Embed Size (px)
Citation preview
Laminin stimulates and guidesaxonal outgrowth via growth conemyosin II activityStephen G Turney1 & Paul C Bridgman2
Guidance cues and signal transduction mechanisms acting at
the nerve growth cone are fairly well understood, but the
intracellular mechanisms operating to change the direction of
axon outgrowth remain unknown. We now show that growth
cones integrate myosin II–dependent contraction for rapid,
coordinated turning at borders of laminin stripes in response to
signals from laminin-activated integrin receptors; in the absence
of myosin II activity, outgrowth continues across the borders.
Signals derived from integrin and other guidance cue receptors con-verge and act to change vertebrate growth cone motility, coordinatingthe direction and rate of outgrowth1–3. The molecular mechanismsthat link and drive structural rearrangements of the growth coneactin and microtubule cytoskeleton during turning are not fullydefined4–6. Most models of turning emphasize control through thedynamics of peripheral extensions that do not depend on contractilemyosin (myosin II (ref. 7)). To determine if myosin II has a role,we modified a turning assay for growth cones8 and imaged neuritesgrowing at borders of substrate-bound laminin-1 stripes in cultureslacking myosin IIB activity or after pharmacologically blockingmyosin II activity.
We grew peripheral nerve explants and dissociated cells fromembryonic day (E) 13.5 mouse embryos on coverslips that containedalternating stripes of poly-l-ornithine plus laminin-1 (PLO+LN1) andpolyornithine (PLO). Cells from individual embryos were plated
0 0 0 0
2 h 7 h
6 h 21 h
5 h 3.5 h
15 h 13.5 h
a b c
d e f g
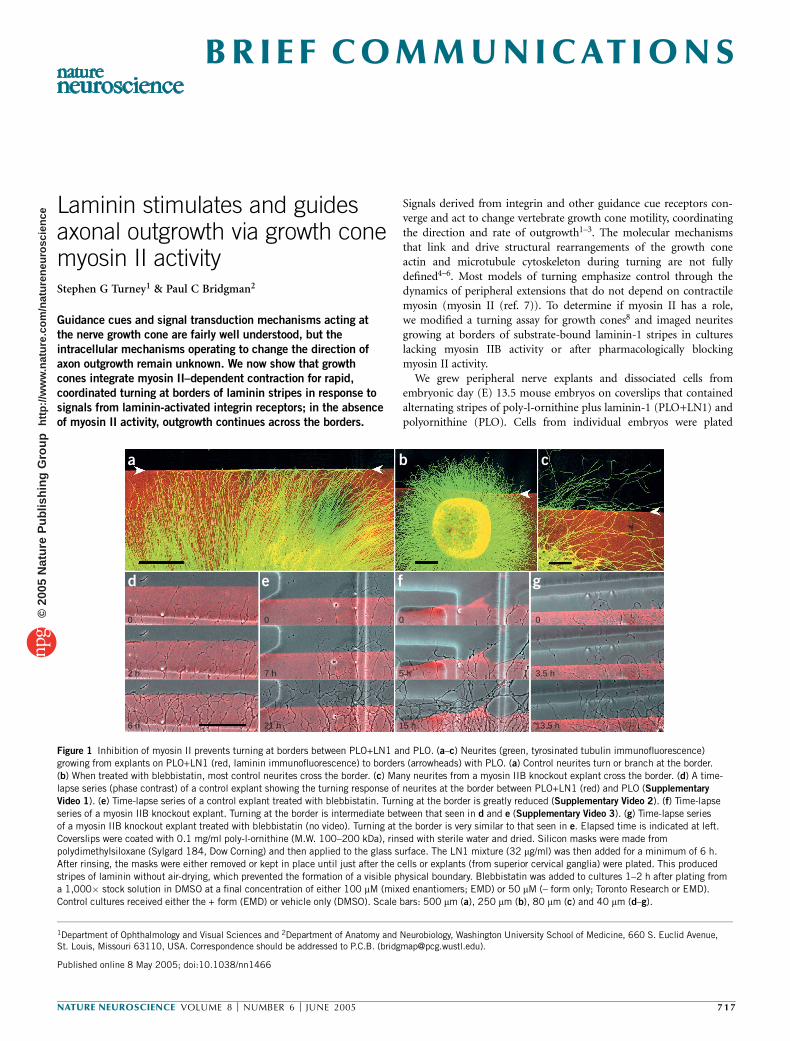
Figure 1 Inhibition of myosin II prevents turning at borders between PLO+LN1 and PLO. (a–c) Neurites (green, tyrosinated tubulin immunofluorescence)growing from explants on PLO+LN1 (red, laminin immunofluorescence) to borders (arrowheads) with PLO. (a) Control neurites turn or branch at the border.
(b) When treated with blebbistatin, most control neurites cross the border. (c) Many neurites from a myosin IIB knockout explant cross the border. (d) A time-
lapse series (phase contrast) of a control explant showing the turning response of neurites at the border between PLO+LN1 (red) and PLO (Supplementary
Video 1). (e) Time-lapse series of a control explant treated with blebbistatin. Turning at the border is greatly reduced (Supplementary Video 2). (f) Time-lapse
series of a myosin IIB knockout explant. Turning at the border is intermediate between that seen in d and e (Supplementary Video 3). (g) Time-lapse series
of a myosin IIB knockout explant treated with blebbistatin (no video). Turning at the border is very similar to that seen in e. Elapsed time is indicated at left.
Coverslips were coated with 0.1 mg/ml poly-l-ornithine (M.W. 100–200 kDa), rinsed with sterile water and dried. Silicon masks were made from
polydimethylsiloxane (Sylgard 184, Dow Corning) and then applied to the glass surface. The LN1 mixture (32 mg/ml) was then added for a minimum of 6 h.
After rinsing, the masks were either removed or kept in place until just after the cells or explants (from superior cervical ganglia) were plated. This produced
stripes of laminin without air-drying, which prevented the formation of a visible physical boundary. Blebbistatin was added to cultures 1–2 h after plating from
a 1,000� stock solution in DMSO at a final concentration of either 100 mM (mixed enantiomers; EMD) or 50 mM (– form only; Toronto Research or EMD).
Control cultures received either the + form (EMD) or vehicle only (DMSO). Scale bars: 500 mm (a), 250 mm (b), 80 mm (c) and 40 mm (d–g).
Published online 8 May 2005; doi:10.1038/nn1466
1Department of Ophthalmology and Visual Sciences and 2Department of Anatomy and Neurobiology, Washington University School of Medicine, 660 S. Euclid Avenue,St. Louis, Missouri 63110, USA. Correspondence should be addressed to P.C.B. ([email protected]).
NATURE NEUROSCIENCE VOLUME 8 [ NUMBER 6 [ JUNE 2005 7 1 7
B R I E F CO M M U N I C AT I O N S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e
separately to allow comparison between those that were expressingmyosin IIB (wild-type and heterozygous) and myosin IIB knockouts9.Myosin IIB knockout neurons still contain myosin IIA. Blebbistatin canbe used to eliminate myosin IIA activity10.
Neurites of control explants growing on PLO+LN1 changed direc-tions at the border, only rarely crossing onto the PLO (Fig. 1a). Forexplants that were about 500 mm from the border (n ¼ 14), individualneurites could be followed, and 90–100% turned at the border. Neuritesof explants (control, n ¼ 10; myosin IIB knockout, n ¼ 8) that weretreated with blebbistatin rarely changed direction and frequentlycrossed the borders when growth was from PLO+LN1 to PLO(Fig. 1b). Explants from myosin IIB knockout embryos (n ¼ 9)showed an intermediate response to borders. Many neurites crossedthe border without changing direction (Fig. 1c). This suggests thatmyosin II activity is required for a growth cone–mediated turningresponse at LN1 borders. The efficiency and speed of turning alsodepends upon the amount or isoform of myosin II activity. Myosin IIBmay be required for rapid and efficient turning.
We used time-lapse recordings to characterize the turning response(Supplementary Note). To identify the substrate border, LN1 wasconjugated with the fluorochrome Cy3. Control growth cones growingon PLO+LN1 turned or branched upon contacting the border withPLO (Fig. 1d, Supplementary Video 1). After turning, neurites grewalong the border on the side containing LN1. In a representativeexperiment, growth cones responded immediately to the border (thebase of the growth cone never left the PLO+LN1) in one of three ways(Table 1a). The majority extended processes on the PLO+LN1 parallelto the border and continued growing in this direction. Some branchedafter contacting the border and then continued growing along it.Usually one branch retracted. Others sidestepped parallel to the border.
Periodic probing of the border by processes extending onto the PLOwas also observed. The few that crossed the border grew onto the PLOfor a short distance with growth cones spreading and retractingmultiple times before retracting. In blebbistatin-treated cultures, con-trol and myosin IIB knockout neurons grew in irregular curved pathsand rarely detected borders (Fig. 1e,g, Supplementary Video 2,Table 1a). Most neurites continued to grow unimpeded across theborder. All others paused, branched or turned at the border beforeeventually crossing. The behavior of myosin IIB knockout neurites wasin-between: many growth cones crossed the border without turning,but they paused and usually formed a varicosity at the border site.Although some neurites branched or turned at borders, many even-tually crossed at an angle and then sometimes returned to thePLO+LN1 (Fig. 1f, Supplementary Video 3, Table 1a). A consistentprobing of the border was not observed.
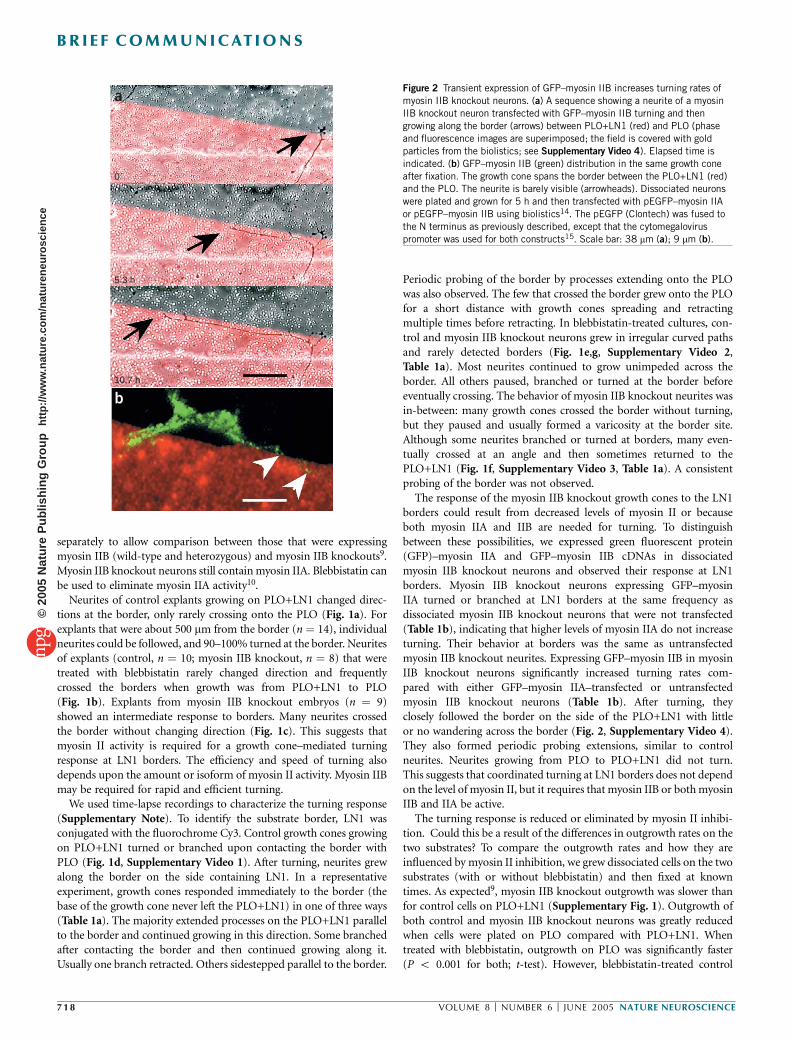
The response of the myosin IIB knockout growth cones to the LN1borders could result from decreased levels of myosin II or becauseboth myosin IIA and IIB are needed for turning. To distinguishbetween these possibilities, we expressed green fluorescent protein(GFP)–myosin IIA and GFP–myosin IIB cDNAs in dissociatedmyosin IIB knockout neurons and observed their response at LN1borders. Myosin IIB knockout neurons expressing GFP–myosinIIA turned or branched at LN1 borders at the same frequency asdissociated myosin IIB knockout neurons that were not transfected(Table 1b), indicating that higher levels of myosin IIA do not increaseturning. Their behavior at borders was the same as untransfectedmyosin IIB knockout neurites. Expressing GFP–myosin IIB in myosinIIB knockout neurons significantly increased turning rates com-pared with either GFP–myosin IIA–transfected or untransfectedmyosin IIB knockout neurons (Table 1b). After turning, theyclosely followed the border on the side of the PLO+LN1 with littleor no wandering across the border (Fig. 2, Supplementary Video 4).They also formed periodic probing extensions, similar to controlneurites. Neurites growing from PLO to PLO+LN1 did not turn.This suggests that coordinated turning at LN1 borders does not dependon the level of myosin II, but it requires that myosin IIB or both myosinIIB and IIA be active.
The turning response is reduced or eliminated by myosin II inhibi-tion. Could this be a result of the differences in outgrowth rates on thetwo substrates? To compare the outgrowth rates and how they areinfluenced by myosin II inhibition, we grew dissociated cells on the twosubstrates (with or without blebbistatin) and then fixed at knowntimes. As expected9, myosin IIB knockout outgrowth was slower thanfor control cells on PLO+LN1 (Supplementary Fig. 1). Outgrowth ofboth control and myosin IIB knockout neurons was greatly reducedwhen cells were plated on PLO compared with PLO+LN1. Whentreated with blebbistatin, outgrowth on PLO was significantly faster(P o 0.001 for both; t-test). However, blebbistatin-treated control
a
b
0
5.3 h
10.7 h
Figure 2 Transient expression of GFP–myosin IIB increases turning rates of
myosin IIB knockout neurons. (a) A sequence showing a neurite of a myosin
IIB knockout neuron transfected with GFP–myosin IIB turning and then
growing along the border (arrows) between PLO+LN1 (red) and PLO (phase
and fluorescence images are superimposed; the field is covered with gold
particles from the biolistics; see Supplementary Video 4). Elapsed time is
indicated. (b) GFP–myosin IIB (green) distribution in the same growth cone
after fixation. The growth cone spans the border between the PLO+LN1 (red)and the PLO. The neurite is barely visible (arrowheads). Dissociated neurons
were plated and grown for 5 h and then transfected with pEGFP–myosin IIA
or pEGFP–myosin IIB using biolistics14. The pEGFP (Clontech) was fused to
the N terminus as previously described, except that the cytomegalovirus
promoter was used for both constructs15. Scale bar: 38 mm (a); 9 mm (b).
71 8 VOLUME 8 [ NUMBER 6 [ JUNE 2005 NATURE NEUROSCIENCE
B R I E F C O M M U N I C AT I O N S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e

neurons on PLO never reached the high outgrowth rates observed inuntreated control cells on PLO+LN1. This indicates that myosin II actsto inhibit outgrowth on PLO independent of isoform, and myosin IIBactivity is necessary for achieving the maximal outgrowth rates onPLO+LN1.
Is turning a consequence of the abrupt change in outgrowth rates atthe border, or does it depend on differential activation of integrinreceptors? To distinguish between these possibilities, we grew cellson alternating stripes of substrates that differed in their capacitiesfor integrin activation but that produced the same proportionaldifference in outgrowth rates. The frequency that growth cones turnedand the degree to which they remained close to the border did notcorrelate with differences in outgrowth rates (Supplementary Table 1),but it did depend upon substrate-bound laminin, consistent withintegrin activation.
We used immunofluorescence to determine if peripheral neuronsalso express myosin IIC (ref. 11). Growth cones were positive formyosin IIC, indicating that they contain all three myosin II isoforms(Supplementary Fig. 2). Blebbistatin is likely to inhibit all forms ofmyosin II (ref. 12).
To determine if LN1 affects the location of myosin II, we comparedthe distribution of myosin IIA and IIB in cells growing on stripes of thetwo substrates. There was no obvious difference in the distribution ofmyosin IIA in growth cones on the two substrates or at borders.However, myosin IIB consistently changed from a random distributionin growth cones on PLO to the more characteristic concentration in thetransition zone in growth cones on PLO+LN1 (ref. 13). For growthcones that had recently turned or were growing along the LN1 side ofthe border, myosin IIB was often asymmetrically concentrated on theside of the growth cone closest to the border (Supplementary Fig. 2).
These results indicate that integrin activation is linked to myosin II(mainly myosin IIB)-dependent changes in growth cone motility.Depending upon the level and location of activation by integrins,local myosin II contractile activity causes growth cones to turn, branch,retract, stall or advance with precision along a laminin border. Becausethe preference for laminin substrates was strong only when the fullcomplement of myosin II (A, B and C) was present, the growth conemust integrate the activity for rapid, coordinated turning. Thesequence of activity is likely to include integrin activation, signaling,adhesive complex formation, coupling to the actin cytoskeleton,redistribution of actin filaments, local formation of myosin II bipolarfilaments, contraction and development of traction forces. Thus, otherguidance cues modulating the location or degree of integrin activationcan have direct effects on myosin II–dependent growth cone behaviorand can profoundly affect guidance through a contractile mechanism.
Note: Supplementary information is available on the Nature Neuroscience website.
ACKNOWLEDGMENTSG. Philips and D. Morris provided technical support. We thank J. Cooper forcomments on the manuscript. The US National Institutes of Health providedfinancial support (NS26150 to P.C.B.)
COMPETING INTERESTS STATEMENTThe authors declare that they have no competing financial interests.
Received 1 April; accepted 21 April 2005
Published online at http://www.nature.com/natureneuroscience/
1. Hopker, V.H., Shewan, D., Tessier-Lavigne, M., Poo, M. & Holt, C. Nature 401, 69–73(1999).
2. Stevens, A. & Jacobs, J.R. J. Neurosci. 22, 4448–4455 (2002).3. Nakamoto, T., Kain, K.H. & Ginsberg, M.H. Curr. Biol. 14, R121–R123 (2004).4. Suter, D.M. & Forscher, P. J. Neurobiol. 44, 97–113 (2000).5. Mitchison, T. & Kirschner, M. Neuron 1, 761–772 (1988).6. Schmucker, D. Neuron 40, 4–6 (2003).7. Dent, E.W. & Gertler, F.B. Neuron 40, 209–227 (2003).8. Gundersen, R.W. Dev. Biol. 121, 423–431 (1987).9. Bridgman, P.C., Dave, S., Asnes, C.F., Tullio, A.N. & Adelstein, R.S. J. Neurosci. 21,
6159–6169 (2001).10. Straight, A.F. et al. Science 299, 1743–1747 (2003).11. Golomb, E. et al. J. Biol. Chem. 279, 2800–2808 (2004).12. Limouze, J., Straight, A.F., Mitchison, T. & Sellers, J.R. J. Muscle Res. Cell Motil. 25,
337–341 (2004).13. Rochlin, M.W., Itoh, K., Adelstein, R.S. & Bridgman, P.C. J. Cell Sci. 108, 3661–3670
(1995).14. Bridgman, P.C., Brown, M.E. & Balan, I. Methods Cell Biol. 71, 353–68 (2003).15. Brown, M.E. & Bridgman, P.C. J. Cell Sci. 116, 1087–1094 (2003).
Table 1 Responses to laminin-1 borders
(a) Response of neurites of peripheral neuron explants at laminin-1 borders from
time-lapse recordings.
Initial response at border (%)
Phenotype Blebbistatin treatment Cross Turn Branch Sidestep n
Control � 10 45 27 15 20
Control + 72 11a 17a 0 36
KO � 36 28 36 0 28
KO + 75 6a 19a 0 16
(b) Turning frequencies of dissociated neurons transfected with GFP–myosin IIA or
GFP–myosin IIB compared with untransfected neurons of fixed cultures.
Phenotype Growth direction Turning frequency (%) n
Control PLO+LN1 to PLO 93 89
Control PLO to PLO+LN1 0 30
KO + GFP–myosin IIA PLO+LN1 to PLO 47 93
KO + GFP–myosin IIB PLO+LN1 to PLO 75b 47
KO PLO+LN1 to PLO 48 139
KO PLO to PLO+LN1 0 23
aAfter turning or branching and growing along the border, most of these eventually crossedduring the recording.Direction was PLO+LN1 to PLO in all cases; KO, myosin IIB knockout.bDifference compared with untransfected myosin IIB knockout and myosin IIA–transfectedknockout neurons was significant (w2, P o 0.01 for both).For analysis of turning frequencies in fixed cells, only two categories were used: (i) turned orbranched, and (ii) crossed or undetermined (that is, when a growth cone was just at the border,and we could not determine its future direction). The growth direction was determined byfollowing the individual neurites to their cell bodies. KO, myosin IIB knockout.
NATURE NEUROSCIENCE VOLUME 8 [ NUMBER 6 [ JUNE 2005 7 1 9
B R I E F C O M M U N I C AT I O N S©
2005
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eneu
rosc
ienc
e





























