Embed Size (px)
Citation preview

Brain Research 863 (2000) 52–58www.elsevier.com/ locate /bres
Research report
Morphine-induced suppression of saccharin intake is correlated withelevated corticosterone levels
*Felipe Gomez , Nicole A. Leo, Patricia S. GrigsonDepartment of Behavioral Science, Penn State College of Medicine, Hershey, PA 17033, USA
Accepted 1 February 2000
Abstract
Rats suppress intake of a saccharin conditioned stimulus (CS) when paired with a drug of abuse. This phenomenon, however, is notuniform across all subjects and is greater following exposure to stress and in animals that more readily self-administer drugs of abuse. Thepresent study was designed to examine these individual differences in intake suppression following seven saccharin–morphine pairings.Plasma corticosterone also was evaluated both before and after conditioning in order to determine whether the magnitude of CSsuppression is, or is not, related to circulating corticosterone levels. The findings indicated that, while all rats were exposed to the samenumber of saccharin–morphine pairings, only half of these animals actually suppressed intake of the saccharin CS. Moreover, the resultsshowed that greater suppression of CS intake was associated with higher corticosterone levels at test (r520.84, P,0.0001). Takentogether, the results demonstrate that individual differences affect not only the reduction in CS intake following taste-drug pairings, butalso the associated cue-induced elevation in circulating corticosterone. 2000 Elsevier Science B.V. All rights reserved.
Themes: Neural basis of behavior
Topics: Drugs of abuse: opioids
Keywords: CTA; Conditioned taste aversion; Morphine; Corticosterone; Saccharin; Anticipatory contrast; Individual differences
1. Introduction hypothesis is in keeping with a similar behavioral phenom-enon, referred to as anticipatory contrast, where rats
Intake of a gustatory conditioned stimulus (CS) is suppress intake of the very same saccharin CS as it comessuppressed when paired with the administration of a drug to predict the availability of a highly preferred sucroseof abuse such as morphine, cocaine, amphetamine, or unconditioned stimulus (US) following daily saccharin–heroin [6,7,11,24,31]. Despite the well known reinforcing sucrose pairings [12,13].properties of such drugs, their suppressive effects have Behavioral and physiological evidence supports thisbeen interpreted as conditioned taste aversions (CTAs) for reward comparison hypothesis. Indeed, when evaluatedover 25 years ([35], for review see [19] and [25]). More across a number of tasks, the suppressive effects of drugsrecently, however, it has been suggested that the reinforc- of abuse do not resemble those of the aversive agent, LiCl,ing rather than the aversive properties of the drugs of but are more like those induced by a rewarding sucroseabuse may be responsible for suppressing intake of the solution. First, rats suppress intake of either a saccharin orgustatory CS following taste-drug pairings [20]. Specifical- a salt CS when paired with LiCl-induced illness, but onlyly, Grigson proposed that rats decrease intake of saccharin of a saccharin CS when paired with a standard dose offollowing saccharin–morphine pairings, for example, be- morphine or cocaine [4,20]. Second, the suppressivecause the value of the gustatory CS pales in comparison to effects of morphine, but not LiCl, are prevented whenthe anticipated, highly rewarding, drug of abuse. This using a sucrose CS and water-deprived rats [23]. Similarly,
the use of a caloric sucrose CS in food-deprived rats ismore disruptive to the suppressive effects of sucrose,*Corresponding author. Tel.: 11-717-531-7676; fax: 11-717-531-morphine, and cocaine, than it is to the suppressive effects6916.
E-mail address: [email protected] (F. Gomez) of LiCl [14,18]. Third, Lewis rats, which show a greater
0006-8993/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0006-8993( 00 )02093-X

F. Gomez et al. / Brain Research 863 (2000) 52 –58 53
preference for drugs of abuse than Fischer 344 rats, also individual stainless steel cages in a colony room in whichevidence greater cocaine- and sucrose-induced suppression the temperature and humidity were automatically reg-of saccharin intake, while LiCl-induced CTAs are equiva- ulated. Lighting was maintained on a 12/12 h light /darklent between the two strains ([17,21,29], but see [28]). cycle and all manipulations where conducted 3 h into theFinally, bilateral lesions of the gustatory thalamus prevent light phase of the cycle. The rats received free access tosucrose-, and morphine-, but not LiCl-induced suppression dry Harlan Teklad rodent diet and water, except whereof CS intake in rats [22,39,41]. noted otherwise.
Although progress is being made in our conceptualiza-tion of this phenomenon, little is known about the neural 2.2. Apparatusmechanisms mediating drug-induced suppression of CSintake. A link can be established, however, between drug- The experiment was conducted in the home cages usinginduced suppression and physiological factors that predict inverted Nalgene graduated cylinders with silicone stop-the propensity to self-administer drugs of abuse. First, pers and stainless steel spouts affixed to the front of thethere is evidence that the activity of the hypothalamic cage. Total intake was recorded to the nearest 0.5 ml.pituitary adrenocortical (HPA) axis may enhance both theacquisition of drug self-administration and drug-induced 2.3. Drugs and solutionssuppression of CS intake. For example, rats that react mostto novelty stress show greater corticosterone (CORT) Morphine was prepared prior to testing by disolvingrelease and faster acquisition of drug self-administration morphine sulfate in physiological saline at a concentration[37]. Furthermore, treatment with corticosterone (CORT) of 15 mg/ml and injected in a dose of 15 mg/kg. The drugcan elicit drug self-administration in rats that otherwise are was provided by the National Institute on Drug Abuse.not prone to the behavior [32,37] and, as with drug Saccharin was obtained from the Sigma Chemical Com-self-administration, drug-induced suppression of CS intake pany, St. Louis, MO and was presented at room tempera-also is exaggerated by exposure to stress in out bred rats ture.[5,30]. Second, as stated, the Lewis rat, is predisposed toboth drug self-administration and to drug-induced suppres- 2.4. Proceduresion of CS intake [2,17,21,29]. Finally, individual differ-ences in drug-induced suppression of CS intake are evident Deprivation state: The rats were placed on a water-in non-stressed Sprague–Dawley rats [10,40,43] and data deprivation schedule for 10 days prior to the start of thefrom Wise et al. [45] indicate that rats showing the greatest experiment in which they received access to distilled waterdrug-induced suppression of CS intake reportedly self- (dH O) for 5 min in the morning and for 1 h in the2
administer the most of the drug of abuse when given the afternoon. Baseline corticosterone: Once intake stabilizedopportunity to do so [45]. within a range between 7 and 13 ml /5 min, all rats
Taken together, the data indicate that rats will suppress received 5 min access to dH O, and after a 15 min2
intake of a saccharin CS following taste-drug pairings, that interval, blood samples (approximately 0.15 ml) werethe magnitude of suppression differs across animals, and collected by tail cut (between 700 h and 800 h) and storedthat the degree of suppression is exaggerated by exposure for hormone analysis. Conditioning: Two days later, allto stress and by a propensity to self-administer drugs of rats were given 5 min access to a 0.15% saccharin CS, andabuse. Although evidence suggests that exposure to stress after a 5 min interval, were injected intraperitoneally (ip)can augment drug-induced suppression of CS intake [5,30], with either saline (n54) or 15 mg/kg morphine sulfateit is not yet known whether the individual differences in (n512). There was one taste-drug pairing a day (betweenCS intake in out bred Sprague–Dawley rats are directly 800 h and 900 h), occurring every other day, for a total ofrelated to individual differences in the activity of the HPA seven trials. All subjects received 5 min access to dH O2
axis. Thus, the present study is designed to determine the each morning between conditioning trials and 1 h accessdegree to which individual differences in CS intake relate every afternoon to rehydrate. Post-conditioning corticos-to circulating CORT either before (i.e., during baseline), or terone: A second blood sample (approximately 0.15 ml)after, taste-drug conditioning. was collected from the tail 15 min after the seventh CS
access period (between 700 h and 800 h). The US (i.e.,saline or morphine) was not administered on this final test
2. Methods day and corticosterone levels were measured as describedbelow.
2.1. Subjects2.5. Hormone assay
The subjects were 16 male Sprague–Dawley rats(Charles River, Wilmington, MA) weighing 250–400 g at Blood samples were maintained on ice and then cen-the beginning of the experiment. They were housed in trifuged at 3000 rpm for 10 min. The serum collected from
54 F. Gomez et al. / Brain Research 863 (2000) 52 –58
each was equiliquoted and stored at 2808C until the assay into two distinct populations, see Fig. 1, right panel. Halfwas performed. Corticosterone concentrations were de- of the rats strongly suppressed intake of the gustatory CStermined using a standard radioimmunoassay kit (ICN (i.e., they consumed less than 5 ml /5 min) following sevenBiomedicals). The samples were run in duplicate, while the taste-drug pairings, while the others showed almost no signstandards were run in triplicate. of suppression (i.e., they consumed more than 5 ml /5 min
throughout training). As a consequence, the data werereanalyzed with group (saline, morphine small suppressers,
3. Results and morphine large suppressers) as a factor in the analysis.The results of this 337 ANOVA revealed that, under these
3.1. CS Intake circumstances, the main effect of group was highly signifi-cant, F(2,13)522.82, P,0.0001, as was the Group3Trials
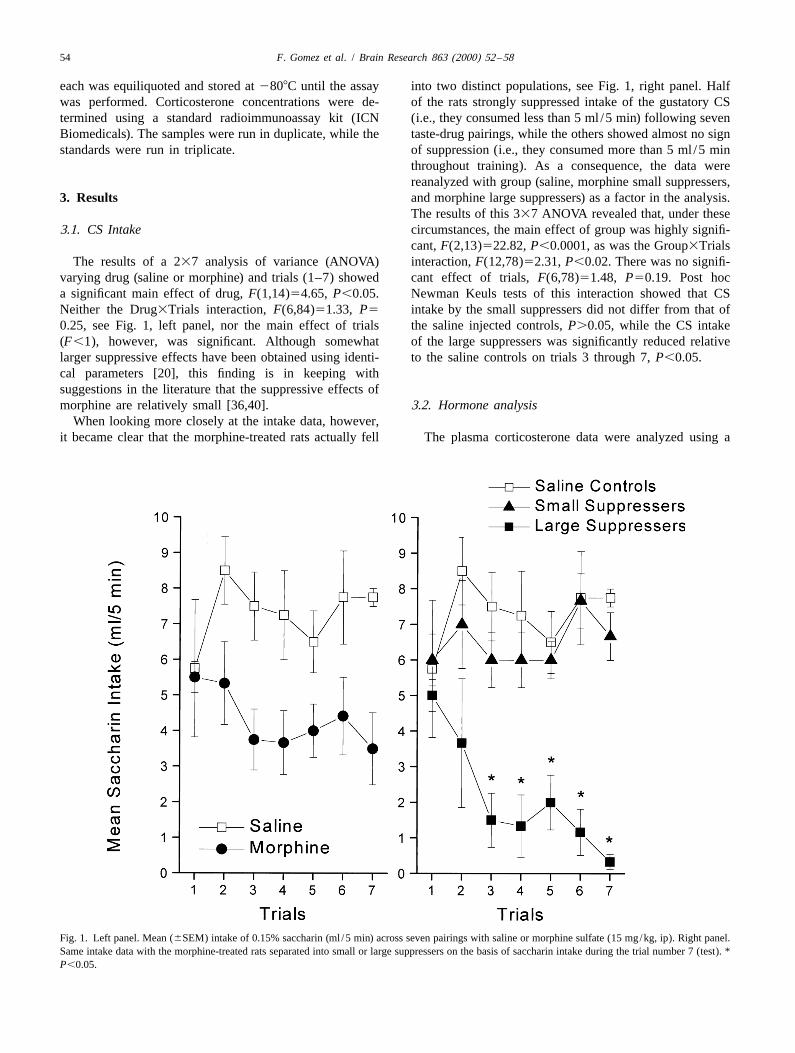
The results of a 237 analysis of variance (ANOVA) interaction, F(12,78)52.31, P,0.02. There was no signifi-varying drug (saline or morphine) and trials (1–7) showed cant effect of trials, F(6,78)51.48, P50.19. Post hoca significant main effect of drug, F(1,14)54.65, P,0.05. Newman Keuls tests of this interaction showed that CSNeither the Drug3Trials interaction, F(6,84)51.33, P5 intake by the small suppressers did not differ from that of0.25, see Fig. 1, left panel, nor the main effect of trials the saline injected controls, P.0.05, while the CS intake(F,1), however, was significant. Although somewhat of the large suppressers was significantly reduced relativelarger suppressive effects have been obtained using identi- to the saline controls on trials 3 through 7, P,0.05.cal parameters [20], this finding is in keeping withsuggestions in the literature that the suppressive effects ofmorphine are relatively small [36,40]. 3.2. Hormone analysis
When looking more closely at the intake data, however,it became clear that the morphine-treated rats actually fell The plasma corticosterone data were analyzed using a
Fig. 1. Left panel. Mean (6SEM) intake of 0.15% saccharin (ml /5 min) across seven pairings with saline or morphine sulfate (15 mg/kg, ip). Right panel.Same intake data with the morphine-treated rats separated into small or large suppressers on the basis of saccharin intake during the trial number 7 (test). *P,0.05.

F. Gomez et al. / Brain Research 863 (2000) 52 –58 55
332 repeated measures ANOVA varying group (saline, relationship between CS intake at test and preconditioningmorphine small suppressers, and morphine large suppres- (i.e., baseline) CORT levels, P.0.05.sers) and trials (pre and post conditioning). The results ofthis analysis revealed a significant main effect of group,F(2,13)57.76, P,0.006, and trials, F (1,13)540.6, P,
0.0001, and a significant Group3Trials interaction, 4. DiscussionF(2,13)59.95, P,0.002, see Fig. 2, left panel.
Post hoc Newman Keuls tests of this interaction indi- Large individual differences in CS intake have beencated that baseline CORT levels did not differ significantly observed following saccharin–cocaine, saccharin–heroinacross the three groups, P.0.05. Levels of CORT did, and, as stated, saccharin–morphine pairings [10,24,40].however, change as a function of training. Specifically, Indeed, Riley et al. (1978) reported low, intermediate, orCORT levels were significantly elevated from pre- to high CS intake when saccharin was paired with a potentpost-training for both the small, P,0.05, and the large, dose of morphine [40]. The results of the present reportP,0.0001, suppressers, but not for the saline injected both support and extend these findings. Specifically, whilecontrols. Furthermore, during post-training, CORT levels rats in the saccharin–morphine group suppressed CS intakewere greater for the small suppressers than for the saline overall, there were clear individual differences. Whencontrols and greater for the large suppressers than for the separated on the median score, two distinct populationssmall suppressers (P,0.05). Finally the results of a emerged such that half of the animals strongly avoided thecorrelational analysis revealed a strong negative relation- saccharin CS following saccharin–morphine pairings,ship between CS intake and plasma corticosterone at test, while the other half did not. Moreover, by combining ther520.84, P,0.0001, see Fig. 2, right panel. That is, at intake data with the CORT data it became evident thattest, greater suppression of CS intake was significantly although the individual differences in CS intake were notcorrelated with higher levels of circulating CORT. A related to baseline CORT levels, they were highly corre-similar correlational analysis failed to reveal a significant lated with post-training CORT levels such that greater
Fig. 2. Left panel. Mean (6SEM) plasma corticosterone level (ng /ml) for the three groups of animals (saline, small suppressers, and large suppressers) 15min after the access period to water (pre-training) or 15 min after the 5 min access period to saccharin at test (post-training). Right panel. Intake of thesaccharin conditioned stimulus as a function of corticosterone levels at test for the three groups.

56 F. Gomez et al. / Brain Research 863 (2000) 52 –58
suppression of CS intake was associated with higher levels low fluid intake at test. While CORT levels can be elevatedof circulating CORT at test. To the authors’ knowledge, by water restriction [1], the data argue against thisthese findings represent the first assessment of CORT interpretation. First, CORT levels also were elevated in thefollowing pairings of a gustatory CS with a drug of abuse. small suppressers which consumed an equal volume of the
Interpretation of the data requires that we consider why saccharin CS as the saline injected controls. Second, werats suppress intake of a saccharin CS following sac- have recently obtained similar elevations in the largecharin–morphine pairings, why circulating CORT levels suppressers, when using a two-bottle test (i.e., when bothare elevated at test, and whether these two indices reflect a the saccharin CS and water were available).similar process. As discussed in the Introduction, there is If the changes in CORT levels are not due to un-some controversy about why rats suppress intake of a conditioned effects, then the possibility must be consideredsaccharin CS when it predicts the availability of a drug of that the elevation reflects a conditioned or learned effect.abuse ([20]; see [19] and [25] for a review). Evidence is This, in fact, is the most likely alternative. It is well knownaccumulating, however, to suggest that drugs of abuse that cues are readily associated with drugs of abuse viasuppress CS intake via their reinforcing, rather than their classical conditioning and that these cues can elicit cravingaversive, US properties. For example, Parker [36] has in drug experienced humans [9,16]. Using a runway, itshown that unlike LiCl-induced CTAs, the suppressive already has been demonstrated that rats can use aneffects of drugs of abuse are not accompanied by an olfactory discriminative stimulus to predict the availabilityincrease in aversive orofacial behaviors such as gapes and of heroin [33] or contextual cues to predict the availabilitychin rubs [36]. In what had previously been considered the of cocaine [15,27]. Furthermore it is known that presenta-most paradoxical of findings, it has been determined that tion of the contextual cues can elicit the conditionedthe decrease in CS intake following taste-drug pairings is release of dopamine in the nucleus accumbens [15,27] orparalleled with a concomitant increase in preference for the the conditioned release of CORT induced by context cueslocation paired with the drug, an increase in running speed associated with cocaine administration [8]. Finally, notin a runway, and an increase in drug self-administration only contextual cues, but also a saccharin cue, will support[38,44,45]. Moreover, Wise et al. found that the rats that a conditioned analgesic response following saccharin–mor-exhibited the greatest reduction in CS intake also exhibited phine pairings [3] or the conditioned release of CORTthe most avid drug self-administration behavior. These following saccharin–LiCl [1].data add to those described in the Introduction to suggest Taken together, these data suggest that the gustatory cuethat drugs of abuse are highly rewarding. As such, the data comes to predict the availability of the drug of abuse viasupport the suggestion that rats suppress intake of a classical conditioning. Indeed, evidence for this conclusionsaccharin CS following taste-drug pairings because the was provided some time ago when Sherman et al. [42]value of the saccharin cue pales in anticipation of the showed that the reduction in CS intake does not occuravailability of the highly rewarding drug of abuse much as when the morphine US is presented before the gustatoryit does when predicting the availability of a highly CS [42]. The present data also suggest that multiple taste-preferred sucrose solution [12,13,20]. drug pairings lead not only to a conditioned suppression of
Given the paucity of data, it is not immediately clear CS intake, but to a conditioned elevation of CORT as well.why circulating CORT levels are elevated at test and why At this juncture it is not possible to determine whether thethe levels differ between the small and the large suppres- elevation in CORT reflects cue-induced anticipation ofsers. A number of explanations can be considered involv- drug administration, ’craving’, or even withdrawal. What ising either unconditioned or conditioned effects of the drug. markedly clear, however, is that some animals are veryRegarding the possible unconditioned effects, the first sensitive to this manipulation while others are not. Somepossibility is that the elevation of CORT reflects general animals are more likely to avoid the natural saccharindifferences in basal CORT levels. This, however, clearly is reward and to exhibit a greater cue-induced release ofnot the case because baseline CORT levels were not CORT following taste-drug pairings. According to thesignificantly different across the three groups (saline, reward comparison hypothesis, these rats avoid the sac-small, and large suppressers) and baseline CORT was not charin CS because they are anticipating the availability ofcorrelated with the magnitude of drug-induced suppression the preferred drug of abuse. Consistent with this interpreta-of CS intake at test. A second possibility is that the tion, Wise at al. [45] showed that those animals that avoidelevation in CORT at test occurred simply because of the saccharin CS are, in fact, more likely to self-administerrepeated exposure to the drug of abuse. Chronic morphine the drug of abuse [45]. These facts reflect what we suspecttreatment, however, has been associated with a decrease, is a lethal combination that may contribute to the intract-rather than an increase, in circulating CORT [26,34]. able nature of drug addiction as the predisposed individualFurther, CORT levels differed between the small and the is attracted by a strong desire for the drug of abuse,large suppressers, despite the fact that all animals had releases CORT which can, itself, facilitate drug self-ad-identical experience with the drug of abuse. A final ministration behavior, and remains unmotivated by naturalconsideration is that the elevation in CORT resulted from rewards which are, by comparison, deemed unsatisfactory.

F. Gomez et al. / Brain Research 863 (2000) 52 –58 57
[20] P.S. Grigson, Conditioned taste aversion and drugs of abuse: AAcknowledgementsreinterpretation, Behav. Neurosci. 1 (1997) 129–136.
[21] P.S. Grigson, C.S. Freet, The suppressive effects of sucrose andThis research was supported by the U.S. Public Health cocaine, but not lithium chloride, are greater in Lewis than Fisher
Service Grants DA 12933, DA 09815 and DC 02016. We rats: Evidence for the reward comparison hypothesis, Behavioralthank the National Institute on Drug Abuse for generously Neuroscience, in press.
[22] P.S. Grigson, P. Lyuboslavsky, Tanase, Bilateral lesions of theproviding the morphine sulfate.gustatory thalamus disrupt morphine, but not LiCl-induced con-ditioned taste aversions in rats: Evidence for the reward comparisonhypothesis, Brain Res., in press.
References [23] P.S. Grigson, P. Lyuboslavsky, D. Tanase, R.A. Wheeler, Waterdeprivation prevents morphine, but not LiCl-induced, suppression ofsucrose intake, Physiol. Behav. 67 (1999) 277–286.[1] R. Ader, Conditioned adrenocortical steroid elevations in the rat, J.
[24] P.S. Grigson, R.C. Twining, R.M. Carelli, Heroin-induced suppres-Comp. Physiol. Psychol. 90 (1976) 1156–1163.sion of saccharin intake in water-deprived and free-feeding rats,[2] E. Ambrosio, S. Goldberg, G. Elmer, Behavior genetic investigationPharmacol. Biochem. Behav., in press.of the relationship between spontaneous locomotor activity and the
[25] T. Hunt, Z. Amit, Conditioned taste aversion induced by self-acquisition of morphine self-administration behavior, Behavioraladministered drugs: Paradox revisited, Neurosci. Biobehav. Rev. 11Pharmacol. 6 (1995) 229–237.(1987) 107–130.[3] M.T. Bardo, J.M. Valone, Morphine-conditioned analgesia using a
[26] D.M. Ignar, C.M. Kuhn, Effects of specific mu and kappa opiatetaste cue: dissociation of taste aversion and analgesia, Psycho-tolerance and abstinence on hypothalamo–pituitary–adrenal axispharmacology 114 (1994) 269–744.secretion in the rat, J. Pharmacol. Exp. Ther. 255 (1990) 1287–[4] R.A. Bevins, T.A. Delzer, M.T. Bardo, Second order conditioning1295.detects unexpressed morphine-induced salt aversion, Anim. Learn.
[27] E.A. Kiyatkin, E.A. Stein, Conditioned changes in nucleus accum-Behav. 24 (1996) 221–229.bens dopamine signal established by intravenous cocaine in rats,[5] W.J. Bowers, M.A. Gingras, Z. Amit, Time-dependent exacerbationNeurosci. Lett. 211 (1996) 73–76.of amphetamine-induced taste aversion following exposure to
footshock, Psychopharmacology 125 (1996) 43–49. [28] T.A. Kosten, M.J.D. Miserendino, S. Chi, E.J. Nestler, Fisher and[6] H. Cappell, A.E. LeBlanc, Conditioned aversion to saccharin by Lewis rat strains show differential cocaine effects in conditioned
single administrations of mescaline and d-amphetamine, Psycho- place preference and behavioral sensitization but not locomotor orpharmacologia 22 (1971) 352–356. conditioned taste aversion, J. Pharmacol. Exp. Ther. 269 (1994)
[7] H. Cappell, A.E. LeBlanc, L. Endrenyi, Aversive conditioning by 137–144.psychoactive drugs: Effect of morphine, alcohol and chloro- [29] T.A. Kosten, M.J.D. Miserendino, C.N. Haile, J.L. DeCaprio, P.L.diazepoxide, Psychopharmacology 29 (1973) 239–246. Jatlow, E.J. Nestler, Acquisition and maintenance of intravenous
[8] A.C. De Vries, S.E. Taymans, J.M. Sundstrom, A. Pert, Conditioned cocaine self-administration in Lewis and Fischer inbred rat strains,release of corticosterone by contextual stimuli associated with Brain Res. 778 (1997) 418–429.cocaine is mediated by corticotropin-releasing factor, Brain Res. 786 [30] P.S. Lasiter, J.J. Braun, Shock facilitation of taste aversion learning,(1998) 39–46. Behav. Neural Biol. 32 (1981) 277–281.
[9] R.N. Ehrman, S.J. Robbins, A.R. Childress, C.P. O’Brian, Con- [31] J. Le Magnen, Peripheral and systemic actions of food in the caloricditioned responses to cocaine-related stimuli in cocaine abuse regulation of intake, Ann. NY Acad. Sci. 157 (1969) 1126–1157.patients, Psychopharmacology 107 (1992) 523–529. [32] J.R. Manstsh, D. Saphier, N.E. Goeders, Corticosterone facilitates
[10] P.D. Farber, J.E. Gorman, L.D. Reid, Morphine injections in the the acquisition of self-administration in rats: Opposite effect of thetaste aversion paradigm, Psysiol. Psych. 4 (1976) 365–368. type II glucocorticoid receptor agonist dexamethasone, J. Phar-
[11] C.M. Ferrari, B.F. O’Connor, A.L. Riley, Cocaine-induced taste macol. Exp. Ther. 287 (1998) 72–80.aversion: Effects of route of administration, Pharmacol. Biochem. [33] K. McFarland, A. Ettenberg, Reinstatement of drug-seeking be-Behav. 38 (1991) 267–271. havior produced by heroin-predictive environmental stimuli, Psy-
[12] C.F. Flaherty, S. Checke, Anticipation of incentive gain, Anim. chopharmacology 131 (1997) 86–92.Lern. Behav. 10 (1982) 177–182. [34] M.V. Milanes, M.L. Laorden, M. Chapleur-Chateau, A. Burlet,
[13] C.F. Flaherty, P.S. Grigson, From contrast to reinforcement: Role of Differential regulation of corticotropin-releasing factor and vas-response contingency in anticipatory contrast, J. Exp. Psychol. opressin in discrete brain regions after morphine administration:Anim. Behav. Proc. 14 (1988) 165–176. correlations with hypothalamic noradrenergic activity and pituitary-
[14] C.F. Flaherty, P.S. Grigson, S. Checke, K.C. Hant, Deprivation state adrenal response, Naunyn Schmiedebergs Arch. Pharmacol. 356and temporal horizons in anticipatory contrast, J. Exp. Psychol. (1997) 603–610.Anim. Behav. Proc. 14 (1991) 503–518. [35] M. Nachman, D. Lester, J. Le Magnen, Alcohol aversion in rats:
[15] D.J. Fontana, R.M. Post, A. Pert, Conditioned increases in mesolim- Behavioral assessment of noxious drugs effects, Science 168 (1970)bic dopamine overflow by stimuli associated with cocaine, Brain 1244–1246.Res. 629 (1993) 31–39. [36] L.A. Parker, Taste reactivity responses elicited by reinforcing drugs:
[16] I.H. Franken, V.M. Hendriks, Predicting outcome of inpatient A dose-response analysis, Behav. Neurosci. 105 (1991) 955–964.detoxification of substance abusers, Psychiatr. Serv. 50 (1999) [37] P.V. Piazza, S. Maccari, J.M. Deminiere, M. LeMoal, P. Mormede,813–817. H. Simon, Corticosterone levels determine individual vulnerability
[17] J.R. Glowa, A.E. Shaw, A.L. Riley, Cocaine-induced conditioned to amphetamine self-administration, Proc. Natl. Acad. Sci. USA 88taste aversions: comparison between effects in LEW/N and F344/N (1991) 2088–2092.rat strains, Psychopharmacology 114 (1994) 229–232. [38] M.A. Reicher, E.W. Holman, Location preference in flavor aversion
[18] F. Gomez, P.S. Grigson, The suppressive effects of LiCl, sucrose, reinforced by amphetamine in rats, Anim. Learn. Behav. 5 (1977)and drugs of abuse are modulated by sucrose concentration in food 343–346.deprived rats, Physiol. Behav. 67 (1999) 351–357. [39] S. Reilly, T.C. Pritchard, Gustatory thalamus lesions in the rat: II-
[19] A.J. Goudie, Aversive stimulus properties of drugs, Neurophar- Aversive and appetitive taste conditioning, Behav. Neurosci. 110macology 18 (1979) 971–979. (1996) 1355–1363.

58 F. Gomez et al. / Brain Research 863 (2000) 52 –58
[40] A.L. Riley, W.J. Jacobs, V.M. LoLordo, Morphine-induced taste [43] S.D. Turenne, C. Miles, L.A. Parker, S. Siegel, Individual differ-aversions: a consideration of parameters, Physiol. Psychol. 6 (1978) ences in reactivity to the rewarding/aversive properties of drugs:96–100. assessment by taste and place conditioning, Pharmacol. Biochem.
[41] G. Scalera, P.S. Grigson, R. Norgren, Gustatory functions, sodium Behav. 53 (1996) 511–516.appetite, and conditioned taste aversion survive excitotoxic lesions [44] N. White, L. Sklar, Z. Amit, The reinforcing action of morphine andof the thalamic taste area, Behav. Neurosci. 111 (1997) 633–645. its paradoxical side effects, Psychopharmacology 52 (1977) 63–66.
[42] J.E. Sherman, C. Pickman, A. Rice, J.C. Lieveskind, E.W. Holman, [45] R.A. Wise, R.A. Yokel, H. DeWit, Both positive reinforcement andRewarding and aversive effects of morphine: Temporal and pharma- conditioned aversion from amphetamine and from apomorphine incological properties, Pharmacol. Biochem. Behav. 13 (1980) 501– rats, Science 191 (1976) 1273–1275.505.