
Immunotoxins in VivoOdile Rostaing-Capaillon and Pierre Casellas1
Department of Immunologi', Sanofi Recherche, rue du Professeur J. Blayac, 34082 Montpellier Cedex, France
ABSTRACT
The tumor localization of various anti-CDS ¡mmunotoxins(ITs) wasstudied in vivo in BALB/c nude mice bearing human ascitic Ichikawatumor cells. Plasma and ascitic clearance of IT as well as the number ofintact IT molecules taken up per tumor cell was measured with a highlysensitive immunoenzymometric assay. This assay allowed the quantification of as few as 400 molecules per cell, either bound to cell surfaceantigens or internalized. Tumor cell localization of an i.v.-administeredIT composed of a whole IgG linked by a disulfide bond to native ricin Achain (RTA) was extremely small. The highest binding level of this ITto ascitic cells did not exceed 1,600 molecules per cell, whereas up to25,000 molecules per cell were quantified after i.p. injection. To improveIT localization after i.v. injection, both the size of the monoclonal antibodymoiety and glycosylation of the ricin A chain were examined. Two majoradditive improvements were obtained by (a) Fc removal and (b) partialdeglycosylation of RTA. The localization of F(ab')2-deglycosylated RTAIT was 5.5 times greater than that of IgG-RTA IT, resulting from reducedplasma clearance and better extravasation of the conjugate. However, theIT targeting observed in vivo after i.v. administration was unexpectedlylower than the theoretical number of bound IT molecules, calculated fromthe concentration of soluble IT molecules in the ascitic fluid. Thisdiscrepancy was found to be related to a dramatic decrease in the antigenbinding capacity of the antibody moiety in ascitic fluid.
INTRODUCTION
Nonspecific damages to normal tissues are one of the majorproblems of conventional cancer chemotherapy. MAbs2 nowprovide high specificity for tumor recognition, but their anti-tumoral efficacy is rather limited (1). To improve their antitu-moral effect, MAbs have been conjugated to plant and bacterialtoxins, such as the phytotoxin ricin (2). However, if the wholetoxin is used, nonspecific toxicity appears, because the B chainof the toxin binds to cell surface galactose residues present onevery cell (3). Many studies have now been undertaken withconjugates prepared with the A chain alone which carries aribosome-inactivating function responsible for cell toxicity.These conjugates, called ITs, have already proved to be usefultools in vitro for the elimination of selected cell populations,and clinical trials have shown good efficacy in bone marrowpurging before transplantation (4-6). However, disappointingresults have been obtained in vivo (7). This may result partiallyfrom low site-specific delivery because of the rapid clearance ofricin A chain conjugates from the blood stream after i.v. injection. The rapid clearance is due to high mannose receptor-mediated uptake in the reticuloendothelial system of the liver(8, 9) and, in the mouse, to the recognition of a 1-3 fucoseresidues on the A chain by receptors on parenchymal liver cells(10). In accordance with these results, it has been shown that
Received 6/19/89; revised 1/8/90.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
' To whom requests for reprints should be addressed.2The abbreviations used are: MAb, monoclonal antibody; IT. immunotoxin;
RTA, ricin A chain; dRTA, deglycosylated ricin A chain: ALL, acute lympho-blastic leukemia; BAA, biological activity assay; IEMA, immunoenzymometricassay; PBS, phosphate-buffered saline; TCA, trichloroacetic acid; FITC, fluores-cein isothiocyanate; MFI. mean fluorescence intensity.
IT pharmacokinetics is significantly improved after chemicaldeglycosylation of the ricin A chain (11, 12), which reduces itsuptake by liver cells (13).
There is little information in the literature about actual invivo targeting. In the present study in vivo localization of an ITcomposed of anti-CD5 monoclonal antibody and ricin A chainlinked by a disulfide bond was investigated for 24 h after i.p.and i.v. injections. These studies were carried out in an ascitictumor model of human T-leukemia in congenitally athymicnude mice. The model is homologous to human ovarian cancerwhich develops only in the peritoneal cavity in 85 to 95% ofthe patients.
A highly precise and sensitive method was developed toquantify intact IT molecules specifically associated with tumorcells. IT molecules, bound to CDS antigen on the cell surfaceor internalized, were first solubilized by cell lysis and measuredby IEMA; the levels of soluble IT in the plasma and asciticfluid were also measured. To improve IT localization after i.v.injection, the immunoreactive part and the toxin were modified.An important benefit was obtained with IT prepared from anantibody fragment and partially deglycosylated RTA. However,a discrepancy was observed between the in vivo localization andthe theoretical binding level expected from ascitic concentrations. Analysis of IT molecules in the ascitic fluid showed thattheir antibody binding capacity was reduced after in vivo administration. These results demonstrate that the clearance rateis not the only parameter responsible for low IT targeting invivo.
MATERIALS AND METHODS
Cell Line
The Ichikawa cell line of human T-ALL origin was provided by S.Watanabe, Tokyo, Japan (14). Line IP 12/2 is a subclone of Ichikawacells and grows in vitro as well as in vivo. This cell line carries the CDSantigen, and the number of sites has been determined as 30,000 perIP 12/2 cell according to the method previously described (15). Cellswere cultured in a 5% CO2 atmosphere at 37°Cin RPMI 1640 medium
(Gibco) supplemented with 10% heat-inactivated fetal calf serum (FlowLaboratories) and antibiotics (100 Me/ml of streptomycin and 100units/ml of penicillin).
Establishment of Tumors in Mice
Six- to 8-wk-old female BALB/c (nu/nu) mice (Iffa-Credo, Lyon,France) were used. They were housed in sterile filter-top cages withsterile bedding. Mice were given injections i.p. with the Ichikawa cellsuspension (25 x IO6 cells in 0.2 ml of Hanks' solution). Ascites
appeared 20 to 25 days later in 70 to 80% of the animals and wasmaintained by serial transplantation. After ten transplantations, cultured cells were used to obtain new ascites. The ascitic volume, 20 to25 days post injection, varied from 1 to 5 ml per mouse. The asciticfluid contained 250 to 350 x IO6cells per ml, and 95% of these tumor
cells were CDS positive as determined by flow cytometry.
Monoclonal Antibodies
Two different anti-CDS MAbs were used, which recognize differentepitopes on the CDS antigen. T101 is a mouse IgG2a purchased from
2909
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

TUMOR LOCALIZATION OF RICIN A CHAIN IMMUNOTOXINS
Hybritech, Inc. (San Diego, CA) and Fl 11.98 is a mouse IgGl (Sanofi,Montpellier, France). MAbs were purified from ascitic fluid by affinitychromatography on a column of Staphylococcus aureus Protein Acoupled to Sepharose. Scatchard analysis showed that T101 andFill.98 MAbs have an affinity of 10'°ivT1for the CDS antigen.
Modification of Ricin A Chain Carbohydrate Moiety
The method used for withdrawing terminal carbohydrate structureswas a modification of the procedure described by Thorpe et al. (16).Before oxidation with sodium metaperiodate, the free thiol groups wereblocked for 30 min at 20°Cwith 0.1 M dithio-5,5'-dinitro-2,2'-diben-
zoic acid (Merck) in 125 mM phosphate buffer with 1 mM EDTAadjusted to pH 7.0 with NaOH. The solution of modified RTA wasfirst dialyzed against the same buffer to remove excess dithio-5,5'-dinitro-2,2'-dibenzoic acid and then incubated with 13 miviNalO4 and
40 mM NaCNBH3 in the dark for 10 h. Oxidation was stopped byadding one volume of 50% glycerol in 0.2 M acetate buffer, pH 6.0, to67 volumes of the RTA solution. The mixture 10 min later was thenadjusted to pH 7.0, and dialysis was performed in 125 mM phosphatebuffer with l mM EDTA, pH 7.O. The carbohydrate compositionshowed an alteration of fucose, xylose, and mannose residues but notof the TV-acetylglucosamine residues of the A chain (11). The inhibitoryactivity of RTA was preserved during the deglycosylation procedure,and dRTA was conjugated to MAb as indicated below.
Immunotoxins
Ricin A chain preparation and conjugation to MAbs were performedas previously described (17). Briefly, activated disulfide radicals wereintroduced into the MAb by reaction with A'-succinimidyl (2-pyridyI-
dithio)propionate. Following dialysis, modified MAb and RTA weremixed and allowed to react overnight before purification by gel filtrationchromatography. Different ITs were prepared using T101 MAb andFill.98 [F(ab')2] fragment. The F(ab')2 fragment was produced by
digestion with pepsin (18) and was purified by gel filtration on ACA44. These ITs are referred to as T101 (IgG)-RTA and Fl 11.98[F(ab')2]-RTA. Partially deglycosylated RTA was prepared as describedabove and conjugated to the Fl 11.98 F(ab')2 fragment. The biological
activities of ITs were determined by measuring their capacity to suppress protein synthesis in Ichikawa cells in the presence of 50 nMmonensin, as previously described (19). The biochemical characteristicsand cytotoxic activities of these ITs are summarized in Table 1.
Radioiodination
T101 MAb and T101 (IgG)-RTA conjugate were labeled by thechloramine T method as described by Hunter and Greenwood (20).Specific activities ranging from 2 to 7 pC\/n% were obtained.
Fluorometric Immunoenzymometric Assay
A fluorometric two-site immunoenzymometric assay was developedto quantify intact IT. The method comprised two steps: a specificbinding of ITs to purified sheep anti-RTA antibodies and then detectionof bound IT molecules with an anti-mouse IgG conjugated to ß-galactosidase. Anti-mouse IgG antibodies were not used in the firststep to prevent binding of autologous mouse immunoglobulins on theimmunoadsorbent. Anti-RTA antibodies were adsorbed for 20 h atroom temperature on a solid phase composed of plastic studs on lidsof Nunc TSP screening systems (Polylabo). The studs were saturatedwith fetal calf serum for 2 h. Specific binding of molecules to bemeasured was complete after 3 h of incubation at room temperature.The immunoadsorbent was washed in 10 mM PBS, pH 7.4, and dipped
Table 1 Anti-CDS IT characterization
IT MAbAffinity Coupling
Isotype (M~') ratio IC50a(M)
T101FillFill(IgG)-RTA.98[F(ab')2].98[F(ab')2]-RTA-dRTAT101FillFill.98.98IgG2aKIgGlIgGl105x5
x1010"10»1.52.31.91.2
X2.0x2.8
Xio-11Ü"1IO'13313" ICso, concentration required for 50% inhibition of the maximal proliferative
response. In vitro toxicity was determined in the presence of 50 nM monensin.
into wells containing an immunoenzymatic conjugate, i.e., F(ab')2-/3-galactosidase from sheep anti-mouse IgG, for detection of IT (Amer-sham). The enzymatic conjugate was diluted in 10 mM PBS, pH 7.4,containing 100 mM NaCl, 1 mM MgCl2, 100 mg/liter of bovine serumalbumin, and 1 g/liter of NaN3.
After incubation for 3 h at room temperature and washing of thesolid phase in PBS, the bound conjugate was incubated with 200 n\ ofa substrate solution [20 mg of 4-methyl-umbelliferyl-fi-D-galactoside(Sigma) dissolved in 2 ml of /V,jV-dimethylformamide, left for 5 h at37°C,and then diluted in 98 ml of the buffer described above]. Theenzymatic reaction was carried out in the dark for 1 h at 37"C, and 100
^1 of 100 mM glycine-NaOH buffer, pH 10.3, were added to each well.Fluorescence intensity was measured with a Fluoroscan device (FlowLaboratories, MacLean, United Kingdom). The wavelengths for excitation and emission of 4-methyl-umbelliferone, a product of the enzymatic reaction, are 355 nm and 480 nm. An absolute scale of fluorescence intensity was used, with a range of 0 to 10.000 fluorescence units.
Animal Experiments
Experiments were performed using ascitic tumor-bearing nu/nu miceweighing 32 to 38 g. One hundred fifty ^g of MAb alone or conjugatedto RTA were injected into each mouse. Equivalent amounts of fragmentmolecules were administered when F(ab')2-ITs were studied.
Localization of Radioiodinated Molecules
MAb clearance and tumor distribution were investigated using iodi-nated T101 MAb (5 to 10 x IO6cpm) in Hanks' solution, injected i.p.
or i.v. via the tail vein. Blood samples were collected by heart puncturein heparinized tubes at the times indicated in the figures, and plasmawas separated by centrifugation. Ascitic fluid was obtained from thesame mice, and IP12/2 cells in 500 p\ of fluid were immediately washed4 times with 50 ml of cold PBS containing 0.1% bovine serum albuminand 0.1% NaNj to remove unbound material. Cells were counted andviability was determined using trypan blue; more than 95% of themwere viable. The radioactivity in IO7 cells was then measured in a
gamma counter, and the number of MAb molecules bound per cell wascalculated using the specific activity of the injected solution. Plasmaand ascitic fluid samples were counted to measure total radioactivity,and the proteins were then precipitated with an equal volume of ice-cold 12.5% TCA for 30 min at 4°C.Total TCA-precipitable counts
were converted to estimate the concentration of intact molecules. Thesame procedure was followed with radiolabeled T101 (IgG)-RTA IT.
Localization of Intact IT
Studies of IT pharmacokinetics and binding after i.p. or i.v. injectionwere performed with native molecules in Hanks' solution. Blood and
ascitic fluid were collected, and the concentrations of soluble IT in theplasma and the ascitic fluid were determined by IEMA. Plasma clearance and ascitic levels are expressed as plasma and ascitic fluid concentrations versus time. IT molecules associated with ascitic tumor cellswere also quantified. Cells were first washed to remove unbound IT,and then IO7 cells were homogenized to release cell-associated ITs bytreating the cells for 30 min at 4°Cwith 500 ß\of detergent mixture
consisting of 1% Triton X-100 (Sigma), 0.5% NP-40 (Sigma), and 0.5%sodium deoxycholate (Merck) in PBS supplemented with 3.7 mg/ml ofiodoacetamide (Merck) and 2 mM phenylmethanesulfonyl fluoride(Sigma). The lysate was centrifuged for 10 min at 1000 x g beforequantification of solubilized ITs. Ninety % of the IT molecules wererecovered in the supernatant as determined using a I2il-labeled IT. The
number of IT molecules per cell (N|T/«n)was collected from the concentration (C,) of solubilized IT in the supernatant of the lysate,measured by IEMA, as follows
C. (mol/ml) x A2 x e x n,*,,
where e is the lysis efficiency (e = 0.9), A is the Avogadro number, and«ecuis the number of homogenized cells.
The theoretical number of bound molecules per cell was calculated
2910
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
TUMOR LOCALIZATION OF RICIN A CHAIN IMMUNOTOXINS
according to the "mass action law": K, = B/(C - B) X F, where K, is
the affinity constant, B is the concentration of bound MAb or IT, F isthe concentration of free molecules in the ascitic fluid determined byIEMA. and C is the concentration of CDS antigen. The number ofmolecules per cell is given by the ratio B x N/C, where N is the averagenumber of CDS antigens per cell (30,000 molecules per IP12/2 cell).
IT Binding Analysis
Antibody Binding Capacity of IT after in Vivo Administration. Theantibody binding capacity of soluble dRTA-IT from the ascitic fluidcollected after i.v. injection was studied in vitro. The percentage ofactive Fill.98 [F(ab')2)-dRTA IT was determined by measuring the
number of unbound IT molecules after immunoadsorption on an excessof target cells. IP 12/2 cells (IO8 per ml) were incubated in the presenceof low IT concentrations (around 10~'°M) for 4 h at 4°Cwith gentle
shaking. Cells were centrifuged at 400 x g, and the unbound ITs presentin the supernatant were measured by IEMA.
Antibody Binding Ability of IT in Ascitic Fluid in Vitro. The bindingcapacity of the control IT, before in vivo administration, was comparedin culture medium and pure ascitic fluid. A constant number of cells(IO6 per ml) were incubated with different concentrations of control ITfor l h at 4°C.After three washes in PBS containing 0.1% bovine
serum albumin and 0.1% sodium azide, cells were stained by indirectimmunofluorescence for l h at 4°Cusing 100 n\ of a monoclonal anti-
A chain-FITC conjugate at 1/40 dilution. The analysis was carried outon a FACSTAR PLUS cell sorter (Becton Dickinson, Sunnyvale, CA).A minimum of 10,000 IP12/2 cells was evaluated. For each sample theMFI was calculated over the 255-channel scale. Negative controls wereobtained using IT-free medium or by incubation for l h with excessnonconjugated Fill.98 before adding IT. The percentage of bindingwas evaluated by comparing each sample with the positive controlincubated at saturating doses of IT and taken as 100%. The percentageof binding was calculated as follows.
% of binding =
MFI of sample - MFI of negative controlMFI of positive control —MFI of negative control x 100.
RESULTS
Development of a Highly Sensitive Immunoenzymometric Assay for in Vivo Quantification of IT. To quantify intact IT inthe plasma and ascitic fluid of treated animals as well as ITuptake in target cells, a fluorometric immunoenzymometricassay was developed. Preliminary studies have shown that theassay can be used to determine IT concentrations in variousphysiological or experimental fluids. The detection limit inplasma and ascitic fluid was as low as 1 ng/ml of RTA. For thequantification of IT associated with tumor cells, unbound ITmolecules in the ascitic fluid were removed by washing, and IO7
cells were homogenized using a detergent mixture to releasebound ITs. The concentration of solubilized IT was measuredby IEMA to determine the number of molecules in the lysate.The standard curve was not modified when the assay wasperformed in detergent mixture or in cell lysate (IO7 cells in
500 n\). The number of IT molecules per cell was then calculatedas indicated in "Materials and Methods." As few as 400 RTA-
IT molecules could be detected per cell with this method. Thewithin-run and between-run variation coefficients of the assaywere both below 10%, in the range of 1 to 300 ng/ml of RTA.A comparative measurement of T101 (IgG)-RTA IT with theIEMA and a biological activity assay was performed. Briefly,samples containing various amounts of IT were incubated with4 x IO4 IP12/2 cells for 18 h at 37°C.The protein synthesiswas then measured by [14C]leucine incorporation into cell pro
tein during 5 h. IT concentrations were related to the inhibition291
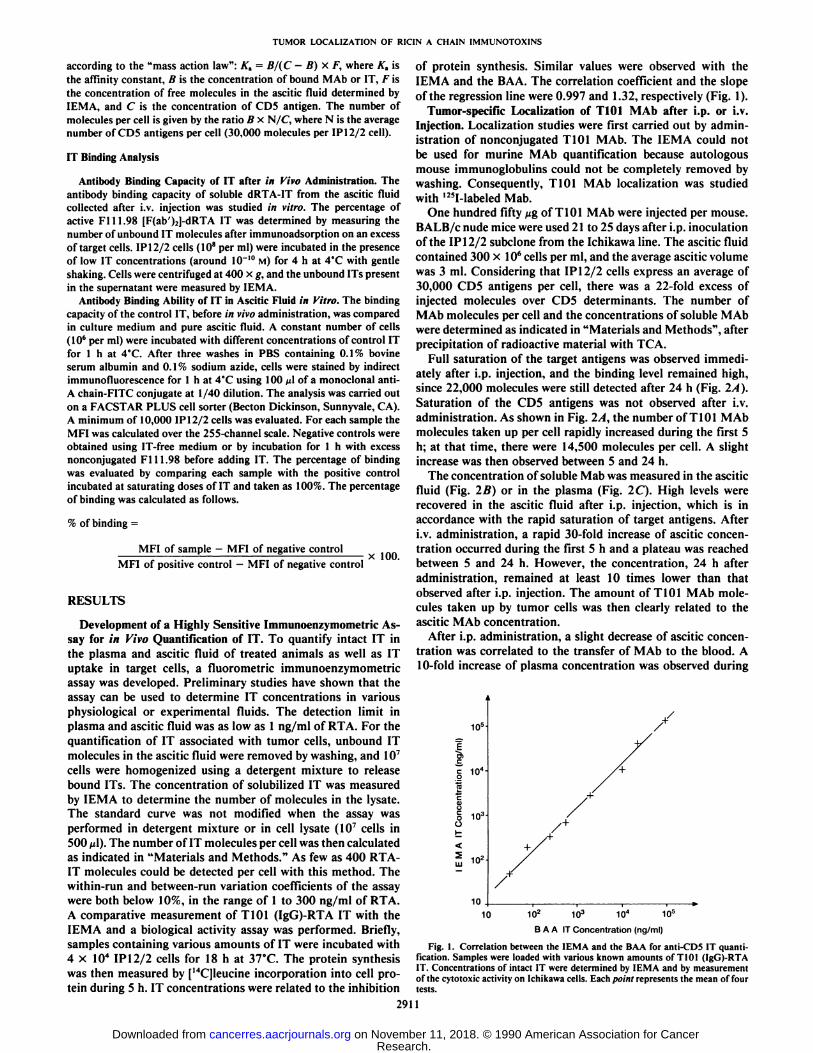
of protein synthesis. Similar values were observed with theIEMA and the BAA. The correlation coefficient and the slopeof the regression line were 0.997 and 1.32, respectively (Fig. 1).
Tumor-specific Localization of T101 MAb after i.p. or i.v.Injection. Localization studies were first carried out by administration of nonconjugated T101 MAb. The IEMA could notbe used for murine MAb quantification because autologousmouse immunoglobulins could not be completely removed bywashing. Consequently, T101 MAb localization was studiedwith 125I-labeled Mab.
One hundred fifty ng of T101 MAb were injected per mouse.BALB/c nude mice were used 21 to 25 days after i.p. inoculationof the IP 12/2 subclone from the Ichikawa line. The ascitic fluidcontained 300 x IO6cells per ml, and the average ascitic volume
was 3 ml. Considering that IP12/2 cells express an average of30,000 CDS antigens per cell, there was a 22-fold excess ofinjected molecules over CDS determinants. The number ofMAb molecules per cell and the concentrations of soluble MAbwere determined as indicated in "Materials and Methods", after
precipitation of radioactive material with TCA.Full saturation of the target antigens was observed immedi
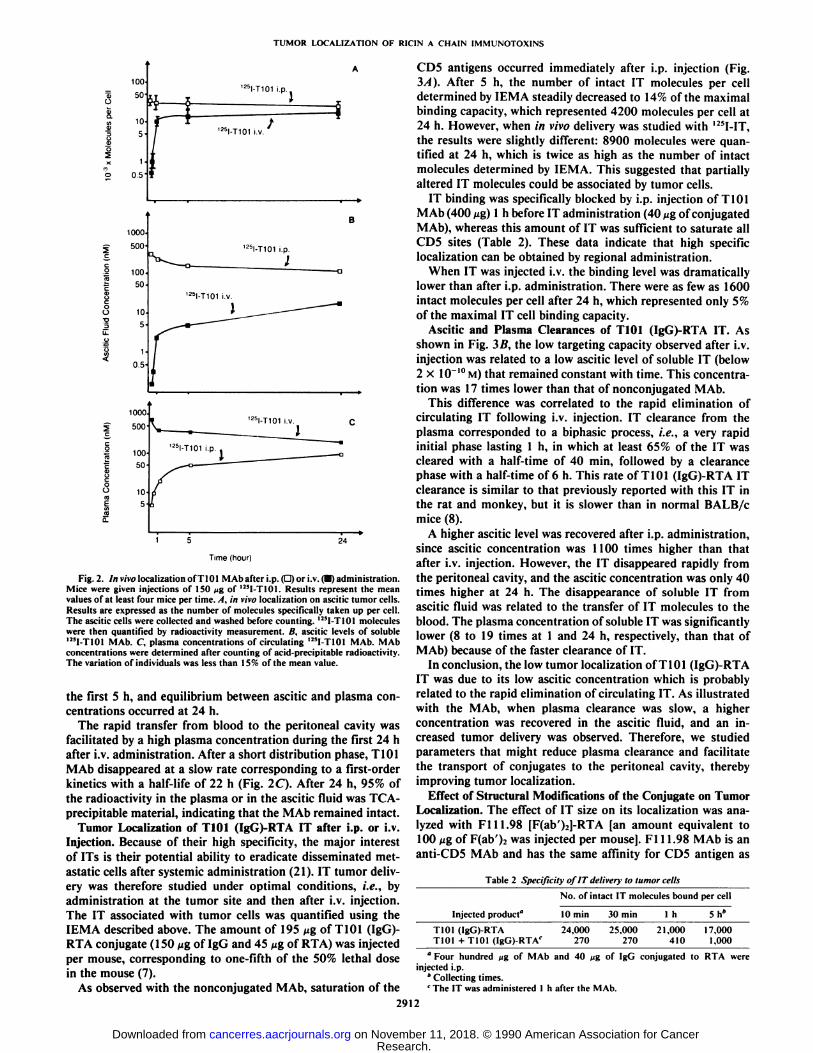
ately after i.p. injection, and the binding level remained high,since 22,000 molecules were still detected after 24 h (Fig. 2A).Saturation of the CDS antigens was not observed after i.v.administration. As shown in Fig. 2A, the number of T101 MAbmolecules taken up per cell rapidly increased during the first 5h; at that time, there were 14,500 molecules per cell. A slightincrease was then observed between 5 and 24 h.
The concentration of soluble Mab was measured in the asciticfluid (Fig. 2B) or in the plasma (Fig. 2C). High levels wererecovered in the ascitic fluid after i.p. injection, which is inaccordance with the rapid saturation of target antigens. Afteri.v. administration, a rapid 30-fold increase of ascitic concentration occurred during the first 5 h and a plateau was reachedbetween 5 and 24 h. However, the concentration, 24 h afteradministration, remained at least 10 times lower than thatobserved after i.p. injection. The amount of T101 MAb molecules taken up by tumor cells was then clearly related to theascitic MAb concentration.
After i.p. administration, a slight decrease of ascitic concentration was correlated to the transfer of MAb to the blood. A10-fold increase of plasma concentration was observed during
10=
10"
10=
102
1010 102 103 10" 105
B A A IT Concentration (ng/ml)
Fig. 1. Correlation between the IEMA and the BAA for anti-CD5 IT quantification. Samples were loaded with various known amounts of T101 (IgG)-RTAIT. Concentrations of intact IT were determined by IEMA and by measurementof the cytotoxic activity on Ichikawa cells. Each point represents the mean of fourtests.
1
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
TUMOR LOCALIZATION OF RICIN A CHAIN IMMUNOTOXINS
fU
1DO
SO
10-
5
1.0.5'
125I-T101 i.p
125I-T101 i.V.
Og
1000
500
10050
0.5'
1000
500
100
50
I-T101 i.p.
I
I!ll-T101 i.V.
I-T101 i.V.
24
Time (hour)
Fig. 2. In vivo localization of T101 MAb after i.p. (D) or i.v. (•)administration.Mice were given injections of 150 ^g of 125I-T101. Results represent the meanvalues of at least four mice per time.. I. in vivo localization on ascitic tumor cells.Results are expressed as the number of molecules specifically taken up per cell.The ascitic cells were collected and washed before counting. U5I-T101 moleculeswere then quantified by radioactivity measurement. B. ascitic levels of soluble12*1-T101MAb. C. plasma concentrations of circulating I2!I-T101 MAb. MAbconcentrations were determined after counting of acid-precipitable radioactivity.The variation of individuals was less than 15% of the mean value.
the first 5 h, and equilibrium between ascitic and plasma concentrations occurred at 24 h.
The rapid transfer from blood to the peritoneal cavity wasfacilitated by a high plasma concentration during the first 24 hafter i.v. administration. After a short distribution phase, T101MAb disappeared at a slow rate corresponding to a first-orderkinetics with a half-life of 22 h (Fig. 2C). After 24 h, 95% ofthe radioactivity in the plasma or in the ascitic fluid was TCA-precipitable material, indicating that the MAb remained intact.
Tumor Localization of T101 (IgG)-RTA IT after i.p. or i.v.Injection. Because of their high specificity, the major interestof ITs is their potential ability to eradicate disseminated met-astatic cells after systemic administration (21). IT tumor delivery was therefore studied under optimal conditions, i.e., byadministration at the tumor site and then after i.v. injection.The IT associated with tumor cells was quantified using theIEMA described above. The amount of 195 ng of T101 (IgG)-RTA conjugate (150 ¿¿gof IgG and 45 //g of RTA) was injectedper mouse, corresponding to one-fifth of the 50% lethal dosein the mouse (7).
As observed with the nonconjugated MAb, saturation of the
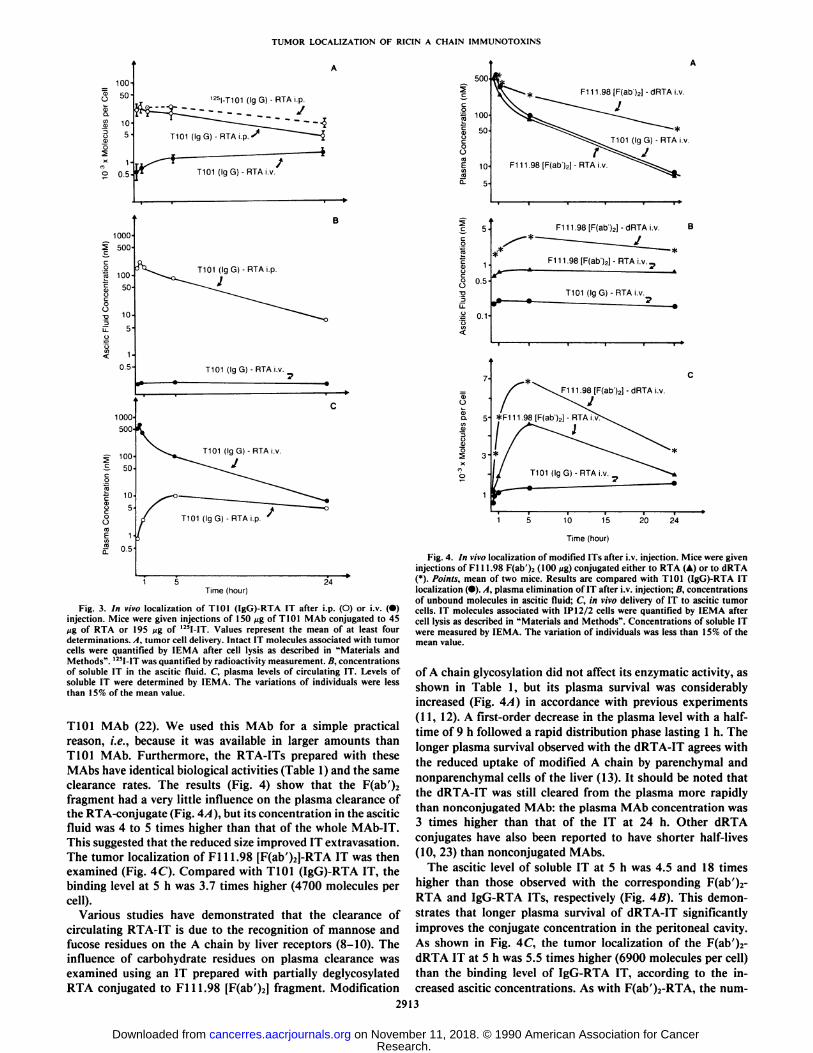
CDS antigens occurred immediately after i.p. injection (Fig.3A). After 5 h, the number of intact IT molecules per celldetermined by IEMA steadily decreased to 14% of the maximalbinding capacity, which represented 4200 molecules per cell at24 h. However, when in vivo delivery was studied with 125I-IT,
the results were slightly different: 8900 molecules were quantified at 24 h, which is twice as high as the number of intactmolecules determined by IEMA. This suggested that partiallyaltered IT molecules could be associated by tumor cells.
IT binding was specifically blocked by i.p. injection of T101MAb (400 ng) 1 h before IT administration (40 ng of conjugatedMAb), whereas this amount of IT was sufficient to saturate allCDS sites (Table 2). These data indicate that high specificlocalization can be obtained by regional administration.
When IT was injected i.v. the binding level was dramaticallylower than after i.p. administration. There were as few as 1600intact molecules per cell after 24 h, which represented only 5%of the maximal IT cell binding capacity.
Ascitic and Plasma Clearances of T101 (IgG)-RTA IT. Asshown in Fig. 3B, the low targeting capacity observed after i.v.injection was related to a low ascitic level of soluble IT (below2 x 10~'°M) that remained constant with time. This concentra
tion was 17 times lower than that of nonconjugated MAb.This difference was correlated to the rapid elimination of
circulating IT following i.v. injection. IT clearance from theplasma corresponded to a biphasic process, i.e., a very rapidinitial phase lasting 1 h, in which at least 65% of the IT wascleared with a half-time of 40 min, followed by a clearancephase with a half-time of 6 h. This rate of T101 (IgG)-RTA ITclearance is similar to that previously reported with this IT inthe rat and monkey, but it is slower than in normal BALB/cmice (8).
A higher ascitic level was recovered after i.p. administration,since ascitic concentration was 1100 times higher than thatafter i.v. injection. However, the IT disappeared rapidly fromthe peritoneal cavity, and the ascitic concentration was only 40times higher at 24 h. The disappearance of soluble IT fromascitic fluid was related to the transfer of IT molecules to theblood. The plasma concentration of soluble IT was significantlylower (8 to 19 times at 1 and 24 h, respectively, than that ofMAb) because of the faster clearance of IT.
In conclusion, the low tumor localization ofTlOl (IgG)-RTAIT was due to its low ascitic concentration which is probablyrelated to the rapid elimination of circulating IT. As illustratedwith the MAb, when plasma clearance was slow, a higherconcentration was recovered in the ascitic fluid, and an increased tumor delivery was observed. Therefore, we studiedparameters that might reduce plasma clearance and facilitatethe transport of conjugates to the peritoneal cavity, therebyimproving tumor localization.
Effect of Structural Modifications of the Conjugate on TumorLocalization. The effect of IT size on its localization was analyzed with Fill.98 [F(ab')i]-RTA [an amount equivalent to100 ßgof F(ab'); was injected per mouse]. Fl 11.98 MAb is an
anti-CDS MAb and has the same affinity for CDS antigen as
Table 2 Specificity of IT deliver)- to tumor cells
No. of intact IT molecules bound per cell
Injected product" 10 min 30 min l h 5h"
T101 (IgG)-RTAT101 +T101 (IgG)-RTAf
24,000270
25.000270
21.000410
17.0001.000
°Four hundred /jg of MAb and 40
injected i.p.* Collecting times.c The IT was administered I h after the MAb.
of IgG conjugated to RTA were
2912
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
TUMOR LOCALIZATION OF RICIN A CHAIN IMMUNOTOXINS
'25I-T101 (IgG) - RTAi.p.
--- . J
_ 100°5 500
5 100-
OB
< 10.5
1000'
500
^ 100S 50
T101 (IgG)- RTAi.p.
T101 (lg G) - RTAi.v. _
Time (hour)
Fig. 3. In vivo localization of T101 (IgG)-RTA IT after i.p. (O) or i.v. (•)injection. Mice were given injections of 150 ng of T101 MAb conjugated to 45jig of RTA or 195 »jgof 1!!I-IT. Values represent the mean of at least fourdeterminations. A, tumor cell delivery. Intact IT molecules associated with tumorcells were quantified by IEMA after cell lysis as described in "Materials andMethods". 125I-ITwas quantified by radioactivity measurement. B, concentrations
of soluble IT in the ascitic fluid. C, plasma levels of circulating IT. Levels ofsoluble IT were determined by IEMA. The variations of individuals were lessthan 15% of the mean value.
T101 MAb (22). We used this MAb for a simple practicalreason, i.e., because it was available in larger amounts thanT101 MAb. Furthermore, the RTA-ITs prepared with theseMAbs have identical biological activities (Table 1) and the sameclearance rates. The results (Fig. 4) show that the F(ab')2
fragment had a very little influence on the plasma clearance ofthe RTA-conjugate (Fig. 4A), but its concentration in the asciticfluid was 4 to 5 times higher than that of the whole MAb-IT.This suggested that the reduced size improved IT extravasation.The tumor localization of Fl 11.98 [F(ab')2]-RTA IT was thenexamined (Fig. 4C). Compared with T101 (IgG)-RTA IT, thebinding level at 5 h was 3.7 times higher (4700 molecules percell).
Various studies have demonstrated that the clearance ofcirculating RTA-IT is due to the recognition of mannose andfucose residues on the A chain by liver receptors (8-10). Theinfluence of carbohydrate residues on plasma clearance wasexamined using an IT prepared with partially deglycosylatedRTA conjugated to Fill.98 [F(ab')2J fragment. Modification
F111.98[F(ab')2]-dRTAi.v.
1
5-
0.5-
0.1
F111.98[F(ab')2]-dRTAi.v. B
F111.98[F(ab')2]- RTAi.v. _
T101 (IgG) -RTAi.v._
Ooo- 5
lI
J=111.98 [F(ab')2] - dRTA i.v
10 15
Time (hour)
20 24
Fig. 4. In rivo localization of modified ITs after i.v. injection. Mice were giveninjections of Fl 11.98 F(ab')2 (100 pg) conjugated either to RTA (A) or to dRTA(*). Points, mean of two mice. Results are compared with T101 (IgG)-RTA ITlocalization (•).A, plasma elimination of IT after i.V.injection; B, concentrationsof unbound molecules in ascitic fluid; C, in vivo delivery of IT to ascitic tumorcells. IT molecules associated with IP12/2 cells were quantified by IEMA aftercell lysis as described in "Materials and Methods". Concentrations of soluble IT
were measured by IEMA. The variation of individuals was less than 15% of themean value.
of A chain glycosylation did not affect its enzymatic activity, asshown in Table 1, but its plasma survival was considerablyincreased (Fig. 4A) in accordance with previous experiments(11, 12). A first-order decrease in the plasma level with a half-time of 9 h followed a rapid distribution phase lasting 1 h. Thelonger plasma survival observed with the dRTA-IT agrees with
the reduced uptake of modified A chain by parenchyma! andnonparenchymal cells of the liver (13). It should be noted thatthe dRTA-IT was still cleared from the plasma more rapidly
than nonconjugated MAb: the plasma MAb concentration was3 times higher than that of the IT at 24 h. Other dRTAconjugates have also been reported to have shorter half-lives(10, 23) than nonconjugated M Abs.
The ascitic level of soluble IT at 5 h was 4.5 and 18 timeshigher than those observed with the corresponding F(ab')2-
RTA and IgG-RTA ITs, respectively (Fig. 4B). This demonstrates that longer plasma survival of dRTA-IT significantlyimproves the conjugate concentration in the peritoneal cavity.As shown in Fig. 4C, the tumor localization of the F(ab')2-
dRTA IT at 5 h was 5.5 times higher (6900 molecules per cell)than the binding level of IgG-RTA IT, according to the increased ascitic concentrations. As with F(ab')i-RTA, the num-
2913
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

TUMOR LOCALIZATION OF RICIN A CHAIN 1MMUNOTOXINS
ber of molecules per cell notably increased during the first 5 h,and approximately half of the molecules detected at 5 h disappeared between 5 and 24 h.
Comparison of Experimental and Theoretical IT Binding toTumor Cells. We next compared the localization in vivo, experimentally determined, with the theoretical number of MAb orIT molecules bound per cell calculated from the ascitic fluidconcentrations. After i.v. injection, a surprising discrepancywas observed between experimental and theoretical values. Thedifference was noted with all the ITs tested and the nonconju-gated MAb. The experimental number of bound molecules percell 30 min after injection (Table 3) was 10 to 30 times lowerthan the expected level. Similar results were obtained at different times after injection (from 30 min to 24 h after injection;data not shown). These contradictory results obtained after i.v.injection could be due either to a decrease in MAb affinity invivo or to a partial inactivation of the antibody binding site.When MAb or IT was administered i.p., the experimental datawere well correlated with the theoretical values, but the asciticconcentrations of soluble molecules were 50 times higher thanthe saturating concentrations. This large excess could mask thepartial alteration of the MAb moiety.
Antibody Binding Capacity of IT. To discriminate between adecrease in MAb affinity and a partial inactivation of the MAbbinding site, we studied the binding capacity of ascitic fluidsamples containing Fill.98 [F(ab')2]-dRTA IT molecules, 30
min after i.v. injection. Samples were first incubated with anexcess of target IP12/2 cells (IO8 cells/ml). Unbound IT was
then quantified in the supernatant by IEMA. As shown in Table4, 87% of the IT molecules from the ascitic fluid remainedunbound, compared with 15% of the control IT, before in vivoadministration. These data indicate that a large portion (73%)of the IT molecules lost their antibody binding capacity after invivo injection, suggesting that the low tumor localization afteri.v. administration could be also related to inactivation of theantibody binding site.
Influence of Ascitic Fluid on IT Binding. We next examinedwhether this inhibition could be due to the ascitic fluid itself.The binding capacity of control Fl 11.98 [F(ab'):]-dRTA con
jugate to IP12/2 cells was studied in vitro in pure ascitic fluidor in culture medium. Bound molecules were revealed with ananti-A chain-FITC conjugate by flow cytometry. As shown inFig. 5, the dose-binding curve was dramatically shifted to higherconcentrations when binding occurred in ascitic fluid. Sevenfold higher doses were required in the IP 12/2 ascitic fluid thanin culture medium, to bind 50% of the CDS antigens. Toexamine the possibility that inactivation of MAb might be
related to the presence of a specific inhibitor such as solubleCDS antigens, the binding was performed in the ascitic fluidgenerated by a nonrelevant hybridoma producing anti-myosinMAb. A weaker but significant 4-fold inhibition was observed,suggesting that nonspecific inhibition also occurred in asciticfluid. These results strongly suggest that the inhibition ofantibody binding in vivo is induced by ascitic fluid.
DISCUSSION
ITs have been designed for in vivo cancer therapy. The toxicsubstance conjugated to MAbs directed against tumor-associated antigens is specifically targeted to tumor cells and doesnot damage normal tissues. However, despite their high in vitrocytotoxicity, the therapeutic efficacy of ITs has rarely beendemonstrated in vivo (21, 24, 25). Low tumor localization couldbe an important reason for this poor activity.
The cytotoxic efficacy of ITs is strongly dependent on thenumber of molecules bound to tumor cells. Laurent et al. (26)have demonstrated that fresh leukemia cell sensitivity to T101(IgG)-RTA IT is related to CDS antigen density. More recently,Preijers et al. (27) showed a strong correlation between theCDS density of different cell lines and the cytotoxic efficacy ofthe same IT. These studies suggest that a threshold level isrequired for IT activity and that IT efficiency increases with ITbinding. These in vitro indications should be true in vivo aswell.
Biodistribution studies (23, 28) have been carried out withlabeled ITs, but the method did not provide any informationabout the actual integrity of bound molecules. We thereforereaddressed the question using an IEMA designed to quantifythe immunologically intact IT associated with target cells. Byassaying IT in cell lysate, both the IT bound to the cell surfaceand the IT internalized in the cell were measured, which makesthis method a choice procedure for quantitative localizationanalysis.
The study was performed in an ascitic model involving alarge tumor burden of about 900 x IO6 Ichikawa cells dissemi
nated in the peritoneal cavity of BALB/c nude mice, with avery high cell density (300 x IO6 per ml). IT localization was
compared with the targeting capacity of nonconjugated MAbafter i.p. or i.v. injection.
i.p. Administration. A complete saturation of CDS antigenson tumor cells was achieved when therapeutic doses of IT orMAb were injected i.p. within the tumor burden. This is consistent with: (a) the high affinity of the MAbs used (10'°M~'),
(b) the 22-fold excess of molecules injected over target antigens.
Table 3 Comparison of the theoretical and experimental numbers of MAb or IT molecules bound per cell in vivo
T101i.p.T101(IgG)-RTAi.p.T101
i.v.T101(IgG)-RTAi.v.FllI.98[F(ab')2J-RTAi.v.F111.98[F(ab')2]-dRTAi.v.Ascitic
fluid Theoreticalconcentration value
(F)°(mol/liter)(N*«J*2.0
x10-2.1x10"2.9
x10-1.9x10-6.5x 10-30,00030.0000
22,250019.5800
22,9401.8x IO' 27,130Experimental
value(N„„.)c34,00025,0008505901,4003,100NItao./N..p.ratio0.91.226.233.216.48.8
°Concentrations were measured by IEMA.6 The number of bound molecules was calculated from ascitic fluid concentrations measured 30 min after injection, as follows.
_ K. x n x FN""°~ i + A:,x F
where A",is the affinity constant of the antibody moiety, and n is the average number of CD5 antigens per cell.e The number of molecules per cell was determined by IEMA as indicated in "Materials and Methods."
2914
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

TUMOR LOCALIZATION OF RICIN A CHAIN 1MMUNOTOXINS
Table 4 Antibody binding capacity of Fl 11.98 [F(ab')J-dRTA IT
IT concentration"IT concentration" remaining in cell
before in vitro supernatant % of unboundbinding (ng/ml) (ng/ml) molecules
IT controlIT from ascitic fluid*
207
3.16.1
14.887.1
a Concentrations were determined by IEMA.* The ascitic fluid was collected 30 min after i.v. injection.
10-
IT Concentration (M)Fig. 5. IT binding on IP12/2 cells in vitro. The binding of FI 11.98 (F(ab');]-
dRTA IT was examined in culture medium (x), in IP12/2 ascitic fluid generatedby i.p. injection of IPI2/2 cells (*). and in nonrelevant ascitic fluid obtained byi.p. injection of an anti-myosin hybridoma (+). Bound IT molecules were detectedwith an FITC-anti-A chain conjugate after incubation of IP 12/2 cells with variousconcentrations of IT at 4'C. Fluorescence-activated cell sorter analysis andcalculations were performed as described in "Materials and Methods". The
variation of individuals was less than 10% of the mean value.
and (c) the easy diffusion of molecules in the peritoneal cavity.The saturation obtained with the IT shortly after injection wastransient, and the number of molecules associated with tumorcells, as determined by IEMA, steadily decreased with timedespite saturating concentrations of free IT in the ascitic fluid.As suggested by results obtained with I25l-labeled IT, which
showed a higher IT uptake than observed with IEMA, partialdegradation of IT might have occurred, probably after endocy-tosis.
i.v. Administration. The targeting capacity of IT injected i.v.was very limited, in contrast with the high tumor localizationobserved with T101 MAb administered by the same route.These observations are in accordance with the increased lifespan of peritoneal tumor-bearing mice after i.p. injection of IT,whereas i.v. injection had no effect on tumor growth (29-32).The difference between the IT and the nonconjugated MAb wasdue to the rapid transfer of T101 MAb from the blood to theascitic fluid resulting in ascitic fluid concentrations 25 or 100times higher than that of IT at 5 and 24 h. The poor extravasation of IT was mainly due to its lower plasma concentration,but it could also be related to its large molecular size. Indeed,removal of the Fc fragment of the antibody allowed fastertransfer of the conjugate to the peritoneal cavity, and thereforebetter tumor delivery. The binding level of Fill.98 [F(ab')2]-
RTA IT was 3.7 times higher than that of T101 (IgG)-RTA IT,at equivalent affinity constants.
As expected, the reduced plasma clearance of Fill.98[F(ab')2]-dRTA IT facilitated its extravasation and localization,
which was 1.5 times higher at 5 h than the targeting of thecorresponding IT with native RTA. Hence, the combination ofthe F(ab')2 fragment and modified RTA resulted in a 5.5-fold
improvement in tumor delivery compared with that of IgG-
RTA IT.It should be noted that, although F(ab')z-dRTA IT was the
most effective IT at localizing, its binding level was still lowerthan that of T101 MAb. This difference was probably relatedto the longer survival of the MAb in the plasma, suggestingthat the chemically removed carbohydrate residues were not theonly element responsible for plasma clearance of RTA-IT. Thiswas confirmed using IT prepared with nonglycosylated recombinant RTA whose plasma clearance was similar to that ofdRTA-IT (data not shown). The instability of the disulfide bondbetween the MAb and the toxin may account for the differencein the clearance rate with the Mab (33, 34). The use of differentcross-linking agents might overcome this drawback (35, 36).
An important and unexpected finding in this study is theinhibition of specific binding to CDS antigens detected in vivo.We found that the low binding capacity in vivo was related torapid inactivation of the antibody binding site of most IT orMAb molecules by the ascitic fluid. A nonspecific inhibition ofIT binding was observed in vitro in nonrelevant ascitic fluid,but the inhibition was greater in IP 12/2 ascitic fluid, suggestingthe presence of a specific inhibiting factor, such as a free CDSantigen shed from IP 12/2 cells. This particular point wasexamined by the quantification of free CDS molecules in theascitic fluid, using a double determinant immunoassay with twoanti-CDS MAbs directed against different epitopes (T101 MAband Fill.98 MAb). The concentration of free CDS antigensdetected in the ascitic fluid (10~'°M; data not shown) is not
sufficient to explain the specific binding inhibition in vivo.However, we cannot exclude the possibility of a shedding ofaltered CDS molecules that are no longer simultaneously recognized by both T101 and Fl 11.98 MAbs. In conclusion, theseresults clearly demonstrate that MAb and IT localization invivo are impaired by specific and/or nonspecific inhibition ofantibody binding capacity.
Whatever the factor involved in inactivating the antibodybinding site, which remains to be characterized, it is clearlyimportant to examine whether this inactivation occurs in bloodand with all ITs. It should be noted that a similar decrease inantibody binding capacity has previously been described (a) inmouse plasma and (b) with another IT anti-murine IgD (23).However, when high IT concentrations are achieved in vivo,this inhibition in ascitic fluid can be overcome or significantlyreduced, as demonstrated by i.p. injection. We also show herethat higher concentrations can be obtained by structural modifications that do not alter the cytotoxic properties of the conjugate. These results confirm the importance of studying theparameters that influence IT extravasation and IT targeting,which should lead to further improvements in the therapeuticefficacy of IT.
ACKNOWLEDGMENTS
We thank H. Vidal and D. Dussossoy for preparing ITs. We aregrateful to P. Carayon for expert fluorescence-activated cell sortinganalyses. We also thank C. Clausner for excellent secretarial assistance.
REFERENCES
1. Oldham. R. K. Monoclonal antibodies: does sufficient selectivity to cancercells exist for therapeutic application? J. Biol. Response Modifiers. 6: 227-234, 1987.
2. Jansen, F. K.. Blythman, H. E., Carrière, D.. Casellas. P., Gros. O.. Gros,P., Laurent. J. C.. Paolucci. F.. Pau. B.. Poncelet. P.. Richer, G., Vidal. H.,and Voisin, G. A. Immunotoxins: hybrid molecules combining high specificity and potent cytotoxicity. Immunol. Rev.. 62: 185-216, 1982.
2915
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

TUMOR LOCALIZATION OF RICIN A CHAIN IMMUNOTOX1NS
3. Youlc, R. J., and Colombatti, M. Immunotoxins: monoclonal antibodieslinked to toxic proteins for bone marrow transplantation and cancer therapy.In: i. A. Roth (ed.). Monoclonal Antibodies in Cancer: Advances in Diagnosis 22.and Treatment, pp. 173-213. Mount Kisco. NY': Futura Publishing Com
pany, Inc.. 1986.4. Casellas, P., Canat, X., Fauser, A. A., Gros, O., Laurent, G., Poncelet, P.,
and Jansen. F. K. Optimal elimination of leukemic T-cells from human bone 23.marrow with TlOl-ricin A-chain immunotoxin. Blood. 65: 289-297, 1985.
5. Fauser, A. A., Shustik, C., Langleben, A., Laurent, G., Kanz. L.. Spurii, G.M., Price, G., Ahlgren, P. D., and Cooper, B. A. T-cell depletion with ricinA-chain-T101 in allogeneic bone marrow transplantation to prevent severe 24.graft-vm«j-host disease. Clin. Invest. Med., //: 40-46, 1988.
6. Casellas, P., Ravel, S., Bourne, B. J. P.. Derocq, J. M., Jansen, F. K.,Laurent, G.. and Gros, P. T-lymphocyte killing by TlOl-ricin A-chainimmunotoxin: pH-dependent potentiation with lysosomotropic amines. 25.Blood, 72: 1197-1202, 1988.
7. Griffin, T. W.. Morgan, A. C., and Blythman, H. E. Immunotoxin therapy:assessment by animal models. In: A. E. Frankel (ed.), Immunotoxins. pp.433-455. Not-well, MA: Kleuwer Academic Publishers, 1988.
8. Bourne, B. J. P., Casellas. P.. Blythman. H. E., and Jansen, F. K. Study ofthe plasma clearance of antibody-ricin A-chain immunotoxins: evidence for 26.specific recognition sites on the A-chain that mediate rapid clearance of theimmunotoxin. Eur. J. Biochem., 155: 1-10, 1986.
9. Byers, V. S., Pimm, M. V., Pawluczyk, I. Z. A., Lee, H. M., Scannon, P. J.. 27.and Baldwin. R. W. Biodistribution of ricin toxin A-chain-monoclonal antibody 791T/36 immunotoxin and influence of hepatic blocking agents. CancerRes., 47: 5277-5283, 1987.
10. Blakey. D. C., Skilleter, D. N., Price, R. J., and Thorpe. P. E. Uptake of 28.native and deglycosylated ricin A-chain immunotoxins by mouse liver paren-chymal and non-parenchymal cells in vitro and in vivo. Biochim. Biophys.Acta, 96«:172-178. 1988.
11. Casellas. P., Bourrié.B.. Canat, X.. Blythman, H. E., and Jansen. F. K.Pharmacology of antibody-ricin A-chain immunotoxins. In: Membrane-me- 29.dialed Cytotoxicity. pp. 185-196. New York: A. R. Liss. Inc., 1987.
12. Blakey, D. C, Watson, G. J., Knowles, P. P.. and Thorpe, P. E. Effect ofchemical deglycosylation of ricin A-chain on the in vivo fate and cytotoxicactivity of an immunotoxin composed of ricin A-chain and anti-Thy 1.1 30.antibody. Cancer Res., 47: 947-952, 1987.
13. Blakey, D. C., and Thorpe, P. E. Effect of chemical deglycosylation on thein vim fate of ricin A-chain. Cancer Drug Deliv., 3: 189-196. 1986. 31.
14. Watanabe. S.. Shimosato. Y., Kameya, T., Kuroki, M., Kitahara, T., Minato,K., and Shimoyama, M. Leukemic distribution of a human acute lymphocyticleukemia cell line (Ichikawa strain) in nude mice conditioned with whole-body irradiation. Cancer Res., 38: 3494-3498. 1978. 32.
15. Poncelet, P., and Carayon, P. Cytofluorometric quantification of cell surfaceantigens by indirect immunofluorescence using monoclonal antibodies. J.Immunol. Methods, 85: 65-74, 1985. 33.
16. Thorpe, P. E., Detre, S. I., Foxwell, B. M. J., Brown, A. N. F., Skilleter. D.N., Wilson, G., Forrester, J. A., and Stirpe, F. Modification of the carbohydrate in ricin with metaperiodate-cyanoborohydride mixtures. Effects ontoxicity and in vivo distribution. Eur. J. Biochem., 147: 197-206. 1985.
17. Gros, O., Gros, P.. Jansen, F. K., and Vidal, H. Biochemical aspects of IT 34.preparation. J. Immunol. Methods, 81: 283-297. 1985.
18. Stanworth. D. R.. and Turner. M. W. Immunochemical analysis of immu-noglobulins and their subunits. In: D. M. Weir (ed.). Handbook of Experimental Immunology. Chap. 10. p. 1. London: Blackwell Scientific, 1973. 35.
19. Casellas, P., Bourrié,B. J. P., Gros, P., and Jansen, F. K. Kinetics ofcytotoxicity induced by immunotoxins. Enhancement by lysosomotropicamines and carboxylic ionophores. J. Biol. Chem.. 259.-9359-9364. 1984.
20. Hunter, W. M.. and Greenwood. F. C. Preparation of iodine-131-labeled 36.human growth hormone of high specific activity. Nature (Lond.). 194: 495-496, 1962.
21. Roth, J. A., Ames. R. S., Fry, K., Lee, H. M., and Scannon, P. J. Mediationof reduction of spontaneous and experimental pulmonary métastasesby ricin
A-chain immunotoxin 45-2D9-RTA with potentiation by systemic monensinin mice. Cancer Res.. 48: 3496-3501, 1988.Carrière, D., Fontaine, C.. Benoit, J. M., Thurneyssen. O.. and Carrel, S.Four different epitopes on the CDS (67 kDa) antigen are defined by MoAbsof the workshop CDS panel. Tissue antigens. Histocompat. Immunogenet.,33: 102, 1989.Fulton. R. J., Tucker. T. F., Vitetta, E. S., and Uhr, J. W. Pharmacokineticsof tumor-reactive immunotoxins in tumor-bearing mice: effect of antibodyvalency and deglycosylation of the ricin A-chain on clearance and tumorlocalization. Cancer Res., 48: 2618-2625, 1988.Thorpe, P. E., Brown. A. N. F., Brenner, J. A. G., Foxwell. B. M. J., andStirpe. F. An immunotoxin composed of monoclonal anti-Thy 1.1 antibodyand a ribosome-inactivating protein from Saponaria officinali!-, potent anti-tumor effects in vitro and in vivo. J. Nati. Cancer Inst., 75: 151-159, 1985.Spitler, L. E., del Rio. M., Khentigan, A., Wedel, N. I., Brophy, N. A.,Miller, L. L.. Harkonen. W. S., Rosendorf. L. L., Lee, H. M., Mischak, R.P., Kawahata, R. T.. Stoudemire, J. B., Fradkin. L. B., Bautista, E. E., andScannon. P. J. Therapy of patients with malignant melanoma using amonoclonal antimelanoma antibody-ricin A-chain immunotoxin. CancerRes., 47: 1717-1723. 1987.Laurent, G.. Kuhlein, E., Casellas, P.. Canat. X., Carayon. P.. Poncelet, P.,Correli, S., Rigai, F.. and Jansen. F. K. Determination of sensitivity of freshleukemia cells to immunotoxins. Cancer Res., 46: 2289-2294, 1986.Preijers, F. W. M. B., Tax, W. J. M., De Witte. T.. Janssen. A., Heijden, H.V. D., Vidal, H.. Wessels, J. M. C., and Capei. P. J. A. Relationship betweeninternalization and cytotoxicity of ricin A chain immunotoxins. Br. J. Hae-matol.. 70: 289-294." 1988.
Scoti, C. F., Goldmacher, V. S., Lambert, J. M., Chari, R. V. J., Bolender,S., Gauthier, M. N., and Blätter.W. A. The antileukemic efficacy of animmunotoxin composed of a monoclonal anti-Thy 1antibody disulfide-linkedto the ribosome-inactivating protein gelonin. Cancer Immunol. Immunother.,25:31-40. 1987.Fitzgerald. D. J.. Björn,M. J., Ferris, R. J., Winkelhake, J. L., Frankel, A.E., Hamilton, T. C., Ozols, R. F.. Willingham. M. C., and Pastan, I.Antitumor activity of an immunotoxin in a nude mouse model of humanovarian cancer. Cancer Res., 47: 1407-1410, 1987.Hará,H., and Seon, B. K. Complete suppression of in vivogrowth of humanleukemia cells by specific immunotoxins: nude mouse models. Proc. Nail.Acad. Sci. USA, 84: 3390-3394. 1987.Griffin, T. W.. Richardson, C.. Houston, L. L.. LePage. D.. Bogden. A., andRaso, V. Antitumor activity of intraperitoneal immunotoxins in a nude mousemodel of human malignant mesothelioma. Cancer Res., 47: 4266-4270,1987.Björn,M. J., and Groetsema, G. Immunotoxins to the murine transferrinreceptor: ¡ntracavitarytherapy of mice bearing syngeneic peritoneal tumors.Cancer Res., 47: 6639-6645, 1987.Scott, C. F., Goldmacher. V. S., Lambert, J. M.. Jackson, J. V., and MeIntyre, G. D. An immunotoxin composed of a monoclonal anti-transferrinreceptor antibody linked by a disulfide bond to the ribosome-inactivatingprotein gelonin: potent in vitro and in vivo effects against human tumors. J.Nail. Cancer Inst.. 79: 1163-1172. 1987.Blakey. D. C., Skilleter. D. N.. Price, R. J.. Watson, G. J., Hart, L. L,Newell, D. R., and Thorpe, P. E. Comparison of the pharmacokinetics andhepatotoxic effects of saporin and ricin A-chain immunotoxins on murineliver parenchyma! cells. Cancer Res., 48: 7072-7078, 1988.Worrel, N. R., Cumber, A. J., Parnell, G. D., Mirza, A., Forrester, J. A., andRoss. W. C. J. Effect of linkage variation on pharmacokinetics of ricin A-chain-antibody conjugate in normal rats. Anti-Cancer Drug Design, /: 179-188. 1986.Thorpe, P. E.. Wallace. P. M.. Knowles, P. P.. Reif, M. G., Brown, A. N.F., Watson, G. J., Knyba, R. E., Wawrzynczak, E. J., and Blakey, D. C. Newcoupling agents for the synthesis of immunotoxins containing a hindereddisulfide bond with improved stability in vivo. Cancer Res., 47: 5924-5931,1987.
2916
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

1990;50:2909-2916. Cancer Res Odile Rostaing-Capaillon and Pierre Casellas
in VivoAntibody-Ricin A Chain Immunotoxins Parameters Affecting Tumor-specific Delivery of Anti-CD5
Updated version
http://cancerres.aacrjournals.org/content/50/10/2909
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/50/10/2909To request permission to re-use all or part of this article, use this link
Research. on November 11, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

![TumorBehaviorinTransitionalCellCarcinomaoftheBladderinRela ...cancerres.aacrjournals.org/content/47/24_Part_1/6800.full.pdf · [CANCERRESEARCH47,6800-6805,December!5,1987] TumorBehaviorinTransitionalCellCarcinomaoftheBladderinRelationto](https://img.pdfslide.us/doc/110x75/5c085d3a09d3f23a458c00a0/tumorbehaviorintransitionalcellcarcinomaofthebladderinrela-cancerresearch476800-6805december51987.jpg)




![TransientandStableComplementationofUltravioletRepairinXero ...cancerres.aacrjournals.org/content/47/11/2967.full.pdf · [CANCERRESEARCH47,2967-2971.June1,1987] TransientandStableComplementationofUltravioletRepairinXeroderma](https://img.pdfslide.us/doc/110x75/5a77350f7f8b9a4b538dd0bd/transientandstablecomplementationofultravioletrepairinxero-a-cancerresearch472967-2971june11987.jpg)




![ResistancetoAnthrapyrazolesandAnthracyclinesinMultidrug ...cancerres.aacrjournals.org/content/46/9/4352.full.pdf[CANCERRESEARCH46,4352-4356,September1986] ResistancetoAnthrapyrazolesandAnthracyclinesinMultidrug-resistantP388](https://img.pdfslide.us/doc/110x75/5aec16b57f8b9a90318dd526/resistancetoanthrapyrazolesandanthracyclinesinmultidrug-cancerresearch464352-4356september1986.jpg)



![HeterotransplantationofHumanLymphoidNeoplasmsUsingaNudeMou ...cancerres.aacrjournals.org/content/50/10/3078.full.pdf · (CANCERRESEARCH50,3078-3086.May15,1990] HeterotransplantationofHumanLymphoidNeoplasmsUsingaNudeMouse](https://img.pdfslide.us/doc/110x75/5e83f48eaae3144d7c04ca6b/heterotransplantationofhumanlymphoidneoplasmsusinganudemou-cancerresearch503078-3086may151990.jpg)







![VirologieStudiesinHumanLeukemiaandLymphoma:The …cancerres.aacrjournals.org/content/28/7/1311.full.pdf · [CANCERRESEARCH28,1311-1318,July1968] VirologieStudiesinHumanLeukemiaandLymphoma:The](https://img.pdfslide.us/doc/110x75/5b98f37b09d3f219118d06a7/virologiestudiesinhumanleukemiaandlymphomathe-cancerresearch281311-1318july1968.jpg)


![InVitroHematopoiesisfollowingInductionChemotherapyforAcute ...cancerres.aacrjournals.org/content/45/11_Part_2/5921.full.pdf[CANCERRESEARCH45,5921-5925,November1985] InVitroHematopoiesisfollowingInductionChemotherapyforAcuteLeukemia1](https://img.pdfslide.us/doc/110x75/5b0a316b7f8b9a45518be441/invitrohematopoiesisfollowinginductionchemotherapyforacute-cancerresearch455921-5925november1985.jpg)
![ClinicalValueofSerumGlycoproteinGalactosyltransferaseLevel ...cancerres.aacrjournals.org/content/43/9/4491.full.pdf · [CANCERRESEARCH43,4491-4496,September1983] ClinicalValueofSerumGlycoproteinGalactosyltransferaseLevelsin](https://img.pdfslide.us/doc/110x75/5ac139b17f8b9ac6688d1490/clinicalvalueofserumglycoproteingalactosyltransferaselevel-cancerresearch434491-4496september1983.jpg)

