Embed Size (px)
Citation preview

Appl Microbiol Biotechnol (1989) 30:59-66 Applied Microbiology
Biotechnology © Springer-Verlag 1989
N A D H levels and solventogenesis in Clostridium acetobutylicum: new insights through culture fluorescence
Govind Rao ~ and R. Mutharasan 2
Chemical Engineering Program and Medical Biotechnology Center of the Maryland Biotechnology Institute, University of Maryland Baltimore County, Baltimore, MD 21228, USA 2 Department of Chemical Engineering, Drexel University, Philadelphia, PA 19104, USA
Summary. The metabolic relationship between the solventogenic state in Clostridium acetobutylicum and intracellular NADH levels was investigated using culture fluorescence as a technique for con- tinuous monitoring of in vivo NADH levels. Con- tinuous culture experiments showed that a transi- tion from acidogenic to solventogenic state was accompanied by a decrease in culture fluores- cence, which was interpreted as a decrease in NADH level. It appears that NADH/NAD + turnover rates may be more significant than NADH levels in determining the metabolic state of the cell. This result provides important new in- formation on regulation of the intracellular reduc- tion state in Clostridium acetobutylicum. Culture fluorescence is shown to be a useful technique for non-invasive on-line monitoring of the metabolic state in continuous acetone-butanol fermenta- tions.
Introduction
One of the most important aspects in the acetone- butanol-ethanol fermentation by Clostridium ace- tobutylicum is the regulation of the reduction state (i. e. ratio of NADH to NADH + NAD +) and its relationship to solvent formation. The primary reason for the interest in developing an under- standing of the physiological factors that in- fluence NADH levels in this microorganism is the dependence of alcohol forming pathways on NADH availability. Considerable effort has been directed by several research groups to increase NADH availability and thus increase alcohol for- mation (Jones and Woods 1986).
Offprint requests to: G. Rao
Fermentation by Clostridium acetobutylieum is characterized by two distinct phases. The first phase consists of rapid growth with active hy- drogen evolution and acetic and butyric acid pro- duction. The medium pH decreases to a charac- teristically low value (typically around 4.5) be- cause of acid formation. The second slower growth phase is accompanied by decreased hy- drogen evolution and solvent formation with par- tial uptake of the acids formed during the first growth phase. The two phases are termed acido- genic and solventogenic, respectively (Ennis et al. 1986).
In Fig. 1, the metabolic reactions of Clostrid- ium acetobutylieum are shown. Glucose is fer- mented via the EMP pathway to pyruvate. Pyru- vate oxidation to acetyl CoA is coupled to ferre- doxin (labelled as Fd) reduction and is mediated by pyruvate ferredoxin oxidoreductase (enzyme 1 in Fig. 1), leading to formation of reduced ferre- doxin (FdH2). Acetyl CoA is further metabolized to ultimately yield the end-products of the ace- tone-butanol fermentation. Ethanol formation is NADH dependent while butanol formation de- pends on both NADH and NADPH availability. As in most saccharolytic strict anaerobes, hy- drogen evolution is the primary means by which this organism rids itself of excess reducing equi- valents (Gray and Gest 1965). The enzyme re- sponsible for hydrogen evolution is hydrogenase (enzyme 2 in Figure 1) and is found in several bacteria (Adams et al. 1981).
Common to both acidogenic and solvento- genic phases, FdH2 is oxidized by hydrogenase which regenerates Fd and releases electrons as molecular hydrogen. Under acidogenic condi- tions, in additon to formation by pyruvate oxida- tion, there is a parallel pathway for FdH2 forma- tion. This takes place by NADH oxidation and
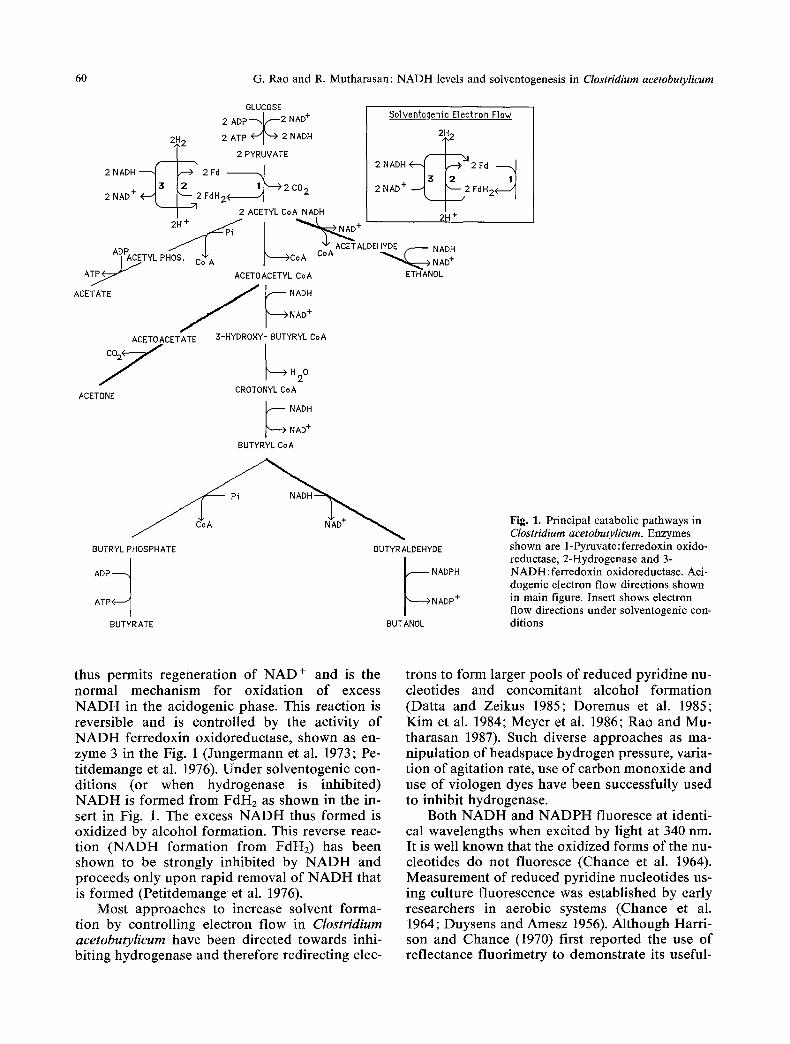
60 G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum
GLUCOSE 2 ADP--',~ -2 NAD+ Solventoqenic Electron Flow
2H 2 2 ATP - - ~ 1 2 NADH 2H2
2 NADH 2 Fd 2 NADH 2 Fd
~ _ ~ 2 2C02 2NAD+ " - - d ~ 2 Fd H 2<'~ 2 NAD + FdH2<
2 ACETYL CoA NADH 2H +
ACETALDEHYDE ADP , CoA NADH ' i ~TYL PHO"~S, Co"LA )CoA L ~ NAD +
ATP~ ~ ACETO ACETYL CoA ETHANOL . / ACETATE ~ ~ NADH
~--~NAD +
ACETOACETATE 3-HYDROXY- BUTYRYL CoA
CROTONYL CoA ACETONE ~ NADH
NAD +
BUTYRYL Co A
BUTRYL PHOSPHATE BUTYRALDEHYDE
ADP~ k NADPH
ATP(-~" r >NADP+
BUTYRATE BUTANOL
Fig. 1. Principal catabolic pathways in Clostridium acetobutylicum. Enzymes shown are 1-Pyruvate:ferredoxin oxido- reductase, 2-Hydrogenase and 3- NADH:ferredoxin oxidoreductase. Aci- dogenic electron flow directions shown in main figure. Insert shows electron flow directions under solventogenic con- ditions
thus permits regeneration of NAD + and is the normal mechanism for oxidation of excess NADH in the acidogenic phase. This reaction is reversible and is controlled by the activity of NADH ferredoxin oxidoreductase, shown as en- zyme 3 in the Fig. 1 (Jungermann et al. 1973; Pe- titdemange et al. 1976). Under solventogenic con- ditions (or when hydrogenase is inhibited) NADH is formed from FdH2 as shown in the in- sert in Fig. 1. The excess NADH thus formed is oxidized by alcohol formation. This reverse reac- tion (NADH formation from FdH2) has been shown to be strongly inhibited by NADH and proceeds only upon rapid removal of NADH that is formed (Petitdemange et al. 1976).
Most approaches to increase solvent forma- tion by controlling electron flow in Clostridium acetobutylicum have been directed towards inhi- biting hydrogenase and therefore redirecting elec-
trons to form larger pools of reduced pyridine nu- cleotides and concomitant alcohol formation (Datta and Zeikus 1985; Doremus et al. 1985; Kim et al. 1984; Meyer et al. 1986; Rao and Mu- tharasan 1987). Such diverse approaches as ma- nipulation of headspace hydrogen pressure, varia- tion of agitation rate, use of carbon monoxide and use of viologen dyes have been successfully used to inhibit hydrogenase.
Both NADH and NADPH fluoresce at identi- cal wavelengths when excited by light at 340 nm. It is well known that the oxidized forms of the nu- cleotides do not fluoresce (Chance et al. 1964). Measurement of reduced pyridine nucleotides us- ing culture fluorescence was established by early researchers in aerobic systems (Chance et al. 1964; Duysens and Amesz 1956). Although Harri- son and Chance (1970) first reported the use of reflectance fluorimetry to demonstrate its useful-

G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum 61
ness in measuring intracellular NAD(P)H levels in continuous cultures of K. aerogenes, this tech- nique has not been widely applied. The principal reason is probably due to the unavailability of a reliable probe for measuring fluorescence on-line for extended periods of time. Although commer- cially available spectrofluorometers can be used to measure the fluorescence of a culture by circu- lating it through a flow cell, the time delay due to the flow from the reactor to the sample cuvette may lead to a significant change in NAD(P)H lev- els resulting in ambiguous measurements (Harri- son and Chance 1970). Another drawback is that most conventional spectrofluorometers use high pressure xenon lamps that offer stable operation for only a few hours at a time and also suffer from relatively short lamp life times (typically less than 500 h). The availability of a new fluorescence probe designed specifically to monitor NAD(P)H reliably and continuously has significantly simpli- fied culture fluorescence measurements.
While it is clear that a higher throughput of NAD(P)H is required for increased alcohol for- mation, it is not clear whether this throughput takes the form of an increased pool size (concen- tration) of NAD(P)H or by an increase in turn- over rate of NAD(P)H/NAD(P) + (Rao and Mu- tharasan 1987). Our focus in all experiments dis- cussed in this paper will simultaneously be on flu- orescence (which is a measure of NAD(P)H pool sizes), and on butanol concentration which is a measure of the consumption of NAD(P)H. It has been reported that in a number of microorgan- isms including a species of Clostridium, NADPH levels are lower than NADH levels by an order of magnitude (London and Knight 1966). Conse- quently, culture fluorescence in Clostridium aceto- butylicum is primarily proportional to intracellu- lar NADH levels. Further, since each mole of bu- tanol formed requires three moles of NADH and only one of NADPH (Jones and Woods 1986), our discussion henceforth will be on NADH lev- els.
In this study we will examine the variation in NADH levels (using culture fluorescence) in steady- state, glucose limited continuous cultures of Clostridium acetobutylicum when solventogenesis was induced. The significance of glucose limita- tion is that it permits altered electron and carbon flow to the products to be readily and unambi- guously discerned. Solventogenesis induction was accomplished in three different ways; i) by addi- tion of methyl viologen (MV) ii) by a change in pH from 6.8 (acidogenic) to 5.1 (solventogenic) and iii) by addition of butyrate to an acidogenic
culture (pH 6.1). All three techniques have been previously shown to induce solventogenesis (Rao and Mutharasan 1987; Jones and Woods 1986; Monot et al. 1983).
Materials and methods
Clostridium acetobutylicum ATCC 824 was maintained at 4°C in Reinforced Clostridial Medium (Oxoid, Basingstoke, Eng- land). A growth medium containing (NH4)2804, KH2PO4, K2HPO4, MgSO4, FeSO4, MnSO4, NaC1 and yeast extract at 3.0, 0.8, 0.8, 0.2, 0.01, 0.01, 0.02 and 4 g - 1-1 respectively in distilled water was used. The feed glucose concentrations for the viologen addition, pH change and butyrate addit ion ex- periments were 17.5, 14.8 and 15 g • l -a, respectively.
Glucose was autoclaved separately from the rest of the medium components. Autoclaving was done at 121°C for 20 min. Steady state was determined to have been reached when hydrogen, glucose and optical density measurements were found to be within 5% over three residence times. This was reconfirmed later with liquid product concentrations.
Fermentat ion was carried out at 500 ml working volume in 1 liter Virtis fermenters (Virtis, Gardiner, New York). pH was measured and logged continuously by an Apple I1 com- puter. The same computer also controlled pH by automatic addition of 4N NaOH. The culture broth was sparged with oxygen-free nitrogen prior to and after inoculation to ensure anoxic conditions. Sparging was maintained throughout all the experiments. All tubing used was made of viton to elimi- nate oxygen entry. Inocolum was 10 percent by volume and in exponential growth phase. Incubation and growth were at 37°C. Samples were centrifuged at 10000 g for 5 rain and the supernatant liquid stored at 4 °C until they could be ana- lyzed.
Liquid products were acidified and analyzed on a Shi- madzu GC (Model 9 A) equipped with a six foot AT-1600 3% T-Port A (Alltech Ass., Deerfield, Ill) column and a FID. The column temperature was held at 100°C for three minutes fol- lowed by a ramp of 10°C/min to 160°C where it was held until the last component (butyric acid) eluted. Injector and de- tector temperatures were at 200 ° C. Glucose was measured on a YSI (Model 27) glucose analyzer. Optical density (OD) was determined at 540 nm on a Shimadzu UV-160 spectrophotom- eter ( 1 0 D unit = 0.293 g/1 cells).
Fluorescence was measured using the FluroMeasure sys- tem (Biochem Technology, Malvern, PA). The FluroMeasure probe consists of a mercury lamp, excitation (360 nm) and emission (460 nm) filters and solid state photodetectors in a self contained unit. The detector signal is read using an IBM PC. Culture fluorescence is reported as Normalized Fluores- cence Units (NFU). One NFU corresponds to a change in flu- orescence caused by a change in NADH concentration of 0.122 lxM at 30°C, pH 8 in the concentration range 1 to 25 txM NADH. The signal from the detector is proportional to the NAD(P)H levels in the culture. The FluroMeasure probe was mounted on the exterior of (in direct contact with) the fer- menter glass (Pyrex) vessel. The optical path of the probe was aligned to be orthogonal to the glass surface. The signal was found to be at tenuated by the glass by approximately 10 NFU. Since fluorescence measurement is interferred with by extra- neous light, the entire apparatus was kept covered with black polyethylene bags throughout all experiments.
In the MV addition experiment, the viologen was added in its oxidized (colorless) form as a 1 g/1 solution. Reduced

62 G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum
MV is blue in color, and reduction took place as soon as the viologen was introduced into the fermenter. For the butyrate addition experiment, butyrate was added as its sodium salt.
Although fluorescence measurements reported here are primarily a measurement of NADH levels, biomass variations can affect the measurements. Consequently, our analysis will focus on specific fluorescence measurements. Specific fluores- cence was calculated as N F U / X where N FU is the instanta- neous fluorescence value and X is the instantaneous cell mass. Additionally, since butanol concentration does not give a pic- ture of the rate at which the metabolic change (if any) is taking place, our discussion will compare variations in specific fluo- rescence with butanol productivity. Butanol productivity, QB, was calculated as (DB + dB/dt)/X where D is the dilution rate, B the instantaneous butanol concentrat ion and dB/dt the rate of change of butanol concentration. Butanol productivity is also a measure of the NADH oxidation rate (only with respect to the related metabolic reactions).
It has previously been shown that for a particular species growing under similar conditions, the total pyridine nucleotide pool (i.e., N A D H + N A D +) size is constant (London and Knight 1966). Since the total pyridine nucleotide pool is rela- tively constant, an increased oxidation rate of NADH can be interpreted as an increased turnover rate of the N A D H / NAD + couple (Rao and Mutharasan 1987).
Results and discussion
Solventogenesis induced by MV addition
One of the problems encountered in the measure- ment of culture fluorescence in presence of MV was the so called "inner filter effect" caused by components in the medium absorbing either (or both) the excitation or the emission light energy. Since MV was in its reduced (colored) form in this experiment, it was determined through another set of experiments performed under identical con- ditions that the additon of 1 mg/1 of MV and its subsequent reduction led to an attenuation in flu- orescence of 5 N F U (Rao 1987).
In Fig. 2A, the fluorescence responce of a con- tinuous culture of Clostridium acetobutylicum at pH 6.55 to a step change in MV concentration is presented. Figure 2B shows the variations in buta- nol and culture fluorescence profiles for the same duration, while Fig. 3 shows the computed pro- files of butanol productivity and specific fluores- cence. For the initial period when MV concentra- tion was constant at 4.3 mg/ l in the fermenter, the fluorescence value increased by about 100 NFU. At the same time, butanol increased from 0 to about 0.3 g/l. When the MV concentration began to increase, the fluorescence value decreased to 400 NFU. In the same period, butanol concentra- tion further increased to 0.72 g/1. A further in- crease in fluorescence from 400 to 625 N F U was seen to take place after MV concentration had
650. 7.0
600 -
550- z
500-
o ,.-.' 450.
400 -
550 40
• , i , , , , ,
s 0 60 70 80 90 100 110 120 150 TIME, (hr)
A
6.5
6.0 E v z" w
5.5~ 2 >
5 . 0 ~ : m T
4.5
4,0 14O
650 - . 0.8 B
, 0 0 + 7
z 0,S
w 500 • 0.4 J
U l <
o~ 0.3 o 450•
0.2
400 - y ~ FLUORESCENCE ~°°'~' 0. I
350 - I I I I I I I I 0.0
40 SO 60 70 80 90 100 110 120 130 140
TIME, (hr)
Fig. 2. A. Fluorescence and MV concentration profiles. B. Fluorescence and butanol concentration profiles. Culture con- ditions: pH =6.55, D=0.195 h -~
reached a constant value. Butanol formation was essentially shut off during this period (although the reason for this is not known at this point). In the last 10 h, fluorescence decreased 125 NFU, while no change in butanol concentration was ob- served. Throughout the experiment, the chemostat was glucose limited as evidenced by the low ef- fluent glucose concentration ( < 0.1 g/l).
As can be seen from Figure 2B, for the most part there is a distinct inverse relationship be- tween butanol concentration and culture fluores- cence. It could be argued that the decrease in flu- orescence observed in this period is caused by the increase in MV concentration giving rise to more reduced MV and causing fluorescence attenua- tion. However, the increase in MV concentration is only 2.5 mg/l and hence it is unlikely that this factor alone was the cause of the decrease in fluo- rescence. It is interesting to note that the relation- ship between specific fluorescence and butanol productivity (Fig. 3) closely parallels the fluores- cence and butanol concentration, suggesting that the uncompensated fluorescence signal alone may be sufficient for on-line sensing of the state of a culture.

G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum 63
520
3 0 0 .
~ 2 8 0 .
~ z 2 6 0 '
~ 240 . a~
2 2 0
200
e_ co 180
150
40
70
00/ \ /\o
\ 77, % _ j \_..... 1o
I I l i I ] I 0 50 60 70 80 90 100 110 120 130 140
TIME, ( h r )
s 0 ~
40
v 30 -
o 20
Fig. 3. Specific fluorescence and butanol productivity profiles for the MV addition experiment shown in Fig. 2. Culture con- ditions: pH =6.55, D=0.195 h -1
The results of the experiment discussed above showed that when the cells produced butanol a) NADH levels could increase (as seen between 50 and 70 h in Fig. 2A and B) with increase in buta- nol production, b) decrease (70 to 120 h) and c) show no change (120 to 130 h). These results did not seem to allow a definite conclusion to be drawm on the relationship between solventogene- sis and the reduction state of the cell.
The known interference of reduced MV also introduced ambiguity on the results, even though the range of fluorescence variation observed was far greather than any effects that MV could have had in the concentration that it was used in.
However, there is supporting evidence from recent measurements of Reardon et al. of fluores- cence of immobilized cells of Clostridium acetobu- tylicum which show a trend similar to the results of the viologen additon experiment (Reardon et al. 1986). They observed that butanol and fluores- cence profiles followed each other monotonically for a few hours. However, when a nitrogen source was added to the medium, fluorescence showed a sharp decrease concomitant with a sharp increase in butanol much like the observations in the vio- logen experiment just described earlier. They con- cluded that there was a change in the mechanism of NADH oxidation causing the sharp decrease in fluorescence. Since there were no color changes in their experiment and they observed similar re- suits, it is quite possible that MV interference in the experiment just discussed may not have been significant. In any case, other experiments were performed whereby solventogenesis was induced by means other than viologen addition so that the fluorescence-butanol relationship could be eluci- dated with greater certainty.
Solventogenesis induced by pH switchin9
To further examine the relationship between sol- ventogenesis and fluorescence, several experi- ments were performed where solventogenesis was induced at steady state by changing culture pH. Similar results were obtained in all of them. In the experiment reported here, the pH was changed from 6.8 (acidogenic) to 5.1 (solventogenic). The pH switch was accomplished by changing the set- point and allowing the system to reach the new pH of its own accord due to acid production. Fig- ure 4A shows the resulting fluorescence and buta- nol profiles. As soon as the pH was decreased, butanol production increased (as expected). Once again, the important observation is that fluores- cence and butanol have an inverse relationship. As in the other experiments, the chemostat was glucose limited during the entire period. Based on the data from Fig. 4A, butanol productivity and the specific fluorescence were calculated. These results are plotted in Fig. 4B and it was observed that as soon as the pH was decreased, there was a sharp increase in butanol productivity concomi- tant with a sharp decrease in specific fluores-
700 • 0.9
BUTANOL m ~ ~ 0.8
n ~ l l • 0,7
.0 .6
.0,4 - * ~ ' ~ ' ~ o " J ~ - 0.3 m
ORESCENCE - 0.2 400 l ~ j m ~ l ~ ' m
-0.1
350 4 - a ' A , ~ i I I l i 0.0 40 60 I " 80 I O0 120 140 160 80
TIME, (h r )
45 280
\ ! ° 35 • t -~' - 240
t~ 30 IFIC FLUORESCE CE , 2 2 0 v z
'o j 25 , 200 ~
20 180~
u~ I 0 . 1 4 0 ~
5 . lZO
0 ' ~ m ~ ---'~e° 810 I I I I 1 O0 40 60 1" 100 120 140 160 180
TIME, (h r )
Fig. 4. A. Fluorescence and butanol concentration profiles. B. Specific fluorescence and butanol productivity profiles for corresponding period. Culture conditions: pH changed from 6.8 to 5.1 at 68.5 h. D = 0 . 1 4 h ]

64 G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum
cence. Between 80 and 100 h the specific fluores- cence was nearly constant while butanol produc- tivity showed a sharp decrease. Around 120 h, for a small decrease in butanol productivity, specific fluorescence showed a large increase.
As in the MV experiment, it appears that al- though there is an inverse relationship for the most part, the magnitude is not constant. It is not clear as to why at some times small changes in butanol productivity produce large changes in flu- orescence, and at other times both show large changes. It should be pointed out that since buta- nol productivity is calculated based on differ- ences in butanol concentrations between consecu- tive samples, it is sensitive to measurement errors in butanol concentration. As a result, small errors lead to inaccurate calculations of productivity and may be partly responsible for the fluctuations seen in the productivity data.
It is interesting to note that in this experiment also, the uncompensated fluorescence value alone can serve as a measure of the metabolic state of a culture.
A brief mention must be made of the relation- ship between pH and fluorescence. It is difficult to characterize pH effects on fluorescence accu-
rately as the chemical properties of individual fluorophores themselves are affected to varying degrees by pH. It is therefore not possible to con- elude if any observed change in fluorescence due to a pH change is an intrinsic change in fluores- cence only or whether it is a result of chemical inactivation of the fluorophore. For example, Reardon et al. (1987) reported that the fluores- cence of (anaerobic) NADH in water was essen- tially independent of pH above pH 6.0. Below pH 6 they observed a (linear) 15% decrease in fluores- cence for a unit decrease in pH, but it is not clear if this is a chemical effect.
However, since culture fluorescence measure- ments reported here are of intracellular NADH and the intracellular pH in Clostridium acetobuty- licum is tightly regulated, it is unlikely that any variation in fluorescence measurements reported here were significantly affected by pH. Huang et al. (1985) measured intracellular pH in Clostrid- ium acetobutylicum and found that varying the ex- ternal pH from 6.0 to 4.5 resulted in the internal pH dropping from 6.46 to 6.17. Other workers have reported similar results (Jones and Woods 1986).
12 0.11 j \ / . \ .o,o I0 0.09
8 ANOL 0.08
~" " 0.06
>- 0.05
4 - 0,04
0.03 2-
0.02
0 I I I I I I i 0.01
0 S 10 1S 20 25 30 35 40 TIME, (hr)
0.30 - 170
/\ /\ 0.25. 180
15o b o.20
~, 0.15. Q 140 z ~ ~" v
150 X 0,I0 o
120
0.05 110
0.00 I O0 {n
SRECI FIC FLUORESCENCE
-O.OS 51 110 IlS 201 251 slo 3 I 5 40 90
TIME, (hr)
Fig. 5. A. Butanol formation in response to butyrate additon. B. Specific fluorescence and butanol productivity profiles for corresponding period. Culture conditions: pH = 6.1, D=0.178 h -1. Butyrate added at 10 h to fermenter
Solventogenesis induced by butyrate addition
In addition to the pH change experiments, buty- rate addition as a means of inducing solventogen- esis was also carried out. In this experiment also, as in the pH switch experiments, a change in physiological state was obtained without the ob- fuscating effects of color changes caused by re- duced MV. The results are shown in Fig. 5A and B. The pH was controlled at 6.1 and glucose limi- tation was observed at all times. Butanol was pro- duced in response to butyrate added (Fig. 5A). When butanol productivity increased, specific flu- orescence showed a small decrease of about 10 units (Fig. 5B). When butanol productivity de- creased in response to the decrease in butyrate concentration, the specific fluorescence showed a corresponding increase of around 60 units.
Having seen the same phenomenon taking place in three different experiments under a vari- ety of conditions and based on similar observa- tions of Reardon et al. (1986) we can generalize that fluorescence (and therefore intracellular NADH levels) and butanol production have an inverse relationship. However, as seen from all the experiments thus far, the relationship does not appear to have any quantitative aspect to it. It does-appear that the qualitative information that

G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum 65
a change in fluorescence continuous fermentation can be applied to detect a corresponding change in butanol productivity.
Regulation of the reduction state in Clostridium acetobutylicum
The observation that butanol and fluorescence show an inverse relationship has an important im- plication on the understanding of the role of the reduction state on the metabolism of Clostridium acetobutylicum. As was pointed out earlier, many investigators have rationalized that since alcohol producing pathways are N A D H dependent, an in- crease in butanol /e thanol yields implies that a larger pool size of N A D H is present. The results presented here belie this conclusion and show that the opposite is generally true. It is seen from culture fluorescence measurement that N A D H pool sizes can be actually smaller when the cul- ture is solventogenic. This observation is rein- forced by the exhaustive in vitro experiments of J~mgermann et al. (1973) and Petitdemange et al. (1976). They have shown that the reaction leading to NADH formation from FdH2 is effectively in- hibited by N A D H at low levels with a K/(inhibi- tion constant) of 0.01 mM. N A D H is believed to be a competitive inhibitor to NAD + and it has been shown that FdH2 oxidation proceeds only in the presence of an NAD + regenerating system or, in other words, an effective sink is required for NADH. In vivo, the alcohol producing pathways are believed to serve as the sinks for NADH (Pe- ti tdemange et al. 1976). However, from a stoi- chiometric analysis of fermentation by Clostrid- ium acetobutylicum, it is clear that there can be a net generation of N A D H from FdH2 under sol- ventogenic conditions (Papoutsakis 1984). We also know that this reaction is strongly inhibited by NADH. Therefore, the only way to satisfy si- multaneously the two conditions of smaller NADH pool size but greater NADH utilization is to have a greatly increased turnover rate of N A D H / N A D + . It is proposed that this is in fact what is happening in vivo whenever fluorescence is observed to sharply decrease with a corre- sponding increase in butanol productivity. As pointed out earlier, butanol productivity is a measure of the turnover rate of NADH. It would thus seem that the N A D H consumption rate would have to increase whenever NADH produc- tion from FdH2 takes place to forestall inhibition by N A D H accumulation.
Additionally, as seen from the experiments re- ported in this paper and those reported by others, N A D H levels can also show relatively small in- creases when butanol is being formed (Reardon et al. 1986).
All the above observations indicate that the regulation of the reduction state is controlled by two regimes. One is when the N A D H pool size is large and is manifest as slightly increasing or al- most constant fluorescence and concomitant in- creases in butanol concentration. The other re- gime is when the NADH turnover rate is high and is evidenced by sharp decreases in fluorescence coupled with sharp increases in butanol forma- tion. A possible mechanism governing which re- gime is operating is likely to be the direction of the reversible reaction:
NADH + Fd ~ NAD + + FdH2 (1)
When the forward reaction dominates, NADH levels in the cell can show increases concomitant with alcohol formation. When the reverse reaction is operative, NADH levels must be kept low to enable the reaction to proceed and consequently, the turnover rate of NADH must be high. The factors that govern the direction of the above reaction are not clear at this point. However, probable factors include be the activities of branch point enzymes, or the intracellular redox potential as given by the N A D H / N A D + ratio or the CoA/Acetyl-CoA ratio, all of which are known to affect the reduction state of the cell. Given the dynamic nature of enzyme regulation in general, it is quite conceivable that a combination of these factors as well as others may be at work to determine the reduction state of the cell.
One of the factors actively being researched is continuous solvent production using Clostridium acetobutylicum. A major problem in this effort is sustaining solvent productivity over extended pe- riods (Ennis et al. 1986). Use of culture fluores- cence as an on-line monitoring (and perhaps con- trol) tool for the metabolic state of the culture may provide information that could lead to con- trol strategies that could be used to stabilize con- tinuous butanol formation. Bioreactor operating strategies to achieve stable and continuous sol- vent production are feasible by using culture fluo- rescence as an on-line sensor to determine if the culture is switching from solventogenic to acido- genic and a corrective action such as lowering pH, CO sparging or viologen additon could be used. Since several commercially important ethanol forming anaerobic microorganisms have electron

66 G. Rao and R. Mutharasan: NADH levels and solventogenesis in Clostridium acetobutylicum
flow pathways similar to that of Clostridium aceto- butylicum, the results presented here can poten- tially be applied to other systems as well.
References
Adams MWW, Morenson LF, Chen JS (1981) Hydrogenase. Biochim Biophys Acta 594:105-176
Chance B, Estabrook RW, Ghosh A (1964) Damped sinusoi- dal oscillations of cytoplasmic reduced pyridine nucleotide in yeast cells. Proc Nat Acad Sci USA 51:1244-1251
Datta R, Zeikus JG (1985) Modulation of acetone-butanol- ethanol fermentation by carbon monoxide and organic acids. Appl Environ Microbiol 49:522-529
Doremus MG, Linden JC, Moreira AR (1985) Agitation and pressure effects on acetone-butanol fermentation. Biotech- nol Bioeng 27:852-860
Duysens LNM, Amesz J (1956) Fluorescence and spectropho- tometry of reduced phosphopyridine nucleotide in intact cells in the near-ultraviolet and visible region. Biochim Biophys Acta 24:19-26
Ennis BM, Gutierrez NA, Maddox IS (1986) The acetone-bu- tanol fermentation: a current assessment. Proc Biochem 21:131-147
Gray CT, Gest H (1965) Biological formation of molecular hy- drogen, Science 148:186-192
Harrison DEF, Chance B (1970) Fluorimetric technique for monitoring changes in the level of reduced nicotinamide nucleotides in continuous cultures of microorganisms. Appl Microbiol 19:446-450
Huang L, Gibbins LN, Forsberg CW (1985) Transmembrane pH gradient and membrane potential in Clostridium aceto- butylicum during growth under acidogenic and solvento- genic conditions. Appl Environ Microbiol 50:1043-1047
Jones DT, Woods DR (1986) Acetone-butanol fermentation revisited. Microbiol Revs 50:484-524
Jungermann K, Thauer RK, Leimenstoll G, Decker K (1973) Function of reduced pyridine nucleotide ferredoxin oxi- doreductase in saccharolytic clostridia. Biochim Biophys Acta 305:268-280
Kim BH, Bellows P, Datta R, Zeikus JG (1984) Control of car- bon and electron flow in Clostridium acetobutylicum fer- mentations: utilization of carbon monoxide to inhibit hy- drogen production and to enhance butanol yields. Appl Environ Microbiol 48:764-770
London J, Knight M (1966) Concentration of nicotinamide nu- cleotide coenzymes in micro-organisms. J Gen Microbiol 44:241-254
Meyer CL, Roos JW, Papoutsakis ET (1986) Carbon monox- ide gassing leads to alcohol production and butyrate up- take without acetone formation in continuous cultures of Clostridium acetobutylicum. Appl Microbiol Biotechnol 24:159-167
Monot F, Engasser JM, Petitdemange H (1983) Regulation of acetone butanol production in batch and continuous cul- tures of Clostridium acetobutylicum. Biotechnol Bioengg Symp 13:207-216
Papoutsakis ET (1984) Equations and calculations for fermen- tations of butyric acid bacteria. Biotechnol Bioengg 26:174-187
Petitdemange H, Cherrier C, Raval G, Gay R (1976) Regula- tion of the NADH and NADPH-ferredoxin oxidoreduc- tases in Clostridia of the butyric Group. Biochim Biophys Acta 421 : 334-347
Rao G (1987) Directed metabolic flow in Clostridium acetobu- tylicum. Ph.D. Thesis. Drexel University, Philadelphia, PA
Rao G, Mutharasan R (1987) Altered electron flow in contin- uous cultures of Clostridium acetobutylicum induced by viologen dyes. Appl Environ Microbiol 53:1232-1235
Reardon KF, Scheper T, Bailey JE (1986) In situ fluorescence monitoring of Clostridium acetobutylicum. Biotech Lett 8:817-822
Reardon KF, Scheper T, Bailey JE (1987) Metabolic pathway rates and culture fluorescence in batch fermentations of Clostridium acetobutylicum. Biotechnol Prog 3:153-167
Received May 13, 1988/Accepted August 24, 1988










![Genome-Wide Analysis of Cell Type-Specific Gene ...aoh/Saujetetal.pdf · also the case for key genes involved in the spore morphogenesis [16,17,20]. In Clostridium acetobutylicum](https://img.pdfslide.us/doc/110x75/5e87518797913649b022fe05/genome-wide-analysis-of-cell-type-specific-gene-aoh-also-the-case-for-key.jpg)


















