Embed Size (px)
Citation preview

Mixed-Strain Mycobacterium tuberculosis Infections and theImplications for Tuberculosis Treatment and Control
Ted Cohen,a,b Paul D. van Helden,c Douglas Wilson,d Caroline Colijn,e Megan M. McLaughlin,a Ibrahim Abubakar,f
and Robin M. Warrenc
Division of Global Health Equity, Brigham and Women’s Hospital, Boston, Massachusetts, USAa; Department of Epidemiology and Center for Communicable DiseaseDynamics, Harvard School of Public Health, Boston, Massachusetts, USAb; DST/NRF Centre of Excellence for Biomedical Tuberculosis Research/MRC Centre for Molecularand Cellular Biology, Division of Molecular Biology and Human Genetics, Faculty of Medicine and Health Sciences, Stellenbosch University, Tygerberg, South Africac;Department of Medicine, Edendale Hospital, Pietermaritzburg, University of KwaZulu-Natal, Durban, South Africad; Department of Mathematics, Imperial College London,London, United Kingdome; and Department of Medicine, Norwich Medical School, University of East Anglia, Norwich, and Tuberculosis Section, Health ProtectionAgency, London, United Kingdomf
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .708EVIDENCE OF MIXED INFECTION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .709CHALLENGES FOR THE DETECTION OF MIXED INFECTIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .709
Specimen Selection and Collection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .709Specimen Handling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .711Typing Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .711
EFFECTS OF MIXED INFECTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .713Individual-Level Effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .713Population-Level Effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .714
CONCLUSIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .714ACKNOWLEDGMENTS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .715REFERENCES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .715
INTRODUCTION
Despite the relative genetic conservation among Mycobacte-rium tuberculosis strains compared with many other bacterial
species (91), numerous approaches for distinguishing mycobac-terial isolates have been successfully developed. Early methodsbased on phage typing were capable of making only crude distinc-tions between mycobacterial strains. More recent developments(e.g., IS6110 restriction fragment polymorphism [RFLP] analysis,spoligotyping, and mycobacterial interspersed repetitive unit–variable-number tandem repeat [MIRU-VNTR] analysis), eachbased on the detection of differences in the numbers or locationsof particular genetic sequences or signatures, permit differentia-tion with much greater resolution (63). In addition to the obviouspublic health applications of these tools for the identification ofclusters of recent transmission, studies using molecular epidemi-ological methods have improved our understanding of the naturalhistory of disease and how tuberculosis (TB) spreads in commu-nities (72, 105).
Early molecular epidemiological studies conducted in settingswhere the incidence of tuberculosis was low found an unexpect-edly high proportion of disease that was attributable to recenttransmission (1, 89). These findings implied that control effortsthat focus on reducing the transmission of disease might have amore important role than previously appreciated (93), even insettings where the incidence of tuberculosis is low. Similarly, stud-ies in settings where the incidence of tuberculosis is high docu-mented that the majority of cases occur as a consequence of recenttransmission (103) and that many of these transmission events aretaking place outside households (108). Similar studies have alsoilluminated the importance of nosocomial (34, 39, 69, 79, 81, 83),penitentiary (17, 102), and casual (54, 71) social settings in disease
spread. These findings further underscore the epidemiologicalimportance of diagnostic delays (due to both patient health-seek-ing and health care provider behaviors) and suggest that morerapid diagnosis and initiation of appropriate drug treatment canplay an enormous role in interrupting transmission.
Molecular epidemiological studies also identified individualswith repeated episodes of disease due to reinfection, thus revealingthe incomplete immunizing effect of previous M. tuberculosis in-fection (18, 107). These data confirm previously reported obser-vational data (15, 45) and statistical arguments (4, 13, 96, 109)suggesting that previous exposure or disease does not confer com-plete protection against subsequent reinfection and have resultedin a paradigm shift in our understanding of the epidemiology oftuberculosis (62).
While reinfection is expected to occur most frequently in set-tings where the incidence of TB is high (since multiple exposuresare most likely) (80, 101, 111), studies in some moderate- andlow-incidence settings among individuals with repeated episodesof disease have found that reinfection also occurs at unexpectedlyhigh rates (14). Several explanations may account for the reinfec-tion documented among those with repeated episodes of tubercu-losis disease in low-incidence areas. Previously diseased individu-
Address correspondence to Ted Cohen, [email protected].
* Present address: Ibrahim Abubakar, Department of Infection and PopulationHealth, University College London, London, United Kingdom.
Supplemental material for this article may be found at http://cmr.asm.org/.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/CMR.00021-12
708 cmr.asm.org Clinical Microbiology Reviews p. 708–719 October 2012 Volume 25 Number 4
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

als may have particular biological characteristics (genetic oracquired immunodeficiency) or acquired or environmental riskfactors (alcoholism or nutritional deficiencies) that render themmore prone to the development of disease after (re)infection. It isalso possible that in low-incidence settings, the risk of exposure isconcentrated within “sociospatial pockets,” such that individualsinfected once are likely to be infected multiple times (19). In othermodels that account for superinfection, because each infection isnot immunizing, reinfection plays a dominant role in disease dy-namics when the disease prevalence increases past a threshold(44).
The fact that previous exposure provides incomplete protec-tion from subsequent infection may have important implicationsfor the implementation of interventions (e.g., treatment of latentinfection) (65) and for the prospect of developing effective pre-ventive or therapeutic vaccines (44). However, the overall impor-tance of reinfection within the context of the tuberculosis epi-demic has been questioned (56). Ongoing doubt concerning theepidemiological importance of reinfection may reflect the fact thatclear documentation of this phenomenon usually requires the col-lection of different strains from two disease episodes within anindividual. This approach underestimates the frequency of rein-fection since it cannot detect individuals who were repeatedly in-fected prior to developing a single episode of disease and will alsomisclassify reinfections as relapses when individuals are repeat-edly infected with similar strains that are predominant in theircommunity or social circle. Previously reported models suggestedthat a substantial fraction of first-time disease episodes is likely tobe the result of reinfection and that this fraction is dependent onboth the local prevalence of disease, which determines the force ofinfection that each individual experiences (19, 65), as well as thedegree of protection afforded by a previous infection (44).
More recently, molecular studies that documented the phe-nomenon of polyclonal or “mixed” infections, in which multiplestrains of M. tuberculosis are retrieved from an individual host at asingle point in time, underscored the potential importance of re-infection in tuberculosis epidemics (Table 1). This phenomenonof mixed infections is not unique to M. tuberculosis and appears tooccur with pathogens of all types (6). At present, accounts ofmixed M. tuberculosis infections are accumulating for various geo-graphic settings, but there have been few efforts to examine thepublic health implications of the fact that a single host can simul-taneously harbor more than one distinct strain of M. tuberculosis.In this paper, we review existing evidence of mixed M. tuberculosisinfections collected from many different epidemiological settings.We highlight why these studies, while very important for docu-menting that these types of infections occur in all settings, maygreatly underestimate the frequency of mixed infections due to thelimited sensitivity of current approaches for the detection of thepresence of mixed infections. We review evidence of the impor-tance of mixed infections for treatment outcomes of tuberculosispatients receiving standard regimens of combination therapy anddiscuss how mathematical models reveal the importance of mixedinfections to the projected trajectory of tuberculosis epidemicsand the effects of interventions. Throughout our review, we aim toidentify areas where additional research is needed to clarify howmixed M. tuberculosis infections modify disease dynamics andcontrol.
EVIDENCE OF MIXED INFECTION
References for this review were identified through searches ofPubMed for articles published by December 2011, by use of thekeywords “(mixed or multiple) AND strain AND tuberculosis.”Articles resulting from these searches and relevant references citedin those articles were reviewed. Articles published in English wereretained if they reported evidence of at least one patient with morethan one strain of M. tuberculosis on a single day. Depending onthe method used to detect mixed strains, more than one sample ortesting of more than one colony from the same day may have beenrequired in order to meet this inclusion criterion.
Selected details of studies documenting mixed M. tuberculosisinfection are provided in Table 1, and a more detailed account ofstudies can be found in Table S1 in the supplemental material; wealso list additional studies that did not meet our strict inclusioncriteria in Table S2 in the supplemental material.
CHALLENGES FOR THE DETECTION OF MIXED INFECTIONS
Figure 1 summarizes laboratory tools that have been used to iden-tify mixed-strain M. tuberculosis infections, each of which has sig-nificant shortcomings. While false-positive results are possible(especially where the risk of cross-contamination is not mini-mized within the laboratory), the most significant obstacle is thelimited sensitivity of these approaches; therefore, current esti-mates of the prevalence of mixed-strain infections serve as a lowerbound, and the actual proportion of cases with mixed infectionsmay be significantly larger. The detection of mixed infections isdependent on collecting a clinical specimen that contains multiplestrains, handling the specimen such that evidence of individualstrains remains intact, and using a typing method that can differ-entiate the strains. We discuss concerns related to each of thesesteps below.
Specimen Selection and Collection
A major impediment to the identification of mixed M. tuberculosisinfections is that it is not usually possible to isolate mycobacteriafrom individuals in the latent stage of infection using current tech-niques because bacillary loads are low and the sites of infection areinaccessible (a notable exception is found for lung resection [33]).Thus, the majority of studies in which mixed M. tuberculosis iso-lates have been identified used sputum samples from individualswith either diagnosed or suspected pulmonary tuberculosis. Themost sensitive methods for the detection of mixed infections ap-pear to be able to detect minority strains constituting �1% of thebacteria within a sample (106). Given that the lung bacillary bur-den in active tuberculosis has been estimated to be around 108 to1010 (43, 88), this means that minority strains may need to bepresent at extremely high numbers (107 to 109) at the time ofsampling to be detected, assuming that the diagnostic specimenrepresents a random sample of bacteria present within the body.
The numbers of specimens examined differ among studies, butmost often, a single sputum specimen is collected. Shamputa et al.found that increasing the number of sputum samples collectedincreased the likelihood of detection of mixed infections (86).Even if multiple sputum specimens are examined, it is likely thatnot all pulmonary lesions may be open to the airways at a giventime, and thus, bacteria present within sputum may not ade-quately reflect the heterogeneity in underlying infections (7). Thetiming of sputum collection relative to the initiation of treatmentis also likely to affect the sensitivity of tests for mixed infections
Mixed-Strain M. tuberculosis Infections
October 2012 Volume 25 Number 4 cmr.asm.org 709
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from
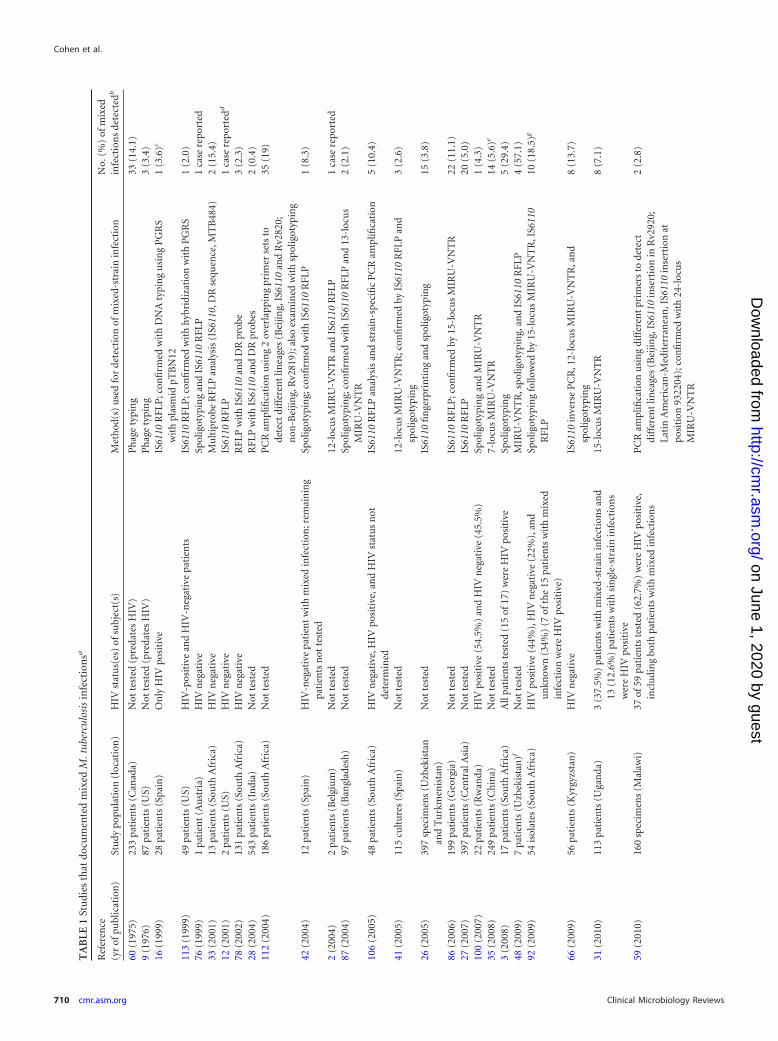
TA
BLE
1St
udi
esth
atdo
cum
ente
dm
ixed
M.t
uber
culo
sis
infe
ctio
nsa
Ref
eren
ce(y
rof
publ
icat
ion
)St
udy
popu
lati
on(l
ocat
ion
)H
IVst
atu
s(es
)of
subj
ect(
s)M
eth
od(s
)u
sed
for
dete
ctio
nof
mix
ed-s
trai
nin
fect
ion
No.
(%)
ofm
ixed
infe
ctio
ns
dete
cted
b
60(1
975)
233
pati
ents
(Can
ada)
Not
test
ed(p
reda
tes
HIV
)P
hag
ety
pin
g33
(14.
1)9
(197
6)87
pati
ents
(US)
Not
test
ed(p
reda
tes
HIV
)P
hag
ety
pin
g3
(3.4
)16
(199
9)28
pati
ents
(Spa
in)
On
lyH
IVpo
siti
veIS
6110
RFL
P;c
onfi
rmed
wit
hD
NA
typi
ng
usi
ng
PG
RS
wit
hpl
asm
idpT
BN
121
(3.6
)c
113
(199
9)49
pati
ents
(US)
HIV
-pos
itiv
ean
dH
IV-n
egat
ive
pati
ents
IS61
10R
FLP
;con
firm
edw
ith
hyb
ridi
zati
onw
ith
PG
RS
1(2
.0)
76(1
999)
1pa
tien
t(A
ust
ria)
HIV
neg
ativ
eSp
olig
otyp
ing
and
IS61
10R
FLP
1ca
sere
port
ed33
(200
1)13
pati
ents
(Sou
thA
fric
a)H
IVn
egat
ive
Mu
ltip
robe
RFL
Pan
alys
is(I
S611
0,D
Rse
quen
ce,M
TB
484)
2(1
5.4)
12(2
001)
2pa
tien
ts(U
S)H
IVn
egat
ive
IS61
10R
FLP
1ca
sere
port
edd
78(2
002)
131
pati
ents
(Sou
thA
fric
a)H
IVn
egat
ive
RFL
Pw
ith
IS61
10an
dD
Rpr
obe
3(2
.3)
28(2
004)
543
pati
ents
(In
dia)
Not
test
edR
FLP
wit
hIS
6110
and
DR
prob
es2
(0.4
)11
2(2
004)
186
pati
ents
(Sou
thA
fric
a)N
otte
sted
PC
Ram
plifi
cati
onu
sin
g2
over
lapp
ing
prim
erse
tsto
dete
ctdi
ffer
ent
linea
ges
(Bei
jing,
IS61
10an
dR
v282
0;n
on-B
eijin
g,R
v281
9);a
lso
exam
ined
wit
hsp
olig
otyp
ing
35(1
9)
42(2
004)
12pa
tien
ts(S
pain
)H
IV-n
egat
ive
pati
ent
wit
hm
ixed
infe
ctio
n;r
emai
nin
gpa
tien
tsn
otte
sted
Spol
igot
ypin
g;co
nfi
rmed
wit
hIS
6110
RFL
P1
(8.3
)
2(2
004)
2pa
tien
ts(B
elgi
um
)N
otte
sted
12-l
ocu
sM
IRU
-VN
TR
and
IS61
10R
FLP
1ca
sere
port
ed87
(200
4)97
pati
ents
(Ban
glad
esh
)N
otte
sted
Spol
igot
ypin
g;co
nfi
rmed
wit
hIS
6110
RFL
Pan
d13
-loc
us
MIR
U-V
NT
R2
(2.1
)
106
(200
5)48
pati
ents
(Sou
thA
fric
a)H
IVn
egat
ive,
HIV
posi
tive
,an
dH
IVst
atu
sn
otde
term
ined
IS61
10R
FLP
anal
ysis
and
stra
in-s
peci
fic
PC
Ram
plifi
cati
on5
(10.
4)
41(2
005)
115
cult
ure
s(S
pain
)N
otte
sted
12-l
ocu
sM
IRU
-VN
TR
;con
firm
edby
IS61
10R
FLP
and
spol
igot
ypin
g3
(2.6
)
26(2
005)
397
spec
imen
s(U
zbek
ista
nan
dT
urk
men
ista
n)
Not
test
edIS
6110
fin
gerp
rin
tin
gan
dsp
olig
otyp
ing
15(3
.8)
86(2
006)
199
pati
ents
(Geo
rgia
)N
otte
sted
IS61
10R
FLP
;con
firm
edby
15-l
ocu
sM
IRU
-VN
TR
22(1
1.1)
27(2
007)
397
pati
ents
(Cen
tral
Asi
a)N
otte
sted
IS61
10R
FLP
20(5
.0)
100
(200
7)22
pati
ents
(Rw
anda
)H
IVpo
siti
ve(5
4.5%
)an
dH
IVn
egat
ive
(45.
5%)
Spol
igot
ypin
gan
dM
IRU
-VN
TR
1(4
.3)
35(2
008)
249
pati
ents
(Ch
ina)
Not
test
ed7-
locu
sM
IRU
-VN
TR
14(5
.6)e
3(2
008)
17pa
tien
ts(S
outh
Afr
ica)
All
pati
ents
test
ed(1
5of
17)
wer
eH
IVpo
siti
veSp
olig
otyp
ing
5(2
9.4)
48(2
009)
7pa
tien
ts(U
zbek
ista
n)f
Not
test
edM
IRU
-VN
TR
,spo
ligot
ypin
g,an
dIS
6110
RFL
P4
(57.
1)92
(200
9)54
isol
ates
(Sou
thA
fric
a)H
IVpo
siti
ve(4
4%),
HIV
neg
ativ
e(2
2%),
and
un
know
n(3
4%)
(7of
the
15pa
tien
tsw
ith
mix
edin
fect
ion
wer
eH
IVpo
siti
ve)
Spol
igot
ypin
gfo
llow
edby
15-l
ocu
sM
IRU
-VN
TR
,IS6
110
RFL
P10
(18.
5)g
66(2
009)
56pa
tien
ts(K
yrgy
zsta
n)
HIV
neg
ativ
eIS
6110
inve
rse
PC
R,1
2-lo
cus
MIR
U-V
NT
R,a
nd
spol
igot
ypin
g8
(13.
7)
31(2
010)
113
pati
ents
(Uga
nda
)3
(37.
5%)
pati
ents
wit
hm
ixed
-str
ain
infe
ctio
ns
and
13(1
2.6%
)pa
tien
tsw
ith
sin
gle-
stra
inin
fect
ion
sw
ere
HIV
posi
tive
15-l
ocu
sM
IRU
-VN
TR
8(7
.1)
59(2
010)
160
spec
imen
s(M
alaw
i)37
of59
pati
ents
test
ed(6
2.7%
)w
ere
HIV
posi
tive
,in
clu
din
gbo
thpa
tien
tsw
ith
mix
edin
fect
ion
sP
CR
ampl
ifica
tion
usi
ng
diff
eren
tpr
imer
sto
dete
ctdi
ffer
ent
linea
ges
(Bei
jing,
IS61
10in
sert
ion
inR
v292
0;La
tin
Am
eric
an-M
edit
erra
nea
n,I
S611
0in
sert
ion
atpo
siti
on93
2204
);co
nfi
rmed
wit
h24
-loc
us
MIR
U-V
NT
R
2(2
.8)
Cohen et al.
710 cmr.asm.org Clinical Microbiology Reviews
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

(59, 78); ideally, pretreatment specimens would be collected, asheterogeneity should be more frequently detected in these unse-lected bacterial populations.
In several cases, mixed-strain M. tuberculosis infections havebeen identified by using specimens sampled from multiple sites(e.g., lung, spleen, blood, and lymph nodes) (9, 16, 74), includingspecimens collected during autopsy (22, 33). It is reasonable toassume that as the numbers and types of specimens included instudies are expanded, especially considering that some lineagesmay exhibit organ tropism (55), the sensitivity for the detection ofmixed infections will increase.
Specimen Handling
Even when multiple distinct strains are initially present in clinicalspecimens collected for analysis, the probability of detectingmixed infections is affected by how specimens are handled beforeand after they enter the laboratory. In most cases of mixed infec-tion, isolates will be present in differing frequencies within speci-mens, and minority variants may be preferentially lost, leading tounderestimates of mixed infections. Delays from collection to lab-oratory testing will likely decrease the sensitivity of methods todetect mixed infections, as the minority strains present may bepreferentially lost. Methods of specimen decontamination thathave been optimized to avoid overgrowth by other organisms maynot be ideal for facilitating the survival of these minority variants,as decontamination processes are known to substantially decreasethe number of viable organisms (53). To our knowledge, there areno existing studies that have examined the effect of delays or dif-ferent decontamination protocols on the yield of mixed-infectionstudies.
While most studies have employed a culture step to increase themycobacterial population prior to attempts to detect mixed infec-tions, the effect of culture on the ability to detect minority variantsis not clear (59). Amplification-based methods that employ lin-eage-specific PCR primers to probe for infections with multipleM. tuberculosis lineages directly from sputum samples (i.e., with-out the use of culture) have successfully documented mixed infec-tions (49, 59). When Martin et al. (61) compared direct methodsand indirect methods that included culture, they observed that inmany samples, the clonal composition changed significantly afterculture, and in some cases, only one strain was detected after cul-ture. Isolates may also exhibit marked differences in their abilitiesto grow and divide in different types of media (solid versus liquidmedium and Lowenstein-Jensen [LJ] versus Middlebrook me-dium, etc.); thus, differing culture methods may result in differentestimates of the prevalence of mixed infections (61).
Methods of collection of mycobacterial material from culturefor typing also differ between studies and are likely to affect thefrequency at which mixed strains are discovered. In some studies,multiple individual clones were isolated from different colonieson solid culture and separately analyzed for evidence of heteroge-neity (35, 41, 87), whereas in other studies, “sweeps” across mul-tiple colonies on solid cultures or samples from liquid cultureswere collectively assessed for evidence of mixed infection (9). Thechoice of how to sample from cultures may be restricted by theculture approach used and the typing method to be employed.
Typing Methods
The method chosen for strain typing affects the likelihood of suc-cessfully detecting mixed infections (Fig. 1). Since the diversity of49
(201
0)18
5pa
tien
ts(T
aiw
an)
1H
IV-p
osit
ive
pati
ent
iden
tifi
edP
CR
ampl
ifica
tion
usi
ng
diff
eren
tpr
imer
sets
tode
tect
diff
eren
tlin
eage
s(B
eijin
g,IS
6110
and
Rv2
820;
non
-B
eijin
g,R
v281
9)
21(1
1.3)
22(2
011)
56pa
tien
ts(S
outh
Afr
ica)
54of
56(9
6%)
HIV
posi
tive
,in
clu
din
gal
lpat
ien
tsw
ith
mix
edin
fect
ion
sSp
olig
otyp
ing
and
24-l
ocu
sM
IRU
-VN
TR
5(9
)
110
(201
1)46
6pa
tien
ts(T
aiw
an)
Not
repo
rted
PC
Ram
plifi
cati
onu
sin
gdi
ffer
ent
prim
erse
tsto
dete
ctdi
ffer
ent
linea
ges
(Bei
jing,
IS61
10an
dR
v282
0;n
on-
Bei
jing,
Rv2
819)
14(3
.0)
74(2
011)
780
pati
ents
(Spa
in)
5of
the
9pa
tien
tsw
ith
mix
edin
fect
ion
wer
eH
IVpo
siti
ve15
-loc
us
MIR
U-V
NT
Ran
d24
-loc
us
MIR
U-V
NT
R;
con
firm
edby
IS61
10R
FLP
h
9(1
.2)
50(2
012)
1,24
8pa
tien
ts(V
ietn
am)
Not
test
edIS
6110
RFL
P,s
polig
otyp
ing,
and
15-l
ocu
sM
IRU
-VN
TR
39(3
.1)i
aP
GR
S,po
lym
orph
icG
C-r
ich
sequ
ence
;DR
,dir
ect
repe
at.
bSo
me
stu
dies
use
dst
aged
sele
ctio
npr
oced
ure
s,so
the
frac
tion
ofsa
mpl
esex
hib
itin
gm
ult
iple
-str
ain
infe
ctio
ns
may
not
bere
pres
enta
tive
ofth
eu
nde
rlyi
ng
popu
lati
on.T
he
nu
mbe
r(p
erce
nt)
ofm
ixed
infe
ctio
ns
was
calc
ula
ted
amon
gth
ose
pati
ents
for
wh
omth
ere
was
the
pote
nti
alto
dete
ctm
ixed
stra
ins.
cT
he
orig
inal
stu
dyre
port
edan
addi
tion
aln
ine
pati
ents
wit
hm
ixed
infe
ctio
ns
for
wh
omis
olat
esco
llect
edat
two
diff
eren
tti
mes
wer
eco
mpa
red.
dT
he
orig
inal
stu
dyre
port
edan
addi
tion
alpa
tien
tfo
rw
hom
isol
ates
colle
cted
15da
ysap
art
wer
eco
mpa
red.
eT
hos
eau
thor
su
sed
two
orm
ore
alle
les
ata
sin
gle
MIR
Ulo
cus
tode
fin
em
ixed
infe
ctio
ns.
fP
atie
nts
wer
ese
lect
edto
un
derg
oM
IRU
-VN
TR
anal
ysis
ifth
eyh
addo
cum
ente
dh
eter
ores
ista
nce
.Am
ong
35pa
tien
tsst
udi
ed,7
pati
ents
had
docu
men
ted
het
eror
esis
tan
ce.
gIn
the
orig
inal
pape
r,u
sin
gtw
oor
mor
eal
lele
sat
asi
ngl
eM
IRU
locu
sto
defi
ne
am
ixed
infe
ctio
n,t
he
nu
mbe
rm
ixed
infe
ctio
ns
dete
cted
was
15(2
7.8%
).T
enof
thes
esa
mpl
esh
adtw
oor
mor
eal
lele
sat
two
orM
IRU
loci
.Th
eor
igin
alst
udy
repo
rted
data
for
252
pati
ents
.Sam
ples
un
derw
ent
MIR
U-V
NT
Ran
alys
isif
they
had
one
ofth
etw
om
ost
com
mon
spol
igot
ypes
.Th
ispr
oced
ure
may
hav
ele
dto
eith
eran
un
dere
stim
atio
n(s
ince
stra
ins
wit
h“m
ixed
spol
igot
ypes
”w
ould
beex
clu
ded)
oran
over
esti
mat
ion
(ift
he
two
mos
tpr
eval
ent
linea
ges
are
mor
elik
ely
tobe
invo
lved
inm
ixed
infe
ctio
ns)
ofth
epr
opor
tion
ofm
ixed
infe
ctio
ns.
hT
he
MIR
U-V
NT
Rm
eth
ods
use
dby
thos
eau
thor
sch
ange
ddu
rin
gth
eco
urs
eof
the
stu
dy;a
ssu
ch,t
he
sen
siti
vity
for
the
dete
ctio
nof
mix
edin
fect
ion
sm
ayn
oth
ave
been
con
stan
t.iT
hir
ty-n
ine
pati
ents
had
mix
edin
fect
ion
s,w
hic
hw
ere
defi
ned
byth
ose
auth
ors
asis
olat
esw
ith
both
diff
eren
tR
FLP
patt
ern
san
ddi
ffer
ent
spol
igot
ypes
.Th
ose
auth
ors
also
repo
rted
that
60pa
tien
tsw
ere
fou
nd
toh
ave
poss
ible
mix
edin
fect
ion
s,w
hic
hw
ere
defi
ned
asdo
ubl
eal
lele
sat
atle
ast
two
loci
inth
eM
IRU
-VN
TR
patt
ern
.Oft
hes
e60
pati
ents
,29
had
mix
edin
fect
ion
sco
nfi
rmed
byR
FLP
and
spol
igot
ypin
g.
Mixed-Strain M. tuberculosis Infections
October 2012 Volume 25 Number 4 cmr.asm.org 711
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

mycobacteria differs among settings (40), it is difficult to quantifythe operating characteristics of these tests for mixed infections.However, each approach has clear limitations that depend on thelimits of detection and the discriminatory capacity of each test.Tests based on mycobacteriophages are capable of only a gross
classification of strains and are expected to exhibit poor sensitivityfor the detection of mixed infections (90). IS6110-based ap-proaches for the detection of mixed infections in a single isolatecan be somewhat subjective since they require the identification ofmultiple strains based on different intensities of banding patterns,
FIG 1 Methods for detecting mixed infections (9, 28, 30, 41, 51, 57, 63, 76, 95, 104, 106, 112).
Cohen et al.
712 cmr.asm.org Clinical Microbiology Reviews
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

which may be difficult to reliably discern (30). The detection ofmixed infections by IS6110 analysis also requires that at least 10%of mycobacterial DNA originate from the minority strain. SomePCR-based approaches have utilized primers specific only at thelevel of lineages; as such, in these studies, it was not possible todetect mixed infections that occurred as a result of infection withmultiple strains within the same lineage or sublineage (59, 106,112), and lineages for which primers are not included would bemissed. Heteroresistance, which occurs as the result of sporadicresistance-conferring mutations arising throughout the course ofinfection (38) in clonal disease (and, thus, is not a true mixedinfection as we have defined here), will also be missed unless spe-cific methods to detect sequence diversity in resistance-relatedgenes are employed (94).
MIRU-VNTR-based methods, currently the most widely usedmethod for the detection of mixed infections, examine heteroge-neity only within a limited set of loci, which necessarily limitssensitivity. Furthermore, it is difficult to draw clear distinctionsbetween clonal heterogeneity (defined as clonal diversity occur-ring as the result of mutation events within the infected host) andmixed infections (defined as diversity occurring as the result ofmultiple infections) based on differences in MIRU-VNTR pat-terns (22, 73). There are vast differences between the rapidity withwhich these two mechanisms produce within-host diversity;clonal heterogeneity involves the sporadic appearance of sequen-tial single-nucleotide polymorphisms through mutation, whilemixed infection allows a host to acquire, wholesale, an entirelynew M. tuberculosis genome. Furthermore, the rate of acquisitionof mixed infections depends on the force of infection in the com-munity and the diversity of strains in that environment (i.e.,mixed infections depend on population-level events), whereasclonal heterogeneity depends entirely on within-host processes.
Whole-genome sequencing of M. tuberculosis bacteria collectedfrom individuals offers resolution that is not possible with othertyping methods. The ability to distinguish highly related but ge-netically distinct strains will soon enable better estimates of thetrue extent of mixed-strain infections. These data will also facili-tate the study of microevolution events and the functional impactof previous infection on superinfection and the progression toactive disease (37). The application of these methods is currentlylimited by the cost, the complexity of data processing and analysis,and the limitations of short sequencing reads to account for repeatregions (84). However, the rapidly falling costs of sequencers, theemergence of new-generation sequencers with longer reads, andthe development of new tools for bioinformatic analysis will helpto address these challenges in the very near future.
EFFECTS OF MIXED INFECTION
Despite the limited sensitivity of current methods for the detec-tion of mixed infections, researchers have been able to identifydeleterious effects of mixed infections for individuals who are af-fected and have begun to explore the potential consequences fordisease dynamics and control.
Individual-Level Effects
For some individuals with mixed-strain tuberculosis, the presenceof multiple strains will result in poor treatment outcomes (5, 75,98). van Rie et al. (106) described a clear example of the negativeindividual-level impact of a mixed infection by using stored spec-imens collected from a series of patients with multidrug-resistant
(MDR) tuberculosis. Using strain-specific PCR to detect mixedinfections, those researchers identified patients who harboredboth MDR and susceptible strains; detailed treatment data al-lowed them to describe how the MDR strain was able to persistand grow during treatment with first-line regimens and, upon aswitch to second-line regimens, how the susceptible strain wasable to reemerge. In addition to demonstrating relative fitnessdeficits of MDR strains in these patients, that study found thatmixed infections with strains of different resistance phenotypescould compromise treatment outcomes using standard combina-tion treatment regimens. Since the presence of underlying resis-tance (even monoresistance) erodes treatment success and in-creases the risk of acquired resistance with standardizedcombination therapy (58), we expect that mixed infections withresistant strains may be associated with poor outcomes in othersettings as well. Furthermore, that study, from South Africa,showed that apparent cases of sequential reinfection may be moreaccurately categorized as mixed infection using methods with agreater sensitivity for the detection of polyclonality. The within-host “unmasking” of MDR strains after first-line therapy may beconsidered a novel mechanism by which drug-resistant diseasemay occur. From a programmatic perspective, the unmasking ofunderlying drug-resistant strains that arose through transmissionwould result in a misclassification of these events as occurrences ofacquired drug resistance, thereby unfairly judging the quality oftreatment programs. Since national tuberculosis programs in thevast majority of settings with a high tuberculosis burden have alimited capacity to perform mycobacterial culture (with evenmore restrictions for the use of drug susceptibility testing), thelikelihood that tuberculosis patients receiving therapy will havefollow-up drug susceptibility testing performed during treatmentis low. Accordingly, the presence of an occult mixed infection atthe time of the initiation of therapy in areas where drug resistanceis prevalent or emerging poses a threat for most treatment pro-grams.
In addition to the ability of multiple-strain infections to com-promise the effectiveness of treatment, it is also possible that su-perinfection may occur some time after the primary infection,initiating disease progression and the endogenous reactivation ofthe primary infection (33). This implies that the primary infectionis unable to confer protection against a secondary infection. Asimilar conclusion was drawn from mouse model experimentsafter sequential infections with different M. tuberculosis strains(67, 77) or repeated infections with the same strain (47). Althoughthe role of reinfection in the pathogenic mechanism of progres-sion of tuberculosis is not well understood, it has been suggestedthat a secondary “infection” with the vaccine strain Mycobacte-rium bovis BCG may enhance the risk of developing tuberculosisin patients who probably had a previous M. tuberculosis infection(36). This implies that the vaccination process (equivalent to asecondary infection) altered the immune state of the host, therebyallowing M. tuberculosis (equivalent to the primary infection) toactively replicate and cause disease (68, 97). Alternatively, the ex-pression of resuscitation-promoting factors (RPFs) may be re-sponsible for stimulating the growth of a previously quiescentinfection (11, 29, 32, 46, 52, 70, 82). Deciphering these mecha-nisms may have important implications for the understanding ofprotective immunity and the development of new vaccines.
Data from animal models suggest that mycobacterial strainsexhibit complicated within-host dynamics. Elegant studies using
Mixed-Strain M. tuberculosis Infections
October 2012 Volume 25 Number 4 cmr.asm.org 713
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

fish, frogs, and mice found that superinfecting mycobacteria pref-erentially invade existing granulomas (24, 25). These findingsdemonstrate complex interactions between the pathogen andhost, whereby the pathogen “tricks” the host macrophages intotransporting the bacilli to an environment where replication mayreadily occur. However, in a mixed-infection study, the presenceof a hypervirulent strain was unable to alter the growth rate of thecoinfecting hypovirulent strain (7). Similarly, the overabundanceof a strain that induced a strong Th1 response was unable to con-trol the growth of the hypervirulent strain. These findings haveimportant implications for vaccine development, as it has longbeen thought that strains which induce a Th1 cytokine profile aregood vaccine candidates (7). The above-described studies did notexclude the possibility of synergy or antagonism between infectingstrains, as the strains used in the above-described study did notreflect the population structure of M. tuberculosis in communitysettings.
The relationship between HIV-associated immune compro-mise and the risk of harboring multiple strains of M. tuberculosishas not been well elucidated. In a study in Uganda using MIRU-VNTR analysis to detect mixed infections in 113 smear- andculture-positive patients, a higher proportion of patients withmixed-strain infections were HIV positive than patients with sin-gle-strain infections (37.5% versus 16.6%). In theory, we expectthat impaired cell-mediated immunity might increase the proba-bility of harboring multiple infections, but the shorter delay be-tween infection and disease and the higher risk of extrapulmonarydisease could theoretically offset the probability of either acquir-ing or detecting multiple-strain infections when they are present.
Population-Level Effects
Investigations of the potential population-level effects of mixedinfections require the use of mathematical models. While allmathematical models of tuberculosis are limited by uncertaintiesin the natural history of tuberculosis infection and disease (whichaffect both model structure and parameterization), models to ex-plore the consequences of mixed-strain M. tuberculosis infectionsrequire additional features that are difficult to model given ourlimited understanding of this phenomenon. Examples of addi-tional areas of uncertainty that are introduced into models thatconsider mixed infections include mechanisms of strain interac-tion/competition within coinfected hosts, the strain specificity ofthe immune responses, and the impact of previous infections onthe likelihood and consequences of reinfection. Nevertheless,models can play an essential role in helping to prioritize the ques-tions that must be answered to improve our ability to understandthe population-level consequences of mixed-strain infections andmay offer new qualitative insights even when we have substantialuncertainty about model structure and parameterization.
For example, it is clear from the simplest mathematical modelsof the dynamics of multistrain pathogens that when strains are instrict competition with one another for susceptible hosts (definedwhere each host can be infected by only a single strain), only thestrain with the highest reproductive number will persist. Relaxingthese models to allow that hosts may simultaneously harbor morethan one strain can substantially expand the potential behaviors ofthe system, allowing for the long-term persistence of strains thatwould have been outcompeted in simpler models (23, 114). Thisdiversity-promoting effect of mixed infections is similar to thepromotion of diversity in metapopulation models in ecology. In
contrast to simple resource-based ecological models, in which thespecies making the most effective use of a resource excludes com-peting species, metapopulation models have the flexibility to allowfor differences in the relative ability of plant species to grow withina patch (i.e., the ability of strains to expand within a host) and therelative ability of plants to be dispersed between patches (i.e., theability of strains to be transmitted between hosts). This can permitthe stable coexistence of species (99). A recent mathematicalmodel that allowed for differences in the abilities of M. tuberculosisstrains to compete within and between hosts determined thatmodels that allow for mixed infections can permit the long-termpersistence of relatively less transmissible drug-resistant strains ifthese strains benefitted from some competitive advantage withincoinfected hosts (85).
Mixed infections may also affect the projected population-levelimpact of antituberculosis interventions. For example, Colijn etal. (23) used a simple mathematical model to demonstrate howmixed infections expand the conditions under which the wide-spread use of isoniazid preventive therapy (IPT) selects for isoni-azid resistance at the population level. A model of the potentialeffects of new tuberculosis vaccines found that mixed infections,by facilitating the coexistence of multiple strain types, could beexpected to erode the expected benefits of a new vaccine that doesnot adequately cover all circulating strain types, since the vaccinewould be more likely to lead to strain replacement (20).
CONCLUSIONS
Previous reviewers have questioned the overall importance ofmixed infections for individuals with tuberculosis or for the dy-namics of tuberculosis within communities (10). In this paper, wepresent an overview of studies that documented mixed-strain in-fections, enumerate challenges for the detection of mixed infec-tions, and discuss the individual-level and potential population-level implications of this phenomenon.
The low sensitivity of most current approaches for the detec-tion of mixed infections constitutes a serious obstacle to our abil-ity to understand their importance. Our inability to reliably doc-ument existing mixed infections stems from challenges withspecimen collection (we may not retrieve all strains when obtain-ing our clinical sample), specimen handling (minority variantspresent in the clinical sample may be lost during transit, decon-tamination, culture, or sampling), and typing (the strain typingmethod may not be sensitive enough to pick up rare variants ormay not have adequate discriminatory power).
Despite these serious limitations, investigators have docu-mented that a substantial minority of tuberculosis patients (10 to20%) in certain settings are infected with multiple strains; thisstrongly suggests that mixed infections are very common. Givenexisting, though admittedly limited, evidence of the effect ofmixed infections on treatment outcomes (particularly whenstrains with different drug resistance phenotypes are involved)and the model-based arguments for the importance of mixed in-fections in tuberculosis dynamics and control, we believe thatmixed infections are an important subject for future research.
Proximal practical questions of interest include assessingwhich methods of specimen collection and processing are mostsensitive for documenting mixed infections. Extensions of currentcollection methods, including the collection of 24-h sputum sam-ples or the collection of strains from multiple anatomical sites,may result in clinical samples that better reflect the within-host
Cohen et al.
714 cmr.asm.org Clinical Microbiology Reviews
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

population structure of mycobacteria. Decontamination and cul-ture steps, while necessary prerequisites for many of the currenttyping methods, may also erode test sensitivity, and new methodsthat can type strains directly from sputum samples will be valu-able. Deep sequencing, which we expect to be used routinely forthe detection of mixed infections in the near future, is likely todisplace many other methods and should generate better data tounderstand many aspects of within-host diversity (37). New ani-mal studies of mixed infection can identify whether certain strainsare more capable of superinfection than others and clarify whetherprevious infection shapes the host immune response in such a waythat makes reinfection easier (47). The relevant scale of straininteractions for both individual- and population-level conse-quences of mixed infections is not known; direct competition be-tween individual organisms likely arises by local interactionswithin a single granuloma (24), whereas more indirect forms ofcompetition may occur as a result of interactions between strainsconfined to different granulomas or organs (33). Studies of thesetypes of interactions would allow for a more complete under-standing of strain competition and even the potential for strainsynergy.
Despite the limitations of current methods of detection andunanswered questions about effects, the current data suggest thatmixed infections are likely to have an important impact on accu-rate disease diagnosis, the effective treatment of individuals, andthe control of tuberculosis in populations. New studies aiming tomeasure the prevalence of mixed infections that use methods ofspecimen collection and handling that are likely to sample andprotect minority strains in combination with the most sensitivetyping methods are needed to provide better estimates of howcommon these mixed infections are. Laboratory, animal model,and epidemiological studies that focus on the within-host com-plexity of M. tuberculosis infection and disease can contributecomplementary information that improves our ability to effec-tively combat this pathogen. For example, an understanding of thestrengths and mechanisms of within-host strain interactions willallow us to better project the trajectories of tuberculosis epidemicsand the expected diversity of pathogen populations (8, 23, 85),understand the effects of the scaling up of existing interventions(e.g., IPT) (21), and evaluate the potential effectiveness of novelvaccines (20, 64).
ACKNOWLEDGMENTS
T.C. received support through award number DP2OD006663 from theOffice of the Director, U.S. National Institutes of Health. I.A. receivessupport from the United Kingdom National Institute for Health Researchthrough a senior research fellowship (award number SR-2011-04-001).
We thank Maria Kempner for assistance with an initial literaturesearch while we were exploring this topic.
The content is solely the responsibility of the authors and does notnecessarily represent the official views of the Office of the Director of theU.S. NIH or the NIH.
REFERENCES1. Alland D, et al. 1994. Transmission of tuberculosis in New York City. An
analysis by DNA fingerprinting and conventional epidemiologic meth-ods. N. Engl. J. Med. 330:1710 –1716.
2. Allix C, Supply P, Fauville-Dufaux M. 2004. Utility of fast mycobacte-rial interspersed repetitive unit-variable number tandem repeat geno-typing in clinical mycobacteriological analysis. Clin. Infect. Dis. 39:783–789.
3. Andrews JR, et al. 2008. Exogenous reinfection as a cause of multidrug-
resistant and extensively drug-resistant tuberculosis in rural South Af-rica. J. Infect. Dis. 198:1582–1589.
4. Andrews JR, et al. 2012. Risk of progression to active tuberculosis fol-lowing reinfection with Mycobacterium tuberculosis. Clin. Infect. Dis.54:784 –791.
5. Baldeviano-Vidalon GC, et al. 2005. Multiple infection with resistantand sensitive M. tuberculosis strains during treatment of pulmonarytuberculosis patients. Int. J. Tuberc. Lung Dis. 9:1155–1160.
6. Balmer O, Tanner M. 2011. Prevalence and implications of multiple-strain infections. Lancet Infect. Dis. 11:868 – 878.
7. Barczak AK, et al. 2005. In vivo phenotypic dominance in mouse mixedinfections with Mycobacterium tuberculosis clinical isolates. J. Infect.Dis. 192:600 – 606.
8. Basu S, Orenstein E, Galvani AP. 2008. The theoretical influence ofimmunity between strain groups on the progression of drug-resistanttuberculosis epidemics. J. Infect. Dis. 198:1502–1513.
9. Bates JH, Stead WW, Rado TA. 1976. Phage type of tubercle bacilliisolated from patients with two or more sites of organ involvement. Am.Rev. Respir. Dis. 114:353–358.
10. Behr MA. 2004. Tuberculosis due to multiple strains: a concern for thepatient? A concern for tuberculosis control? Am. J. Respir. Crit. CareMed. 169:554 –555.
11. Biketov S, et al. 2007. The role of resuscitation promoting factors inpathogenesis and reactivation of Mycobacterium tuberculosis during in-tra-peritoneal infection in mice. BMC Infect. Dis. 7:146. doi:10.1186/1471-2334-7-146.
12. Braden CR, et al. 2001. Simultaneous infection with multiple strains ofMycobacterium tuberculosis. Clin. Infect. Dis. 33:e42– e47. doi:10.1086/322635.
13. Brooks-Pollock E, Becerra MC, Goldstein E, Cohen T, Murray MB.2011. Epidemiologic inference from the distribution of tuberculosiscases in households in Lima, Peru. J. Infect. Dis. 203:1582–1589.
14. Caminero JA, et al. 2001. Exogenous reinfection with tuberculosis on aEuropean island with a moderate incidence of disease. Am. J. Respir.Crit. Care Med. 163:717–720.
15. Canetti G, Sutherland I, Svandova E. 1972. Endogenous reactivationand exogenous reinfection: their relative importance with regard to thedevelopment of non-primary tuberculosis. Bull. Int. Union Tuberc. 47:116 –134.
16. Chaves F, Dronda F, Alonso-Sanz M, Noriega AR. 1999. Evidence ofexogenous reinfection and mixed infection with more than one strain ofMycobacterium tuberculosis among Spanish HIV-infected inmates.AIDS 13:615– 620.
17. Chaves F, et al. 1997. A longitudinal study of transmission of tubercu-losis in a large prison population. Am. J. Respir. Crit. Care Med. 155:719 –725.
18. Chiang CY, Riley LW. 2005. Exogenous reinfection in tuberculosis.Lancet Infect. Dis. 5:629 – 636.
19. Cohen T, Colijn C, Finklea B, Murray M. 2007. Exogenous re-infectionand the dynamics of tuberculosis epidemics: local effects in a networkmodel of transmission. J. R. Soc. Interface 4:523–531.
20. Cohen T, Colijn C, Murray M. 2008. Modeling the effects of straindiversity and mechanisms of strain competition on the potential perfor-mance of new tuberculosis vaccines. Proc. Natl. Acad. Sci. U. S. A. 105:16302–16307.
21. Cohen T, Lipsitch M, Walensky RP, Murray M. 2006. Beneficial andperverse effects of isoniazid preventive therapy for latent tuberculosisinfection in HIV-tuberculosis coinfected populations. Proc. Natl. Acad.Sci. U. S. A. 103:7042–7047.
22. Cohen T, Wilson D, Wallengren K, Samuel EY, Murray M. 2011.Mixed-strain Mycobacterium tuberculosis infections among patientsdying in a hospital in KwaZulu-Natal, South Africa. J. Clin. Microbiol.49:385–388.
23. Colijn C, Cohen T, Murray M. 2009. Latent coinfection and the main-tenance of strain diversity. Bull. Math. Biol. 71:247–263.
24. Cosma CL, Humbert O, Ramakrishnan L. 2004. Superinfecting myco-bacteria home to established tuberculous granulomas. Nat. Immunol.5:828 – 835.
25. Cosma CL, Humbert O, Sherman DR, Ramakrishnan L. 2008. Traf-ficking of superinfecting Mycobacterium organisms into establishedgranulomas occurs in mammals and is independent of the Erp and ESX-1mycobacterial virulence loci. J. Infect. Dis. 198:1851–1855.
26. Cox HS, et al. 2005. The Beijing genotype and drug resistant tubercu-
Mixed-Strain M. tuberculosis Infections
October 2012 Volume 25 Number 4 cmr.asm.org 715
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

losis in the Aral Sea region of Central Asia. Respir. Res. 6:134. doi:10.1186/1465-9921-6-134.
27. Cox HS, et al. 2007. Risk of acquired drug resistance during short-coursedirectly observed treatment of tuberculosis in an area with high levels ofdrug resistance. Clin. Infect. Dis. 44:1421–1427.
28. Das S, et al. 2004. Simultaneous infection with multiple strains of My-cobacterium tuberculosis identified by restriction fragment length poly-morphism analysis. Int. J. Tuberc. Lung Dis. 8:267–270.
29. Davies AP, et al. 2008. Resuscitation-promoting factors are expressed inMycobacterium tuberculosis-infected human tissue. Tuberculosis(Edinb.) 88:462– 468.
30. de Boer AS, et al. 2000. Genetic heterogeneity in Mycobacterium tuber-culosis isolates reflected in IS6110 restriction fragment length polymor-phism patterns as low-intensity bands. J. Clin. Microbiol. 38:4478 – 4484.
31. Dickman KR, et al. 2010. Detection of multiple strains of Mycobacte-rium tuberculosis using MIRU-VNTR in patients with pulmonary tuber-culosis in Kampala, Uganda. BMC Infect. Dis. 10:349. doi:10.1186/1471-2334-10-349.
32. Downing KJ, et al. 2005. Mutants of Mycobacterium tuberculosis lack-ing three of the five rpf-like genes are defective for growth in vivo and forresuscitation in vitro. Infect. Immun. 73:3038 –3043.
33. du Plessis DG, Warren R, Richardson M, Joubert JJ, van Helden PD.2001. Demonstration of reinfection and reactivation in HIV-negativeautopsied cases of secondary tuberculosis: multilesional genotyping ofMycobacterium tuberculosis utilizing IS 6110 and other repetitive ele-ment-based DNA fingerprinting. Tuberculosis (Edinb.) 81:211–220.
34. Edlin BR, et al. 1992. An outbreak of multidrug-resistant tuberculosisamong hospitalized patients with the acquired immunodeficiency syn-drome. N. Engl. J. Med. 326:1514 –1521.
35. Fang R, et al. 2008. Mixed infections of Mycobacterium tuberculosis intuberculosis patients in Shanghai, China. Tuberculosis (Edinb.) 88:469 –473.
36. Fine PE. 2001. BCG: the challenge continues. Scand. J. Infect. Dis. 33:243–245.
37. Ford C, et al. 2012. Mycobacterium tuberculosis— heterogeneity re-vealed through whole genome sequencing. Tuberculosis (Edinb.) 92:194 –201.
38. Ford CB, et al. 2011. Use of whole genome sequencing to estimate themutation rate of Mycobacterium tuberculosis during latent infection.Nat. Genet. 43:482– 486.
39. Frieden TR, et al. 1996. A multi-institutional outbreak of highly drug-resistant tuberculosis: epidemiology and clinical outcomes. JAMA 276:1229 –1235.
40. Gagneux S, Small PM. 2007. Global phylogeography of Mycobacteriumtuberculosis and implications for tuberculosis product development.Lancet Infect. Dis. 7:328 –337.
41. Garcia de Viedma D, Alonso Rodriguez N, Andres S, Ruiz Serrano MJ,Bouza E. 2005. Characterization of clonal complexity in tuberculosis bymycobacterial interspersed repetitive unit–variable-number tandem re-peat typing. J. Clin. Microbiol. 43:5660 –5664.
42. Garcia de Viedma D, Marin M, Ruiz MJ, Bouza E. 2004. Analysis ofclonal composition of Mycobacterium tuberculosis isolates in primaryinfections in children. J. Clin. Microbiol. 42:3415–3418.
43. Gillespie SH. 2002. Evolution of drug resistance in Mycobacterium tu-berculosis: clinical and molecular perspective. Antimicrob. Agents Che-mother. 46:267–274.
44. Gomes MG, Franco AO, Gomes MC, Medley GF. 2004. The reinfectionthreshold promotes variability in tuberculosis epidemiology and vaccineefficacy. Proc. Biol. Sci. 271:617– 623.
45. Grzybowski S, Galbraith JD, Dorken E. 1976. Chemoprophylaxis trialin Canadian Eskimos. Tubercle 57:263–269.
46. Gupta RK, Srivastava BS, Srivastava R. 2010. Comparative expressionanalysis of rpf-like genes of Mycobacterium tuberculosis H37Rv underdifferent physiological stress and growth conditions. Microbiology 156:2714 –2722.
47. Henao-Tamayo M, et al. 2012. A mouse model of tuberculosis reinfec-tion. Tuberculosis (Edinb.) 92:211–217.
48. Hofmann-Thiel S, et al. 2009. Mechanisms of heteroresistance to isoni-azid and rifampin of Mycobacterium tuberculosis in Tashkent, Uzbeki-stan. Eur. Respir. J. 33:368 –374.
49. Huang HY, et al. 2010. Mixed infection with Beijing and non-Beijingstrains and drug resistance pattern of Mycobacterium tuberculosis. J.Clin. Microbiol. 48:4474 – 4480.
50. Huyen MN, et al. 2012. Mixed tuberculosis infections in rural SouthVietnam. J. Clin. Microbiol. 50:1586 –1592.
51. Kamerbeek J, et al. 1997. Simultaneous detection and strain differenti-ation of Mycobacterium tuberculosis for diagnosis and epidemiology. J.Clin. Microbiol. 35:907–914.
52. Kana BD, et al. 2008. The resuscitation-promoting factors of Mycobac-terium tuberculosis are required for virulence and resuscitation fromdormancy but are collectively dispensable for growth in vitro. Mol. Mi-crobiol. 67:672– 684.
53. Kent PT, Kubica GP. 1985. Public health mycobacteriology: a guide forthe level III laboratory. Centers for Disease Control, Atlanta, GA.
54. Klovdahl AS, et al. 2001. Networks and tuberculosis: an undetectedcommunity outbreak involving public places. Soc. Sci. Med. 52:681– 694.
55. Kong Y, et al. 2007. Association between Mycobacterium tuberculosisBeijing/W lineage strain infection and extrathoracic tuberculosis: in-sights from epidemiologic and clinical characterization of the three prin-cipal genetic groups of M. tuberculosis clinical isolates. J. Clin. Micro-biol. 45:409 – 414.
56. Lambert ML, et al. 2003. Recurrence in tuberculosis: relapse or reinfec-tion? Lancet Infect. Dis. 3:282–287.
57. Lazzarini LC, et al. 2012. Mycobacterium tuberculosis spoligotypes thatmay derive from mixed strain infections are revealed by a novel compu-tational approach. Infect. Genet. Evol. 12:798 – 806.
58. Lew W, Pai M, Oxlade O, Martin D, Menzies D. 2008. Initial drugresistance and tuberculosis treatment outcomes: systematic review andmeta-analysis. Ann. Intern. Med. 149:123–134.
59. Mallard K, et al. 2010. Molecular detection of mixed infections of My-cobacterium tuberculosis strains in sputum samples from patients inKaronga District, Malawi. J. Clin. Microbiol. 48:4512– 4518.
60. Mankiewicz E, Liivak M. 1975. Phage types of Mycobacterium tuber-culosis in cultures isolated from Eskimo patients. Am. Rev. Respir. Dis.111:307–312.
61. Martin A, Herranz M, Ruiz Serrano MJ, Bouza E, Garcia de ViedmaD. 2010. The clonal composition of Mycobacterium tuberculosis in clin-ical specimens could be modified by culture. Tuberculosis (Edinb.) 90:201–207.
62. Mathema B, Kurepina N, Fallows D, Kreiswirth BN. 2008. Lessonsfrom molecular epidemiology and comparative genomics. Semin. Re-spir. Crit. Care Med. 29:467– 480.
63. Mathema B, Kurepina NE, Bifani PJ, Kreiswirth BN. 2006. Molecularepidemiology of tuberculosis: current insights. Clin. Microbiol. Rev. 19:658 – 685.
64. McShane H, et al. 2012. BCG: myths, realities, and the need for alterna-tive vaccine strategies. Tuberculosis (Edinb.) 92:283–288.
65. Mills HL, Cohen T, Colijn C. 2011. Modelling the performance ofisoniazid preventive therapy for reducing tuberculosis in HIV endemicsettings: the effects of network structure. J. R. Soc. Interface 8:1510 –1520.
66. Mokrousov I, et al. 2009. Penitentiary population of Mycobacteriumtuberculosis in Kyrgyzstan: exceptionally high prevalence of the Beijinggenotype and its Russia-specific subtype. Infect. Genet. Evol. 9:1400 –1405.
67. Mollenkopf HJ, Kursar M, Kaufmann SH. 2004. Immune response topostprimary tuberculosis in mice: Mycobacterium tuberculosis and My-cobacterium bovis bacille Calmette-Guerin induce equal protection. J.Infect. Dis. 190:588 –597.
68. Moreira AL, et al. 2002. Mycobacterial antigens exacerbate disease man-ifestations in Mycobacterium tuberculosis-infected mice. Infect. Im-mun. 70:2100 –2107.
69. Moro ML, et al. 1998. An outbreak of multidrug-resistant tuberculosisinvolving HIV-infected patients of two hospitals in Milan, Italy. ItalianMultidrug-Resistant Tuberculosis Outbreak Study Group. AIDS 12:1095–1102.
70. Mukamolova GV, et al. 2002. A family of autocrine growth factors inMycobacterium tuberculosis. Mol. Microbiol. 46:623– 635.
71. Munch Z, et al. 2003. Tuberculosis transmission patterns in a high-incidence area: a spatial analysis. Int. J. Tuberc. Lung Dis. 7:271–277.
72. Murray M, Nardell E. 2002. Molecular epidemiology of tuberculosis:achievements and challenges to current knowledge. Bull. World HealthOrgan. 80:477– 482.
73. Narayanan S, et al. 2010. Impact of HIV infection on the recurrence oftuberculosis in South India. J. Infect. Dis. 201:691–703.
74. Navarro Y, et al. 2011. Systematic survey of clonal complexity in tuber-
Cohen et al.
716 cmr.asm.org Clinical Microbiology Reviews
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

culosis at a populational level and detailed characterization of the isolatesinvolved. J. Clin. Microbiol. 49:4131– 4137.
75. Niemann S, Richter E, Rusch-Gerdes S, Schlaak M, Greinert U. 2000.Double infection with a resistant and a multidrug-resistant strain of My-cobacterium tuberculosis. Emerg. Infect. Dis. 6:548 –551.
76. Pavlic M, Allerberger F, Dierich MP, Prodinger WM. 1999. Simulta-neous infection with two drug-susceptible Mycobacterium tuberculosisstrains in an immunocompetent host. J. Clin. Microbiol. 37:4156 – 4157.
77. Repique CJ, Li A, Collins FM, Morris SL. 2002. DNA immunization ina mouse model of latent tuberculosis: effect of DNA vaccination on re-activation of disease and on reinfection with a secondary challenge. In-fect. Immun. 70:3318 –3323.
78. Richardson M, et al. 2002. Multiple Mycobacterium tuberculosis strainsin early cultures from patients in a high-incidence community setting. J.Clin. Microbiol. 40:2750 –2754.
79. Rivero A, et al. 2001. High rate of tuberculosis reinfection during anosocomial outbreak of multidrug-resistant tuberculosis caused by My-cobacterium bovis strain B. Clin. Infect. Dis. 32:159 –161.
80. Romeyn JA. 1970. Exogenous reinfection in tuberculosis. Am. Rev. Re-spir. Dis. 101:923–927.
81. Rullan JV, et al. 1996. Nosocomial transmission of multidrug-resistantMycobacterium tuberculosis in Spain. Emerg. Infect. Dis. 2:125–129.
82. Russell-Goldman E, Xu J, Wang X, Chan J, Tufariello JM. 2008. AMycobacterium tuberculosis Rpf double-knockout strain exhibits pro-found defects in reactivation from chronic tuberculosis and innate im-munity phenotypes. Infect. Immun. 76:4269 – 4281.
83. Sacks LV, Pendle S, Orlovic D, Blumberg L, Constantinou C. 1999. Acomparison of outbreak- and nonoutbreak-related multidrug-resistanttuberculosis among human immunodeficiency virus-infected patients ina South African hospital. Clin. Infect. Dis. 29:96 –101.
84. Schurch AC, van Soolingen D. 2012. DNA fingerprinting of Mycobac-terium tuberculosis: from phage typing to whole-genome sequencing.Infect. Genet. Evol. 12:602– 612.
85. Sergeev R, Colijn C, Cohen T. 2011. Models to understand the popu-lation-level impact of mixed strain M. tuberculosis infections. J. Theor.Biol. 280:88 –100.
86. Shamputa IC, et al. 2006. Mixed infection and clonal representativenessof a single sputum sample in tuberculosis patients from a penitentiaryhospital in Georgia. Respir. Res. 7:99. doi:10.1186/1465-9921-7-99.
87. Shamputa IC, et al. 2004. Genotypic and phenotypic heterogeneityamong Mycobacterium tuberculosis isolates from pulmonary tubercu-losis patients. J. Clin. Microbiol. 42:5528 –5536.
88. Shimao T. 1987. Drug resistance in tuberculosis control. Tubercle68:5–18.
89. Small PM, et al. 1994. The epidemiology of tuberculosis in San Fran-cisco. A population-based study using conventional and molecularmethods. N. Engl. J. Med. 330:1703–1709.
90. Snider DE, Jr, Jones WD, Good RC. 1984. The usefulness of phagetyping Mycobacterium tuberculosis isolates. Am. Rev. Respir. Dis. 130:1095–1099.
91. Sreevatsan S, et al. 1997. Restricted structural gene polymorphism in theMycobacterium tuberculosis complex indicates evolutionarily recentglobal dissemination. Proc. Natl. Acad. Sci. U. S. A. 94:9869 –9874.
92. Stavrum R, et al. 2009. High diversity of Mycobacterium tuberculosisgenotypes in South Africa and preponderance of mixed infectionsamong ST53 isolates. J. Clin. Microbiol. 47:1848 –1856.
93. Stead WW. 1967. Pathogenesis of a first episode of chronic pulmonary
tuberculosis in man: recrudescence of residuals of the primary infectionor exogenous reinfection? Am. Rev. Respir. Dis. 95:729 –745.
94. Streicher EM, et al. 2012. Mycobacterium tuberculosis populationstructure determines the outcome of genetics-based second-line drugresistance testing. Antimicrob. Agents Chemother. 56:2420 –2427.
95. Supply P, et al. 2001. Automated high-throughput genotyping for studyof global epidemiology of Mycobacterium tuberculosis based on myco-bacterial interspersed repetitive units. J. Clin. Microbiol. 39:3563–3571.
96. Sutherland I. 1976. Recent studies in the epidemiology of tuberculosis,based on the risk of being infected with tubercle bacilli. Adv. Tuberc. Res.19:1– 63.
97. Taylor JL, et al. 2003. Pulmonary necrosis resulting from DNA vacci-nation against tuberculosis. Infect. Immun. 71:2192–2198.
98. Theisen A, et al. 1995. Mixed-strain infection with a drug-sensitive andmultidrug-resistant strain of Mycobacterium tuberculosis. Lancet 345:1512. doi:10.1016/S0140-6736(95)91073-5.
99. Tilman D. 1994. Competition and biodiversity in spatially structuredhabitats. Ecology 75:2–16.
100. Umubyeyi AN, et al. 2007. Molecular investigation of recurrent tuber-culosis in patients from Rwanda. Int. J. Tuberc. Lung Dis. 11:860 – 867.
101. Uys PW, van Helden PD, Hargrove JW. 2009. Tuberculosis reinfectionrate as a proportion of total infection rate correlates with the logarithm ofthe incidence rate: a mathematical model. J. R. Soc. Interface 6:11–15.
102. Valway SE, et al. 1994. Outbreak of multi-drug-resistant tuberculosis ina New York State prison, 1991. Am. J. Epidemiol. 140:113–122.
103. van der Spuy GD, et al. 2003. Use of genetic distance as a measure ofongoing transmission of Mycobacterium tuberculosis. J. Clin. Microbiol.41:5640 –5644.
104. van Embden JD, et al. 1993. Strain identification of Mycobacteriumtuberculosis by DNA fingerprinting: recommendations for a standard-ized methodology. J. Clin. Microbiol. 31:406 – 409.
105. van Helden PD. 2002. Molecular epidemiology of TB: challenging dog-mas and asking new questions. IUBMB Life 53:219 –223.
106. van Rie A, et al. 2005. Reinfection and mixed infection cause changingMycobacterium tuberculosis drug-resistance patterns. Am. J. Respir.Crit. Care Med. 172:636 – 642.
107. van Rie A, et al. 1999. Exogenous reinfection as a cause of recurrenttuberculosis after curative treatment. N. Engl. J. Med. 341:1174 –1179.
108. Verver S, et al. 2004. Proportion of tuberculosis transmission that takesplace in households in a high-incidence area. Lancet 363:212–214.
109. Vynnycky E, Fine PE. 1997. The natural history of tuberculosis: theimplications of age-dependent risks of disease and the role of reinfection.Epidemiol. Infect. 119:183–201.
110. Wang JY, et al. 2011. Mixed infection with Beijing and non-Beijingstrains in pulmonary tuberculosis in Taiwan: prevalence, risk factors, anddominant strain. Clin. Microbiol. Infect. 17:1239 –1245.
111. Wang JY, et al. 2007. Prediction of the tuberculosis reinfection propor-tion from the local incidence. J. Infect. Dis. 196:281–288.
112. Warren RM, et al. 2004. Patients with active tuberculosis often havedifferent strains in the same sputum specimen. Am. J. Respir. Crit. CareMed. 169:610 – 614.
113. Yeh RW, Hopewell PC, Daley CL. 1999. Simultaneous infection withtwo strains of Mycobacterium tuberculosis identified by restriction frag-ment length polymorphism analysis. Int. J. Tuberc. Lung Dis. 3:537–539.
114. Zhang P, Sandland GJ, Feng Z, Xu D, Minchella DJ. 2007. Evolution-ary implications for interactions between multiple strains of host andparasite. J. Theor. Biol. 248:225–240.
Continued next page
Mixed-Strain M. tuberculosis Infections
October 2012 Volume 25 Number 4 cmr.asm.org 717
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

Ted Cohen, M.D., M.P.H., D.P.H., is an epide-miologist whose research focuses on the trans-mission dynamics of infectious diseases. Dr.Cohen’s primary research focus is on the emer-gence and control of drug-resistant tuberculo-sis. He uses a combination of mathematicalmodeling techniques and traditional methodsfor epidemiological analysis in his research andis currently working on research projects inSouth America and sub-Saharan Africa. Dr. Co-hen has carried out studies that documentmixed infections and evaluate the potential population-level effects of mixedinfections. Dr. Cohen is an Assistant Professor in the Department of Epide-miology at Harvard School of Public Health and an Assistant Professor inMedicine in the Division of Global Health Equity at Brigham and Women’sHospital. Dr. Cohen holds an M.P.H. from the University of North Carolinaat Chapel Hill, an M.D. from the Duke University School of Medicine, and aD.P.H. from the Harvard School of Public Health.
Paul D. van Helden, Ph.D., is the Director ofthe DST/NRF Centre of Excellence for Biomed-ical Tuberculosis Research at Stellenbosch Uni-versity. Dr. van Helden is one of the world’smost prolific TB researchers. He started work-ing on tuberculosis in about 1990 and in thattime has focused the attention of the depart-ment mainly on this disease. In July 2009, Dr.van Helden was featured in an article onThompson Reuters’ Sciencewatch, citing him asthe fourth-highest-ranked scientist in the worldin the field of tuberculosis. He has been the recipient of a number of awards,including Stellenbosch University’s Vice Chancellors award for Excellence inResearch, Gold Medals by the SASBMB and the Academy of Science of SouthAfrica, the Silver medal of the MRC, and the NSTF award for outstandingcontribution to Science.
Douglas Wilson, M.B.Ch.B., is a clinician-re-searcher at Edendale Hospital, KwaZulu-Natal,South Africa. He, along with Dr. Cohen, hasconducted research documenting mixed-strainM. tuberculosis infection in South Africa. Dr.Wilson has also published on the diagnosis oftuberculosis and the treatment of tuberculosisin patients with HIV coinfection.
Caroline Colijn, M.E.S., Ph.D., is an appliedmathematician and Lecturer in Biomathemat-ics at the Imperial College London. Dr. Colijnworks on mathematical modeling applied to ep-idemiology. She is interested in coinfectingpathogens and the interplay between coinfec-tion, competition between pathogen variants,and pathogen evolution. She is also working onmathematical and computational models ofbacterial metabolism, with applications to M.tuberculosis and Bordetella pertussis. Dr. Colijn,along with Dr. Cohen, has evaluated the potential population-level conse-quences of mixed infections. Dr. Colijn earned her master of EnvironmentalStudies from York University and her Ph.D. in applied mathematics from theUniversity of Waterloo.
Megan M. McLaughlin, M.P.H., is a ResearchAssistant in the Division of Global Health Eq-uity at Brigham and Women’s Hospital. Sheconducts research in collaboration with clini-cians at the nonprofit organization Partners inHealth, where her focus has been on the evalu-ation of treatment outcomes and adverse effectsin patients treated for multidrug-resistant tu-berculosis and HIV coinfection in Lesotho. Ms.McLaughlin earned her M.P.H. from YaleUniversity.
Ibrahim Abubakar, M.B.B.S., M.Sc., Ph.D.,D.P.H. F.R.C.P., is a Clinical Professor andco-Director of the Centre for Infectious Dis-ease Epidemiology at University College Lon-don. He is also a Consultant Infectious Dis-ease Epidemiologist with responsibility fornational tuberculosis epidemiology and sur-veillance at the Health Protection Agency,London, United Kingdom. His research in-terests include the epidemiology of tubercu-losis, HIV, and hepatitis, evaluation of publichealth interventions and diagnostics, and the use of molecular epidemi-ology to inform the control of infectious disease. He holds an M.B.B.S.from Ahmadu Bello University, an M.Sc. from the London School ofHygiene and Tropical Medicine, a D.P.H. from the University of Cam-bridge, and a Ph.D. from the University of East Anglia.
Cohen et al.
718 cmr.asm.org Clinical Microbiology Reviews
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from

Robin M. Warren, Ph.D., is an Associate Pro-fessor in the Department of Medical Bio-chemistry at Stellenbosch University. His re-search interests focus on the disease dynamicsof the M. tuberculosis epidemic in two vastlydifferent communities, the evolution of theM. tuberculosis genome, the global spread ofdefined M. tuberculosis strains, the develop-ment of new epidemiological markers, the di-agnosis of MDR tuberculosis, the populationstructure of M. tuberculosis strains in the host,and the identification of mechanisms leading to drug resistance. Dr. War-ren has published over 160 papers and holds 5 patents. He is a coremember of the DST/NRF Centre of Excellence for Biomedical Tubercu-losis Research. Dr. Warren is a worldwide authority on mixed-strain M.tuberculosis infections. He, along with Dr. Paul van Helden, was amongthe first researchers to document mixed-strain M. tuberculosis infections,using DNA fingerprinting and spoligotyping, and to establish their role incompromising an individual’s response to treatment.
Mixed-Strain M. tuberculosis Infections
October 2012 Volume 25 Number 4 cmr.asm.org 719
on June 1, 2020 by guesthttp://cm
r.asm.org/
Dow
nloaded from