Embed Size (px)
Citation preview

[CANCER RESEARCH 44, 4403-4408, October 1984]
Lack of Correlation between Natural Killer Activity and Tumor Growth
Control in Nude Mice with Different Immune Defects
Oystein Fodstad,1 Carl T. Hansen, Grace B. Cannon, Charles N. Statham, Gary R. Lichtenstein, and
Michael R. Boyd
Laboratory of Experimental Therapeutics and Metabolism, Developmental Therapeutics Program, Division of Cancer Treatment [0. F., C. N. S., G. R. L, M. R. B.] andVeterinary Resources Branch, Division of Research Services [C. T. H.J, NIH, Bethesda, Maryland 20205, and Department of Immunology, Litton Bionetics, Inc.[G. B. C.], Kensington, Maryland 20814
ABSTRACT
To elucidate the in vivo role of natural killer (NK) cells, thegrowth of several murine and human tumors was studied in fourvariants of athymic, nude mice with different levels of NK activity.
Beige-nude mice, homozygous for both the beige and the
nude genes, had very low levels of NK activity, and their responseto the B-cell mitogen, bacterial lipopolysaccharide, was lowerthan that of high-NK, adult NIH nude mice. Young and adult NIH
nudes had different NK levels and showed different response inassays for K-cell, T-cell, and B-cell activity. The B-cell-defectiveNIH-II mice had slightly lower NK levels than adult NIH animals,but much lower response in the antibody-dependent cell-me
diated cytotoxicity assay.No correlation was found between host NK activity and the
s.c. growth of various human (LOX, CEM, K562) and murine(YAC-1) tumor cells. Low NK activity was not associated withincreased lung colony formation in a metastasis model using i.v.-
injected human (LOX) and murine (B16F10) melanoma cells. Norelationship was found between host NK activity and the rate ofelimination of i.v.-injected 5-iodo-2'-deoxyuridine-labeled LOX,
B16F10, and YAC-1 cells from lungs, liver, or spleen.
The results fail to support the view that NK cells exert significant direct effects on tumor cells in vivo.
INTRODUCTION
It is widely believed that NK2 cells have an important role in
immune surveillance against tumors (12,19, 36). The data supporting this theory have been obtained in mice with differentlevels of NK activity (23, 26), or in models where the animalswere treated with drugs that are supposed to selectively affectNK function (11, 14, 15, 24). However, differences in hostimmune response other than in NK activity may have influencedthe results. Therefore, definitive studies, proving the in vivo roleof NK cells, are lacking (28), and there is a need for better models(33, 34).
Recently, we have reported on viable beige-nude mice with
very low NK activity, compared to that of adult, regular NIH nudemice.3 Apart from the NK difference, the 2 types of nude micediffered immunologically only in a lower response to the B-cell
1To whom requests for reprints should be addressed, at Norsk Hydros Institute
for Cancer Research, The Norwegian Cancer Society and The Norwegian RadiumHospital, Montebello 0310 Oslo 3, Norway.
2The abbreviations used are: NK, natural killer; LPS, lipopolysaccharide; ADCC,antibody-dependent cell-mediated cytotoxicity; PFC, plaque-forming cell; IdUrd, 5-iodo-2 ' -deoxyuridine.
30. Fodstad, C. T. Hansen, G. B. Cannon, and M. R. Boyd. Immune characteristics of the beige-nude mouse, a model for studying immune surveillance. Scand.J. Immunol., submitted for publication, 1984.
Received February 17,1984; accepted June 1,1984.
mitogen, LPS, in the beige than in NIH animals. The beige-nudes,therefore, seemed to offer new possibilities for investigating thein vivo importance of NK cells.
In the present study, we have compared cellular immuneresponse and tumor growth in beige-nude mice and in 3 other
variants of athymic, nude mice with different immune characteristics. Surprisingly, no correlation between NK activity and tumorgrowth control was found.
MATERIALS AND METHODS
Mice
Four groups of immune-deficient, athymic mice were studied: (a) adult
(5 to 7 weeks old) N:NIH(S) nude mice, which have high NK activity (18);(b) immunologically immature 2- to 3-week-old N:NIH(S) nudes (14, 15,18); (c) low-NK beige-nude mice, developed on C57BL/6N-beige andN:NIH(S)-nude background3; and (d) B-cell-defective N:NIH(S)-ll nude
mice(1).All animals were bred and kept in specific pathogen-free facilities until
they had reached the desired age and body weight (adult mice, 5 to 7weeks old), when they were transferred to conventional facilities for thetumor experiments. There, the mice were kept in disposable plasticcages with autoclaved filter tops, placed in laminar air-flow benches. The
food and bedding were autoclaved before use, and the animals weregiven tap water ad libitum in sterilized bottles.
Breeding of Beige-Nude Mice
Five mating steps were needed to obtain beige-nude mice, (a) FemaleC57BL/6N-bg bg/bg +/+ mice were mated to male, nude N:NIH(S) +/+
nú/númice, resulting in bg/+ nu/+ offspring, agouti in color, (b) The fag/+ nu/+ mice were crossed back to C57BL/6N-bg, yielding 50% homo
zygous bg/bg beige mice. Half of these were nu¡+, and half were +/+,but these could not be distinguished by eye. (c) When such beige micewere mated to either nu/+ females or nu/nu males, the bg/bg nu/+parent could be identified, since they had at least one nude animal intheir litter, (d) Female and male bg/bg nu/+ mice were mated to eachother, producing one-fourth beige-nude bg/bg nu/nu, two-fourths bg/bgnu/+, and one-fourth bg/bg +/+ animals, (e) To identify the bg/bg nu/+mice, beige-nude males were mated to bg/bg nonnude females. Those
females that did not produce any nude offspring were bg/bg +/+ andtherefore discarded, whereas the remaining bg/bg nu/+ females wereused to establish the breeding colony, together with the bg/bg nu/numales. Since the progeny were either bg/bg nu/+ or bg/bg nu/nu, inequal proportions, no further testing was necessary.
Tumors
Two murine (YAC-1 and B16F10) and 3 human (K562, CEM, andLOX) tumors were used. The YAC-1 is a lymphoma, originally induced
in A/Sn mice, and the cell line is commonly used as a sensitive target fortesting NK cell activity in vitro (4). The B16F10 melanoma cell line,isolated by Fidler (7), was obtained from E. M. Jensen, Mason Research
OCTOBER 1984 4403
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

0. Fodstad et al.
Institute, Worcester, MA. The human, undifferentiated T-cell line, CEM,
was a gift from Kevin Scanlon, Mt. Sinai Medical Center, New York, NY.The K562 line is a chronic myelogenic cell line, originally established byLozzio and Lozzio (29). LOX is a human malignant melanoma, passagedas a xenograft tumor line in nude mice, and also established as a cell
line in vitro.The tumor cells were kept in continuous culture in RPM11640 medium
(Flow Laboratories, Rockville, MD) with 5% fetal calf serum for at least3 to 4 weeks, to ensure a high sensitivity to NK cells (2, 13), beforebeing used in the experiments.
The tumors were shown to be free of Mycoplasma and the followingpathogenic viruses: (a) K-virus; (b) reovirus type 3; (c) pneumonia virusof mice; (d) Theiler's encephalitis virus; (e) Sendai virus; (f) minute virusof mice; (g) mouse adenovirus; (h) mouse hepatitis virus; (/') lymphocyticchoriomeningitis virus; and (/') ectromelia virus (Microbiological Associ
ates, Bethesda, MD).
Cellular Immune Response In Vitro
NK and ADCC Cytotoxicity Assays. In the NK cell assay, spleen cellsfrom the different groups of mice were dispersed (0.1 ml) at differentconcentrations into the round-bottomed wells of a microtiter plate. 5'Cr-
labeled YAC-1 tumor cells (0.1 ml) were added into the appropriate wellsand incubated at 37°for 4 hr. The ADCC assay, regarded to reflect the
activity of killer cells (34), was performed essentially as described (16),but with Chang cells as targets. The rabbit anti-Chang antiserum was a
gift from Julie Djeu, and was used in an optimal concentration of 1/10.The antibody-coated 51Cr-labeled Chang cells were mixed with the spleeneffector cells and incubated at 37° for 18 hr. For both assays, thesupernatants in each well containing released 51Cr were collected and
counted in a -»-counter.Spontaneous release was always less than 10%
of total release obtained with 2% sodium dodecyl sulfate. NK activityand ADCC are expressed as:
% of cytotoxicity = Test cpm- spontaneous cpm/cpm initially incorporated
The tests were performed with 3 different effector/target ratios: (a) 50/1; (b) 100/1; and (c) 200/1. Only the results with the ratio 200/1 aregiven.
Lymphocyte Proliferation Assays. Viable spleen cells were culturedin 4 replicates in the round-bottomed wells of a microtiter plate (2.5 x105 cells/well) in RPMI 1640 medium, supplemented with 25 mM 4-{2-
hydroxyethyl)-1-piperazineethanesulfonicacid buffer, 1% glutamine, and10% heat-inactivated fetal bovine serum. Optimal concentrations of
purified phytohemagglutinin (0.25 jig/well), concanavalin A (0.5 ^g/well),or bacterial LPS (20 MO/well) were added, and the cultures were incubated for 3 days at 37°.Six hr prior to the culture termination, 1.0 #tCiof [3H]thymidine (0.05 ml) was added to each culture. The cells were
harvested with a multiple sample harvester onto the strips of fiberglasspaper, and [3H]thymidine incorporation into DNA was determined by
liquid scintillation spectroscopy. The background values were 700 ±54cpm (S.D.), and the data are expressed as net cpm (cpm in stimulatedcultures minus cpm in cultures with no mitogen).
RFC Assay. Spleens were removed from mice sacrificed 4 days afteri.v. injection of 2 x 109 (0.1 ml of 10% suspension) sheep RBC. Single-cell suspensions were tested in Cunningham's modification of the Jeme
plaque assay; a mixture of either 1 x 107 or 1 x 10" spleen cells (0.2
ml), 0.4 ml of sheep RBC, 0.1 ml of guinea pig complement, and 0.3 mlof medium was pipetted into Cunningham chambers. The chamberswere sealed with petroleum jelly and were incubated at 37°for 1 hr, and
plaques were counted using an inverted microscope. The data wereexpressed as PFCs per spleen.
Tumor Studies in Vivo
Tumor Growth s.c. Tumor cells were harvested from exponentiallygrowing cultures with mild trypsinization, washed twice in ice-cold me
dium without fetal calf serum, diluted in appropriate concentrations, andinjected s.c. into the flanks of nude mice in 0.5-ml volumes. The mice
were checked daily for tumor take, and when the tumors were growing,2 perpendicular diameters were measured 2 to 3 times weekly, andmean tumor diameter for each group was calculated.
Assay for Growth of Experimental Pulmonary Métastases. TheB16F10 cells were harvested from in vitro cultures as described above.Single cell suspensions of the LOX tumor were in this case obtainedfrom s.c. xenografts growing in nude mice, removed under sterile conditions, cut into pieces of 4.5 mm, and minced using a Stomacher 80Lab-Blender, Seward Surgical, London, England. The cell suspension
was passed through sterile gauze, and viable cells, determined by Trypanblue exclusion, were diluted in medium to appropriate concentrationsand injected i.v. in 0.2-ml volumes. The mice were checked daily, and
the survival time of mice that died was recorded. In some experiments,one-half of the animals within each group were killed on Day 25 (LOX)
or Day 18 (B16F10) after tumor cell injection. The lungs were removedand fixed in Bouin's solution, and the number of tumor colonies was
determined.Assay of Radiolabeled Tumor Cell Retention in Different Tissues.
The tumor cells were labeled as described by Riccardi et al. (32). Shortly,5 x 10" tumor cells in 50 ml of RPMI medium, supplemented with 20%fetal calf serum, were incubated for 14 hr at 37°in a 5% C02 incubatorin the presence of 5 to 10 mg of 5-fluoro-2'-deoxyuridine and 20 ¿iCiof125ldUrd (Radiochemical Centre, Amersham, England). The cells were
extensively washed with medium, diluted to appropriate concentrationsof viable cells, and injected i.v. in 0.2-ml volumes, containing 1 x 106LOX cells (-75,000 cpm), 5 x 10* B16F10 cells (-60,000 cpm), and 5x 104 YAC-1 cells (-50,000 cpm). At intervals, ranging from 10 min to
48 hr after i.v. tumor cell inoculation, animals (4 to 5/group) were killed,and the radioactivity in the lungs, liver, and spleen was measured in a ->-
counter. Each experiment was performed twice. The results were expressed as retained radioactivity in percentage of radioactivity injected.The radioactivity injected into each mouse was determined by countingquadruplicate samples of 0.2-ml tumor cell suspensions.
RESULTS
Immune Characteristics of Different Nude Mouse Genotypes
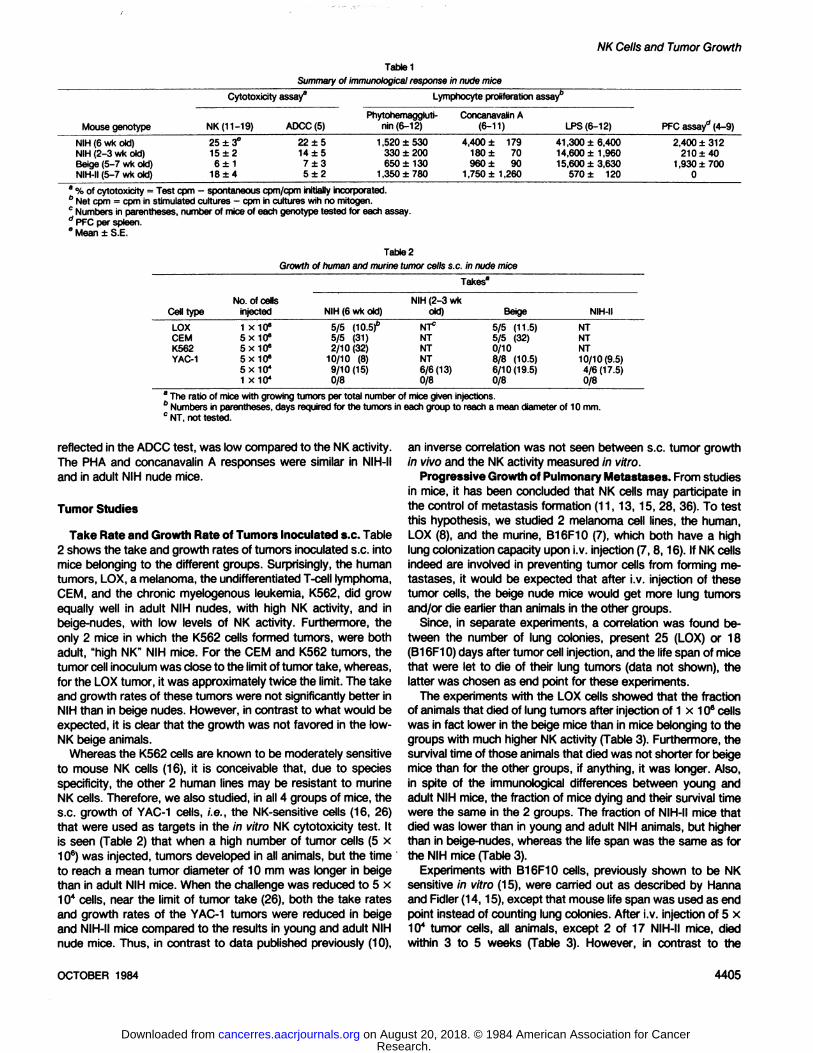
Adult NIH nude mice are known to have high NK activity3 (19),
whereas young (2 to 3 weeks old) nude mice have low NKactivity (11, 13, 14). However, the results in Table 1 show thatthe young NIH nudes gave lower values than adult NIH nudes,not only in the NK test, but also in the assays for K-cell (ADCC),T-cell (phytohemagglutinin and concanavalin A), and B-cell (LPS)activity, as well as in T-dependent B-cell response (PFC) assay.
For the beige-nude mice, the NK activity was much lower than
for adult NIH nudes (Table 1). The difference is statisticallysignificant in the Wilcoxon test (p < 0.01). The NK activity inbeige-nudes was also appreciably lower than in the "low NK"
(14,15,18), 2- to 3-week-old NIH nude mice (Table 1). Compat
ible with the suggestion that a relationship may exist betweenthe effector cells in the NK and ADCC assays (33,40), the valuesin these tests were closely similar for beige as well as for NIHnudes. The response to the T-mitogens was lower, although not
significantly, in beige than in adult NIH mice, whereas the LPSresponse was similar in beige and in 2- to 3-week-old NIH nude
mice, and the PFC values were similar in beige and in adult NIHanimals.
The NIH-II mice have a defective B-cell response, due to the
acquisition of the XID gene (1), as reflected here in the LPS andPFC results (Table 1). In contrast to the situation in the othervariants of nude mice, the activity in NIH-II mice of K-cells, as
4404 CANCER RESEARCH VOL. 44
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
NK Cells and Tumor Growth
Table 1Summaryof immunologicalresponse in nude mice
Cytotoxicityassay8Mouse
genotypeNIH
(6 wk old)NIH (2-3 wk old)Beige (5-7 wk old)NIH-II(5-7 wk old)NK
(11-19)25±3°
15±26±1
18 ±4ADCC
(5)22
±514 ±57±35±2Lymphocyte
proliferationassay"Phytohemaggluti-
nin(6-12)1,520
±530330 ±200650 ±130
1,350 ±780Concanavalin
A(6-11)4,400
± 179180 ± 70960 ± 90
1,750±1,260LPS(6-12)41
,300 ±6,40014,600 ±1,96015,600 ±3,630
570 ± 120RFC
assay"(4-9)2,400
±312210 ±40
1,930 ±7000
a % of cytotoxicity = Test com - spontaneous cpm/cpm initially incorporated.6 Net com = cpm in stimulated cultures - cpm in cultures wih no mitogen.0 Numbers in parentheses,number of mice of each genotype tested for each assay." PFC per spleen." Mean ±S.E.
Table2Growth ol humanand murine tumor cells s.c. in nude mice
Takes8Cell
typeLOX
OEMK562YAC-1No.
of cellsinjected1
x10»5x10«5x10«5x10«5x 10*1 X104NIH
(6 wkold)5/5(10.5)6
5/5 (31)2/10(32)
10/10 (8)9/10(15)0/8NIH
(2-3 wkold)NT*
NTNTNT6/6(13)0/8Beige5/5
(11.5)5/5 (32)0/108/8 (10.5)6/10(19.5)0/8NIH-IINT
NTNT
10/10(9.5)4/6(17.5)0/8
aThe ratio of mice with growing tumors per total number of mice given injections." Numbers in parentheses,days required for the tumors in each group to reach a meandiameter of 10 mm.c NT, not tested.
reflected in the ADCC test, was low compared to the NK activity.The PHA and concanavalin A responses were similar in NIH-II
and in adult NIH nude mice.
Tumor Studies
Take Rate and Growth Rate of Tumors Inoculated s.c. Table2 shows the take and growth rates of tumors inoculated s.c. intomice belonging to the different groups. Surprisingly, the humantumors, LOX, a melanoma, the undifferentiated T-cell lymphoma,
CEM, and the chronic myelogenous leukemia, K562, did growequally well in adult NIH nudes, with high NK activity, and inbeige-nudes, with low levels of NK activity. Furthermore, the
only 2 mice in which the K562 cells formed tumors, were bothadult, "high NK" NIH mice. For the CEM and K562 tumors, the
tumor cell inoculum was close to the limit of tumor take, whereas,for the LOX tumor, it was approximately twice the limit. The takeand growth rates of these tumors were not significantly better inNIH than in beige nudes. However, in contrast to what would beexpected, it is clear that the growth was not favored in the low-
NK beige animals.Whereas the K562 cells are known to be moderately sensitive
to mouse NK cells (16), it is conceivable that, due to speciesspecificity, the other 2 human lines may be resistant to murineNK cells. Therefore, we also studied, in all 4 groups of mice, thes.c. growth of YAC-1 cells, i.e., the NK-sensitive cells (16, 26)
that were used as targets in the in vitro NK cytotoxicity test. Itis seen (Table 2) that when a high number of tumor cells (5 x106) was injected, tumors developed in all animals, but the time
to reach a mean tumor diameter of 10 mm was longer in beigethan in adult NIH mice. When the challenge was reduced to 5 x104 cells, near the limit of tumor take (26), both the take rates
and growth rates of the YAC-1 tumors were reduced in beigeand NIH-II mice compared to the results in young and adult NIH
nude mice. Thus, in contrast to data published previously (10),
an inverse correlation was not seen between s.c. tumor growthin vivo and the NK activity measured in vitro.
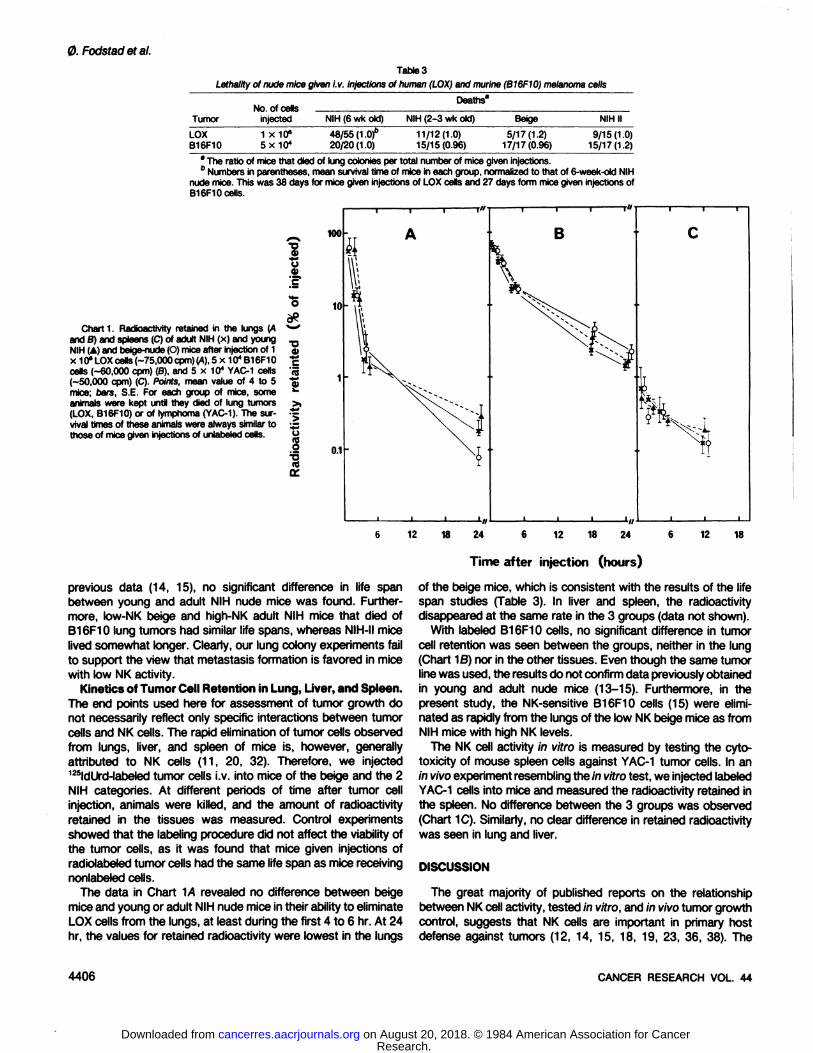
Progressive Growth of Pulmonary Métastases.From studiesin mice, it has been concluded that NK cells may participate inthe control of metastasis formation (11, 13, 15, 28, 36). To testthis hypothesis, we studied 2 melanoma cell lines, the human,LOX (8), and the murine, B16F10 (7), which both have a highlung colonization capacity upon i.v. injection (7,8,16). If NK cellsindeed are involved in preventing tumor cells from forming métastases, it would be expected that after i.v. injection of thesetumor cells, the beige nude mice would get more lung tumorsand/or die earlier than animals in the other groups.
Since, in separate experiments, a correlation was found between the number of lung colonies, present 25 (LOX) or 18(B16F10) days after tumor cell injection, and the life span of micethat were let to die of their lung tumors (data not shown), thelatter was chosen as end point for these experiments.
The experiments with the LOX cells showed that the fractionof animals that died of lung tumors after injection of 1 x 106 cells
was in fact lower in the beige mice than in mice belonging to thegroups with much higher NK activity (Table 3). Furthermore, thesurvival time of those animals that died was not shorter for beigemice than for the other groups, if anything, it was longer. Also,in spite of the immunological differences between young andadult NIH mice, the fraction of mice dying and their survival timewere the same in the 2 groups. The fraction of NIH-II mice thatdied was lower than in young and adult NIH animals, but higherthan in beige-nudes, whereas the life span was the same as for
the NIH mice (Table 3).Experiments with B16F10 cells, previously shown to be NK
sensitive in vitro (15), were carried out as described by Hannaand Fidler (14,15), except that mouse life span was used as endpoint instead of counting lung colonies. After i.v. injection of 5 x104 tumor cells, all animals, except 2 of 17 NIH-II mice, died
within 3 to 5 weeks (Table 3). However, in contrast to the
OCTOBER 1984 4405
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
0. Fodstad et al.
Table 3
Lethality of nude mice given i.v. injections of human (LOX) and murine (B16F10) melanoma cells
TumorLOX
B16F10No.
of cellsinjected1
X10"5X104Deaths'NIH
(6 wkold)48/55(1
.Of20/20(1.0)NIH
(2-3 wkold)11/12(1.0)
15/15(0.96)Beige5/17(1.2)17/17(0.96)NIH
II9/15(1.0)
15/17(1.2)* The ratio of mice that died of lung colonies per total number of mice given injections." Numbers in parentheses, mean survival time of mice in each group, normalized to that of 6-week-oid NIH
nude mice. This was 38 days for mice given injections of LOX cells and 27 days form mice given injections ofB16F10 cells.
Chart 1. Radioactivity retained in the lungs (Aand B) and spleens (C) of adult NIH (x) and youngNIH (A) and beige-nude (O) mice after injection of 1x 10" LOX cells (-75,000 cpm) (A), 5 x 104 B16F10cells (-60,000 cpm) (B), and 5 x 104 YAC-1 cells
(-50,000 cpm) (C). Points, mean value of 4 to 5mice; bare, S.E. For each group of mice, someanimals were kept until they died of lung tumors(LOX, B16F10) or of lymphoma (YAC-1). The sur
vival times of these animals were always similar tothose of mice given injections of unlabeled cells.
1
•o
l'SS>•*rf
j>"o
I
KM
10
0.1
12 18 24 12 18 24 12 18
Time after injection (hours)
previous data (14, 15), no significant difference in life spanbetween young and adult NIH nude mice was found. Furthermore, low-NK beige and high-NK adult NIH mice that died ofB16F10 lung tumors had similar life spans, whereas NIH-II mice
lived somewhat longer. Clearly, our lung colony experiments failto support the view that metastasis formation is favored in micewith low NK activity.
Kinetics of Tumor Cell Retention in Lung, Liver, and Spleen.The end points used here for assessment of tumor growth donot necessarily reflect only specific interactions between tumorcells and NK cells. The rapid elimination of tumor cells observedfrom lungs, liver, and spleen of mice is, however, generallyattributed to NK cells (11, 20, 32). Therefore, we injected125ldUrd-labeled tumor cells i.v. into mice of the beige and the 2
NIH categories. At different periods of time after tumor cellinjection, animals were killed, and the amount of radioactivityretained in the tissues was measured. Control experimentsshowed that the labeling procedure did not affect the viability ofthe tumor cells, as it was found that mice given injections ofradiolabeled tumor cells had the same life span as mice receivingnonlabeled cells.
The data in Chart 1/4 revealed no difference between beigemice and young or adult NIH nude mice in their ability to eliminateLOX cells from the lungs, at least during the first 4 to 6 hr. At 24hr, the values for retained radioactivity were lowest in the lungs
of the beige mice, which is consistent with the results of the lifespan studies (Table 3). In liver and spleen, the radioactivitydisappeared at the same rate in the 3 groups (data not shown).
With labeled B16F10 cells, no significant difference in tumorcell retention was seen between the groups, neither in the lung(Chart ^B) nor in the other tissues. Even though the same tumorline was used, the results do not confirm data previously obtainedin young and adult nude mice (13-15). Furthermore, in thepresent study, the NK-sensitive B16F10 cells (15) were elimi
nated as rapidly from the lungs of the low NK beige mice as fromNIH mice with high NK levels.
The NK cell activity in vitro is measured by testing the cyto-toxicity of mouse spleen cells against YAC-1 tumor cells. In an
in vivo experiment resembling the in vitro test, we injected labeledYAC-1 cells into mice and measured the radioactivity retained in
the spleen. No difference between the 3 groups was observed(Chart 1C). Similarly, no clear difference in retained radioactivitywas seen in lung and liver.
DISCUSSION
The great majority of published reports on the relationshipbetween NK cell activity, tested in vitro, and in vivo tumor growthcontrol, suggests that NK cells are important in primary hostdefense against tumors (12, 14, 15, 18, 19, 23, 36, 38). The
4406 CANCER RESEARCH VOL. 44
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

present results do not support this notion. Thus, in 4 variants ofnude mice, low NK activity was not associated with increasedgrowth of s.c.-inoculated human (LOX, GEM, K562) and murine(YAC-1) tumor cells, nor was low NK activity associated withincreased lung colony formation in a metastasis model using i.v.-
injected human (LOX) and murine (B16F10) melanoma cells.Moreover, there was no relationship between host NK activityand the rate of elimination of 125ldUrd-labeled LOX, B16F10, and
YAC-1 cells from lungs, liver, or spleen.
In several of the previous reports (10, 15, 22, 23, 36), theselectivity of the NK impairment was not carefully investigated,and immune differences other than in NK activity may haveinfluenced the results. This could be the case when increasedtumor growth in young, compared to in adult, NIH nude micewas ascribed to differences in NK activity, and considered asevidence for the in vivo significance of NK cells (11, 13, 14).Thus, in light of the other immunological differences demonstrated here (Table 1), this conclusion may not be valid.
Beige-nude mice with low NK activity have previously been
suggested to be a valuable tool for studies of tumor growth andmetastasis, and for evaluating the In vivo role of NK cells inimmune surveillance against tumors (5, 22). However, until nowsuch studies have not been reported, possibly due to problemswith the viability of the mice (22). Our beige-nudes were healthy
and had a life span similar to that of regular nude mice. Througha comparison of the immune status of both parental strains,3
and as shown in Table 1, it was established that the beige-nude
mice had immune characteristics that were promising for suchtumor experiments.
To our surprise, the growth of the human and mouse tumorstested was not increased in these low-NK mice compared to innude mice with high-NK activity. Beige mice are not completely
devoid of NK cells, but the differences in NK function betweenthe different types of nude mice here tested were highly significant, and the present results strongly question the view that NKcells have important cytotoxic effects In vivo. It is well-known
that tumor inoculations, as well as interferon and the copolymerof polyinosinic and polycytidylic acids, can induce a transientincrease in NK activity, and this increase could theoretically bedifferent in different nude strains. It seems reasonable to assume,however, that the magnitude of this response will not be largerin beige-nude mice with deficient NK function than in mice of
genotypes with normal or high initial NK values.The different nude genotypes examined did not differ in NK
function only (Table 1). However, the other immune defects foundwould be expected, if anything, to favor tumor growth in thebeige-nude animals. Macrophage function was not tested, and
since macrophages may be involved in natural resistance totumors (6, 35), possible differences in macrophage activity between the mouse genotypes could perhaps influence tumorgrowth and help to explain the present data. However, it isknown that the macrophages in regular beige mice show abnormal granules, defective chemotaxis, and delayed antitumor activity in vitro (31), and it is likely that this is the case also in beigenude mice. If so, this would be thought to favor, not suppress,tumor take and growth in the beige animals.
It is well-known that host immune response may easily be
affected by viral infections in the animals. Therefore, it should beemphasized that no virus infection was detected during thepresent studies, a fact probably reflected by the overall relatively
NK Cells and Tumor Growth
low NK values (14,15,18) for the different groups of mice (Table1).
The B-cell-deficient NIH-II nude mice had much lower values
in the ADCC assay than in the NK test. This finding is notconsistent with the view that the NK and K-effector cells are thesame (33, 34, 41). NIH-II mice might, therefore, be useful instudying the lineage of the K-cells, and the results raise thequestion of whether K-cell function may depend partly on anormal B-cell activity.
No simple answer is available to the question of why thepresent data are inconsistent with previous results (11, 12, 15,23,24,36) on the in vivo role of NK cells. If NK cells indeed havean important role in vivo, one would have expected to finddifferences between the mice in take and growth rates of s.c.-inoculated tumors (10). Furthermore, the claim that NK cells areimportant in preventing metastasis formation (11,13,15,28,36)was not supported by the results of our lung colony experimentswith the human LOX and the murine B16F10 melanomas. In thepresent study, life span was used as end point, whereas Hannaand Fidler (15), in similar experiments with the B16F10 tumor,counted the number of lung colonies present 3 to 5 weeks aftertumor cell injection. The use of different end points cannot explainthe discrepancy between the results, as, in our experiments, acorrelation was found between the number of lung colonies andthe life span of the animals. It may be of importance, however,that whereas Hanna and Fidler (15) reported that adult, regularnude mice had developed very few lung colonies 3 to 5 weeksafter injection of 5 x 10* B16F10 cells (15), a high number of
pulmonary métastaseswas found at Day 18 in our adult NIHnude mice, which started to die of lung tumors approximately 3weeks after injection of the cells. The beige-nude mice used in
our experiments do in part have the C57BL genetic background,and since the B16F10 tumor is syngeneic for C57BL (7), theB16F10 cells could possibly have a growth advantage in thebeige-nude mice. Nevertheless, no difference in lung colony
growth between beige and NIH nude mice was seen.In contrast to the results of previous studies on B16F10 cells
(15), a lack of relationship was found here between host NKactivity and the rate of elimination of 125ldUrd-labeled tumor cells
from lungs, liver or spleen. However, since consistent resultswere obtained with all 3 tumors studied, our data stronglyindicate that NK cells do not have an important role in early hostdefense against tumor cells.
In agreement with data recently obtained in conventional mice(35), the overall conclusion of the present study is that, althoughNK cells clearly are cytotoxic in vitro, they do not seem to exertany significant direct effect on tumor cells in vivo. The mechanisms involved in host defense against tumors are apparentlymore complex than previously appreciated. Thus, the demonstrated differences in tumor growth control between the 4 typesof mice cannot easily be explained on the basis of the immunological data. It should be noted, however, that in all strains ofnude mice tested, minor responses to the T-cell mitogens wereseen (Table 1). It is possible that nude mice have T-like cells (27,30) that may become functional after induction (9, 21, 37). Thedistribution of such cells may differ between the nude genotypes.Also, it is conceivable that interactions between different immunecells (20,39,41 ), possibly including NK cells, or factors producedby them (9,17, 25), are important in the regulation of the immuneresponse (3). Therefore, it seems reasonable to suggest that,
OCTOBER 1984 4407
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

0. Fodstad et al.
rather than attempting to explain tumor growth data in vivo by¡mmunologicaldifferences restricted to one single parameter, theinteraction between different immune cells should be furtherinvestigated. In such studies, nude mice with various combinations of immune defects may prove valuable.
ACKNOWLEDGMENTS
We thank ChartesJ. Sanders, Mary McMenamin,and Larry Jerome for excellenttechnicalassistance,and Alexander Pihl for valuablediscussion.
REFERENCES
1. Azar, H. A., Hansen,C. T., and Costa, J. N:NIH(S)ll-nu/numice with combinedimmunodeficiency.A new model for human tumor heterotransplantation. J.Nati. Cancer Inst . 65: 421-430,1980.
2. Becker, S., Kiessling, Ft., Lee, N., and Klein, G. Modulation of sensitivity tonatural killer cell lysis after in vitro explantation of a mouse lymphoma.J. Nati.Cancer Inst., 61: 1495-1498,1978.
3. Beverly, P. Understandingnatural killer cells. Nature (Lond.),297: 288,1981.4. Cikes, M., Friberg, S., Jr., and Klein, G. Progressive bss of H-2 antigen(s)
determinedby Moloney leukemiavirus in cultured murine lymphomas.J. Nati.Cancer Inst., 50: 347-362.1973.
5. Clark, E. A., Schultz, L. D., and Pollack, S. B. Mutations in mice that influencenatural killer (NK)cell activity. Immunogenetics, 12: 601-613,1981.
6. Evans. R. Macrophagesand neoplasms: new insights and their implication intumor immunobiology Cancer Metastasis Rev., 1: 227-239,1982.
7. Fidler, I. J. General considerations for studies of experimental metastasis.Methods Cancer Res., 75: 399-439,1978.
8. Fodstad, 0., Aamdal, S., Tveit, K. M., and Pihl, A. Lung colony formation inadult nude mice upon intravenous injection of cells from a human melanomaxenograft. Proc. Am. Assoc. Cancer Res., 23: 223,1982.
9. Gillis, S., and Watson, J. lnterteukin-2 induction of hapten-specifìccytolytic Tcells in nude mice. J. Immuro!., J26:1245-1248,1981.
10. Gorelik. E., and Herberman,R. B. Radioisotopeassay for evaluationof in vivonatural cell-mediatedresistanceof mice to local transplantation of tumor cells.Int. J. Cancer, 27: 709-720, 1981.
11. Gorelik, E., Wiltrout, R. H., Okumura, K., Habu,S., and Herberman,R. B. Roteof NK cells in the control of metastatic spread and growth of tumor cells inmice. Int. J. Cancer,30:107-112,1982.
12. Haller,0., Hansson,M., Kiessling,R., and Wigzell.H. Roleof non-conventionalnatural killer cells in resistance against syngeneic tumour cells in vivo. Nature(Lond.),270.609-611,1977.
13. Hanna. N. Expression of metastatic potential of tumor cells in young nudemice is correlated with low levels of natural killer cell-mediated cytotoxicity.Int. J. Cancer, 26: 675-680, 1980.
14. Hanna, N. Rote of natural killer cells in control of cancer metastasis. CancerMetastasis Rev.. 1: 45-64,1982.
15. Hanna,N., and Fidler. I. J. Expression of metastatic potential of allogene«andxenogeneicneoplasms in young nude mice. Cancer Res., 41: 438-444,1981.
16. Hansson, M., Karre, K., Bakacs, T., Kiessling, R., and Klein. G. Intra- andinterspectesreactivityof humanand mousenatural kilter(NK)cells.J. Immunol.,72?. 6-12, 1978.
17. Henney, C. S., Kuribayashi,K., Kern, D. E., and Gillis, S. lnterteukin-2naturalkiltercell activity. Nature (Lond.), 297:335-338,1981.
18. Herberman, R. B., and Holden, H. T. Natural cell-mediated immunity. AdvCancer Res., 27: 305-377,1978.
19. Herbennan, R. B., and HokJen,H. T. Natural killer cells as antitumor effectorcells. J. Nati. Cancer Inst.. 62: 441-445,1979.
20. Herberman,R. B., and Ortaldo, J. R. Natural kiltercells: their rote in defensesagainst disease. Science(Wash. DC). 214:24-30,1981.
21. Hünig,T., and Bevan, M. J. Specificity of cytotoxic T cells from athymic mice.J. Exp. Med., 752: 688-702,1980.
22. Karre, K., Klein, G. O., Kiessling, R., Klein, G., and Roder, J. C. In vitro NK-activity and in vivo resistance to leukemia: studies of beige, beige/nude andwild-type hosts on C57BL background. Int. J. Cancer,26: 789-797,1980.
23. Karre, K., Klein,G. 0., Kiessling,R., Klein, G., and Roder,J. C. Low natural invivo resistance to syngeneic leukaemiasin natural kilter-deficientmice. Nature(Lond.),284: 624-626, 1980.
24. Kasai, M., Yoneda.T., Habu, S., Maruyama, Y., Okumura, K., and Tokunaga.T. in vivo effect of anti-asialo GM1 antibody on natural kilter activity. Nature(Lond.),297:334-335,1981.
25. Kawase, I., Urdal, D. L., Newman, W., and Henney, C. S. The mechanismofaugmentation of natural kilter cell activity by syngeneic tumor cells: rote ofmacrophage-denvedfactor in NK boosting. Int. J. Cancer,37: 365-372,1983.
26. Kiessling, R., Petranyi, G., Klein, G., and Wigzell, H. Non-T-cell resistanceagainst a mouse Moloney lymphoma. Int. J. Cancer, 77: 275-281,1976.
27. Kindred, B. T cell functions in nude mice. Exp. Cell Biol., 50: 306, 1982.28. Koren, S. K., and Herberman,R. B. Natural killing—presentand future (sum
mary of workshop on natural killer cells). J. Nati. Cancer Inst., 70: 785-786,1983.
29. Lozzio. C. B., and Lozzio. B. B. Human chronic myelogenous leukemia cell-line with positive Philadelphiachromosome. Blood, 45: 321-334,1975.
30. MacDonató,H. R. Phenotypic and functional characteristics of T-like" cells in
nude mice. Exp. Cell Bid., 50:305,1982.31. Mahoney, K. H., Morse, S. S., and Morahan, P. S. Macrophage functions in
beige (Chédiak-Higashisyndrome)mice. Cancer Res., 40: 3934-3939,1980.32. Riccardi, C., Puccetti, P., Santoni, A., and Herberman, R. B. Rapid in vivo
assayof mouse natural killer cell activity. J. Nati. Cancer Inst., 63:1041-1045,1979.
33. Roder,J. C., and Duwe, A. The beige mutation in the mouse selectivelyimpairsnatural killer cell function. Nature (Lond.),278: 451-453,1979.
34. Roder,J. C., Lohmann-Matthes,M.-L, Domzig,W., and Wigzell,H. The beigemutation in the mouse. II. Selectivity of the natural kilter (NK) cell defect. J.Immunol., 723: 2174-2182,1979.
35. Stutman. O. Natural antitumor resistance in immune-deficientanimals ExpCell Biol.. 50: 308,1982.
36. Talmadge.J. E., Meyers, K. M., Prieur, D. J., and Starkey, J. R. Rote of NKcells in tumour growth and metastasis in beige mice. Nature (Lond.), 284:622-624,1980.
37. Wagner, H., Hardt, C., Heeg, K., Rollinghof, M., and Pfizenmater,K. T-ce»-derived helper factor allows in vivo induction of cytotoxic T cells in nu/nu mice.Nature (Lond.), 284: 278-280,1980.
38. Warner, J. F., and Dennert, G. Effects of a cloned cell line with NK activity onbone marrow transplants, tumor development and metastasis in vivo. Nature(Lond.),300: 31-34,1982.
39. Welsh, R. M. Mouse natural kilter cells: induction specificity, and function. J.Immunol., 727:1631-1635,1978.
40. Zölter,M., Andrighetto, G. C., Heyman, B., Lamon, E. W., and Wigzell, H.Characterizationof effector cells mediating IgG and IgM antibody-dependentcellularcytotoxicity. Scand. J. Immunol., 77:19-27,1983.
41. Zöller,M., and Wigzell, H. Normally occurring inhibitory cells for natural kiltercell activity. II. Characterization of the inhibitory cell. Cell. Immunol., 74: 27-39.1982.
4408 CANCER RESEARCH VOL. 44
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

1984;44:4403-4408. Cancer Res Øystein Fodstad, Carl T. Hansen, Grace B. Cannon, et al. Growth Control in Nude Mice with Different Immune DefectsLack of Correlation between Natural Killer Activity and Tumor
Updated version
http://cancerres.aacrjournals.org/content/44/10/4403
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/44/10/4403To request permission to re-use all or part of this article, use this link
Research. on August 20, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from





























