Embed Size (px)
Citation preview

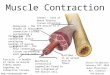
Muscle Physiology and Contraction TheoriesBy RICHARD J. PODOLSKY, PH.D.
The structural basis of current contraction theories is the double array of thick and thinmyofilaments revealed by the electron microscope. The physiologic properties that characterizethe contractile process are natural consequences of this structure if (a) during shorteningthe 2 sets of filaments move relative to each other and (b) the flux of chemical energythrough the contractile mechanism is limited by interaction between complementary sitesdistributed along the 2 sets of filaments. Contraction models fitted to these ideas differlargely in the mechanism by which force is generated. In the sliding model, force isdeveloped by mechanical interaction between the thick and thin filaments, and filamentlength remains constant during contraction. In the folding model, force is developedin the thin filament, which shortens during contraction. Both models quantitatively accountfor the force-velocity relation and the Fenn effect. They also accommodate the quick-releasephenomenon and predict, at least qualitatively,seen after quick release from tetanic tension.
B IOCHEMICAL PROCESSES in musclecells have 2 exceptional characteristics.
The first, of course, is that they generatelarge forces. The second, which is not quiteso obvious, is that they proceed at a rate whichdepends on the motion of the cell. Manyproperties of contracting muscle can be tracedback to the influence of the motion of the cellon the chemical processes driving the con-tractile mechanism, an idea first proposed byFenn.1
It seems very natural that a muscle shouldlift a light load more rapidly than a heavierload. However, as Fenn wrote some yearsago, "the more one tries to explain these sim-ple facts, the less obvious do they seem to be. 11
The "simple" fact is shown in figure 1, theforce-velocity curve for the classical striatedmuscle, the frog sartorius. The circles aredata from an experiment we shall discusslater. The smooth curve is A. V. Hill's force-velocity relation.3 The question is: Why doesthe contractile force fall with the velocity?
In the early twenties, the force-velocityrelation was explained with a viscoelasticmodel4 along the following lines: Upon acti-vation, muscle becomes an elastic filament,like a spring. The spring is immersed in aviscous fluid. When the muscle shortens agiven distance, the available potential energy
From the Naval Medical Research Institute,National Naval Medical Center, Bethesda, Maryland.
the isotonic velocity transients that can be
appears either as work or as heat. The morequickly the muscle moves, the greater theviscous force, the greater the heat produc-tion, and the smaller will be the energy avail-able for work. Thus the fall in force withspeed.The essential part of the viscoelastic model
is that, in shortening a given distance, theavailable energy is independent of the me-chanical conditions of contraction. The corol-lary, that the amount of heat produced in acontraction increases with the speed, wasdoubted as long ago as 1864.5 The unambig-uous experiments of A. V. Hill confirmedthese doubts.
Figure 2, taken from Hill's classical 1938paper,3 shows the critical experiment.* CurveE is the total heat produced as a function oftime when the frog sartorious is tetanized iso-metrically. Curves F, G, H, and J show theheat liberated when the muscle is released,at the time indicated by the first arrow, andallowed to shorten a given distance. Althoughthe speed of contraction in J, say, is 5 timesgreater than in F, the total extra heat due toshortening does not change. Since it wasclear that heat production did not increasewith speed, the viscoelastic theory wasdemolished.
*Figure 2 reproduced from Hill: Proc. Roy. Soc.London s.B 126: 136, 1938.' By permission of theRoyal Society of London.
Circulation, Volume XXIV, August 1961 399
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

PODOLSK Y
P/P mox
FROG SARTORIUS00c
V/V moxFigure 1
Relation between force, P, and velocity, V, in liv-ing muscle. Experimental points are calculatedfrom data of figure 12; smooth curve is the force-velocity equation of A. V. Hill.3
The error, of course, is that the energyavailable during contraction is not constant.The flux of chemical energy into the muscleis somehow affected by the motion itself. Inother words, some kind of mechanochemicalcontrol is built into the contractile mechanism.The force-velocity relation might be a re-flection of this mechanochemical control ifthe tension generating process were continu-ously opposed, or inactivated, by the mechan-ical motion.
This idea is schematized in figure 3. Weassume that a sequence of chemical reactionsdrives the contractile mechanism. One ofthese reactions, M, is closely linked to thecontractile mechanism. Since the chemicalreactions are themselves linked like a trainof gears, the extent of the reaction during acontraction is limited by the turnover of M,which, in turn, is some function of the short-ening velocity. The total energy producedduring contraction depends on the extent ofthe reaction A + M. This chemical energy*
*In this paper, " energy " denotes the enthalpyof reaction, AIH=AU + PAV. The energy availablefor mechanical work is the free energy, AH - TAS.In these expressions, U is internal energy, P ispressure, V is volume, T is temperature, and S isentropy.
(a.)
/ ~~~~~~~~(ad
/ I I ~~~I I -I1
Time marks:O 2sec
Figure 2Heat production of living muscle. Tetanicallystimulated frog sartorius at 0 C. (a) A: isometriccontraction. B, C, D: 1.2 sec. after start of stimu-lus, muscle is released and allowed to shorten vari-ous distances (B < C < D) under constant load.(b) E: isometric contraction. F, G, H, J: muscleis allowed to shorten the same distance undervarious loads (F > G > H > J). (From Hill.3)
is partitioned between work and heat. Con-versely, the rate of chemical reaction can beinferred from the rate of total energy pro-duction. The trick in devising contractiontheories is to make the link between mechani-cal and chemical processes such that both thetotal energy production and its partition intowork and heat depend on mechanical motionin just the right way. We shall first showwhat the "right way" is, and then describehow several models manage to do this.
Before leaving figure 3, I should like tomention that there is independent evidence,from certain heat measurements, that thisrepresentation of the sequence of events isclose to the truth.6 7 Perhaps the best justi-fication, though, is that it accommodates thephysiology of muscle very comfortably.
Energy ProductionHow does the rate of chemical reaction
change with the rate of mechanical contrac-tion in living muscle? This can be inferredfrom the velocity dependence of the energyflux (fig. 4).
Consider the heat first. You will recallthat Hill's heat measurements showed thatthe heat of shortening is independent ofcontraction speed (fig. 2). This means that
Circulation. Volume XXIV, August 1961
400
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

MIUSCLE PHYSIOLOGY AND CONTRACTION THEORIES
NET CHEMICAL REACTION = A+M
A M
B C ( 0 /; \4 ....................contractilemechanism
HEAT WORKFigure 3
Scheme for chemical processes associated withmuscular contraction.
the rate of heat production must increaselinearly with speed (fig. 4, open region).The rate of work production also depends
on speed: the dependence can be calculatedfrom the force-velocity relation. When themuscle does not move (V = 0) it produces nowork. Also, when it is unloaded (P = 0) nowork is done. This ties down the ends ofthe work-flux curve (fig. 4, shaded region).Adding the heat to the work, we see that thetotal energy flux increases monotonically, butnot linearly, with speed. The rate of therate-limiting chemical reaction, the one linkedto the contractile mechanism, must also in-crease with speed in exactly the same way.To be acceptable, a contraction theory shouldyield this relation naturally and quantita-tively.
Structural Basis of Contraction TheoriesWhat mechanism regulates muscle chemis-
try according to speed? The double arrayof filaments revealed in Hugh Huxley's beau-tiful electron micrographss provides a clue.Figure 5 shows the characteristic thick andthin filaments.* The thick filaments definethe A-band. A second set of thinner filamentsextend from the Z-line, through the I-band,into the A-band, there interdigitating with
*Figure 5 reproduced from H. E. Huxley: J. Bio-phys. & Biochem. Cytol. 3: 631, 1957.8 By permissionof the Journal of Biophysical and Biochemical Cy-tology.
Circulation, Volume XXIV, August 1961
ENERGY FLUX
4
3
2
1.0V/V max
Figure 4Relation between energy flux and velocity in livingmuscle. Open region: rate of heat production.Shaded region: rate of work production.
the thick filaments. The part of the structurethat interests us is diagrammed at the top offigure 6.Andrew Huxley and Rolf Neidergerke dem-
onstrated that when living muscle shortens,the decrease in length takes place almost en-tirely in the light I-bands, the width of thedark A-bands remaining constant.9 Thereare two obvious ways for this to come about:the filaments could slide along each other or,after anchoring its ends, the thin filamentcould shorten by folding (fig. 6). In bothschemes only the I-band shortens. Also, andthis is important in what follows, in bothschemes there is relative motion between the2 sets of filaments. This means that if reac-tive sites were distributed along both thethick and the thin filaments, and if some kindof interaction between these sites were stochio-metrically linked to the driving chemical re-action, the relative motion of the myofilaments-and therefore the sites-would provide anatural basis for introducing velocity as aparameter in the chemical kinetics.
Implications of Relative Motion:Energy Production
The basic idea is shown in figure 7. Sites
401
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

PODOLSKY
Figure 5Double array of filavments in striated muscle. Electron micrograph of longitudinal sectionof saromere (length = 2.5 microns). (From H. E. Huxley.8)
D are distributed along the thick filament andcomplementary sites, K, are distributed alongthe thin filament. We assume that interac-tion may take place when these sites pass eachother. If interaction does take place, a sub-strate molecule, M, is used up, and the drivingchemical reaction proceeds one step.*
Proximity Time Is Rate LimitingTo understand the influence of velocity in
the kinetics of such systems, consider first acase in which the probability of interactiondepends on the time K spends in the neighbor-hood of D: the shorter the time, that is, thehigher the speed, the lower the probabilitywill be. The interrupted line in figure 8shows how the probability decreases withspeed if the K-D interaction is first order in
*In the following, a helpful mnemonic is to readD as " Dragon," K as "Knight," and M as" Maiden. "
time. Since at each interaction the drivingchemical reaction proceeds 1 step, the inter-action probability in a transit of K past Dwill also be proportional to the energy re-leased for a given amount of shortening. Thedecrease in this quantity with velocity can beinterpreted as the Fenn effect.'To calculate the number of interactions per
unit of time, we must remember that the num-ber of chances a given K site will have tointeract with D sites-the encounter rate-isproportional to the velocity (dotted line, fig.8). The number of successful interactionswill be the product of the probability and theencounter rate (solid line, fig. 8). Both thisfunction and the rate of the driving chemicalreaction in living muscle, calculated from theenergetics (fig. 4), depend on velocity inthe same way. We conclude, then, that ifcomplementary discrete sites were distributedalong the 2 kinds of myofilaments, the rela-
Circulation, Volume XXIV, August 1961
402
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

MUSCLE PHYSIOLOGY AND CONTRACTION THEORIES
I A I1 7 11 11 - lo >7
L n L//,,,,,,,,,,,,z,, _ _ ,
A,,,,,,,,,,,,,,,,, _ _ Ad,..............
=~~~~~~~~,I
= ~ ~~ ~ ~
I~~~,
Sliding Mechanism
acti vation I
shorteningg\aio= h=i
1=
Folding MechanismFigure 6
Hypothetical mechanisms for m uscultr contraction. Upper: Configuration of thick andthin filaments in resting muscle (after fig. 5). Lower: Change inz filamlent shape andconfiguration in sliding and folding contraction models.
tive velocity of the mnyofilanments could (con-trol the rate of interaction between thesesites in exactly the same way as it controlsthe release of chemical energy in livingmuscle.
Filling Time Is Rate Limiting
A physically different way of describinginteraction between sites on movinog filamentsfocuses on the time spent betwwecni, ratherthan at, encounters. In this case we suppose
that the substrate molecule, M, is carried byK past D. If K is loaded with M1, an inter-action takes place at D, emptying K, regard-less of the speed. The rate-limiting step inthis scheme is the filling of K with another Mafter it has been emptied by D. The fillingprobability will be lower the shorter the timespent between D sites, that is, the higher thespeed. It turns out that if the law for fillingempty sites is exponential in time, which isnot unreasonable, the probability factor willhave exactly the same form as in the previouscase, in which "proximity time" rather than"filling timee" is rate limiting.10 This mneansthat there will be no difference in the kinetics
Circulation, Volume XXIV, August 1961
THICK FILAMENT
THIN FILAMENT
Figure 7Distribution of sites along the thick and thin fila-mnents. See text for explanation.
of the 2 schemes for steady motions; bothexplain equally well how motion controls therate of energy release. The different structureof the schemes will become important, how-ever, when we consider velocity and forcetransients.The question of whether the thin filaments
fold or slide during shortening has been side-stepped. This, could be done because insteady motions the mathematics proves to besubstantially the same for both cases.7
ForceWhat generates the contractile force? In
a sliding model, since the lengths of both the
403
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

PODOLSKY
PROBABILITY
1.0 r
/ENCOUNTER,' RATEl
V/VmoxFigure 8
Chemical kinetics for reaction sites moving withrelative velocity. V. Interrupted line: probabilityof interaction (rate limited by proximity time) orof filling (rate limited by filling time). Dottedline: encounter rate. Solid line: reaction rate =probability X encounter rate.
thick and thin filaments remain the same, thecontractile force must stem from mechanicalinteraction between the filaments. In a fold-ing model, however, the contractile force isgenerated in the thin filament and its lengthis supposed to decrease during shortening.A Sliding ModelThe mechanical and chemical properties of
a sliding mechanism in which the D site onthe thick filament has the mechanochemicalproperties diagrammed in figure 9 was workedout by Andrew Huxley.ll* D oscilates backand forth. When the K site passes, it caninteract with D to form a mechanical connec-tion. Since the probability of forming a con-nection in a D-K transit depends on therelative speed, this is a special case of inter-action in which "proximity time" is ratelimiting. Each connection is, in time, broken.One step of the driving chemical reaction isassociated with each connect-disconnect cycle.
If D and K connect, the elastic elementsholding D to the thick filament will exert a
*Figures 9 and 10 reproduced from A. F. Huxley:Progr. Biophysics 7: 255, 1957." By permission ofPergamon Press Inc.
force on the thin filament that is proportionalREACTIJN to the distance of D from 0. If the thin fila-4 RATE ment is moving to the left, connections with
D to the right of 0 will make a positive con-tribution to the force. Conversely, if motionof the thin filament carries D to the left of 0,a force will be developed that retards the
- motion.To develop a net positive force-that is,
to ensure that there will be a greater numberof pulling than retarding connections-Hux-ley postulates that D can connect to K onlywhen it is to the right of 0. The rate con-stant for breaking a connection also dependson x: it is small when D is to the right of 0but large when it is to the left.The force developed by the model depends
on speed because the number of attachmentsand their distribution about the equilibriumposition of the D site depends on speed. Thisis shown in figure 10, taken from Huxley'spaper.11 The ordinate is the fraction of D sitesat a given displacement from the equilibriumposition that are connected to K sites. Whenthe filaments do not move, all the connectionsare on the pulling side of the equilibrium posi-tion: links can be made only on this side andthere is no motion to carry them to the otherside. In steady motion, the number of pullinglinks decreases; the retarding links tend toincrease, and the force drops. At the maxi-mum speed, the pulling and retarding forcesbalance and there is no net force. Huxleyshowed that, as the relative speed increases,these shifts in both number and distributionof links between filaments can account quanti-tatively for the force-velocity relation in liv-ing muscle.The diagram also shows why there must be
steady motion for a less-than-maximum forceto remain constant. Consider the distribu-tion of connections when the speed say, is one-tenth of the maximum. If the motion shouldstop, after some time pulling links wouldform to the right of 0, retarding links wouldopen to the left of 0, and the net forcewould increase. The original force could bere-established by relaxing the pulling links
Circulation, Volume XXIV, August 1961
40ni4
l
12
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

MUSCLE PHYSIOLOGY AND CONTRACTION THEORIES
or stretching the retarding links, that is, bydisplacing the "connection contour" to theleft, which, of course, is what happens whenthe thin filament slides past the thick fila-ment. A steady force can be set up only whenthe motion in a given time interval just com-
pensates for the net increase in pulling linksformed in that same period.The diagram also explains what happens
if tetanized muscle is quickly released. Beforerelease, the links are all in the pulling posi-tion, as shown in the distribution at the topof the figure. If the muscle is moved so
quickly that the links do not change theirpoints of attachment, the tension will falllinearly as the links are carried past theequilibrium point and will vanish when thedistribution becomes symmetrical about theorigin. Thus the model perdicts that, if mus-
cle is moved very quickly, the tension willdrop to zero with a small displacement, whichis what actually happens.'2' 13 This is a simplemechanical process and is due to the releaseof strain in the relatively short pulling con-
nections between the thick and thin filaments.Notice that the distribution of links just
after a quick displacement will be reetangu-lar, while the corners are rounded in thesteady state. Since, for a given tension, themotion depends on the shape of the "con-nection contour," this implies that the velo-city just after a sudden drop in tension fromthe maximum to some intermediate value willnot be the same as that after the steady statehas been established. There should be a velo-city transient reflecting the transition ofconnection contour " from a rectangle to
the steady-state shape. We shall return tothis point later.
In summary, in the contraction model ana-
lyzed by Huxley, the 2 sets of myofilamentsslide past each other. The sliding motionarises from mechanical interactions betweencomplementary sites. The chemistry of thesystem reflects the mechanical motion, sinceeach mechanical interaction is coupled to a
chemical reaction. The model explains re-
markably well the structural and energetic
Circulation, Volume XXIV, August 1961
Myosinfilament
Actinfilament
Equilibrium position Eof K site -.4
Figure 9Me(hantochemtical element in sliding model of 4.F. Huxley. x is distance of thin (actin) filamentsite, K, from equilibrium position of thick (myo-sin) filament site, D. See text for further explana-tion. (From A. F. Huxley.11)
changes that take place during contraction.The least satisfactory element in it is therather special nature of the hypotheticalmechanochemical sidepieces on the thick fila-ments, the "pullers." However, "it is notdifficult for Nature to do things in ways whichseem unduly complicated to physiologists. "2A Folding Model
In a folding model the contractile forcearises from a change in state of the thin fila-mnents; the thin filaments become elastic, likea rubber band. To make such a model work,we must assume that force is generated whena substrate molecule, M, binds to (or reactswith) the thin filament at a K site and thatthe magnitude of the force is proportional tothe number of occupied K sites.14 The forcecould arise from an "electrostatic entropic"process, as has been eloquently argued byMorales and Botts,1 5 or from a " polymermelting" process, as suggested by Pryor16and by Flory.17
In an "electrostatic entropiic" process, arubber-like filament is stretched out by themutual repulsion of distributed, electricallycharged groups. At the equilibrium length.the entropic force tending to shorten the fila-ment is just balanced by the electrostaticforce tending to extend it. If some of thecharged groups were neutralized by the bind-ing of oppositely charged molecules, the netelectrostatic force would decrease and thefilament would tend to shorten; if length werekept constant, force would be developed.
In a "polymer melting" process, the fila-ment is initially extended by structural forces,
405
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

PODOLSKY
10
n0.08
06
04
02
-15 -10 -05 x/h 0~5
V. 0.8
06V= Van
-1-5 -10 - 0-5 0 x/h 05
Figure 10Distribution of links between thick and thin fila-ments in steady motion of sliding model of A. F.Huxley: n is fraction of D sites at a distance x
from the equilibrium position that are connected toK sites; V is relative velocity; h is maximum valueof x (see fig. 9). (From A. F. Huxley.11)
such as those that coordinate a crystallinesolid. When a critical temperature is reached,the crystalline structure "melts" and thepolymer becomes like rubber. In this state,if kept isometric, the filament will exert con-
siderable force. The dotted line in fig. 11shows such a phase transition for an idealcrystalline polymer. The melting temperaturewill be changed if substances bind to (orreact with) the polymer; the change will de-pend on the extent of the binding.Sharp phase transitions are characteristic
of the ideal crystalline polymer. Meltingcurves or real polymers are often more grad-ual (solid line, fig. 11) ;17 however, in thiscase too the curve can be shifted by an amountthat depends on the extent of binding. Now,if temperature is kept constant (T =) andthe extent of binding varied, the extent ofmelting will vary. If length is kept constant,the force will increase with binding.
Degree of melting in this process is analo-
gous to charge neutralization in the electro-statie-entropie process; both unlock the rub-ber-like properties of the polymer. Theessential features are that (a) force is gen-erated in a single filament which can shortenby folding and (b) force can change accord-ing to the number of small molecules boundto (or reacted with) the filament.A considerable amount of muscle physi-
ology follows naturally from this hypothesisif we suppose, further, that there are siteson the thick filament that remove the forcegenerating molecules from the thin filamentby interacting chemically with them as theymove by. In other words, we suppose thatthere are interactions between the 2 sets offilaments in which "filling time" is rate limit-ing. We have already shown that this schemeyields up the correct answers for the relationbetween rate of energy production andspeed.*To explain how the force-velocity curve
comes out, consider figure 7 again. Supposethe force is a third of the maximum. Then1 binding (K) site out of 3 will be filled; 2out of 3 will be empty. A substrate moleculefrom the environment will, in time, find itsway to one of the empty sites. When the sitefills, the force in the filament will rise abovethat of the load. To re-establish mechanicalequilibrium, the filament will shorten by fold-ing until 1 of the filled sites passes a D site,which removes an M so that the force willagain be a third of the maximum. The processwill be repeated when another binding site isfilled. In the steady state, with many sites inthe game, the rate of motion will be such thatthe emptying of full K sites passing D sitesis just balanced by filling of empty K sites.The force is the average occupancy of the K
*The energeties of steady motion does not dependon whether "'filling time'" or "'proximity time'" israte limiting. However, in the latter ease (as wasused in an earlier version of the folding model"1),it can be showii that the model does not accommodatethe drop in force after quick release from tetanictension. On the other hand, if "'filling time " israte limiting, the folding model behaves "properly"upon quick release.
Circulation, Volume XXIV, August 1961
406
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

MUSCLE PHYSIOLOGY AND CONTRACTION THEORIES
sites; the dependence of the average occu-pancy on velocity is the force-velocity relation.This turns out to be the same as that in livingmuscle.What happens when a muscle exerting
maximum tension is suddenly released? (Youwill recall that in the sliding model, becausethe " pullers" relaxed, the tension droppedlinearly with shortening.) In this case, ifthe motion is quick relative to the filling time,full K sites will be rapidly emptied by re-acting chemically with passing D sites, andthe tension will drop. The tension will falllinearly with distance only until the forcereaches half maximum. Then, because short-ening is by folding rather than by sliding,it can be shown that further motion will leadto a proportionately smaller drop in force,which is what happens in living muscle.13As in the analogous situation in the sliding
model, just after a quick drop in force, thedistribution of filled sites along the filamentis not the same as it will be somewhat later,after the steady state is established. Thismeans that the isotonic velocity after quickrelease from tetanic tension will generallybe different from the later steady velocity.Some time must pass before the velocity set-tles down to the characteristic steady value.
Mechanical TransientsWe made a series of experiments to look
for this transition phase.18 The study wasmade with tetanically stimulated frog sarto-rius at 0 C, (fig. 12).* The upper trace isdisplacement of one end of the muscle and thelower trace is force at the other end. Afterfull tension was developed, the force on themusele was suddenly lowered to, and thenmaintained at, a given value. Each set oftraces is for a different final value of theforce. The insert (fig. 12g) is a control withthe muscle replaced by a simple spring.
Three regions are of interest. To the leftof the vertical line we see the quick releasephenomenon. The muscle is 35 mm. long;the tension drops to nearly zero when the end
*Figure 12 reproduced from Podolsky: Nature188: 666, 1960.18 By permission of Nature.
Circulation, Volume XXIV, August 1961
TENSION Constant length C2 >C1>0
r TEMPERATURE
Figure 11Tension development according to "polymer-melt-ing" process. Interrupted lines: melting curves forideal crystalline polymers; solid lines: melting(crves for real polymers. The melting tempera-ture decreases with the extent of binding (or re-action) of substrate with the polymer filament.
moves less than 2 per cent of this length (fig.12f). Depending on which model we favor,this could be either relaxation of pulling linksor emptying of binding sites. Hill found aburst of heat during the quick release, whichhe attributed to a high thermoelastic coeffi-cient of what in the sliding model correspondsto the pulling spring.'9 An alternative inter-pretation is that a chemical rather than aphysical process is associated with the loss oftension, as is the' case in the folding model.This interpretation also agrees with the studiesof the insect physiologists, who invoke "in-activation by release"-as opposed to "relaxa-tion by release "-to explain the very highfrequency movements of certain insect mus-cles, such as those driving the noisemaker ofthe locust.20At each force, the velocity ultimately set-
tles down to the characteristic steady value:the lower the force, the more rapid the motion.(The force-velocity curve in figure 1 wasdrawn from these data.) The remarkablelinearity of the displacement traces supportsthe idea that the motion is controlled by afeedback mechanism.
The interesting findings in these experi-ments are the variations ill speed (to theright of the vertical line) before the velocitysettles down to the steady value; there is a
1
407
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

PODOLSKY
mmn0.203
30
20gm
I0
7
10 msec
a
d
-7
b
e
X 11
\11-At
f r.
Figure 12Response of muscle to a sud-den change in force. Uppertrace: displacement; lowertrace: tension; frog sartorius,standard length = 35 mm.,0 C. Muscle is initially at thestandard length and the rec-ord is started after full tetanicforce, Pmax., has developed.P/Pmax: (a) 0.84 (b) 0.69(c) 0.55 (d) 0.40 (e) 0.26(f) 0.10. In (g) the muscleis replaced by a simple spring.(From Podolsky.158)
characteristic isotonic velocity transient foreach tension step. One contribution to thesetransients is the variation in velocity corre-sponding to the setting up of the steady stateof motion in the contraction models we havediscussed. However, since the experimentswere made with the whole sartorius muscle,there is also a contribution due to the inter-action of muscle fibers of different lengths.To sort out these 2 components of the tran-sient seen in the intact muscle, we are repeat-ing these experiments with preparationscontaining only a few fibers; this should re-duce the contribution of fiber interaction.We are also calculating the transients pre-dicted by the sliding and folding contractionmodels, to see which model accommodates theexperimental data better.* In these studiesattention is focused on the approach to thesteady state of motion rather than on thesteady state per se; then the contraction kinet-ics of the 2 models can be distinguished.
ConclusionIn summing up, I should like to point out
that the contraction theories work because of2 basic assumptions. The first-and this canreally be elevated to the status of fact ratherthan assumption-is that the 2 sets of myo-
*This study is being made in collaboration withDr. N. Z. Shapiro of the National Institutes ofHealth.
filaments move relative to each other in short-ening. The second is that the flux of chemicalenergy through the contractile mechanism islimited by interactions between complemen-tary sites distributed along the 2 sets offilaments.Two models were fitted to these ideas. They
differ largely in the mechanism by which forceis generated. In the sliding model, force isdeveloped by mechanical interaction betweenfilaments and there is no change in filamentlength during shortening. In the foldingmodel, force is developed in a single filament,which shortens during contraction. In bothmodels there is interaction between the me-chanical motion and the force-generatingmechanism. Chemical processes tend to in-crease the mechanical force: in the slidingmodel, pulling connections are made; in thefolding model, binding sites, which generateforce if filled, become filled. If the load isconstant, these processes tend to create a me-chanical imbalance which, however, can berighted by shortening: in the sliding model,pulling connections become weaker; in thefolding model, binding sites are emptied.Chemical and mechanical equilibrium areincompatible for forces less than full tension.However, since shortening tends to inactivatethe force generator, a less-than-maximumforce can be maintained if there is steadymotion. Conversely, if the load is less than
Circulation, Volume XXIV, August 1961
408
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

MUSCLE PHYSIOLOGY AND CONTRACTION THEORIES
the maximum force that can be generated,there will be steady shortening. This is theforce-velocity relation.Comparing the models with living muscle,
both of them quantitatively account for thechanges in force and energy flux with velo-city. The quick-release phenomenon is alsoaccommodated: mechanically in the slidingmodel and chemically in the folding model.Both predict, at least qualitatively, the iso-tonic velocity transients that can be seen afterquick release from tetanic tension.
This list of accomplishments suggests thatsome of the devices used to get them mightactually be built into living muscle.
References1. FENN, W. 0.: A quantitative comparison between
the energy liberated and the work performedby the isolated sartorius muscle of the frog.J. Physiol. 58: 175, 1923.
2. FENN, W. 0.: Contractility. In Hbber, R.: Physi-cal Chemistry of Cells and Tissues. Philadel-phia, The Blakiston Co., 1945, pp. 459, 502.
3. HILL, A. V.: The heat of shortening and thedynamic constants of muscle. Proc. Roy. Soc.London s.B 126: 136, 1938.
4. HILL, A. V.: The maximum work and mechanicalefficiency of human muscles, and their mosteconomical speed. J. Physiol. 56: 19, 1922.
5. HEIDENHAIN, R.: Mechanische Leistung, Warm-entwickelung, und Stoffumsatz bei der Mus-keltiatigkeit. Leipzig, Breitkopf u. Hirtel,1864.
6. HILL, A. V.: Recovery heat in muscle. Proc.Roy. Soc. London s.B 127: 297, 1939.
7. PODOLSKY, R. J.: Thermodynamics of muscle.In Structure and Function of Muscle. Edited
by G. H. Bourne. New York, Academic PressInc., 1960, vol. 2, p. 375.
8. HUXLEY, H. E.: The double array of filamentsin cross-striated muscle. J. Biophys. & Biochem.Cytol. 3: 631, 1957.
9. HUXLEY, A. F., AND NIEDERGERKE, R.: Structuralchanges in muscle during contraction. Nature173: 971, 1954.
10. PODOLSKY, R. J.: The nature of the contractilemechanism in muscle. In Biophysics of Physi-ological and Pharmacological Actions. Editedby A. M. Shanes. Washington, D. C., AmericaiAssociation for the Advancement of Science.In press.
11. HUXLEY, A. F.: Muscle structure and theoriesof contraction. Prog. Biophysics 7: 255, 1957.
12. GASSER, H. S., AND HILL, A. V.: The dynamicsof muscular contraction. Proe. Roy. Soc.London s.B 96: 398, 1924.
13. HILL, A. V.: The series elastic component ofmuscle. Proe. Roy. Soc. London s.B 137: 273,1950.
14. PODOLSKY, R. J.: The chemical thermodynamicsand molecular mechanism of muscular contrac-tion. Ann. Newl York Aead. Se. 72: 522, 1959.
15. MORALES, M. F., AND BOTTS, J.: A model forthe elementary process in muscle action. Arch.Biochem. 37: 283, 1952.
16. PRYOR, M. G. M.: Mechanical properties offibers and muscles. Progr. Biophysics 1: 216,1950.
17. FLORY, P. J.: Role of crystallization in polymersand proteins. Science 124: 53, 1956.
18. PODOLSKY, R. J.: The kinetics of muscularcontraction; the approach to the steady state.Nature 188: 666, 1960.
19. HILL, A. V.: The 'instantaneous' elasticity ofactive muscle. Proc. Roy. Soc. London s.B141: 161, 1953.
20. PRINGLE, J. W. S.: The mechanism of the myo-genie rhythm of certain insect striated muscle.J. Physiol. 124: 269, 1954.
Circulation, Volume XXIV, August 1961
409
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from

John V. Taggart and RICHARD J. PODOLSKYMuscle Physiology and Contraction Theories
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 1961 American Heart Association, Inc. All rights reserved.
75231is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TXCirculation
doi: 10.1161/01.CIR.24.2.3991961;24:399-409Circulation.
http://circ.ahajournals.org/content/24/2/399located on the World Wide Web at:
The online version of this article, along with updated information and services, is
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer
of the Web page under Services. Further information about this process is available in thewhich permission is being requested is located, click Request Permissions in the middle columnClearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of the CopyrightCirculationoriginally published in Requests for permissions to reproduce figures, tables, or portions of articlesPermissions:
by guest on May 23, 2018
http://circ.ahajournals.org/D
ownloaded from