Embed Size (px)
Citation preview

Review
Mechanisms of cell recruitment in the immune responseto Mycobacterium tuberculosis
Wendy Peters a, Joel D. Ernst b,*a Gladstone Institute of Cardiovascular Disease, PO Box 419100, San Francisco, CA 94141-9100, USA
b Departments of Medicine and Microbiology, New York University School of Medicine, 550 First Avenue, NB 1658, New York, NY, 10016, USA
Abstract
Recent advances in understanding cell traffic, especially the roles of adhesion proteins, chemokines, and chemokine receptors, provide theopportunity for understanding mechanisms involved in the immune response to tuberculosis. This review concentrates on the roles of thesemolecules and the immune response in tuberculosis, based on studies of humans and mice infected with Mycobacterium tuberculosis.
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords: Tuberculosis; Immunity; Cell traffic
1. Introduction
Adaptive immunity to Mycobacterium tuberculosis isprincipally, if not exclusively, a function of a cellular immuneresponse. Consequently, initiation and maintenance of adap-tive immunity to tuberculosis require a high frequency ofspecific cellular interactions, which in turn require regulatedand efficient mechanisms in order for them to occur. Thepurpose of this review is to describe the present state ofknowledge of the mechanisms of recruitment of cells to sitesof infection with M. tuberculosis during the initiation, matu-ration, and maintenance of protective immunity. In particu-lar, it focuses on the evolving knowledge of the roles ofspecific chemokines, chemokine receptors, and adhesionproteins in the trafficking of macrophages, dendritic cells(DCs), and T lymphocytes to (and from) sites of infectionwith M. tuberculosis.
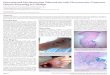
Granulomas are the characteristic histopathologic lesionsof tuberculosis, and can form in any tissue infected withM. tuberculosis. Granulomas exhibit a characteristic mixtureof macrophages and lymphocytes. In tuberculous granulo-mas, multinucleated Langhans giant cells may be found in acentral location, surrounded by macrophages (some of whichadopt an epithelioid morphology) and lymphocytes (Fig. 1).The presence of bacteria in macrophages in granulomas,together with the correlation between the cell populations
found in granulomas and the immune cell subsets that areessential for immune control of tuberculosis, strongly im-plies that granulomas represent the local immune response toinfection and are essential for restricting the growth of M. tu-berculosis.
2. Cellular requirements for protective immunity totuberculosis
At the cellular level, the requirements for immune controlof tuberculosis are reasonably well defined. Both CD4+ andCD8+ T cells are essential for effective control of M. tuber-culosis infection (reviewed in [1]). In mice [2,3] and humans[4], CD4+ T cells are approximately 2-fold more abundantthan are CD8+ T cells at sites of M. tuberculosis infection,and depletion or deletion of CD4+ T cells has an earlier andmore striking effect than elimination of CD8+ T cells on thecontrol of M. tuberculosis in mice [5]. Th1-polarized CD4+
effector cells are much more important than Th2 cells forcontrol of tuberculosis, as indicated by the rapid progressionof infection in mice lacking IFNc [6,7], and the negligibleeffect of the absence of IL-4 [8,9]. In addition to thesepredominant subsets of T lymphocytes that express ab anti-gen receptors, T cells that express cd antigen receptors areactivated in animals and humans infected with M. tuberculo-sis [10,11], and have been found to have both protective [12]and anti-inflammatory [13] roles in experimental tuberculo-sis in mice.
* Corresponding author. Tel.: +1-212-263-5165.E-mail address: [email protected] (J.D. Ernst).
Microbes and Infection 5 (2003) 151–158
www.elsevier.com/locate/micinf
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.DOI: 1 0 . 1 0 1 6 / S 1 2 8 6 - 4 5 7 9 ( 0 2 ) 0 0 0 8 2 - 5

While essential roles for T lymphocytes are well estab-lished in the control of M. tuberculosis, the roles of B lym-phocytes are less striking and much less well understood. Bcells are recruited to the lungs of mice infected with M. tuber-culosis and contribute to granuloma formation, yet mice thatlack mature B cells are able to control the growth of thebacteria in the lungs [14,15]. The observation that B-cell-deficient mice recruit fewer neutrophils, macrophages, andCD8+ T lymphocytes to their lungs implies a role for B cellsin the regulation of chemokine and/or adhesion protein ex-pression after infection with M. tuberculosis. Similarly, tran-sient depletion of neutrophils does not affect the survival ofmice or microbiological course of infection with M. tubercu-losis [16], but neutrophils clearly contribute to the formationof normal granulomas (see below).
Macrophages are commonly regarded as the phagocyticcells that initially ingest M. tuberculosis, and provide animportant cellular niche during infection.At some point earlyin the infection, M. tuberculosis also disseminates from thealveoli [17]. It is presently unclear as to whether this is due totransport from the alveoli by motile cells, or due to the directinvasion of alveolar epithelial cells and endothelial cells bysome of the bacteria, although recent in vitro studies supportboth models [18]. Macrophages are also considered to beimportant effector cells of the cellular immune response intuberculosis. Macrophages can be activated by cytokines andby direct contact with activated T cells to restrict the growthof M. tuberculosis and thereby control the infection once anadaptive immune response has developed [19,20].
DCs have only recently been characterized as having arole in the immune response to tuberculosis, and little infor-mation exists with respect to their roles during in vivo infec-tion. DCs are considered to be professional antigen-presenting cells (APCs), due to their ability to endocytoseantigens and express abundant quantities of MHC class II,costimulating molecules, and cytokines (reviewed in [21]).Recent studies have revealed that human and murine DCs caningest M. tuberculosis, and that DCs exposed to M. tubercu-losis undergo a typical maturation program in vitro andupregulate their antigen-presenting activities [22–24]. In ad-dition, it was recently shown that DCs, but not macrophages,
infected with M. tuberculosis are capable of driving Th1polarization of naïve CD4+ T cells [25]. Therefore, it islikely, albeit unproven, that DCs in the lung play an impor-tant role in initiating the immune response in tuberculosis.An additional property of DCs that contributes to their effec-tiveness in initiating immune responses in vivo is their abilityto migrate from peripheral tissues to secondary lymphoidtissues after acquiring antigens in the presence of proinflam-matory stimuli (detailed below).
In summary, the challenge to the immune system to gen-erate a protective response and control M. tuberculosis infec-tion requires the delivery of the bacteria and/or bacterialantigens by resident and/or recruited APCs to secondarylymphoid tissues, where the APCs must migrate to the re-gions enriched in naïve CD4+ and CD8+ T cells (the paracor-tical zone of lymph nodes or the periarteriolar lymphoidsheath in the spleen). After the activation of naïve T cells viatheir antigen receptors and costimulatory receptors in thepresence of polarizing cytokines such as IL-12, the activatedT cells must migrate via the bloodstream to the site(s) ofinfection, emigrate from the intravascular space into thetissue, and deliver macrophage-activating cytokines and/orkill infected cells by direct contact (Fig. 2). In a chronicinfection such as tuberculosis, this results in the formation ofgranulomas, and effective cell recruitment must be sustainedfor the lifetime of the host in order to maintain control of theinfection. These processes require active signals and sensors,as well as mechanisms to allow cells to adhere and migrateappropriately.
3. Chemokines and chemokine receptors
Chemokines are small-molecular-mass chemotactic cy-tokines (8–14 kDa) that mediate constitutive and inflamma-tory recruitment of leukocytes from the blood into tissues.
Fig. 1. Photomicrograph of a granuloma from a human with tuberculosis(hematoxylin and eosin stain). Original magnification 400×.
Fig. 2. Cell traffic in the immune response in tuberculosis. Specific cellsubsets are indicated by numbers: (1) monocyte, (2) dendritic cell precursor,(3) tissues macrophage, (4) tissue (immature) dendritic cell, (5) maturedendritic cell, (6) T-cell progenitor, (7) naïve T cell, and (8) effector T cell.
152 W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158

Chemokines (approximately 40 have been identified to date[26]) exert their effects through seven-transmembranedomain/G protein-coupled receptors (of which 19 areknown). Chemokines are grouped into four structurally re-lated families (designated CC, CXC, C, or CX3C), deter-mined by the numbers of amino acids that separate the twocysteine residues closest to the amino terminus. Chemokinereceptor nomenclature has been based upon these familyclassifications; the receptors that interact with the CCchemokines are termed CCR1-11, whereas the chemokinereceptors that interact with the CXC chemokines areCXCR1-5. The chemokine nomenclature system uses a simi-lar method, using “L” for ligand. Since this is still a fairlynew system, this review will give the former chemokinenames as well as the designations from the new classifica-tion. For nearly every receptor, there is more than one ligand,and many chemokines have more than one receptor. Thesefeatures suggest chemokine redundancy and promiscuity,respectively—phenomena that may represent different ana-tomical and kinetic expression of chemokines that signalthrough the same receptor.
3.1. Chemokine biology: constitutive vs. inflammatoryleukocyte recruitment
Recruitment of leukocytes into tissues can be mediatedeither in a constitutive fashion or induced by inflammation.Constitutive recruitment accounts for the accumulation ofnaïve and central memory T cells and B cells in secondarylymphoid tissues such as lymph nodes and the spleen, as wellas for establishing resident populations of tissue macroph-ages and DCs. Inflammatory recruitment accounts for theaccumulation of mononuclear phagocytes, lymphocytes andeffector cells at sites of local infection or inflammation.These distinct events are generally mediated by distinctchemokines and their receptors. For instance, the constitutiverecruitment of naïve T cells from the high endothelialvenules (HEVs) into secondary lymphoid organs is carriedout through CCL21/SLC expression on the HEV and CCR7on the T cells [27]. CXCL12/SDF-1 and CXCR4 have alsobeen implicated in this process [28]. In contrast, numerouschemokines can recruit effector T cells into inflammatorytissues. For instance, Th1 cells express CCR1, CCR2, CCR5,CXCR3, and CXCR5. This allows effector CD4+ T cells torespond to chemokines induced by intracellular infections,especially CCL2, 7, 8, 12 and 13 (the monocyte chemoattrac-tant proteins (MCPs 1–5), CCL5/RANTES, CCL3/MIP1a,CCL4/MIP1b, CXCL10/IP-10, CXCL9/mig and CXCL11/I-TAC) (for a review see [29]).
Monocytes and macrophages, like T cells, express distinctchemokine receptors, depending on their state of differentia-tion and activation. In many infections and immune-mediated inflammatory responses, recruitment of monocytesand macrophages into inflamed tissues is mediated by CCR2and CCR5, responding to the MCPs, CCL3/MIP1a,CCL4/MIP1b, and/or CCL5/RANTES [30]. In contrast, con-stitutive trafficking of monocytes to establish populations of
resident tissue macrophages may be mediated byCXCL14/BRAK [31], a constitutively expressed chemokinefor which a receptor has yet to be identified.
DCs exist in different subsets, depending in part on theiranatomical location. Monocyte-derived DCs express CCR1and CCR5 and respond to CCL3/MIP1a and CCL4/MIP1b,whereas CD11c+ DCs in the blood respond to the MCPs viaCCR2 [3]. After phagocytosing or endocytosing antigens inthe presence of an inflammatory stimulus (whether exog-enous and recognized by a Toll-like receptor, or endogenous,such as TNFa), DCs mature and upregulate CCR7 expres-sion. Upon expression of CCR7, mature antigen-loaded DCsmigrate from peripheral tissues to the paracortical regions oflocal draining lymph nodes, in response to CCL21/SLC andCCL19/ELC, which are constitutively expressed in lym-phatic endothelium and lymph node paracortex [32–34].Once they have migrated to the paracortex, antigen-loadedDCs can contact naïve T lymphocytes and initiate their acti-vation.
3.2. Studies of chemokines and tuberculosis
Several studies have established that M. tuberculosis caninduce the expression of chemokines, and that certainchemokines can be detected in clinical samples from peoplewith active tuberculosis. These studies are summarized inTable 1.
3.3. Studies of tuberculosis in chemokine and chemokinereceptor knockout mice
Although it is recognized that the accumulation of leuko-cytes in the lungs is critical to the establishment of granulo-mas and cell-mediated immunity to M. tuberculosis, themechanisms of recruitment have been largely unknown. Theuse of chemokine and chemokine receptor knockout micehas been useful in this regard. At this time, data fromCCR2–/– and MCP-1–/– mice have been reported. WhenCCR2–/– mice were infected with M. tuberculosis, they werefound to have a rapid and progressive infection that resultedin the death of 90% of the mice by day 24 post-infection(compared to no deaths in the infected wild-type controls)[3]. The number of bacteria in the lungs of the CCR2–/– micereflected this susceptibility, with 100-fold more colony-forming units in their lungs than in the CCR2+/+ controls.The lack of CCR2 caused an early and persistent decrease inthe number of macrophages recruited to the lungs over thecourse of infection, even though resident populations ofmacrophages were equivalent in uninfected CCR2–/– andCCR2+/+ mice. In addition to the decrease in macrophagerecruitment to the lungs, the lung-draining mediastinallymph node (MLN) also had fewer numbers of macrophagesat early, intermediate and late time points after infection. Inaddition, there were fewer DCs recruited to the MLNthroughout the infection. Since both these cell types canserve as APCs, it was not surprising that IFNc productionafter purified protein derivative (PPD) restimulation of MLN
153W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158

cells was reduced in CCR2–/– compared to CCR2+/+ mice.This probably reflected fewer T cells being activated, due tothe deficiency of APCs, since similar numbers of CD4+ andCD8+ T cells were present in the MLN of the CCR2–/– andCCR2+/+ controls. The accumulation of activated CD4+ andCD8+ and IFNc-producing CD4+ and CD8+ T cells to thelungs of infected CCR2–/– mice was also defective, whichwas especially marked later in the infection when T-cellrecruitment to the lungs of CCR2+/+ mice was maximal. Thiscould have reflected a direct effect of the lack of CCR2 oneffector T cells or could be indirectly related to the earlydefect in macrophage trafficking to the lungs and the activa-tion of T cells in the MLN. To resolve these two possibilities,we have recently used a mixed bone-marrow transfer schemeto obtain data that indicate that the defect in the recruitmentof T cells to the lungs of M. tuberculosis-infected CCR2–/–
mice is not due to the deficiency of CCR2 on T cells. Inmixed bone-marrow chimeras, the accumulation of T cells inthe lungs after M. tuberculosis infection correlated with thepresence of CCR2 on APCs (macrophages and DCs), but notthe presence of CCR2 on T lymphocytes (WP and JDE,unpublished). Therefore, the susceptibility of the CCR2–/–
mice to M. tuberculosis infection is probably due to a com-bination of a deficiency in both innate and adaptive immuneresponses, beginning at the level of the macrophage.
Another report supports an important role for CCR2 incontrol of M. tuberculosis infection. When transgenic micethat constitutively expressed CCL2/MCP-1 at moderate lev-els were infected with M. tuberculosis, they were more sus-ceptible to the disease than wild-type mice, similar to thefindings with CCR2–/– mice [41]. This effect of constitutiveexpression of CCL2/MCP-1 may be due to desensitization ofCCR2 and consequent hyporesponsiveness to all CCR2ligands, or to the lack of a concentration gradient ofCCL2/MCP-1 which would disable directed migration ofcells to the site of infection, or both. Since CCR2 is the onlyknown receptor for CCL2/MCP-1, these findings are consis-tent with those in CCR2–/– mice.
Interestingly, in contrast to the markedly defective re-sponse to M. tuberculosis in CCR2–/– mice, MCP-1–/– mice(lacking the major ligand for CCR2) were only slightly less
able than wild-type mice to control M. tuberculosis, withonly a transient excess of bacterial growth in their lungs andsimilar numbers of bacteria in their livers and spleens [42].The difference in susceptibility between the MCP-1–/– andthe CCR2–/– mice may be explained by the existence ofCCL7, 8 and 12/MCP-2, 3 and 5 that may compensate for thelack of CCL2/MCP-1. Furthermore, although CCR2–/– micecan have an IFNc production defect, MCP-1–/– mice havebeen reported to possess a Th2, but not a Th1, cytokine defect[43]. Therefore, the difference in the survival of these twomutant strains of mice may be due to differential defects intheir T-cell cytokine responses. These studies strongly sup-port an essential role for CCR2 and its ligands in the recruit-ment of macrophages to the lungs in tuberculosis, and implythat CCR2-dependent cell recruitment is required for optimalinitiation and polarization of T-lymphocyte responses.
Activated neutrophils have been found to secrete theCXCR3 ligands mig/CXCL9, IP-10/CXCL10, andI-TAC/CXCL11 [44], and a recent study has revealed anessential role for CXCR3 ligands in establishing normalgranuloma architecture in M. tuberculosis-infected mice.Transient depletion of neutrophils at the time of infectionwas found to have a long-lasting effect on granuloma archi-tecture, which correlated with a deficiency of mig/CXCL9and, to a lesser extent, I-TAC/CXCL11. The importance ofthese transient chemokine deficiencies was supported byfinding a similar defect in granuloma formation in M. tuber-culosis-infected CXCR3–/– mice (P. Seiler and S. Kaufmann,personal communication). Therefore, proper temporal andspatial expression of CXCR3 ligands is required for localorganization of cells in granulomas, and a transient defect inthese chemokines can have long-lasting effects on the orga-nization of the immune response.
4. Adhesion proteins
As noted above, leukocyte recruitment to sites of infectionor inflammation requires expression of one or more vascularaddressins on endothelial cells, and on chemokine-inducedactivation of integrins on specific leukocytes. One of the
Table 1Chemokines in human tuberculosis
Chemokines Cellular source/clinical source CitationCXCL8 (Interleukin-8) Produced and secreted by M. tuberculosis-stimulated human alveolar macrophages;
detected in bronchoalveolar lavage fluid of patients with active tuberculosis.[35]
CXCL8 Secreted by pulmonary epithelial cells (A549 and normal human bronchial epithelial cells)exposed to M. tuberculosis or supernatants from M. tuberculosis-infected macrophages.
[36]
CCL2 (MCP-1) Spontaneously produced by peripheral blood monocytes of TB patients; also detected byimmunohistochemistry in lymph nodes of patients with active tuberculosis.
[37]
CCL2, CXCL8, CCL3 (MIP1$), CCL5 (RANTES) Produced by M. tuberculosis-stimulated human alveolar macrophages and peripheralblood monocytes; bronchoalveolar lavage fluid from patients with active tuberculosiscontained CCL2, CXCL8, CCL5, but no detectable CCL3.
[38]
CCL3, CCL4 (MIP1b), CCL5 Produced by M. tuberculosis-stimulated human alveolar macrophages [39]CXCL9 (Mig), CXCL10 (IP-10), CXCL11 (I-TAC) Produced by IFNc-stimulated primary bronchial epithelial cells or respiratory epithelial
cell lines; CXCL10 mRNA found in bronchial epithelium and bronchoalveolar lavagecells of TB patients
[40]
154 W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158

vascular addressins that has been found to have a crucial rolein the formation of granulomas in M. tuberculosis-infectedmice is ICAM-1. Mice expressing mutant ICAM-1 mol-ecules that are unable to interact with CD11b/CD18 (amb2
integrin) exhibit defective recruitment of macrophages to thelungs after aerosol infection with M. tuberculosis, fail toform well-organized granulomas, and die after approxi-mately 4 months of infection [45,46]. CD11b/CD18 is con-sidered to be the major monocyte/macrophage integrin thatinteracts with ICAM-1; however, mice lacking CD11b (onany of three genetic backgrounds) have been found to exhibitno alteration in the ability to control the growth of M. tuber-culosis, no apparent difference in granuloma architecture,and no difference in survival after M. tuberculosis infection[47]. Together, the results in ICAM-1 mutant and CD11bknockout mice indicate that an interaction of leukocytes withone or more domains of ICAM-1 is essential for the recruit-ment of macrophages and granuloma formation in responseto M. tuberculosis, but that ligands for ICAM-1 other thanCD11b/CD18 are able to compensate for the absence of thisintegrin in mediating the recruitment of macrophages. Inaddition to the roles played by integrins, CD44, a receptor forhyaluronate, can mediate adhesion of leukocytes to extracel-lular matrix and can mediate cell–cell interactions. A recentanalysis of CD44–/– mice has revealed that they exhibit de-fects in macrophage and CD4+ T-lymphocyte recruitment tothe lungs after infection with virulent M. tuberculosis. Thesecell recruitment defects were associated with a defect inIFNc production in the lungs, poorer control of mycobacte-rial growth in the lungs and liver, poor formation of granulo-mas, and reduced survival of the infected CD44–/– mice (J.C.Leemans and T. van der Poll, personal communication). Thesimilarities in the phenotypes of CD44–/– and CCR2–/– miceafter infection with M. tuberculosis suggest that these mol-ecules may mediate events in a common pathway. Indeed,hyaluronate fragments have been found to induce CD44-dependent release of MCP-1 from human pulmonary mac-rophages [48].
As in the recruitment of macrophages from the blood-stream to tissue sites of infection, the recruitment of T lym-phocytes also depends on chemokines, vascular addressins,and integrins. Nearly all the CD4+ and CD8+ T lymphocytesisolated from the lungs of M. tuberculosis-infected miceexpress markers characteristic of activated, effector cells,rather than those of naïve T cells [49]. While several distinctintegrins have been found to be expressed on activated Tlymphocytes, a4b1 is the most prevalent on CD4+ and CD8+
T cells isolated from the lungs of mice infected with M. tu-berculosis [50]. Administration of an anti-a4 monoclonalantibody beginning 14 d after aerosol infection with M. tu-berculosis (when activated T cells begin to appear in thelung) blocked the accumulation of lymphocytes in the lung,prevented the formation of well-organized granulomas, andcaused early death of the mice. While activated CD4+ andCD8+ T lymphocytes expressing a4b7 integrin were a minorpercentage of lymphocytes isolated from the lungs of M. tu-
berculosis-infected mice, administration of a monoclonalantibody specific for the a4b7 heterodimer had an effect thatclosely resembled that of the antibody to a4 [50]. Thesefindings, together with the observation that the vascular ad-dressin recognized by a4b1 and a4b7 integrins, VCAM-1, isinduced in the pulmonary endothelium of M. tuberculosis-infected mice, indicate that these adhesion molecules play amajor role in the trafficking of activated CD4+ and CD8+ Tlymphocytes to sites of infection in the lung.
Studies of adhesion protein expression in humans withtuberculosis have been limited. However, a recent extensivecharacterization of peripheral blood monocytes, NK cells, aband cd T cells revealed several intriguing observations. Inparticular, a higher proportion of cd but not ab, T cells in theperipheral blood of people with active tuberculosis diseaseexpressed ICAM-1 and VLA-4 (a4-containing integrin) com-pared to healthy controls [10]. Whether the differential ex-pression of these adhesion proteins on cd vs. ab T cells in theperipheral blood is due to differences in the activation of thetwo cell types or in trafficking of the activated T cells fromthe blood to the sites of disease in the lungs, will requireadditional study.
5. Tumor necrosis factor-a and lymphotoxin-a
Defective granuloma formation and inability to controlM. tuberculosis infection has been noted in mice given aTNFa-neutralizing antibody or lacking TNFR p55, indicat-ing a role for TNFa and/or LTa, which both depend onTNFRp55 [51]. Determination of the respective roles ofTNFa and LTa actually revealed that they have closely re-lated, but not identical, roles. Mice selectively lacking TNFaalone are unable to control low-dose aerosol infection withM. tuberculosis, and die between days 28 and 35, with muchhigher bacterial loads in the lungs than in wild-type controls[52]. On day 28 of infection, TNFa-deficient mice exhibit nodifferences compared to wild-type mice in the percentage oftotal or activated CD4+ or CD8+ T cells in the lung, or in theamount of nitrite in the serum, as a measure of macrophageactivation. In contrast, T lymphocytes in the lungs of M. tu-berculosis-infected TNFa-deficient mice failed to migratefrom perivascular and peribronchial locations into granulo-mas in the peripheral regions of the lungs. The analysis of therole of LTa necessitated the study of bone-marrow chimeramice, since LTa and LTb are essential for establishing sec-ondary lymphoid organs, including lymph nodes. When irra-diated RAG1–/– mice were reconstituted with bone-marrowfrom LTa–/– mice, the chimeras exhibited a defective re-sponse to M. tuberculosis that closely resembled that ofTNFa–/– mice [53]. Chimeric mice with LTa–/– hematopoi-etic cells died between days 28 and 38 after low-dose aerosolinfection with M. tuberculosis and had higher bacterial loadsin the lungs than chimeras reconstituted with LTb–/– or wild-type marrow. There were no differences in the rate of recruit-ment or number of CD4+ or CD8+ T cells recruited to thelungs, no difference in the induction of TNFa, and no differ-
155W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158

ence in serum nitrite concentrations. As seen in TNFa–/–
mice, T lymphocytes in the lung were clustered near bloodvessels and airways in LTa–/– chimeras, while they migratedinto the clusters of macrophages in granulomas in LTb–/– andwild-type chimeras. These results together imply that TNFaand LTa have similar (but not redundant) roles in inducingthe migration of activated T lymphocytes to the immediatesite of infection with M. tuberculosis, after T lymphocytesare recruited to the general area by TNFa- and LTa-independent mechanisms. Whether the effects of TNFa andLTa are exerted through the induction or activation of spe-cific chemokines, chemokine receptors, or adhesion proteinsawaits further analysis.
6. Conclusion and directions for future study
The studies reported to date provide a strong foundationfor understanding the mechanisms of immune cell recruit-ment to sites of infection with M. tuberculosis, and provide abackground for additional studies. Among the topics of im-mediate interest are the identification of chemokines andchemokine receptors that direct CD4+ and CD8+ T cells tothe lung in tuberculosis, once an adaptive immune responsehas been initiated. The use of gene expression microarrayanalyses, together with the rapidly increasing availability ofantibodies to chemokine receptors as well as chemokine andchemokine receptor knockout mice, should facilitate identi-fication of the signals and mediators involved in T-cell re-cruitment in tuberculosis. Additional studies can also deter-mine the roles of specific chemokines and receptors requiredfor the maintenance of a local immune response at the site ofchronic infection. Moreover, studies in human subjects canidentify the molecules expressed by macrophages and lym-phocytes in the lungs of individuals with active tuberculosis,and can allow a comparison with those found in the periph-eral blood and at the sites of cutaneous reactions to PPD skintests.
While an approach to studying the mechanisms of recruit-ment to sites of infection in tuberculosis can be generallypatterned after the studies that identify molecules involved inother immune responses, the fact that immune cells areorganized in granulomas in tuberculous lesions should not beforgotten. Granulomas, with a characteristic pattern of cellu-lar architecture, clearly require specific signals for their es-tablishment and maintenance, and a challenge for futurestudies will be to identify the mechanisms involved in thisarchitecture. As the tools become available, conceptual ap-proaches to using them to discern the mechanisms of theimmune response to tuberculosis should reveal a high-resolution view of the effective and ineffective immune re-sponses that lead to the control of infection without its eradi-cation. One can also hope that this higher-resolution viewwill yield clues to the development of more effective vac-cines for tuberculosis, one of the major scourges afflicting thehuman population.
Acknowledgements
We thank Dr. Edward C. Klatt (Florida State UniversityCollege of Medicine) for permission to reproduce the photo-micrograph in Fig. 1, and Drs. Peter Seiler, Stefan Kauf-mann, Jaklien Leemans, and Tom van der Poll for allowing usto cite their results in advance of publication. This study hasbeen supported in part by grants from the NIH (HL51992 andAI46097) and the Sandler Family Foundation.
References
[1] J.L. Flynn, J. Chan, Immunology of tuberculosis, Annu. Rev. Immu-nol. 19 (2001) 93–129.
[2] M. Gonzalez-Juarrero, O.C. Turner, J. Turner, P. Marietta,J.V. Brooks, I.M. Orme, Temporal and spatial arrangement of lympho-cytes within lung granulomas induced by aerosol infection withMycobacterium tuberculosis, Infect. Immun. 69 (2001) 1722–1728.
[3] W. Peters, H.M. Scott, H.F. Chambers, J.L. Flynn, I.F. Charo,J.D. Ernst, Chemokine receptor 2 serves an early and essential role inresistance to Mycobacterium tuberculosis, Proc. Natl. Acad. Sci. USA98 (2001) 7958–7963.
[4] P.S. Randhawa, Lymphocyte subsets in granulomas of humantuberculosis: an in situ immunofluorescence study using monoclonalantibodies, Pathology 22 (1990) 153–155.
[5] T. Mogues, M.E. Goodrich, L. Ryan, R. LaCourse, R.J. North, Therelative importance of T cell subsets in immunity and immunopathol-ogy of airborne Mycobacterium tuberculosis infection in mice, J. Exp.Med. 193 (2001) 271–280.
[6] A.M. Cooper, D.K. Dalton, T.A. Stewart, J.P. Griffin, D.G. Russell,I.M. Orme, Disseminated tuberculosis in interferon gamma gene-disrupted mice, J. Exp. Med. 178 (1993) 2243–2247.
[7] J.L. Flynn, J. Chan, K.J. Triebold, D.K. Dalton, T.A. Stewart,B.R. Bloom, An essential role for interferon gamma in resistance toMycobacterium tuberculosis infection, J. Exp. Med. 178 (1993)2249–2254.
[8] R.J. North, Mice incapable of making IL-4 or IL-10 display normalresistance to infection with Mycobacterium tuberculosis, Clin. Exp.Immunol 113 (1998) 55–58.
[9] B.M. Saunders, A.A. Frank, I.M. Orme, A.M. Cooper, Interleukin-6induces early gamma interferon production in the infected lung but isnot required for generation of specific immunity to Mycobacteriumtuberculosis infection, Infect. Immun. 68 (2000) 3322–3326.
[10] S.I. Behr-Perst, M.E. Munk, T. Schaberg, T. Ulrichs, R.J. Schulz,S.H. Kaufmann, Phenotypically activated gamma delta T lympho-cytes in the peripheral blood of patients with tuberculosis, J. Infect.Dis. 180 (1999) 141–149.
[11] Y. Shen, D. Zhou, L. Qiu, X. Lai, M. Simon, L. Shen, Z. Kou,Q. Wang, L. Jiang, J. Estep, R. Hunt, M. Clagett, P.K. Sehgal, Y. Li,X. Zeng, C.T. Morita, M.B. Brenner, N.L. Letvin, Z.W. Chen, Adap-tive immune response of Vgamma2Vdelta2+ T cells during mycobac-terial infections, Science 295 (2002) 2255–2258.
[12] C.H. Ladel, C. Blum, A. Dreher, K. Reifenberg, S.H. Kaufmann,Protective role of gamma/delta T cells and alpha/beta T cells intuberculosis, Eur. J. Immunol. 25 (1995) 2877–2881.
[13] C.D. D’Souza, A.M. Cooper, A.A. Frank, R.J. Mazzaccaro,B.R. Bloom, I.M. Orme, An anti-inflammatory role for gamma delta Tlymphocytes in acquired immunity to Mycobacterium tuberculosis, J.Immunol. 158 (1997) 1217–1221.
[14] C.M. Johnson, A.M. Cooper, A.A. Frank, C.B. Bonorino, L.J. Wysoki,I.M. Orme, Mycobacterium tuberculosis aerogenic rechallenge infec-tions in B cell-deficient mice, Tuber Lung Dis. 78 (1997) 257–261.
156 W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158

[15] C.M. Bosio, D. Gardner, K.L. Elkins, Infection of B cell-deficientmice with CDC 1551, a clinical isolate of Mycobacteriumtuberculosis: delay in dissemination and development of lung pathol-ogy, J. Immunol 164 (2000) 6417–6425.
[16] P. Seiler, P. Aichele, B. Raupach, B. Odermatt, U. Steinhoff,S.H. Kaufmann, Rapid neutrophil response controls fast-replicatingintracellular bacteria but not slow-replicating Mycobacterium tuber-culosis, J. Infect. Dis 181 (2000) 671–680.
[17] A.A. Chackerian, J.M. Alt, T.V. Perera, C.C. Dascher, S.M. Behar,Dissemination of Mycobacterium tuberculosis is influenced by hostfactors and precedes the initiation of T-cell immunity, Infect. Immun.70 (2002) 4501–4509.
[18] L.E. Bermudez, F.J. Sangari, P. Kolonoski, M. Petrofsky, J. Goodman,The efficiency of the translocation of Mycobacterium tuberculosisacross a bilayer of epithelial and endothelial cells as a model of thealveolar wall is a consequence of transport within mononuclearphagocytes and invasion of alveolar epithelial cells, Infect. Immun. 70(2002) 140–146.
[19] R.F. Silver, Q. Li, W.H. Boom, J.J. Ellner, Lymphocyte-dependentinhibition of growth of virulent Mycobacterium tuberculosis H37Rvwithin human monocytes: requirement for CD4+ T cells in purifiedprotein derivative-positive, but not in purified protein derivative-negative subjects, J. Immunol. 160 (1998) 2408–2417.
[20] K.J. Brill, Q. Li, R. Larkin, D.H. Canaday, D.R. Kaplan, W.H. Boom,R.F. Silver, Human natural killer cells mediate killing of intracellularMycobacterium tuberculosis H37Rv via granule-independent mecha-nisms, Infect. Immun. 69 (2001) 1755–1765.
[21] P. Guermonprez, J. Valladeau, L. Zitvogel, C. Thery, S. Amigorena,Antigen presentation and T cell stimulation by dendritic cells, Annu.Rev. Immunol. 20 (2002) 621–667.
[22] K.A. Bodnar, N.V. Serbina, J.L. Flynn, Fate of Mycobacterium tuber-culosis within murine dendritic cells, Infect. Immun. 69 (2001)800–809.
[23] R.A. Henderson, S.C. Watkins, J.L. Flynn, Activation of human den-dritic cells following infection with Mycobacterium tuberculosis, J.Immunol. 159 (1997) 635–643.
[24] M. Gonzalez-Juarrero, I.M. Orme, Characterization of murine lungdendritic cells infected with Mycobacterium tuberculosis, Infect.Immun. 69 (2001) 1127–1133.
[25] S.P. Hickman, J. Chan, P. Salgame, Mycobacterium tuberculosisinduces differential cytokine production from dendritic cells and mac-rophages with divergent effects on naive T cell polarization, J. Immu-nol. 168 (2002) 4636–4642.
[26] A. Zlotnik, O. Yoshie, Chemokines: a new classification system andtheir role in immunity, Immunity 12 (2000) 121–127.
[27] M.D. Gunn, V.N. Ngo, K.M. Ansel, E.H. Ekland, J.G. Cyster,L.T. Williams, A B-cell-homing chemokine made in lymphoid fol-licles activates Burkitt’s lymphoma receptor-1, Nature 391 (1998)799–803.
[28] R. Phillips, A. Ager, Activation of pertussis toxin-sensitive CXCL12(SDF-1) receptors mediates transendothelial migration of T lympho-cytes across lymph node high endothelial cells, Eur. J. Immunol. 32(2002) 837–847.
[29] F. Sallusto, A. Lanzavecchia, C.R. Mackay, Chemokines and chemok-ine receptors in T-cell priming and Th1/Th2-mediated responses,Immunol. Today 19 (1998) 568–574.
[30] L. Boring, J. Gosling, S.W. Chensue, S.L. Kunkel, R.V. Farese Jr,H.E. Broxmeyer, I.F. Charo, Impaired monocyte migration andreduced type 1 (Th1) cytokine responses in C–C chemokine receptor 2knockout mice, J. Clin. Invest. 100 (1997) 2552–2561.
[31] I. Kurth, K. Willimann, P. Schaerli, T. Hunziker, I. Clark-Lewis,B. Moser, Monocyte selectivity and tissue localization suggests a rolefor breast and kidney-expressed chemokine (BRAK) in macrophagedevelopment, J. Exp. Med. 194 (2001) 855–861.
[32] J.G. Cyster, V.N. Ngo, E.H. Ekland, M.D. Gunn, J.D. Sedgwick,K.M. Ansel, Chemokines and B-cell homing to follicles, Curr. Top.Microbiol. Immunol. 246 (1999) 87–92 (discussion 93).
[33] K. Willimann, D.F. Legler, M. Loetscher, R.S. Roos, M.B. Delgado,I. Clark-Lewis, M. Baggiolini, B. Moser, The chemokine SLC isexpressed in T cell areas of lymph nodes and mucosal lymphoidtissues and attracts activated T cells via CCR7, Eur. J. Immunol. 28(1998) 2025–2034.
[34] R. Forster, A. Schubel, D. Breitfeld, E. Kremmer, I. Renner-Muller,E. Wolf, M. Lipp, CCR7 coordinates the primary immune response byestablishing functional microenvironments in secondary lymphoidorgans, Cell 99 (1999) 23–33.
[35] Y. Zhang, M. Broser, H. Cohen, M. Bodkin, K. Law, J. Reibman,W.N. Rom, Enhanced interleukin-8 release and gene expression inmacrophages after exposure to Mycobacterium tuberculosis and itscomponents, J. Clin. Invest. 95 (1995) 586–592.
[36] M.I. Wickremasinghe, L.H. Thomas, J.S. Friedland, Pulmonary epi-thelial cells are a source of IL-8 in the response to Mycobacteriumtuberculosis: essential role of IL-1 from infected monocytes in aNF-?B-dependent network, J. Immunol. 163 (1999) 3936–3947.
[37] Y. Lin, J. Gong, M. Zhang, W. Xue, P.F. Barnes, Production ofmonocyte chemoattractant protein 1 in tuberculosis patients, Infect.Immun. 66 (1998) 2319–2322.
[38] M.I. Sadek, E. Sada, Z. Toossi, S.K. Schwander, E.A. Rich, Chemok-ines induced by infection of mononuclear phagocytes with mycobac-teria and present in lung alveoli during active pulmonary tuberculosis,Am. J. Respir. Cell Mol. Biol. 19 (1998) 513–521.
[39] J.J. Saukkonen, B. Bazydlo, M. Thomas, R.M. Strieter, J. Keane,H. Kornfeld, Beta-chemokines are induced by Mycobacterium tuber-culosis and inhibit its growth, Infect. Immun. 70 (2002) 1684–1693.
[40] A. Sauty, M. Dziejman, R.A. Taha, A.S. Iarossi, K. Neote, E.A. Gar-cia-Zepeda, Q. Hamid, A.D. Luster, The T cell-specific CXC chemok-ines IP-10, Mig, and I-TAC are expressed by activated human bron-chial epithelial cells, J. Immunol. 162 (1999) 3549–3558.
[41] B.J. Rutledge, H. Rayburn, R. Rosenberg, R.J. North, R.P. Gladue,C.L. Corless, B.J. Rollins, High level monocyte chemoattractantprotein-1 expression in transgenic mice increases their susceptibilityto intracellular pathogens, J. Immunol. 155 (1995) 4838–4843.
[42] B. Lu, B.J. Rutledge, L. Gu, J. Fiorillo, N.W. Lukacs, S.L. Kunkel,R. North, C. Gerard, B.J. Rollins, Abnormalities in monocyte recruit-ment and cytokine expression in monocyte chemoattractant protein1-deficient mice, J. Exp. Med. 187 (1998) 601–608.
[43] L. Gu, S. Tseng, R.M. Horner, C. Tam, M. Loda, B.J. Rollins, Controlof TH2 polarization by the chemokine monocyte chemoattractantprotein-1, Nature 404 (2000) 407–411.
[44] S. Gasperini, M. Marchi, F. Calzetti, C. Laudanna, L. Vicentini,H. Olsen, M. Murphy, F. Liao, J. Farber, M.A. Cassatella, Geneexpression and production of the monokine induced by IFN-gamma(MIG), IFN-inducible T cell alpha chemoattractant (I-TAC), and IFN-gamma-inducible protein-10 (IP-10) chemokines by human neutro-phils, J. Immunol. 162 (1999) 4928–4937.
[45] C.M. Johnson, A.M. Cooper, A.A. Frank, I.M. Orme, Adequateexpression of protective immunity in the absence of granuloma for-mation in Mycobacterium tuberculosis-infected mice with a disrup-tion in the intracellular adhesion molecule 1 Gene, Infect. Immun. 66(1998) 1666–1670.
[46] B.M. Saunders, A.A. Frank, I.M. Orme, Granuloma formation isrequired to contain bacillus growth and delay mortality in micechronically infected with Mycobacterium tuberculosis, Immunology98 (1999) 324–328.
[47] C. Hu, T. Mayadas-Norton, K. Tanaka, J. Chan, P. Salgame, Mycobac-terium tuberculosis infection in complement receptor 3-deficientmice, J. Immunol. 165 (2000) 2596–2602.
[48] C.M. McKee, M.B. Penno, M. Cowman, M.D. Burdick, R.M. Strieter,C. Bao, P.W. Noble, Hyaluronan (HA) fragments induce chemokinegene expression in alveolar macrophages. The role of HA size andCD44, J. Clin. Invest. 98 (1996) 2403–2413.
157W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158

[49] C.G. Feng, A.G.D. Bean, H. Hooi, H. Briscoe, W.J. Britton, Increasein gamma interferon-secreting CD8+, as well as CD4+, T cells inlungs following aerosol infection with Mycobacterium tuberculosis,Infect. Immun. 67 (1999) 3242–3247.
[50] C.G. Feng, W.J. Britton, U. Palendira, N.L. Groat, H. Briscoe,A.G. Bean, Up-regulation of VCAM-1 and differential expansion ofbeta integrin-expressing T lymphocytes are associated with immunityto pulmonary Mycobacterium tuberculosis infection, J. Immunol. 164(2000) 4853–4860.
[51] J.L. Flynn, M.M. Goldstein, J. Chan, K.J. Triebold, K. Pfeffer,C.J. Lowenstein, R. Schreiber, T.W. Mak, B.R. Bloom, Tumor necro-sis factor-alpha is required in the protective immune response againstMycobacterium tuberculosis in mice, Immunity 2 (1995) 561–572.
[52] A.G. Bean, D.R. Roach, H. Briscoe, M.P. France, H. Korner,J.D. Sedgwick, W.J. Britton, Structural deficiencies in granulomaformation in TNF gene-targeted mice underlie the heightened suscep-tibility to aerosol Mycobacterium tuberculosis infection, which is notcompensated for by lymphotoxin, J. Immunol. 162 (1999)3504–3511.
[53] D.R. Roach, H. Briscoe, B. Saunders, M.P. France, S. Riminton,W.J. Britton, Secreted lymphotoxin-alpha is essential for the controlof an intracellular bacterial infection, J. Exp. Med. 193 (2001)239–246.
158 W. Peters, J.D. Ernst / Microbes and Infection 5 (2003) 151–158





















![[Micro] mycobacterium tuberculosis](https://img.pdfslide.us/doc/110x75/55d6fc67bb61ebfa2a8b47ea/micro-mycobacterium-tuberculosis.jpg)





