Embed Size (px)
Citation preview

Linezolid-resistant Staphylococcus aureus strain 1128105, the first known clinical isolate 1
possessing the cfr multidrug resistance gene 2
3
Running title: cfr-positive S. aureus isolate 1128105 4
5
Jeffrey B. Locke1#, Douglas E. Zuill1, Caitlyn R. Scharn3, Jennifer Deane2, Daniel F. Sahm2, 6
Gerald A. Denys4, Richard V. Goering3, and Karen J. Shaw1 7
8
1Trius Therapeutics, Inc., 6310 Nancy Ridge Drive, Suite 105, San Diego, CA 92121 9 10 2Eurofins Global Central Laboratory, 14100 Park Meadow Drive, Suite 110, Chantilly, VA 11 20151 12 13 3Department of Medical Microbiology and Immunology, Creighton University School of 14 Medicine, 2500 California Plaza, Omaha, NE 68178 15 16 4Indiana University Health Pathology Laboratory, 350 West 11th Street, Room 6027B 17 Indianapolis, IN 46202 18 19
#Corresponding author: 20
Jeffrey B. Locke, Ph.D. 21
Cubist Pharmaceuticals, Inc. 22
6310 Nancy Ridge Drive, Suite 105 23
San Diego, CA 92121 24
Phone: (858) 452-0370 x226 25
Fax: (858) 452-0412 26
E-mail: [email protected] 27
Key words: cfr, linezolid, tedizolid, oxazolidinone, Staphylococcus aureus 28
AAC Accepts, published online ahead of print on 25 August 2014Antimicrob. Agents Chemother. doi:10.1128/AAC.03493-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ABSTRACT 29
The Cfr methyltransferase confers resistance to six classes of drugs which target the 30
peptidyl transferase center of the 50S ribosomal subunit, including some oxazolidinones such as 31
linezolid (LZD). The mobile cfr gene was identified in European veterinary isolates from the 32
late 1990s, although the earliest report of a clinical cfr-positive strain was 2005 Colombian 33
MRSA isolate CM05. Here, through retrospective analysis of LZDr clinical strains from a US 34
surveillance program, we identified a cfr-positive MRSA isolate, 1128105, from January 2005, 35
predating CM05 by 5 months. Molecular typing of 1128105 revealed a unique PFGE profile 36
most similar to USA100, spa type t002 and multilocus sequence type 5 (ST5). In addition to cfr, 37
LZD resistance in 1128105 is partially attributed to the presence of a single copy of the 23S 38
rRNA gene mutation T2500A. Transformation of the ~37 kb conjugative p1128105 cfr-bearing 39
plasmid from 1128105 into S. aureus ATCC 29213 background strains was successful in 40
recapitulating the Cfr antibiogram, as well as resistance to aminoglycosides and trimethoprim. A 41
7 kb cfr-containing region of p1128105 possessed sequence nearly identical to that found in the 42
Chinese veterinary Proteus vulgaris isolate PV-01 and in US clinical S. aureus isolate 1900, 43
although the presence of IS431-like sequences is unique to p1128105. The cfr gene environment 44
in this early clinical cfr-positive isolate has now been identified in Gram-positive and Gram-45
negative strains of clinical and veterinary origin and has been associated with multiple mobile 46
elements, highlighting the versatility of this multidrug resistance gene and its potential for further 47
dissemination. 48
49
50
51
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

52
INTRODUCTION 53
Linezolid (LZD) resistance predominantly occurs through structural alterations to the 54
oxazolidinone binding site in the 50S peptidyl transferase center (PTC) (1). These 55
conformational changes are the result of mutations in genes encoding 23S rRNA (2, 3) or 56
ribosomal proteins L3 and L4 (4, 5) or via posttranscriptional modification of 23S rRNA base 57
A2503 by the Cfr methyltransferase (6). Highly LZDr isolates have been identified that possess 58
both the cfr-gene and chromosomally-encoded mutations (7-10). 59
The horizontally-transferrable, plasmid-borne nature of cfr makes this resistance 60
determinant inherently more worrisome than chromosomally-encoded resistance mechanisms 61
that must arise independently and/or disseminate clonally (11, 12). Adding to the potential for 62
spread is the low fitness cost of this gene (13) and the broad spectrum of resistance conferred by 63
Cfr to drugs included in the PhLOPSA phenotype (phenicol, lincosamide, oxazolidinone, 64
pleuromutilin, and streptogramin A class antibiotics) (6, 12, 14) as well as 16-member-ring 65
macrolides (15). Within the oxazolidinone class there are differences in susceptibility to strains 66
possessing cfr depending on structural features at both ends of the molecule. The addition of D-67
ring systems that pick up additional binding interactions in the PTC and the substitution of an A-68
ring C-5 hydroxymethyl groups (in place of the bulkier acetamide-containing substituent found 69
on LZD and other oxazolidinones) allow oxazolidinones such as tedizolid (16) to retain greater 70
potency than LZD in the presence of Cfr methylation (17). 71
The cfr gene was first identified on the pSCFS1 plasmid in a Staphylococcus sciuri 72
isolate recovered from a florfenicol-treated calf in Bavaria in 1997 (18). Since then, a variety of 73
other veterinary staphylococci have been identified possessing cfr-bearing plasmids sharing 74
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

either similarity to pSCFS1 or to one of two other groups possessing regions containing the cfr 75
gene flanked by either IS256 or IS21-558 mobile elements (19). The first clinical cfr-positive 76
strains reported were Staphylococcus aureus isolates recovered in 2005 from Ireland and 77
Colombia. Irish MRSA isolate M05/0060 (no specific collection date given) (20) possessed the 78
cfr-bearing pSCFS7 plasmid which has similarity to IS21-558-carrying veterinary plasmid 79
pSCFS3 (21) although only a truncated portion of the IS21-558 element remained (22). 80
Colombian MRSA isolate CM05 was recovered in May of 2005 from a patient in Medellin who 81
had briefly undergone LZD therapy (2 doses) (6, 23). The cfr gene in CM05 was chromosomally 82
located and shared flanking sequence with high similarity to the pSM19035 Gram-positive 83
multidrug resistance plasmid, including an IS21-558 element (24, 25). Subsequently, additional 84
cfr-positive clinical staphylococci and enterococci have been recovered from diverse 85
geographical origins: United States (26), Belgium (27), Germany (28), France (29), Italy (30), 86
Spain (31), Mexico (32), Panama (33), Brazil (34), Thailand (35), Canada (36), India (37), and 87
China (38). The cfr gene has also been found in Gram-negative species including Escherichia 88
coli (39) and Proteus vulgaris (40), although in these species the Gram-negative IS26 element 89
appears to have been involved in cfr mobility (19). 90
To investigate the potential existence of clinical cfr-positive strains predating CM05 we 91
screened historical LZDr Gram-positive isolates that were collected as part of an ongoing 92
surveillance initiative conducted by Eurofins Medinet to monitor antimicrobial resistance trends 93
among key Gram-positive pathogens encountered across the USA. Among these was a 2005 94
LZDr MRSA isolate (strain # 1128105) that was selected for further phenotypic and genotypic 95
analysis. Here we characterize the genetic determinants conferring LZD resistance in 1128105, 96
describe the genetic environment of the cfr gene and assess its transmissibility. 97
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

98
CASE REPORT 99
In January 2005, a 36-year-old female patient with end-stage cystic fibrosis presented to 100
the emergency room (Indianapolis, IN) with increased cough, low grade fever (100.8 ºF), 101
decreased appetite, and left sided chest pain. Chest X-ray on admission showed acute left mid 102
lung airspace disease with left pleural effusion. The S. aureus respiratory isolate resistant to 103
linezolid (strain 1128105) recovered during the course of her hospitalization is described below. 104
The patient had numerous prior hospitalizations with a complicated history of end stage 105
cystic fibrosis, cystic fibrosis related diabetes, pulmonary hypertension, hemoptysis, hypoxemia, 106
and chronic renal failure. The patient was admitted to the ICU on 1-2-05 requiring oxygen and 107
Albuterol and Pulmozyme nebulizer treatments. She was also started on IV ceftazidime and 108
tobramycin because previous hospitalization cultures grew multi-drug resistant Pseudomonas 109
aeruginosa. The patient’s lung disease showed improvement on X-ray and she was transferred 110
to the floor on 1-4-05. Over a course of several days the patient’s pulmonary functions 111
worsened. Repeat X-rays showed chronic lung changes, and sputum cultures from 1-12-05 grew 112
P. aeruginosa and MRSA susceptible to linezolid (MIC 4 μg/ml). Oral linezolid was started on 113
1-15-05. The patient completed 13 days of oral linezolid and was continued on IV ceftazidime 114
and tobramycin throughout hospitalization. Repeat sputum cultures from 1-20-05 showed no 115
changes with both P. aeruginosa and MRSA present. The MIC of linezolid, however, was 116
elevated (16 μg/ml) from the previous culture on 1-12-05. The patient was stable on 1-28-05 and 117
discharged home on IV ceftazidime and tobramycin and Albuterol, colistin, and Pulmozyme 118
nebulizer treatments. 119
120
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

MATERIALS AND METHODS 121
Bacterial strains and culture conditions. S. aureus strains 1128105 (initially 122
designated as 3133832) (41), ATCC 29213, ATCC 29213 T2500A, RN4220 and transformants 123
thereof were cultured aerobically at 37°C on cation-adjusted Mueller-Hinton II agar (MHA, 124
Becton Dickinson, Franklin Lakes, NJ) or in MH broth (MHB). The ATCC 29213 T2500A 125
strain possesses a single copy (allele 4) of the 23S rRNA gene mutation T2500A (E. coli 126
numbering) and was selected as a spontaneous mutant with reduced LZD susceptibility in a 127
previous study (42). For conjugation experiments, RN4220 was transduced to novobiocin 128
resistance from S. aureus strain U9NO (43) as an additional strain selection marker. 129
Antimicrobial agents. Vancomycin (VAN), florfenicol (FFC), erythromycin (ERY), 130
clindamycin (CLI), tobramycin (TOB), trimethoprim (TMP), ciprofloxacin (CIP), oxacillin 131
(OXA), ceftazidime (CAZ), and novobiocin (NOV) were purchased from Sigma-Aldrich (St. 132
Louis, MO). Other compounds were obtained as follows: TZD (Trius Therapeutics, San Diego, 133
CA), LZD (ChemPacific Corp., Baltimore, MD), tiamulin (TIA, Wako Pure Chemical Industries, 134
Ltd., Richmond, VA), daptomycin (DAP, TSZ CHEM, Framingham, MA). All compounds were 135
prepared fresh in DMSO, ethanol or deionized water prior to use in MIC assays or selective 136
media. 137
MIC assays. MIC assays were performed via broth microdilution in accordance with 138
CLSI guidelines (44) with the exception that test compounds were made up at 50X concentration 139
(2 µl added to 98 µl of broth + cells) and that MIC values were determined visually through 140
detection with alamarBlue® (Invitrogen Corp., Carlsbad, CA) as previously described (45). 141
Enumeration of colony-forming units was performed by serially diluting bacteria in PBS and 142
plating on MHA. MIC assays containing DAP used MHB supplemented with 50 mg/l Ca2+. 143
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

Plasmid isolation and transformation. Plasmid DNA was isolated from 1128105 via 144
lysostaphin digestion and subsequent processing using a miniprep kit (Qiagen, Venlo, 145
Netherlands). Competent cell preparation and transformation of ATCC 29213 and RN4220 146
background strains was performed as previously described (46). Transformant cells were plated 147
on MHA containing 5 µg/ml of TIA. 148
PCR and plasmid sequencing. De novo plasmid sequencing of p1128105 was 149
performed using Illumina HiSeq (Genewiz, South Plainfield, NJ). 23S rRNA alleles from rrn 150
operons 1, 3, 4, and 5 were amplified by PCR using primers previously described (47). Allele 2 151
was amplified with primers rrn6_RF (reverse complement to rrn6_R (48, 49)) and rrn2_R (47) 152
and allele 6 was amplified using 16S_F and rrn6_R (48, 49). Genes encoding ribosomal proteins 153
L3 (rplC) and L4 (rplD) were amplified as previously described (42). Sequence data were 154
analyzed using Vector NTI Advance 11TM software (Invitrogen) and annotated based on 155
homology to sequence data from NCBI BLAST analyses (50). 156
Molecular typing. For pulsed-field gel electrophoresis (PFGE), chromosomal DNA was 157
prepared in agarose plugs, digested with SmaI restriction endonuclease, and analyzed as 158
previously described (51). Staphylococcal protein A (spa) typing and multi-locus sequence 159
typing (MLST) were also performed using published protocols (52, 53). 160
Filter mating. Conjugation experiments were performed by filter mating as described 161
previously (54). Briefly, overnight donor and recipients cultures were resuspended in 5 ml 0.5% 162
NaCl to an optical density (OD540) of 1.0. The suspensions were combined and collected on a 163
0.45 μm nitrocellulose filter which was incubated upright on a brain heart infusion (BHI) 164
(Becton, Dickinson and Company, Sparks, MD) agar plate overnight at 37°C. The filter was 165
resuspended in 10 ml BHI broth, vigorously vortexed, and the cells recovered by centrifugation 166
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

for 10 minutes at 2,700 × g. The resulting cell pellet was resuspended in 1.0 ml BHI broth and 167
plated on BHI agar containing 5 μg/ml TIA and 10 μg/ml NOV for transconjugant selection after 168
overnight incubation at 37°C. Resulting colonies were further subcultured twice on BHI agar 169
with TIA and NOV followed by PFGE analysis to confirm the transconjugant lineage. 170
Plasmid analysis and Southern hybridization. Plasmid DNA was isolated by the 171
method of Holmes and Quigley (55) and visualized on 0.8% SEAKems LE agarose gels 172
(LONZA, Rockland, ME) with electrophoresis for 12 hours at 6V/cm. A digoxigenin-labeled 173
full length cfr-specific 1,050 bp probe was synthesized using cfr_F/cfr_R primers (25) and the 174
Roche PCR DIG Probe Synthesis Kit (Roche Diagnostics, Mannheim, Germany). Southern 175
hybridization was performed essentially by the method of Sambrook and Russell (56) using the 176
Roche DIG Nucleic Acid Detection Kit following the manufacturer’s instructions. 177
Nucleotide sequence accession number. A 7,020 bp region of the p1128105 plasmid 178
containing the cfr gene has been deposited in the NCBI database under the GenBank accession 179
number KJ866414. 180
181
RESULTS 182
1128105 has a molecular profile most similar to USA100. Molecular analysis of S. aureus 183
strain 1128105 revealed a PFGE banding pattern most similar to that of healthcare-associated 184
MRSA USA100 (88% related to USA100 by UPGMA/Dice Coefficient) (Fig. 1). Results of spa 185
typing (i.e., t002) and MLST (clonal complex 5, sequence type 5) were also consistent with this 186
relationship (57). 187
The 1128105 antibiogram is consistent with the presence of cfr. Sequence analysis of LZD 188
resistance determinants in 1128105 (cfr, 23S rRNA, rplC, rplD) identified the presence of the cfr 189
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

gene and a single copy of the T2500A 23S rRNA gene mutation (allele 4). T2500A has been 190
previously associated with reduced susceptibility to oxazolidinones (42, 48) and a single copy of 191
this mutation is consistent with a TZD MIC value of 1 μg/ml in S. aureus (42). The 1128105 192
antibiogram coincides with the PhLOPSA phenotype conferred by Cfr, including resistance to 193
LZD, FFC, TIA, and CLI (Table 1) (14, 17). Additionally, 1128105 demonstrated resistance to 194
TOB, TMP, CIP, ERY, OXA, and CAZ. Of the drugs tested, 1128105 was only susceptible to 195
VAN and DAP. 196
The 1128105 cfr gene is plasmid-borne. To determine whether the cfr gene was located on a 197
plasmid or the chromosome, total 1128105 plasmid DNA was transformed into the S. aureus 198
ATCC 29213 wild-type and 23S rRNA T2500A mutant backgrounds and plated on selective 199
media containing TIA. The presence of TIAr colonies that were PCR-positive for cfr suggested 200
that the cfr gene was plasmid-borne. The transformant strains recapitulated the MIC profile of 201
1128105 for drugs falling within the Cfr resistance spectrum (i.e. LZD, CLI, TIA, and FFC) 202
(Table 1). In addition, no change in MIC was observed for TZD in either the S. aureus ATCC 203
29213 wild-type (TZD MIC 0.5 µg/ml) or ATCC 29213 23S rRNA T2500A (MIC 1 µg/ml) 204
strains transformed with cfr, consistent with previous findings (17). The resulting MIC values 205
for TZD and LZD (1 and 16 µg/ml, respectively) in the ATCC 29213 T2500A background were 206
identical to the combination of the cfr and T2500A resistance determinants in the 1128105 207
isolate. Southern hybridization confirmed the location of the cfr gene on a plasmid ca. 37 kb in 208
size in S. aureus strain 1128105 and RN4220 + p1128105 transconjugants (Fig. 2, lanes 2-3 and 209
6-7, respectively). Transformation of p1128105 in the ATCC 29213 wild-type and T2500A 210
backgrounds also conferred resistance to TMP and TOB (Table 1), likely accounted for by the 211
dfrA and aacA-aphD genes identified in other de novo plasmid sequencing contigs. 212
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

The p1128105 cfr plasmid is conjugative. Filter mating experiments demonstrated conjugal 213
transfer of the cfr plasmid (p1128105) from S. aureus strain 1128105 to recipient RN4220 at a 214
frequency ranging from 1.0 × 10-8 to 4.0 × 10-9 transconjugates per recipient cell. Agarose gel 215
electrophoresis of plasmid preparations from transconjugants confirmed the presence of 216
p1128105 (data not shown). 217
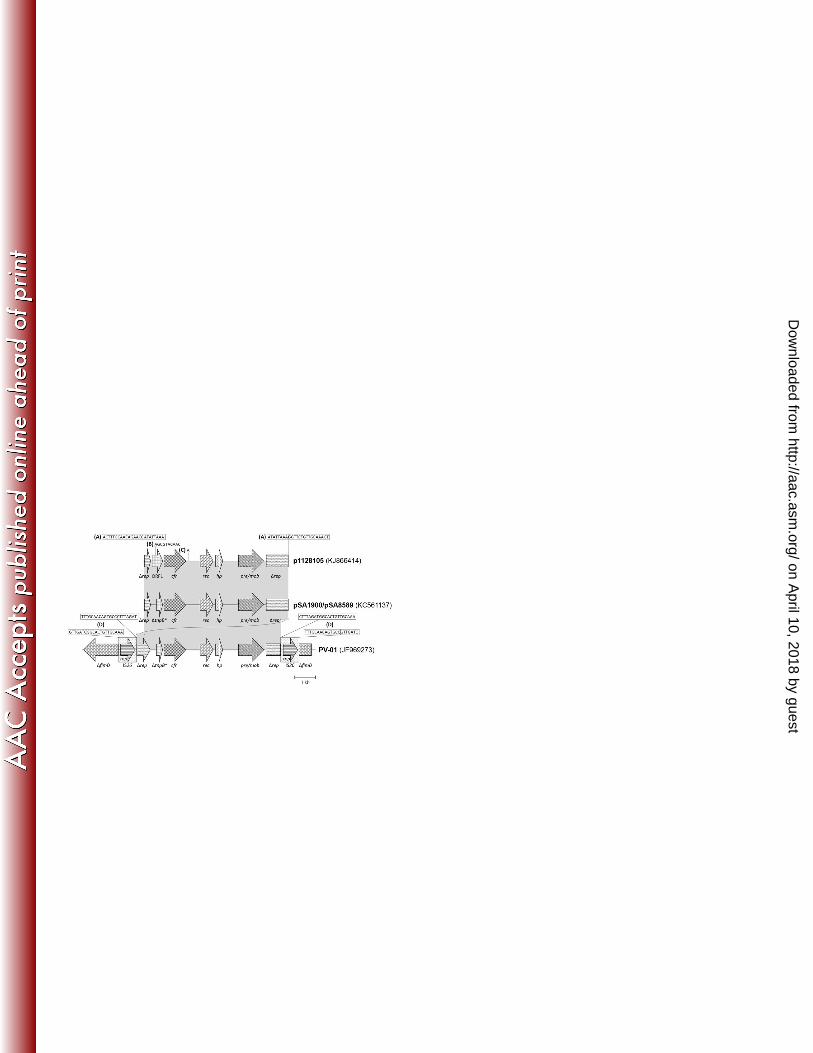
The p1128105 cfr gene environment has high similarity to those found in P. vulgaris PV-01 218
and S. aureus 1900 isolates. A 7,020 bp contig containing the cfr gene was identified from de 219
novo sequencing of 1128105 total plasmid DNA. This 7 kb region of p1128105 was nearly 220
identical to that found chromosomally in the Chinese PV-01 P. vulgaris veterinary isolate 221
(GenBank accession no. JF969273) (40) and in the pSA1900 plasmid of the recently described 222
clinical S. aureus isolate 1900 recovered from an Ohio hospital in during a 2006-2007 cfr 223
outbreak (GenBank accession no. KC561137; deposited as “pSA8589”) (58) (Fig. 3). In both 224
1128105 and 1900, the cfr gene is plasmid-borne whereas it is chromosomally located in PV-01, 225
although it is thought to have arisen through integration of a cfr-bearing plasmid via 226
recombination between an IS26 element present in a plasmid and one inserted into the 227
chromosomal fimD gene (40). 228
The p1128105 sequence contains two internal sequence discrepancies with PV-01 and 229
pSA1900/pSA8589: 1) 5’-AGCGTACAAC-3’ insertion in p1128105 311 bp upstream from cfr 230
not present in pSA1900/pSA8589 or PV-01 (Fig. 3B), and 2) an “A” present in p1128105 and 231
PV-01 58 bp downstream of cfr but not found in pSA1900/pSA8589 (Fig. 3C). The 5’-232
AGCGTACAAC-3’ insertion within the 300-bp “∆tnpB” ORF of PV-01 and pSA1900/pSA8589 233
results in the generation of a 513 bp ORF in p1128105. This ORF, as well as the “∆tnpB” ORFs 234
of PV-01 and pSA1900/pSA8589, however lack any significant sequence similarity with the 235
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

tnpB gene of Tn554 (GenBank accession no. X03216) (59). The p1128105 ORF shares 97% 236
sequence identity with the 226 bp 5’ portion of the 357 bp “∆tnpB” ORF of the pSCFS1 cfr 237
plasmid (GenBank accession no. AJ579365) (60), although this similarity ends around the 3’ 140 238
bp of pSCFS1 ∆tnpB sequence which has 100% nucleotide identity with the 3’ end of the Tn554 239
tnpB gene, hence this ORF has been designated simply as a hypothetical protein (“ORF1”) in 240
p1128105. The p1128105 ORF1 sequence, including the 5’-AGCGTACAAC-3’ insertion, also 241
has high similarity with sequences found in multidrug resistance plasmids pKKS825 from S. 242
aureus (GenBank accession no. FN377602) (61) and pDB2011 from Listeria innocua (GenBank 243
accession no. KC456362) (62). 244
Both PV-01 and p1128105 cfr environments contain the ORF1-cfr-rec-hp-pre/mob gene 245
insertion within identical rep gene sequences; however the rep gene is disrupted in different 246
locations. In p1128105 insertion of a mobile element resulted in the direct duplication of the 5’-247
ATATTAAA-3’ insertion site sequence within rep which is flanked by the 16-bp perfect 248
terminal inverted repeats (TIRs) (5’-ACTTTGCAACAGAACC-3’ and 5’-249
GGTTCTGTTGCAAAGT-3’), while PV-01 contains a direct duplication of the 5’-CTTTAGAT-250
3’ rep insertion site sequence flanked by IS26 14-bp TIRs (5’-TTTGCAACAGTGCC-3’ and 5’-251
GGCACTGTTGCAAA-3’) (Fig. 3A, D) (40, 63). The p1128105 TIRs as well as flanking 252
sequence of the contig both upstream (5’-TTTT-3’) and downstream (5’-TGA-3’) of the 5’ and 253
3’ TIRs are identical to sequences of IS431 (64, 65). The pSA1900/pSA8589 plasmid also 254
shares the conserved ORF1-cfr-rec-hp-pre/mob genetic composition, however this plasmid 255
contains an uninterrupted rep gene. Two additional, larger cfr-bearing plasmids are present in S. 256
aureus 1900 and it is unknown how this 7 kb plasmid may integrate into those plasmids and 257
whether such integrations may involve mobile elements inserting within rep (58). 258
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

259
DISCUSSION 260
Linezolid-resistant S. aureus 1128105 represents the earliest documented cfr-positive 261
clinical isolate, predating CM05 by 5 months. Of the three 2005 cfr clinical isolates described to 262
date (CM05, M05/0060, and 1128105,) all have originated from geographically-distinct regions 263
and possess unique cfr gene environments. The cfr gene in CM05 is found in a pSM19035-like 264
chromosomally-integrated region (25); the cfr environment of M05/0060 is a pSCFS3-like 265
background (22); while here, we show that the p1128105 cfr-flanking sequence is highly similar 266
to the cfr gene chromosomal insertion in P. vulgaris (40) and the pSA1900/pSA8589 plasmid of 267
S. aureus 1900 (58), although unique in its association with IS431-like sequences. The IS431 268
element has only been found in one other cfr-bearing plasmid, pERGB (66), although it is an 269
element commonly found in SCCmec cassettes and Gram-positive plasmids (67). The TIRs of 270
IS431 are identical to those found in Gram-positive insertion sequences ISSau10 (68), IS257 (69) 271
and are highly similar to those of the Gram-negative IS26 element (63, 64). These associations 272
further support the emerging picture of cfr as a widely disseminated multidrug resistance drug 273
gene found in a diverse set of genetic environments in a wide variety of clinical and veterinary 274
pathogens (19). The ongoing use of drug classes such as phenicols and pleuromutilins in 275
veterinary medicine (i.e. florfenicol, tiamulin, valnemulin) will provide continued positive 276
selective pressure for cfr and increases the likelihood of transfer of this gene into clinical strains 277
where other members of these drug classes (i.e. chloramphenicol, retapamulin) are used in 278
human medicine. 279
Analysis of the remainder of p1128105 sequence could provide further clues about the 280
origins of early clinical cfr plasmids and the likely transfer from genetic environments found in 281
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

veterinary pathogens to those found in clinical pathogens and/or transfer between Gram-positive 282
and Gram-negative species. Although the first documented occurrence of cfr was in a veterinary 283
isolate of S. sciuri, there have not been worldwide longitudinal surveys of veterinary or 284
environmental isolates to identify the origin or comprehensive species distribution of the gene in 285
non-clinical settings. Therefore, the abundance of cfr in P. vulgaris, and whether the gene 286
originated from or was transferred to staphylococci, is still unknown. More conclusive 287
documentation of the early dissemination of cfr however will perhaps require a different strategic 288
approach than was taken here (analyzing linezolid-resistant strains) since: 1) a recent report that 289
describes an E. faecalis veterinary isolate that failed to confer the typical Cfr antibiogram profile 290
likely due to unknown strain factors, as the gene itself was still transcribed and translated (70), 2) 291
LZD surveillance studies in recent years that have identified LZDs cfr-positive S. aureus clinical 292
isolates (MIC=4 µg/ml) (71, 72), and 3) the identification of cfr in Gram-negative species (39, 293
40) which are intrinsically resistant to some drugs within the Cfr resistance spectrum, such as 294
oxazolidinones and pleuromutilins. Identifying potential pre-2005 clinical cfr isolates would 295
therefore benefit from the use of MIC-independent genotypic screening strategies. 296
Although the strain was not retained, the S. aureus isolate collected from the patient on 1-297
12-05, 3 days prior to starting LZD therapy, had a LZD MIC value of 4 µg/mL, which is 298
consistent with the MIC of the ATCC 29213 background single-copy 23S rRNA T2500A mutant 299
strain used in this study and could represent a cfr-negative background of 1128105 or one in 300
which the cfr gene was not expressed. The subsequent therapeutic regiment containing LZD and 301
TOB would have provided dual selective pressure for the acquisition and/or expression of the 302
cfr-containing multidrug resistance plasmid p1128105. The conjugative potential of p1128105 303
adds credence to the horizontal transmissibility of this particular cfr plasmid. It is interesting to 304
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

note that the 1900 S. aureus isolate, which also possesses a plasmid-borne P. vulgaris-like cfr 305
gene environment, was isolated as part of an outbreak of cfr strains from 2006-2007 in a 306
bordering state (Ohio). 1900, like 1128105, was also multilocus sequence type 5, spa type t002 307
and has a PFGE profile most similar to USA100 (58). 308
Similarly to 1128105, the cfr gene has been found in a large number of strains with co-309
occurring chromosomal mutations in 23S rRNA and/or ribosomal protein L3 and L4 mutations 310
that selectively confer reduced susceptibility to oxazolidinones (8, 10, 73). The presence of cfr 311
in strains with such background mutations may favor the propagation of these strains even in the 312
absence of selective pressure from LZD, since any member of the 6 classes of agents falling 313
within the Cfr resistance spectrum could select for its retention. As demonstrated with previous 314
structure-activity relationship studies, MIC values of TZD were maintained in the presence of cfr 315
likely due to the presence of a potency-adding D-ring system and reduced steric interaction of 316
the methylated A2503 23S rRNA with the hydroxymethyl A-ring C-5 substituent of TZD, 317
relative to the more bulky C-5 acetamide-containing group of linezolid (17). 318
The expansive drug resistance profile of 1128105 highlights the importance of continuing 319
to develop Gram-positive agents which maintain activity against a wide variety of resistance 320
mechanisms. VAN and DAP were the only two anti-staphylococcal agents tested in this study to 321
which 1128105 remained susceptible. The β-lactam and aminoglycoside resistance mechanisms 322
in this strain additionally conferred resistance to broad spectrum agents such as ceftazidime and 323
tobramycin used to target the patient’s co-occurring P. aeruginosa infection. Although it is the 324
earliest clinical cfr-positive isolate described to date, 1128105 possesses a diverse set of 325
mechanisms capable of evading many contemporary antimicrobial therapeutics. 326
327
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ACKNOWLEDGEMENTS 328
Portions of this work were presented at the 23rd European Congress of Clinical Microbiology 329
and Infectious Diseases (41). This study was funded in part by Trius Therapeutics. Jeffrey B. 330
Locke is a current employee of Cubist Pharmaceuticals. Douglas E. Zuill and Karen J. Shaw 331
were employees of Trius Therapeutics at the time this study was conducted. All authors had full 332
access to the data. The authors had final responsibility for the decision to submit for publication. 333
334
TABLES AND FIGURES 335
Table 1. MIC profiling of S. aureus 1128105 and ATCC 29213 transformants possessing the 336
p1128105 cfr-bearing plasmid 337
338
TZD, tedizolid; LZD, linezolid; FFC, florfenicol; TIA, tiamulin; CLI, clindamycin; TOB, 339
tobramycin; TMP, trimethoprim; CIP, ciprofloxacin; ERY, erythromycin; OXA, oxacillin; CAZ, 340
ceftazidime; VAN, vancomycin; DAP, daptomycin. 341
342
343
FIG 1 PFGE profile comparison of 1128105 with USA100 and USA800 reference strains. 344
345
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

346
FIG 2 Southern blot analysis of cfr in 1128105 and the RN4220 p1128105 transformant. 347
Agarose gel electrophoresis of plasmid preparations (A) and Southern hybridization analysis 348
using a cfr-specific probe (B) with S. aureus strains 1128105 (lanes 2 and 3), RN4220 recipients 349
(lanes 4 and 5) and TIAr/NOVr RN4220 p1128105 transconjugants (lanes 6 and 7) (see text). S. 350
aureus USA300 strain FPR3757 (lane 1) (74) was included as a plasmid size reference. To 351
demonstrate reproducibility, donor and recipient isolates were analyzed in duplicate lanes. 352
353
354
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

355
FIG 3 Comparative analysis of the p1128105 cfr locus. Alignment of cfr gene environments 356
from S. aureus 1128105 plasmid p1128105 (7,020 bp), S. aureus 1900 plasmid 357
pSA1900/pSA8589 (6,962 bp) (58), and P. vulgaris PV-01 chromosomal sequence (11,228 bp) 358
(40) reveal almost 7 kb of near 100% identity (gray shading). Direct repeats (DRs) and adjacent 359
terminal inverted repeats (TIRs) are shown in boxes. Key differentiating sequence features 360
among the three cfr loci include: (A) rep gene 8-bp direct repeats in p1128105 at the site of 361
insertion of a IS431-like element flanked by 16-bp TIRs, (B) 10-bp insertion in p1128105 but not 362
present in pSA1900/pSA8589 or PV-01, (C) a single adenine base present in p1128105 and PV-363
01 but not in pSA1900/pSA8589, (D) 8-bp DRs in the PV-01 rep gene and 8-bp fimD DRs at the 364
site of insertion of IS26 and its 14-bp TIRs. The ORFs located upstream of cfr in 365
pSA1900/pSA8589 and PV-01 are labeled here as “ΔtnpB*” rather than their “∆tnpB” 366
designation in GenBank due to a lack of sequence similarity with the tnpB gene of Tn554 367
(GenBank accession no. X03216) (59). In p1128105, the 5’-AGCGTACAAC-3’ sequence 368
present upstream of cfr extends this putative ORF (“ORF1”), which has been designated as a 369
hypothetical protein, beyond the length of the ΔtnpB* ORFs of pSA1900/pSA8589 and PV-01. 370
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

The pSA1900/pSA8589 sequence is a complete circular plasmid that was linearized here for 371
illustrative purposes by splitting the rep gene to optimize alignment with the linear p1128105 372
and PV-01 sequences. 373
374
REFERENCES 375
1. Long KS, Vester B. 2012. Resistance to linezolid caused by modifications at its binding 376 site on the ribosome. Antimicrob. Agents Chemother. 56:603-612. 377
2. Gonzales RD, Schreckenberger PC, Graham MB, Kelkar S, DenBesten K, Quinn 378 JP. 2001. Infections due to vancomycin-resistant Enterococcus faecium resistant to 379 linezolid. Lancet 357:1179. 380
3. Tsiodras S, Gold HS, Sakoulas G, Eliopoulos GM, Wennersten C, Venkataraman L, 381 Moellering RC, Ferraro MJ. 2001. Linezolid resistance in a clinical isolate of 382 Staphylococcus aureus. Lancet 38:207-208. 383
4. Locke JB, Hilgers M, Shaw KJ. 2009. Mutations in ribosomal protein L3 are associated 384 with oxazolidinone resistance in staphylococci of clinical origin. Antimicrob. Agents 385 Chemother. 53:5275-5278. 386
5. Wolter N, Smith AM, Farrell DJ, Schaffner W, Moore M, Whitney CG, Jorgensen 387 JH, Klugman KP. 2005. Novel mechanism of resistance to oxazolidinones, macrolides, 388 and chloramphenicol in ribosomal protein L4 of the pneumococcus. Antimicrob. Agents 389 Chemother. 49:3554-3557. 390
6. Toh SM, Xiong L, Arias CA, Villegas MV, Lolans K, Quinn J, Mankin AS. 2007. 391 Acquisition of a natural resistance gene renders a clinical strain of methicillin-resistant 392 Staphylococcus aureus resistant to the synthetic antibiotic linezolid. Mol. Microbiol. 393 64:1506-1514. 394
7. Cercenado E, Marin M, Insa R, Bouza E. 2010. Abstr. 20th Euro. Cong. Clin. Micro. 395 Infect. Dis., abstr P979. 396
8. Locke JB, Morales G, Hilgers M, GC K, Rahawi S, Picazo JJ, Shaw KJ, Stein JL. 397 2010. Elevated linezolid resistance in clinical Staphylococcus aureus cfr isolates is 398 associated with co-occurring mutations in ribosomal protein L3. Antimicrob. Agents 399 Chemother. 400
9. Bonilla H, Huband MD, Seidel J, Schmidt H, Lescoe M, McCurdy SP, Lemmon 401 MM, Brennan LA, Tait-Kamradt A, Puzniak L, Quinn JP. 2010. Multicity outbreak 402 of linezolid-resistant Staphylococcus epidermidis associated with clonal spread of a cfr-403 containing strain. Clin. Infect. Dis. 51:796-800. 404
10. LaMarre J, Mendes RE, Szal T, Schwarz S, Jones RN, Mankin AS. 2013. The 405 genetic environment of the cfr gene and the presence of other mechanisms account for the 406 very high linezolid resistance of the Staphylococcus epidermidis isolate 426-3147L. 407 Antimicrob. Agents Chemother. 57:1173-1179. 408
11. Kehrenberg C, Cuny C, Strommenger B, Schwarz S, Witte W. 2009. Methicillin-409 resistant and -susceptible Staphylococcus aureus strains of clonal lineages ST398 and 410
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ST9 from swine carry the multidrug resistance gene cfr. Antimicrob. Agents Chemother. 411 53:779-781. 412
12. Kehrenberg C, Schwarz S, Jacobsen L, Hansen LH, Vester B. 2005. A new 413 mechanism for chloramphenicol, florfenicol and clindamycin resistance: methylation of 414 23S ribosomal RNA at A2503. Mol. Microbiol. 57:1064-1073. 415
13. LaMarre JM, Locke JB, Shaw KJ, Mankin AS. 2011. Low fitness cost of the 416 multidrug resistance gene cfr. Antimicrob. Agents Chemother. 55:3714-3719. 417
14. Long KS, Poehlsgaard J, Kehrenberg C, Schwarz S, Vester B. 2006. The cfr rRNA 418 methyltransferase confers resistance to phenicols, lincosamides, oxazolidinones, 419 pleuromutilins, and streptogramin A antibiotics. Antimicrob. Agents Chemother. 420 50:2500-2505. 421
15. Smith LK, Mankin AS. 2008. Transcriptional and translational control of the mlr 422 operon, which confers resistance to seven classes of protein synthesis inhibitors. 423 Antimicrob. Agents Chemother. 52:1703-1712. 424
16. Im WB, Choi SH, Park JY, Finn J, Yoon SH. 2011. Discovery of torezolid as a novel 425 5-hydroxymethyl-oxazolidinone antibacterial agent. Eur. J. Med. Chem. 46:1027-1039. 426
17. Locke JB, Finn J, Hilgers M, Morales G, Rahawi S, G CK, Picazo JJ, Im W, Shaw 427 KJ, Stein JL. 2010. Structure-activity relationships of diverse oxazolidinones for 428 linezolid-resistant Staphylococcus aureus strains possessing the cfr methyltransferase 429 gene or ribosomal mutations. Antimicrob. Agents Chemother. 54:5337-5343. 430
18. Schwarz S, Werckenthin C, Kehrenberg C. 2000. Identification of a plasmid-borne 431 chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri. Antimicrob. 432 Agents Chemother. 44:2530-2533. 433
19. Shen J, Wang Y, Schwarz S. 2013. Presence and dissemination of the multiresistance 434 gene cfr in Gram-positive and Gram-negative bacteria. The Journal of antimicrobial 435 chemotherapy 68:1697-1706. 436
20. Rossney AS, Shore AC, Morgan PM, Fitzgibbon MM, O'Connell B, Coleman DC. 437 2007. The emergence and importation of diverse genotypes of methicillin-resistant 438 Staphylococcus aureus (MRSA) harboring the Panton-Valentine leukocidin gene (pvl) 439 reveal that pvl is a poor marker for community-acquired MRSA strains in Ireland. J. Clin. 440 Microbiol. 45:2554-2563. 441
21. Kehrenberg C, Schwarz S. 2006. Distribution of florfenicol resistance genes fexA and 442 cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob. Agents 443 Chemother. 50:1156-1163. 444
22. Shore AC, Brennan OM, Ehricht R, Monecke S, Schwarz S, Slickers P, Coleman 445 DC. 2010. Identification and characterization of the multidrug resistance gene cfr in a 446 Panton-Valentine leukocidin-positive sequence type 8 methicillin-resistant 447 Staphylococcus aureus IVa (USA300) isolate. Antimicrob. Agents Chemother. 54:4978-448 4984. 449
23. Arias CA, Vallejo M, Reyes J, Panesso D, Moreno J, Castaneda E, Villegas MV, 450 Murray BE, Quinn JP. 2008. Clinical and microbiological aspects of linezolid 451 resistance mediated by the cfr gene encoding a 23S rRNA methyltransferase. J. Clin. 452 Microbiol. 46:892-896. 453
24. Behnke D, Golubkov VI, Malke H, Boitsov AS, Totolian AA. 1979. Restriction 454 endonuclease analysis of group A streptococcal plasmids determining resistance to 455 macrolides, lincosamides and streptogramin-B antibiotics. FEMS Microbiol. Lett. 6:5-9. 456
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

25. Locke JB, Rahawi S, Lamarre J, Mankin AS, Shaw KJ. 2012. Genetic environment 457 and stability of cfr in methicillin-resistant Staphylococcus aureus CM05. Antimicrob. 458 Agents Chemother. 56:332-340. 459
26. Mendes RE, Deshpande LM, Castanheira M, Dipersio J, Saubolle M, Jones RN. 460 2008. First report of cfr-mediated resistance to linezolid in human staphylococcal clinical 461 isolates recovered in the United States. Antimicrob. Agents Chemother. 52:2244-2246. 462
27. Fritsche TR, Castanheira M, Mendes RE, Jones RN, Deshpande LM. 2008. Abstr. 463 108th Gen. Meet. Am. Soc. Microbiol., abstr A-047. 464
28. Layer F, Strommenger B, Werner G. 2013. Abstr. 23rd Euro. Cong. Clin. Micro. 465 Infect. Dis., abstr O450. 466
29. Flamm RK, Mendes RE, Ross JE, Sader HS, Jones RN. 2013. An international 467 activity and spectrum analysis of linezolid: ZAAPS Program results for 2011. Diagn. 468 Microbiol. Infect. Dis. 76:206-213. 469
30. Jones RN, Kohno S, Ono Y, Ross JE, Yanagihara K. 2009. ZAAPS International 470 Surveillance Program (2007) for linezolid resistance: results from 5591 Gram-positive 471 clinical isolates in 23 countries. Diagn. Microbiol. Infect. Dis. 64:191-201. 472
31. Morales G, Picazo JJ, Baos E, Candel FJ, Arribi A, Pelaez B, Andrade R, de la 473 Torre MA, Fereres J, Sanchez-Garcia M. 2010. Resistance to linezolid is mediated by 474 the cfr gene in the first report of an outbreak of linezolid-resistant Staphylococcus aureus. 475 Clin. Infect. Dis. 50:821-825. 476
32. Mendes RE, Deshpande L, Rodriguez-Noriega E, Ross JE, Jones RN, Morfin-Otero 477 R. 2010. First report of Staphylococcal clinical isolates in Mexico with linezolid 478 resistance caused by cfr: evidence of in vivo cfr mobilization. J. Clin. Microbiol. 479 48:3041-3043. 480
33. Deshpande L, Vega S, Ross J, Jones R, Mendes R. 2013. Abstr. 23rd Euro. Cong. Clin. 481 Micro. Infect. Dis., abstr P1405. 482
34. Ross J, Flamm R, Sader H, Hogan P, Jones R. 2013. Abstr. 53rd Intersci. Conf. 483 Antimicrob. Agents Chemother., abstr C2-088. 484
35. Diaz L, Kiratisin P, Mendes RE, Panesso D, Singh KV, Arias CA. 2012. Transferable 485 plasmid-mediated resistance to linezolid due to cfr in a human clinical isolate of 486 Enterococcus faecalis. Antimicrob. Agents Chemother. 56:3917-3922. 487
36. Patel SN, Memari N, Shahinas D, Toye B, Jamieson FB, Farrell DJ. 2013. Linezolid 488 resistance in Enterococcus faecium isolated in Ontario, Canada. Diagn. Microbiol. Infect. 489 Dis. 77:350-353. 490
37. Rajan V, Kumar VG, Gopal S. 2014. A cfr-positive clinical staphylococcal isolate from 491 India with multiple mechanisms of linezolid-resistance. Indian J. Med. Res. 139:463-467. 492
38. Cui L, Wang Y, Li Y, He T, Schwarz S, Ding Y, Shen J, Lv Y. 2013. Cfr-mediated 493 linezolid-resistance among methicillin-resistant coagulase-negative Staphylococci from 494 infections of humans. PloS one 8:e57096. 495
39. Wang Y, He T, Schwarz S, Zhou D, Shen Z, Wu C, Ma L, Zhang Q, Shen J. 2012. 496 Detection of the staphylococcal multiresistance gene cfr in Escherichia coli of domestic-497 animal origin. The Journal of antimicrobial chemotherapy 67:1094-1098. 498
40. Wang Y, Wang Y, Wu CM, Schwarz S, Shen Z, Zhang W, Zhang Q, Shen JZ. 2011. 499 Detection of the staphylococcal multiresistance gene cfr in Proteus vulgaris of food 500 animal origin. The Journal of antimicrobial chemotherapy 66:2521-2526. 501
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

41. Locke JB, Zuill DE, Sahm DF, Deane J, Denys GA, Shaw KJ. 2013. Abstr. 23rd Euro. 502 Cong. Clin. Micro. Infect. Dis., abstr O 449. 503
42. Locke JB, Hilgers M, Shaw KJ. 2009. Novel ribosomal mutations in Staphylococcus 504 aureus strains identified through selection with the oxazolidinones linezolid and torezolid 505 (TR-700). Antimicrob. Agents Chemother. 53:5265-5274. 506
43. Pattee PA, Baldwin JN. 1961. Transduction of resistance to chlortetracycline and 507 novobiocin in Staphylococcus aureus. J. Bacteriol. 82:875-881. 508
44. CLSI. 2012. Methods for dilution antimicrobial susceptibility tests for bacteria that grow 509 aerobically; approved standard, CLSI Document M07-A9, ninth ed, vol. 32, number 2. 510
45. Baker CN, Tenover FC. 1996. Evaluation of Alamar colorimetric broth microdilution 511 susceptibility testing method for staphylococci and enterococci. J. Clin. Microbiol. 512 34:2654-2659. 513
46. Schenk S, Laddaga RA. 1992. Improved method for electroporation of Staphylococcus 514 aureus. FEMS Microbiol. Lett. 73:133-138. 515
47. Pillai SK, Sakoulas G, Wennersten C, Eliopoulos GM, Moellering RC, Jr., Ferraro 516 MJ, Gold HS. 2002. Linezolid resistance in Staphylococcus aureus: characterization and 517 stability of resistant phenotype. J. Infect. Dis. 186:1603-1607. 518
48. Meka VG, Pillai SK, Sakoulas G, Wennersten C, Venkataraman L, DeGirolami PC, 519 Eliopoulos GM, Moellering RC, Gold HS. 2004. Linezolid resistance in sequential 520 Staphylococcus aureus isolates associated with a T2500A mutation in the 23S rRNA 521 gene and loss of a single copy of rRNA. J. Infect. Dis. 190:311-317. 522
49. Prunier AL, Malbruny B, Tande D, Picard B, Leclercq R. 2002. Clinical isolates of 523 Staphylococcus aureus with ribosomal mutations conferring resistance to macrolides. 524 Antimicrob. Agents Chemother. 46:3054-3056. 525
50. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment 526 search tool. J. Mol. Biol. 215:403-410. 527
51. Goering RV, Ribot EM, Gerner-Smidt P. 2011. Pulsed Field Gel Electrophoresis: 528 Laboratory and Epidemiologic Considerations for Interpretation of Data, p. 167-177. In 529 Persing DH, Tenover FC, Nolte FS, Hayden RT, van Belkum A (ed.), Molecular 530 Microbiology: Diagnostic Principles and Practice, 2nd ed. ASM Press, Washington, D.C. 531
52. Shopsin B, Gomez M, Montgomery SO, Smith DH, Waddington M, Dodge DE, Bost 532 DA, Riehman M, Naidich S, Kreiswirth BN. 1999. Evaluation of protein A gene 533 polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. 534 Clin. Microbiol. 37:3556-3563. 535
53. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. 2000. Multilocus sequence 536 typing for characterization of methicillin-resistant and methicillin-susceptible clones of 537 Staphylococcus aureus. J Clin Microbiol 38:1008-1015. 538
54. Goering RV, Ruff EA. 1983. Comparative analysis of conjugative plasmids mediating 539 gentamicin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 24:450-540 452. 541
55. Holmes DS, Quigley M. 1981. A rapid boiling method for the preparation of bacterial 542 plasmids. Anal. Biochem. 114:193-197. 543
56. Sambrook J, Russell DW. 2001. Molecular Cloning - A Laboratory Manual. Cold 544 Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 545
57. McDougal LK, Steward CD, Killgore GE, Chaitram JM, McAllister SK, Tenover 546 FC. 2003. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus 547
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

aureus isolates from the United States: establishing a national database. J. Clin. 548 Microbiol. 41:5113-5120. 549
58. Mendes RE, Deshpande LM, Bonilla HF, Schwarz S, Huband MD, Jones RN, Quinn 550 JP. 2013. Dissemination of a pSCFS3-like cfr-carrying plasmid in Staphylococcus aureus 551 and Staphylococcus epidermidis clinical isolates recovered from hospitals in Ohio. 552 Antimicrob. Agents Chemother. 57:2923-2928. 553
59. Murphy E, Huwyler L, de Freire Bastos Mdo C. 1985. Transposon Tn554: complete 554 nucleotide sequence and isolation of transposition-defective and antibiotic-sensitive 555 mutants. The EMBO journal 4:3357-3365. 556
60. Kehrenberg C, Ojo KK, Schwarz S. 2004. Nucleotide sequence and organization of the 557 multiresistance plasmid pSCFS1 from Staphylococcus sciuri. The Journal of 558 antimicrobial chemotherapy 54:936-939. 559
61. Kadlec K, Schwarz S. 2009. Novel ABC transporter gene, vga(C), located on a 560 multiresistance plasmid from a porcine methicillin-resistant Staphylococcus aureus 561 ST398 strain. Antimicrob. Agents Chemother. 53:3589-3591. 562
62. Bertsch D, Anderegg J, Lacroix C, Meile L, Stevens MJ. 2013. pDB2011, a 7.6 kb 563 multidrug resistance plasmid from Listeria innocua replicating in Gram-positive and 564 Gram-negative hosts. Plasmid 70:284-287. 565
63. Mollet B, Iida S, Shepherd J, Arber W. 1983. Nucleotide sequence of IS26, a new 566 prokaryotic mobile genetic element. Nucleic Acids Res. 11:6319-6330. 567
64. Barberis-Maino L, Berger-Bachi B, Weber H, Beck WD, Kayser FH. 1987. IS431, a 568 staphylococcal insertion sequence-like element related to IS26 from Proteus vulgaris. 569 Gene 59:107-113. 570
65. Barberis-Maino L, Ryffel C, Kayser FH, Berger-Bachi B. 1990. Complete nucleotide 571 sequence of IS431mec in Staphylococcus aureus. Nucleic Acids Res. 18:5548. 572
66. Gopegui ER, Juan C, Zamorano L, Perez JL, Oliver A. 2012. Transferable multidrug 573 resistance plasmid carrying cfr associated with tet(L), ant(4')-Ia, and dfrK genes from a 574 clinical methicillin-resistant Staphylococcus aureus ST125 strain. Antimicrob. Agents 575 Chemother. 56:2139-2142. 576
67. Malachowa N, DeLeo FR. 2010. Mobile genetic elements of Staphylococcus aureus. 577 Cell. Mol. Life Sci. 67:3057-3071. 578
68. Kadlec K, Schwarz S. 2010. Identification of a plasmid-borne resistance gene cluster 579 comprising the resistance genes erm(T), dfrK, and tet(L) in a porcine methicillin-resistant 580 Staphylococcus aureus ST398 strain. Antimicrob. Agents Chemother. 54:915-918. 581
69. Rouch DA, Skurray RA. 1989. IS257 from Staphylococcus aureus: member of an 582 insertion sequence superfamily prevalent among gram-positive and gram-negative 583 bacteria. Gene 76:195-205. 584
70. Liu Y, Wang Y, Schwarz S, Wang S, Chen L, Wu C, Shen J. 2014. Investigation of a 585 multiresistance gene cfr that fails to mediate resistance to phenicols and oxazolidinones 586 in Enterococcus faecalis. The Journal of antimicrobial chemotherapy 69:892-898. 587
71. Mendes RE, Hogan PA, Streit JM, Jones RN, Flamm RK. 2014. Zyvox(R) Annual 588 Appraisal of Potency and Spectrum (ZAAPS) Program: report of linezolid activity over 9 589 years (2004-12). The Journal of antimicrobial chemotherapy 69:1582-1588. 590
72. Mendes RE, Flamm RK, Hogan PA, Ross JE, Jones RN. 2014. Summary of Linezolid 591 Activity and Resistance Mechanisms Detected during the 2012 LEADER Surveillance 592 Program for the United States. Antimicrob. Agents Chemother. 58:1243-1247. 593
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

73. Baos E, Candel FJ, Merino P, Pena I, Picazo JJ. 2013. Characterization and 594 monitoring of linezolid-resistant clinical isolates of Staphylococcus epidermidis in an 595 intensive care unit 4 years after an outbreak of infection by cfr-mediated linezolid-596 resistant Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 76:325-329. 597
74. Diep BA, Gill SR, Chang RF, Phan TH, Chen JH, Davidson MG, Lin F, Lin J, 598 Carleton HA, Mongodin EF, Sensabaugh GF, Perdreau-Remington F. 2006. 599 Complete genome sequence of USA300, an epidemic clone of community-acquired 600 meticillin-resistant Staphylococcus aureus. Lancet 367:731-739. 601
602 603
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

strain cfr MIC (µg/ml)
TZD LZD FFC TIA CLI TOB TMP CIP ERY OXA CAZ VAN DAP 1128105 + 1 16 >128 >128 >128 >128 >128 >128 >128 >128 >128 2 0.5
29213 - 0.5 2 8 0.5 0.062 0.5 2 0.5 0.25 0.5 16 1 0.25 29213 + p1128105 + 0.5 16 >128 >128 >128 64 >128 0.5 0.25 0.25 16 1 0.25
29213 T2500A - 1 4 8 1 0.062 0.5 2 0.5 0.25 0.25 16 1 0.25 29213 T2500A + p1128105 + 1 16 >128 >128 >128 64 >128 0.5 0.25 0.25 16 1 0.25
on April 10, 2018 by guest
http://aac.asm.org/
Dow
nloaded from