Embed Size (px)
Citation preview

Introduction
Tuberculosis remains a significant global public healthproblem, representing the major cause of death in adults by asingle bacterial agent.1 A large number of people, one-quarterof the world’s population, is infected with the organism andface a significant life-long risk of developing active disease.Barriers to the control of the disease include the worldwidespread of multidrug resistance2 and the marked susceptibilityof subjects with HIV coinfection.3 New approaches to thecontrol of tuberculosis are urgently needed, including more effective implementation of observed chemotherapy,development of new drugs targeting mycobacteria-specificprocesses and development of new vaccines with greater efficacy than Mycobacterium bovis Bacille Calmette–Guerin(BCG), which has been the only vaccine for over 70 years.Strategies for new vaccines include the development ofsubunit vaccines, such as DNA vaccines expressing dominantsecreted proteins of Mycobacterium tuberculosis,4 andimproved viable vaccines, either by enhancing BCG throughthe high-level expression of genes for specific, immuno-dominant M. tuberculosis antigens or by attenuating virulentM. tuberculosis by deletion of defined genes.5 There havebeen recent major advances in mycobacterial genetics thatwill facilitate the development of new viable vaccines. First,the complete genome sequence of M. tuberculosis is known.6
Second, new genetic techniques have been developed thatallow efficient manipulation of mycobacterial genes, mostimportantly successful homologous recombination and trans-poson mutagenesis in slow-growing mycobacteria.7 Thisraises the possibility that targeted gene mutants of M. tuber-culosis could be developed as new vaccine candidates.
Adaptation of M. tuberculosis to intracellular life
The hallmark of bacterial pathogens is their ability to enter,survive and replicate at sites within the host that are not acces-sible to commensal species. Some of these pathogens, such asM. tuberculosis, are able to survive and replicate within phago-cytic cells that are part of the host antimicrobial defencesystem. Mycobacterium tuberculosis uses a variety of strate-gies to modify its intracellular environment within phagocytes,predominantly through alterations in the characteristics of themacrophage phagosome. The bacilli inhibit fusion with themacrophage lysosome, thus limiting its exposure to degrada-tive lysosomal enzymes.8,9 Vacuoles surrounding M. tuber-culosis lack the proton-pumping ATPase needed for vacuoleacidification,10 thus inhibiting the activation of low pH-dependent lysosomal enzymes. Mycobacterium tuberculosiscan also modify the antigen-presenting ability of infectedmacrophages, contributing to immune evasion.11 While thiscomplex array of methods M. tuberculosis possess to promoteintracellular persistence is impressive, we are hindered by thefact that the molecular mechanisms by which M. tuberculosisresists host antibacterial functions are largely unknown.
What are the possible approaches to define the molecularmechanisms that govern the survival of M. tuberculosisin vivo? Bacterial pathogens must adapt to a wide variety ofchanging environmental conditions as infection proceeds, suchas changes in the type and availability of nutrients and exposure to antimicrobial products and other host cell killingmechanisms. On entry of the host, pathogens express specificgene products that permit persistence and multiplication in thecorrect location and the avoidance of host defence systems.The environmental signals that regulate the control of viru-lence gene expression in pathogens are simple physical andchemical factors, such as temperature, osmolarity, pH, oxygenavailability, reactive oxygen and nitrogen compounds, nutri-ent availability and inorganic ion concentrations. Many ofthese factors change in the transition from the external
Immunology and Cell Biology (2000) 78, 311–317
Special Feature
Life on the inside: Probing Mycobacterium tuberculosis geneexpression during infection
JAMES A TRICCAS 1 and BRIGITTE GICQUEL 2
1Centenary Institute of Cancer Medicine and Cell Biology, Newtown, New South Wales, Australia and 2Unité de Génétique Mycobactérienne, Institut Pasteur, Paris, France
Summary The identification of Mycobacterium tuberculosis genes specifically expressed during infection is akey step in understanding mycobacterial pathogenesis. Such genes most likely encode products required for survival within the host and for progressive infection. Recent advances in mycobacterial genetics have permittedthe development of new techniques and the adaptation of existing methods to analyse mycobacterial in vivo geneexpression and virulence. This has revealed a subset of M. tuberculosis genes that are differentially expressedduring infection and has demonstrated that a number of components contribute to the virulence of the organism.This information is expected to provide new strategies to prevent tuberculosis infection, new targets for anti-microbial therapy and new insights into the infectious process.
Key words: gene, in vivo expression, infection, tuberculosis, vaccine, virulence.
Correspondence: Dr James A Triccas, Centenary Institute ofCancer Medicine and Cell Biology, Locked Bag no. 6, Newtown,NSW 2042, Australia. Email: [email protected]
Received 1 May 2000; accepted 1 May 2000.

environment to the host, between different locations in thehost and between different hosts. Accordingly, examinationof the in vivo induction profile of host-induced genes,largely conducted on the intracellular pathogen Salmonellatyphimurium, has demonstrated that the major genesexpressed in vivo are those involved with the acquisition ofmetals, the synthesis and acquisition of nucleotides and cofactors, DNA repair, membrane modification, thermo-tolerance, osmotic tolerance and acid tolerance.12,13 Further-more, mutations in many in vivo-induced genes confervirulence defects in the organism, suggesting that this classof genes plays an important role in infection and/or pathogenesis.12,13
With this in mind, efforts are now being directed towardsidentifying M. tuberculosis genes of which expression ispreferentially upregulated within the host during infection.The remainder of the present review outlines recent efforts toidentify the repertoire of host-induced genes by M. tuber-culosis and the current status of our knowledge of those components of M. tuberculosis required for life in vivo.
Defining M. tuberculosis genes expressed in vivo
Many bacterial pathogens will selectively express genes onlyin response to signals encountered during infection. Thusmethods to find such genes must allow the direct isolation ofgenes expressed in vivo, as conditions encountered within the host cannot be reproduced accurately within the labora-tory. Methodologies used to study bacterial gene expressionin vivo have been reviewed in detail elsewhere.14 Essentiallyall of these techniques were first developed to study fast-growing, readily manipulated bacteria, such as Salmonellaand Yersinia, and the challenge has been to adapt these tech-niques to the mycobacteria.
Green fluorescent protein as a reporter of gene expressionin vivo
The technique termed differential fluorescence induction(DFI) was first developed to identify promoters of Salmo-nella that are differentially expressed within the intracellularenvironment.13 In this method, a pool of bacteria containingrandom DNA fragments that contain possible differentiallyexpressed promoters are placed upstream of a promoterlesscopy of the gene encoding the green fluorescent protein(GFP) of the jellyfish Aequorea victoria. Green fluorescentprotein is a 27 kDa naturally fluorescent protein for whichfluorescence can be readily detected by fluorescencemicroscopy and conventional flow cytometry, making it auseful marker for the study of gene expression or proteinlocalization within living cells and in real time.15 Culturedcells are then infected with this bacterial pool and cells harbouring bacteria that become fluorescent in this intra-cellular environment are isolated by fluorescence-activatedcell sorting (FACS). Bacteria are isolated, grown ex vivo andthe least fluorescent bacteria (those harbouring promotersless active in vivo) are collected by FACS. These bacteria arethen used to reinfect cells and intracellular-dependent induc-tion of gfp confirmed for individual bacteria. Using this strat-egy, a number of S. typhimurium promoters selectivelyinduced within the macrophage have been identified.13 Some
of these macrophage-induced genes are necessary for sur-vival within the host, displaying a direct link between intracellular gene induction and pathogenicity.
The observation that GFP is efficiently expressed inmycobacteria and that fluorescent bacteria can be directlyobserved in infected macrophages16,17 has suggested that sucha strategy may be applicable to the study of gene induction bymycobacteria in host cells. A similar system to DFI has beenused to identify promoters of Mycobacterium marinum thatshow elevated levels of expression within macrophages com-pared with the extracellular environment.18 We have reporteda variation on the DFI approach where fluorescence of GFPis combined with the counterselectable property of the Bacillus subtilis SacB protein to isolate M. tuberculosispromoters displaying enhanced in vivo activity (Fig. 1).19
SacB is a secreted enzyme that confers sucrose sensitivity onmycobacteria.20 This property has been used to perform efficient allelic exchange and construct transposon mutantlibraries in both the fast- and slow-growing mycobacteria.7,21
In our system, macrophages are infected with M. bovis BCGcontaining a library of M. tuberculosis promoters controllinggfp and sacB expression, and GFP-positive bacteria sorted byFACS. SacB expression is then used to eliminate BCG cloneswith strong promoter activity outside the macrophage,leading to the enrichment of clones containing M. tuberculo-sis promoters differentially expressed intracellularly.
Selective capture of transcribed sequences
A novel approach, termed selective capture of transcribedsequences (SCOTS), has been used to identify RNA synthe-sized by M. tuberculosis in response to phagocytosis bymacrophages (Fig. 1).22 This method attempts to overcomethe inherent limitations in the development of cDNA selec-tion methods to study host-mediated bacterial gene expres-sion. In this method, total microbial and host-cell RNA isextracted from infected cells and converted to cDNA frag-ments with defined terminal sequences for amplification bypolymerase chain reaction (PCR). Microbial cDNA are thenselectively captured by hybridization to biotinylated micro-bial ribosomal DNA (rDNA) fragments. In order to identifybacterial genes that are expressed in response to interactionwith the host, these captured cDNA are hybridized togenomic DNA fragments that have been prehybridized withboth cDNA and rDNA prepared from bacteria grown in vitro.This results in the enrichment of bacterial cDNA that corre-sponds to sequences preferentially transcribed during growthin the host. Unlike methods using GFP, SCOTS directly iden-tifies host-expressed genes rather than promoter sequences.Furthermore, SCOTS may potentially be applied to the detec-tion of in vivo gene expression from cells obtained fromliving tissues in natural disease states.
Signature-tagged mutagenesis to identify in vivoexpressed M. tuberculosis genes required for virulence
Signature-tagged mutagenesis (STM) allows the screening ofa large pool of tagged mutants for those containing insertionsin genes required for growth within the infected host.23
Pooled mutants are used to infect the host, permitted to multiply, and hybridization used to identify those strains
JA Triccas and B Gicquel312

present in the initial inoculum but absent from bacteriarecovered after infection (Fig. 1). While this technique doesnot provide information concerning the differential expres-sion of genes in vivo with respect to ex vivo conditions, invivo expression is assumed due to the requirement of geneexpression for survival within the host.
Signature-tagged mutagenesis requires an efficient inser-tional mutagenesis system to generate tagged mutations rep-resenting the genome of the target organisms. Such a systemis available for pathogens such as S. typhimurium, Staphylo-coccus aureus, Vibrio cholerae and Streptococcus pneu-moniae and have permitted the use of STM to identify in vivoexpressed virulence factors of these organisms.23–27 We havedescribed an efficient transposon mutagenesis system basedon a plasmid carrying a thermosensitive replicon and thecounter selectable marker sacB.7 By using this mutagenesissystem, we have been able to successfully adapt STM toM. tuberculosis, which has permitted the identification of 16 mutants attenuated for their growth in the lungs of mice(Table 1).28
Other possible methods for identifying M. tuberculosisgenes expressed in vivo
The first practical strategy used to identify genes expressedduring bacterial infection was termed in vivo expression tech-nology (IVET).29 This technique uses the animal as the selec-tive medium to identify bacterial genes expressed specificallyduring infection, and provides a powerful tool to investigatethe changes in metabolism, gene expression and cell surfaceproperties that allow microbial infection of host tissues. Thissystem acts as a promoter trap, whereby bacterial promotersare selected that drive the expression of a gene that isrequired for virulence. A promoterless reporter gene is placeddownstream of the virulence gene to allow the elimination ofpromoters that function ex vivo (e.g. on normal laboratorymedium).
Most of the in vivo-induced genes of S. typhimurium havebeen identified by this technique.12 While the lack of a suitable M. tuberculosis gene proven to be essential formycobacterial virulence has hindered the application of theIVET method to the mycobacteria, recent studies havedemonstrated that a number of M. tuberculosis genes are necessary for survival in macrophages or the organs ofinfected mice (see Table 1). Such genes may allow the appli-cation of this powerful technique to M. tuberculosis.
Because the entire genomic sequence of M. tuberculosis isknown,6 the opportunity exists to use microarray technologyto study gene regulation within mycobacteria. Microarrayassays are based on the ability of labelled cDNA sequencesfrom cells and tissues (probes) to hybridize to complemen-tary sequences (targets) immobilized to solid surfaces. Liter-ally thousand of genes can be analysed simultaneously; thusthe 4000 or so genes of M. tuberculosis could be readilyanalysed for a defined condition in a single experiment.Mycobacterium tuberculosis microarrays have already beendeveloped and have been used to analyse drug-induced alter-ations in gene expression in M. tuberculosis30 and to comparethe genomic differences between M. tuberculosis and BCGvaccine strains.31 One could envisage that this technology hasthe potential to provide a wealth of information concerningM. tuberculosis gene expression in response to host infection.
So what do we really know about M. tuberculosis geneexpression during infection?
What goes up?
The identified M. tuberculosis genes for which expression isknown to be enhanced during infection are represented inTable 2. It is worth noting that all of these genes have beenidentified in models using cultured macrophages, and in nocase has their upregulation during infection within theanimal host been confirmed. Although the number of genesidentified is small, inferences regarding the ecology of
M. tuberculosis gene expression during infection 313
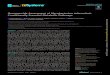
Fig. 1 Methods used for the iso-lation of Mycobacterium tubercu-losis genes expressed in hosttissue. (a) Green fluorescentprotein (GFP)/SacB selection usesfluorescence-activated cell sorting(FACS) to isolate promotersexpressed in vivo and counter-selection by sucrose to eliminatepromoters also expressed ex vivo.(b) Selective capture of tran-scribed sequences (SCOTS) is apositive cDNA selection methodthat identifies sequences preferen-tially transcribed in vivo ( , ).(c) Signature-tagged mutagenesis(STM) selects for mutants that arepresent in the initial inoculumused for infection (d) but do notsurvive in the animal (s). mØ,Macrophage; PCR, polymerasechain reaction.

infection can be made by analysing the proposed function oftheir products. A number of M. tuberculosis genes encodingproteins predicted to be involved in the stress response are
induced in vivo, which is the case for in vivo expressed genesof other bacteria.12 These products include the M. tuberculo-sis transketolase of the pentose phosphate pathway,19 whose
JA Triccas and B Gicquel314
Table 2 Mycobacterium tuberculosis genes displaying enhanced expression within the host
Gene Proposed function Selection/detection method Reference
aceA (Rv0467)* Isocitrate lyase SCOTS 22acr (Rv2031c) Alpha-crystallin homologue Luc/GFP 41ctpV (Rv0969) Cation transport SCOTS 22cysD (Rv1285) Sulfate adenylate transferase subunit 2 GFP/SacB 19fadB4 (Rv3141) 3-hydroxyacyl-CoA dehydrogenase GFP/SacB 19mceD (Rv0170) Cell invasion and persistence SCOTS 22mtrA (Rv3246c) Response regulator GFP 42pks2 (Rv3825c) Polyketide synthesis SCOTS 22sigE (Rv1221) Sigma factor SCOTS 22sigH (Rv3223c) Sigma factor SCOTS 22ponA (Rv3682) Penicillin-binding protein SCOTS 22prrA (Rv0903c) Response regulator SCOTS 22sseA (Rv3283) Putative thiosulfate sulfurtransferase GFP/SacB 19tkt (Rv1449c) Transketolase GFP/SacB 19uvrA (Rv1638) DNA repair SCOTS 22Rv0834c Unknown GFP/SacB 19Rv3070 Unknown: membrane associated? SCOTS 22Rv3483c Unknown: membrane associated? SCOTS 22Rv3718c PE-PGRS subfamily GFP/SacB 19
*Gene number as annotated by Cole et al.6 SCOTS, selective capture of transcribed sequences; GFP/SacB, selection by green fluorescentprotein (GFP) and the Bacillus subtilis SacB protein; Luc/GFP, detection by luciferase and GFP; PE-PGRS, proline-glutamic acid-polymorphicglycine-rich repetitive sequence.
Table 1 Mycobacterium tuberculosis genes required for virulence
Gene Proposed function Tissue Method Reference
acr (Rv2031c)* Alpha-crystallin homologue Macrophage Allelic exchange 41ahpC (Rv2428)† Alkyl hydroperoxide reductase Spleen, liver, lung Antisense RNA 53drrC (Rv2938) ABC transporter Lung, macrophage STM 28erp (Rv3810) Unknown; cell surface exposed Macrophage, lung, spleen Allelic exchange 50fadD26 (Rv2930) Lipid metabolism Lung STM 28fadD28 (Rv2941) Mycocerosic acid synthesis Lung STM 45fbpA (Rv3804c) Mycolyl transferase/fibronectin binding Macrophage Allelic exchange 48glnA1 (Rv2220) Glutamine synthetase Macrophage Enzyme inhibition 49ideR (Rv2711) Iron-dependent repressor Spleen, lung Constitutive gene repression 55katG (Rv1908c)† Catalase/peroxidase Spleen, liver, lung In vivo complementation 52lipF (Rv3487c) Lipid degradation Lung, macrophage STM 28mceD (Rv0170) Cell invasion and persistence Macrophage In vivo complementation 60mbtB (Rv2383c) Hydroxyphenylthiazoline biosynthesis Macrophage Allelic exchange 56mmpL2 (Rv0507c) Proton-dependent transporter Lung STM 28mmpL4 (Rv0450c) Proton-dependent transporter Lung STM 28mmpL7 (Rv2942) Proton-dependent transporter Lung, macrophage STM 28,45modA (Rv1857) Molybdenum fixation Lung STM 28pks6 (Rv0405) Polyketide synthase Lung STM 28purC (0780) Purine biosynthesis Macrophage, spleen, liver, lung Allelic exchange 51SigA (rpoV; Rv2703)† Principal sigma factor Spleen, liver, lung In vivo complementation 54Rv3018c PPE family Lung STM 28Rv1395 Transcriptional activator Lung STM 28Rv0204 Unknown Lung STM 28Rv2452c Unknown Lung STM 28
*Gene number as annotated by Cole et al.6 †Genes identified as virulence factors of Mycobacterium bovis of the M. tuberculosis complex.STM, signature-tagged mutagenesis; ABC, ATP-binding cassette; PPE, proline-proline-glutamic acid.

homologue in Saccharomyces cerevisiae can compensate forthe loss of superoxide dismutase, a key enzyme in the cell’sdefence against toxic oxygen metabolites generated byphagocytic cells in an attempt to eliminate pathogenicinvaders.32 Enhanced expression of M. tuberculosis UvrA of the DNA repair system is typical of a bacterial stressresponse. In both Salmonella and Yersinia, genes of the DNArepair system are induced during infection and appear to berequired for survival within the macrophage.33,34
The extracytoplasmic sigma factors have been shown toregulate gene expression in response to numerous environ-mental conditions in a number of bacterial species.35–37 BothSigE and SigH play a role in the resistance of Mycobacteriumsmegmatis to various stress stimuli, including elevated tem-perature and oxidative stress.38,39 These results imply thatthese sigma factors may be involved in the ability of myco-bacteria to adapt to the variety of stresses encountered in theenvironment of the host. Accordingly, expression of M. tuber-culosis sigE and sigH is markedly enhanced within themacrophage compared with the extracellular environment.22
Furthermore, another extracellular sigma factor, SigF,appears to control expression of the Acr protein,40 itselfinduced within the macrophage and necessary for intra-cellular persistence of M. tuberculosis.41
Bacteria typically respond to changes in environmentalconditions through paired membrane protein sensors andresponse regulators. Expression of two genes encoding postulated response regulators of M. tuberculosis is increasedin vivo, mtrA42 and prrA.22 At this stage we have no knowl-edge of the specific environmental signal that triggers theirexpression and what genes, if any, are under their control. InSalmonella, a large number of genes required for intracellularsurvival are induced by expression of the two-component regulatory system PhoPQ, whose expression is induced bylow Mg2+ concentration in the macrophage phagosome.43
Enzymes involved in cell wall metabolism are differen-tially expressed by a number of pathogens within thehost.24,27,44 Three M. tuberculosis genes thought to beinvolved in cell wall metabolism have been identified asinduced during infection: fadB4 (3-hydroxyacyl-CoAdehydrogenase),19 ponA (class I penicillin binding protein)22
and pks2 (polyketide synthase).22 A large proportion of theM. tuberculosis genome encodes proteins involved in cellwall synthesis and a number of these proteins appear to benecessary for in vivo growth (see Table 1).
Mycobacterium tuberculosis in vivo expressed genesrequired for virulence
Genes confirmed to be necessary for in vivo persistence of M. tuberculosis are outlined in Table 1. Most of this infor-mation has been gained from the two published studies thatused STM to identify virulence determinants.28,45 The moststriking finding is the identification of a cluster of virulencegenes located on a 50 kb chromosomal region. These genes,fadD26, fadD28, drrC and mmplL, are implicated in the for-mation and transport of the lipid phthiocerol dimycocerosate(PDIM), which is found only in pathogenic mycobacteria.46,47
Synthesis and transport of PDIM is required for in vivogrowth, particularly within the lung, because mutations in these genes do not affect growth in other organs45 or in
in vitro macrophage cell culture (L Camacho et al., unpubl.data, 2000). Furthermore, mutations in genes encodingrelated enzymes (mmpL2, mmpL4),28 enzymes involved in the metabolism of lipids (lipF, pks6, fbpA)28,48 and other cellwall structures (glnA)49 attenuate in vivo growth of the bacilli,demonstrating the importance of cell wall components of M. tuberculosis for virulence. While its exact function isunknown, the cell-surface exposed exported repetitive protein(ERP) is also essential for the in vivo persistence of M. tuberculosis.50
A number of processes that are necessary for the in vivosurvival of other bacterial pathogens also appear importantfor the survival of M. tuberculosis within the host. Theseinclude the biosynthesis of purines, presumably lacking in theintracellular environment as indicated by the requirement ofthe purC gene,51 and genes involved in the response to environmental stress, including katG,52 acr 41 and ahpC.53 Theregulation of gene transcription is known to be an importantdeterminant of virulence and mycobacteria are no exception,as mutations in the principal sigma factor SigA54 and a pre-dicted transcriptional activator (Rv1395)28 have an attenuat-ing effect. Whether these two genes actually control theexpression of virulence genes is currently unknown. In con-trast, the mycobacterial iron-dependent repressor is impli-cated in the control of the expression of genes essential forvirulence, as inhibition of its function attenuates M. tuber-culosis.55 The link between M. tuberculosis virulence and ironis further demonstrated by the requirement for the mbtB geneproduct for macrophage persistence, as MbtB is essential forbiosynthesis of the mycobactin siderophores used for ironacquisition.56
Mycobacterium tuberculosis strains lacking in vivoexpressed virulence genes: New vaccine candidates?
Owing to the variable protective efficacy of BCG against theadult forms of tuberculosis, ranging from 0 to 90%,57 effortsare being directed at developing new, improved antituber-culosis vaccines. Because a variety of M. tuberculosis genesare necessary for in vivo virulence, strains lacking thesegenes could be possible candidates as attenuated vaccines.We have investigated the vaccine potential of an auxotrophicmutant of M. tuberculosis that lacks the purC gene and havefound that, while the vaccine was protective in a guinea pigmodel of tuberculosis, this effect was no greater than thatafforded by BCG.51 This raises a major problem associatedwith new anti-TB vaccination strategies: how can we deter-mine if a new vaccine is better than the existing BCG?Bacille Calmette–Guerin induces significant protectionagainst virulent M. tuberculosis in animal models.58 Thus, itmay be necessary to look at other parameters in addition tochanges in bacterial load, the standard method to assessvaccine efficacy. These include the induction of immunolog-ical memory and the long-term persistence of the vaccine’seffect. On this latter point, while BCG is very effective, it isalso highly attenuated, which may explain why BCG vacci-nation in humans wanes after approximately 10 years.59
Therefore, mutant M. tuberculosis strains that are less atten-uated than BCG, such as those identified in the study ofCamacho et al.,28 could be expected to persist longer and possibly provide a more prolonged protective effect. This
M. tuberculosis gene expression during infection 315

requires that the strain is attenuated sufficiently not to induceimmune pathology. Such a hypothesis is yet to be tested.
Gene products that influence survival are potential targetsfor novel antituberculosis drugs, especially those eitherexposed on the cell surface, and thus accessible to antibiotics,or involved in the synthesis of cell wall structures. Aninhibitor of M. tuberculosis glutamine synthetase blocks bacterial multiplication in both media and human mono-nuclear phagocytes and growth inhibition is associated witha reduction in the amount of the virulence-associated cellwall component poly glutamate/glutamine.49 These resultssuggest that targeting extracellular proteins of M. tuber-culosis is a feasible strategy for developing new antibiotics.
Conclusions
Whilst our knowledge of the mechanisms used by M. tuber-culosis to promote infection has increased over the last fewyears, our understanding of these mechanisms at a molecularlevel is still quite limited. Development of new techniques toanalyse mycobacterial in vivo gene expression and virulence,such as SCOTS, and the adaptation of existing techniques tothe mycobacteria, such as DFI and STM, are importantadvances in understanding tuberculosis infection. Such tech-niques permitting investigation of the interaction between M. tuberculosis and the host will provide the key to under-standing the pathogenesis of infection, as well as providingthe basis for designing new anti-infective agents and preven-tative vaccines.
Acknowledgements
We thank members of our laboratories for useful comments.JAT is supported by a Peter Doherty NHMRC Fellowship.
References
1 Raviglione MC, Snider DE, Kochi A. Global epidemiology oftuberculosis. Morbidity and mortality of a worldwide epidemic.JAMA 1995; 273: 220–6.
2 Petrini B, Hoffner S. Drug-resistant and multidrug-resistanttubercle bacilli. Int. J. Antimicrob. Agents 1999; 13: 93–7.
3 Britton WJ, Garsia RJ. Mycobacterial infections in clinicalimmunology. In: Bradley J, McClusky J (eds). ClinicalImmunology. Oxford: Oxford Univerisity Press, 1997; 483–98.
4 Kamath AT, Feng CG, Macdonald M, Briscoe H, Britton WJ.Differential protective efficacy of DNA vaccines expressingsecreted proteins of Mycobacterium tuberculosis. Infect. Immun.1999; 67: 1702–7.
5 Young DB, Duncan K. Prospects for new interventions in thetreatment and prevention of mycobacterial disease. Annu. Rev.Microbiol. 1995; 49: 641–73.
6 Cole ST, Brosch R, Parkhill J et al. Deciphering the biology ofMycobacterium tuberculosis from the complete genomesequence. Nature 1998; 393: 537–44.
7 Pelicic V, Jackson M, Reyrat JM, Jacobs Jr WR, Gicquel B,Guilhot C. Efficient allelic exchange and transposon mutagene-sis in Mycobacterium tuberculosis. Proc. Natl Acad. Sci. USA1997; 94: 10 955–60.
8 Clemens DL. Characterization of the Mycobacterium tubercu-losis phagosome. Trends Microbiol. 1996; 4: 113–18.
9 Ferrari G, Langen H, Naito M, Pieters J. A coat protein on
phagosomes involved in the intracellular survival of myco-bacteria. Cell 1999; 97: 435–47.
10 Sturgill-Koszycki S, Schlesinger PH, Chakraborty P et al. Lackof acidification in Mycobacterium phagosomes produced byexclusion of the vesicular proton-ATPase. Science 1994; 263:678–81.
11 Gercken J, Pryjma J, Ernst M. Defective antigen presentation byMycobacterium tuberculosis-infected monocytes. Infect. Immun.1994; 62: 3472–8.
12 Heithoff DM, Conner CP, Mahan MJ. Dissecting the biology ofa pathogen during infection. Trends Microbiol. 1997; 5: 509–13.
13 Valdivia RH, Falkow S. Fluorescence-based isolation of bacter-ial genes expressed within host cells. Science 1997; 277:2007–11.
14 Handfield M, Levesque RC. Strategies for isolation of in vivoexpressed genes from bacteria. FEMS Microbiol. Rev. 1999; 23:69–91.
15 Cubit AB, Heim R, Adams SR, Boyd AE, Gross LA, Tsien RY.Understanding, improving and using green fluorescent proteins.Trends Biochem. Sci. 1995; 20: 448–55.
16 Dhandayuthapani S, Via LE, Thomas CA, Horowitz PM, DereticD, Deretic V. Green fluorescent protein as a marker for geneexpression and cell biology of mycobacterial interactions withmacrophages. Mol. Microbiol. 1995; 17: 901–12.
17 Kremer L, Baulard A, Estaquier J, Poulain-Godefroy O, Locht C.Green fluorescent protein as a new expression marker inmycobacteria. Mol. Microbiol. 1995; 17: 913–22.
18 Barker LP, Brooks DM, Small PL. The identification ofMycobacterium marinum genes differentially expressed inmacrophage phagosomes using promoter fusions to green fluo-rescent protein. Mol. Microbiol. 1998; 29: 1167–77.
19 Triccas JA, Berthet FX, Pelicic V, Gicquel B. Use of fluores-cence induction and sucrose counterselection to identifyMycobacterium tuberculosis genes expressed within host cells.Microbiology 1999; 145: 2923–30.
20 Pelicic V, Reyrat JM, Gicquel B. Expression of the Bacillus subtilis sacB gene confers sucrose sensitivity on mycobacteria.J. Bacteriol. 1996; 178: 1197–9.
21 Pelicic V, Reyrat JM, Gicquel B. Generation of unmarkeddirected mutations in mycobacteria, using sucrose counter-selectable suicide vectors. Mol. Microbiol. 1996; 20: 919–25.
22 Graham JE, Clark-Curtiss JE. Identification of Mycobacteriumtuberculosis RNAs synthesized in response to phagocytosis by human macrophages by selective capture of transcribedsequences (SCOTS). Proc. Natl Acad. Sci. USA 1999; 96:11 554–9.
23 Hensel M, Shea J, Gleeson C, Jones M, Dalton E, Holden D.Simultaneous identification of bacterial virulence genes by negative selection. Science 1995; 269: 400–3.
24 Mei JM, Nourbakhsh F, Ford CW, Holden DW. Identification ofStaphylococcus aureus virulence genes in a murine model ofbacteraemia using signature-tagged mutagenesis. Mol. Micro-biol. 1997; 26: 399–407.
25 Chiang S, Mekalanos J. Use of signature-tagged transposonmutagenesis to identify Vibrio cholerae genes critical for colo-nization. Mol. Microbiol. 1998; 27: 795–805.
26 Coulter S, Schwan W, Ng E et al. Staphylococcus aureus geneticloci impacting growth and survival in multiple infection envi-ronments. Mol. Microbiol. 1998; 30: 393–404.
27 Polissi A, Pontiggia A, Feger G et al. Large-scale identificationof virulence genes from Streptococcus pneumoniae. Infect.Immun. 1998; 66: 5620–9.
28 Camacho LR, Ensergueix D, Perez E, Gicquel B, Guilhot C.Identification of a virulence gene cluster of Mycobacterium
JA Triccas and B Gicquel316

tuberculosis by signature-tagged transposon mutagenesis. Mol.Microbiol. 1999; 34: 257–67.
29 Mahan MJ, Slauch JM, Mekalanos JJ. Selection of bacterial virulence genes that are specifically induced in host tissues.Science 1993; 259: 686–8.
30 Wilson M, DeRisi J, Kristensen HH et al. Exploring drug-induced alterations in gene expression in Mycobacterium tuber-culosis by microarray hybridization. Proc. Natl Acad. Sci. USA1999; 96: 12 833–8.
31 Behr MA, Wilson MA, Gill WP et al. Comparative genomics ofBCG vaccines by whole-genome DNA microarray. Science1999; 284: 1520–3.
32 Miller RA, Britigan BE. Role of oxidants in microbial patho-physiology. Clin. Microbiol. Rev. 1997; 10: 1–18.
33 Young GM, Miller VL. Identification of novel chromosomal lociaffecting Yersinia enterocolitica pathogenesis. Mol. Microbiol.1997; 25: 319–28.
34 Heithoff DM, Conner CP, Hanna PC, Julio SM, Hentschel U,Mahan MJ. Bacterial infection as assessed by in vivo geneexpression. Proc. Natl Acad. Sci. USA 1997; 94: 934–9.
35 Lonetto MA, Brown KL, Rudd KE, Buttner MJ. Analysis of theStreptomyces coelicolor sigE gene reveals the existence of asubfamily of eubacterial RNA polymerase sigma factorsinvolved in the regulation of extracytoplasmic functions. Proc.Natl Acad. Sci. USA 1994; 91: 7573–7.
36 Deretic V, Schurr MJ, Boucher JC, Martin DW. Conversion ofPseudomonas aeruginosa to mucoidy in cystic fibrosis: Environmental stress and regulation of bacterial virulence byalternative sigma factors. J. Bacteriol. 1994; 176: 2773–80.
37 Hiratsu K, Amemura M, Nashimoto H, Shinagawa H, MakinoK. The rpoE gene of Escherichia coli, which encodes sigma E,is essential for bacterial growth at high temperature. J. Bacteriol.1995; 177: 2918–22.
38 Wu QL, Kong D, Lam K, Husson RN. A mycobacterial extra-cytoplasmic function sigma factor involved in survival follow-ing stress. J. Bacteriol. 1997; 179: 2922–9.
39 Fernandes ND, Wu QL, Kong D, Puyang X, Garg S, Husson RN.A mycobacterial extracytoplasmic sigma factor involved in survival following heat shock and oxidative stress. J. Bacteriol.1999; 181: 4266–74.
40 Manabe YC, Chen JM, Ko CG, Chen P, Bishai WR. Conditionalsigma factor expression, using the inducible acetamidase pro-moter, reveals that the Mycobacterium tuberculosis sigF genemodulates expression of the 16-kilodalton alpha-crystallinhomologue. J. Bacteriol. 1999; 181: 7629–33.
41 Yuan Y, Crane DD, Simpson RM et al. The 16-kDa alpha-crystallin (Acr) protein of Mycobacterium tuberculosis isrequired for growth in macrophages. Proc. Natl Acad. Sci. USA1998; 95: 9578–83.
42 Via LE, Curcic R, Mudd MH, Dhandayuthapani S, Ulmer RJ,Deretic V. Elements of signal transduction in Mycobacteriumtuberculosis: In vitro phosphorylation and in vivo expressionof the response regulator MtrA. J. Bacteriol. 1996; 178:3314–21.
43 Garcia Vescovi E, Soncini FC, Groisman EA. Mg2+ as an extra-cellular signal: Environmental regulation of Salmonella viru-lence. Cell 1996; 84: 165–74.
44 Lowe AM, Beattie DT, Deresiewicz RL. Identification of novel
staphylococcal virulence genes by in vivo expression technol-ogy. Mol. Microbiol. 1998; 27: 967–76.
45 Cox JS, Chen B, McNeil M, Jacobs WR. Complex lipid deter-mines tissue-specific replication of Mycobacterium tuberculosisin mice. Nature 1999; 402: 79–83.
46 Kolattukudy PE, Fernandes ND, Azad AK, Fitzmaurice AM,Sirakova TD. Biochemistry and molecular genetics of cell-walllipid biosynthesis in mycobacteria. Mol. Microbiol. 1997; 24:263–70.
47 Brennan PJ, Nikaido H. The envelope of mycobacteria. Annu.Rev. Biochem. 1995; 64: 29–63.
48 Armitige LY, Jagannath C, Wanger AR, Norris SJ. Disruption ofthe genes encoding antigen 85A and antigen 85B of Myco-bacterium tuberculosis H37Rv: Effect on growth in culture andin macrophages. Infect. Immun. 2000; 68: 767–78.
49 Harth G, Horwitz MA. An inhibitor of exported Mycobacteriumtuberculosis glutamine synthetase selectively blocks the growthof pathogenic mycobacteria in axenic culture and in humanmonocytes: Extracellular proteins as potential novel drugtargets. J. Exp. Med. 1999; 189: 1425–36.
50 Berthet FX, Lagranderie M, Gounon P et al. Attenuation of virulence by disruption of the Mycobacterium tuberculosis erpgene. Science 1998; 282: 759–62.
51 Jackson M, Phalen SW, Lagranderie M et al. Persistence andprotective efficacy of a Mycobacterium tuberculosis auxotrophvaccine. Infect. Immun. 1999; 67: 2867–73.
52 Wilson TM, Lisle GW, Collins DM. Effect of inhA and katG onisoniazid resistance and virulence of Mycobacterium bovis. Mol.Microbiol. 1995; 15: 1009–15.
53 Wilson T, Lisle GW, Marcinkeviciene JA, Blanchard JS, CollinsDM. Antisense RNA to ahpC, an oxidative stress defence gene involved in isoniazid resistance, indicates that AhpC ofMycobacterium bovis has virulence properties. Microbiology1998; 144: 2687–95.
54 Collins DM, de Kawakami RP, Lisle GW, Pascopella L, BloomBR, Jacobs WR. Mutation of the principal sigma factor causesloss of virulence in a strain of the Mycobacterium tuberculosiscomplex. Proc. Natl Acad. Sci. USA 1995; 92: 8036–40.
55 Manabe YC, Saviola BJ, Sun L, Murphy JR, Bishai WR. Atten-uation of virulence in Mycobacterium tuberculosis expressing aconstitutively active iron repressor. Proc. Natl Acad. Sci. USA1999; 96: 12 844–8.
56 De Voss JJ, Rutter K, Schroeder BG, Su H, Zhu Y, Barry CE.The salicylate-derived mycobactin siderophores of Mycobac-terium tuberculosis are essential for growth in macrophages.Proc. Natl Acad. Sci. USA 2000; 97: 1252–7.
57 Colditz GA, Berkey CS, Mosteller F et al. The efficacy ofbacillus Calmette-Guerin vaccination of newborns and infants inthe prevention of tuberculosis: Meta-analyses of the publishedliterature. Pediatrics 1995; 96: 29–35.
58 Orme IM. New vaccines against tuberculosis. The status ofcurrent research. Infect. Dis. Clin. North Am. 1999; 13: 169–85.
59 Sterne JA, Rodrigues LC, Guedes IN. Does the efficacy of BCGdecline with time since vaccination? Int. J. Tuberc. Lung Dis.1998; 2: 200–7.
60 Arruda S, Bomfim G, Knights R, Huima-Byron T, Riley LW.Cloning of an M. tuberculosis DNA fragment associated withentry and survival inside cells. Science 1993; 261: 1454–7.
M. tuberculosis gene expression during infection 317