Embed Size (px)
Citation preview

HUMAN GENE THERAPY 13:1415–1425 (August 10, 2002)© Mary Ann Liebert, Inc.
Intranasal Gene Transfer by Chitosan–DNA NanospheresProtects BALB/c Mice Against Acute Respiratory
Syncytial Virus Infection
MUKESH KUMAR,1 ARUNA K. BEHERA,1 RICHARD F. LOCKEY,1 JIAN ZHANG,1
GURAMAN BHULLAR,1 CRISTINA PEREZ DE LA CRUZ,2 LI-CHEN CHEN,3 KAM W. LEONG,2
SHAU-KU HUANG,3 and SHYAM S. MOHAPATRA1
ABSTRACT
Respiratory syncytial virus (RSV) infection is often associated in infancy with life-threatening bronchiolitis,which is also a major risk factor for the development of asthma. At present, no effective prophylaxis is avail-able against RSV infection. Herein, we describe an effective prophylactic intranasal gene transfer strategyutilizing chitosan–DNA nanospheres (IGT), containing a cocktail of plasmid DNAs encoding all RSV antigens,except L. A single administration of IGT (25 mg/mouse) induces expression of the mRNA and proteins of allantigens in the lung and results in a significant reduction of viral titers and viral antigen load after acute RSVinfection of these mice. IGT-administered mice show no significant change in airway reactivity to metha-choline and no apparent pulmonary inflammation. Furthermore, IGT results in significant induction of RSV-specific IgG antibodies, nasal IgA antibodies, cytotoxic T lymphocytes, and interferon-g production in thelung and splenocytes compared with controls. Together, these results demonstrate the potential of IGT againstacute RSV infection.
1415
OVERVIEW SUMMARY
We describe an intranasal gene transfer approach againstRSV infection, using chitosan nanospheres. Intranasal ad-ministration of chitosan nanospheres, containing plasmidDNAs encoding nine different RSV antigens, into the mouselung reduced RSV titers and induced the production of anti-RSV antibody with neutralizing properties. The therapyalso enhanced interferon-g production in spleen and lungand generated cytotoxic T lymphocyte responses againstRSV. This prophylactic gene expression therapy also re-duced RSV-induced lung inflammation. We conclude thatintranasal gene transfer utilizing chitosan nanospheres maybe useful against acute RSV infection.
INTRODUCTION
RESPIRATORY SYNCYTIAL VIRUS (RSV), the most commoncause of viral lower respiratory tract infections in infants
and children, affects about 4 million children globally andcauses about 100,000 hospitalizations and 4500 deaths per an-num in the United States alone (Centers for Disease Controland Prevention, 1999). Acute RSV infection is associated withepisodes of bronchiolitis, wheezing, and exacerbation of asthmain children (Chanock et al., 1992). In the 1960s, children ad-ministered a formalin-inactivated RSV vaccine developed ex-aggerated disease when subsequent RSV infection occurred(Chanock et al., 1992). The development of a protective RSVvaccine has been a high priority at a global level. No effectivevaccine is currently available for RSV infection.
Experimental vaccines have included subunit, peptide, at-tenuated-live, and RSV DNA vaccines, and some have pro-gressed to clinical trials (Hall, 1994; Brandenburg et al.,2001). Immunization with plasmid DNAs can potentially leadto more efficient antigen processing that induces a strong pro-tective cellular and humoral immune response, as well asgreater safety and cost-effectiveness (Cohen et al., 1998; Don-nelly et al., 1998). Intramuscular injection of pDNA encod-ing the RSV-F or RSV-G protein was effective in mice (Li et
1Division of Allergy and Immunology-Joy McCann Culverhouse Airway Disease Center, James A. Haley Veterans Administration Hospitaland University of South Florida College of Medicine, Tampa, FL 33612.
2Department of Biomedical Engineering, Johns Hopkins University School of Medicine, Baltimore, MD 21205.3Johns Hopkins Asthma and Allergy Center, Baltimore, MD 21224.

al., 1998, 2000); however, the quantity of DNA used per unitbody mass, as much as 10 mg/kg, and the route of adminis-tration chosen are inconvenient for infants and are suboptimalfor inducing mucosal immunity against a pulmonary infection(Guy et al., 2001).
Most, if not all, of the RSV antigens are immunogenic in hu-mans and mice, although only the F and G antigens have beenshown to induce neutralizing antibodies against RSV (Connorset al., 1991; Wyatt et al., 1999). An analysis of the cytotoxicT lymphocyte (CTL) repertoire in humans indicates that the N,F, P, M2, and NS2 proteins are strong target antigens (Nicholaset al., 1990). Similarly, in BALB/c mice, the F, N, and espe-cially the M2 proteins have been shown to be the major targetantigens of CTL activity (Domachowske and Rosenberg, 1999).Both serum and mucosal antibodies and MHC class I-restrictedCTLs are considered to protect against RSV infection (Bran-denburg et al., 2001).
Because the airway epithelium is the major target of RSVinfection, we reasoned that DNA vaccines capable of mount-ing a mucosal immunity against RSV might be more effective.However, the development of such mucosal DNA vaccines hasbeen hindered by inefficient transgene expression of pDNAs inthe airway epithelium. Chitosan, a biodegradable, biocompati-ble, low-toxic polysaccharide has been used as a gene carrierto mucosal sites such as the gastrointestinal tract (Artursson etal., 1994; Richardson et al., 1999; Roy et al., 1999) and by thenasal route (Illum et al., 2001), which induces higher transfec-tion efficiency and ensures more sustained expression of thevaccine antigens. Further, DNA vaccines induce elevated pro-duction of interferon-g (IFN-g), which has an antiviral effectagainst RSV (Kumar et al., 1999). To test potential mucosalgenetic immunization, in this study, we utilized a strategy in-volving an intranasal gene transfer, referred to as IGT, with chi-tosan–DNA nanospheres containing a cocktail of plasmidDNAs (pDNAs) encoding nine immunogenic RSV antigens,against acute RSV infection in a BALB/c mouse model. Theeffectiveness and mechanism of this IGT strategy were inves-tigated. Results demonstrate that IGT is safe and effectiveagainst RSV and significantly attenuates pulmonary inflamma-tion induced by RSV infection.
MATERIALS AND METHODS
Animals
Six-week old female BALB/c mice were purchased fromJackson Laboratory (Bar Harbor, ME) and maintained underpathogen-free conditions at the animal center. All procedureswere reviewed and approved by the University of South Floridaand James A. Haley Veterans Administration Medical CenterCommittee on Animal Research (Tampa, FL).
Cells and virus
HEp-2 (ATCC CCL-23) cells and RSV A2 Long strain (VR-1302) were obtained from the American Type Culture Collec-tion (ATCC, Manassas, VA). HEp-2 is an epithelial-like cervi-cal carcinoma cell line and is used for propagating RSV. Cellswere grown and viral stocks were prepared as described previ-ously (Behera et al., 2001).
Vaccine development and protocol
Individual RSV cDNAs were amplified from an RSV-in-fected mouse lung cDNA library by polymerase chain reaction(PCR), using Vent polymerase (New England BioLabs, Bev-erly, MA), and cloned in the mammalian expression vectorpVAX (Invitrogen, San Diego, CA). The resulting plasmidswere propagated in Escherichia coli DH5a cells. Large-scaleplasmid DNA was prepared with a Qiagen kit (Qiagen,Chatsworth, CA), according to the manufacturer’s specifica-tions. This produced sufficiently pure DNA. Equal quantitiesof pDNAs were mixed to make a cocktail of RSV cDNAs.DNA–chitosan nanospheres were generated as described pre-viously (Roy et al., 1999), with the total DNA concentration inthe solution equally contributed by the nine RSV plasmids; theresulting product is referred to as IGT. Mice were intranasallyadministered IGT (25 mg of total DNA per mouse) under lightanesthesia. Control mice received either phosphate-bufferedsaline (PBS), equivalent quantities of naked DNA, empty vec-tor complexed in chitosan nanospheres (chitosan plus pVAX),or chitosan alone. Sixteen days after vaccination, mice were in-fected intranasally with 1 3 106 PFU of the human RSV A2strain (ATCC) in a 50-ml volume. Five days postinfection, micewere killed and their lungs and spleens were collected asepti-cally for reverse transcriptase (RT)-PCR, histopathologicalstudies, and cytokine and viral plaque analyses. On day 21 af-ter vaccination mice were bled to obtain serum. Nasal washeswere also collected on day 21 for IgA antibody assays as de-scribed previously (Matsuo et al., 2000).
Quantitation of RSV titers and antigen in lung
To quantify RSV titers in mouse lung, whole lungs were firstweighed and placed immediately in Eagle’s minimal essentialmedium (EMEM) supplemented with 10% fetal bovine serum(FBS). Lungs were homogenized and centrifuged at 10,000 rpmfor 10 min at 4°C, and the clear supernatants were used forplaque assays by the shell vial technique (Domachowske andBonville, 1998). RSV antigen load was determined as describedpreviously (Kumar et al., 1999).
RNA extraction and RT-PCR analysis
Total cellular RNA was isolated from lung tissue with TRI-zol reagent (Life Technologies, Gaithersburg, MD), accordingto the manufacturer’s instructions. RT-PCR was carried out fordifferent RSV genes, as described previously (Behera et al.,2001). The primers used for individual cDNA amplification andtheir corresponding sizes are listed in Table 1.
Pulmonary function
To evaluate the pulmonary function in vaccinated and con-trol groups, mice were administered IGT, as described previ-ously. Three days later, airway responsiveness was assessednoninvasively in conscious, unrestrained mice in a whole bodyplethysmograph (Buxco Electronics, Troy, NY) as previouslydescribed (Matsuse et al., 2000). With this system, the volumechanges that occur during a normal respiratory cycle arerecorded as the pressure difference between an animal-con-taining chamber and a reference chamber. The resulting signalis used to calculate respiratory frequency, minute volume, tidal
KUMAR ET AL.1416

volume, and enhanced pause (Penh). Penh was used as the mea-sure of bronchoconstriction and was calculated according to thefollowing formula: Penh 5 pause 3 (peak expiratory pres-sure/peak inspiratory pressure), where pause is the ratio of timerequired to exhale the last 30% of tidal volume relative to thetotal time of expiration. Mice were placed in the plethysmo-graph and the chamber was equilibrated for 10 min. They wereexposed to aerosolized PBS (to establish baseline) followed byincremental doses (6, 12.5, 25, and 50 mg/ml) of methacholine(Sigma Chemicals, St. Louis, MO). Each dose of methacholinewas aerosolized for 5 min, and respiratory measurements wererecorded for 5 min afterward. During the recording period, anaverage of each variable was derived from every 30 breaths (or30 sec, whichever occurred first). The maximum Penh value af-ter each dose was used to measure the extent of bronchocon-striction.
Bronchoalveolar lavage, spleen cell culture, and assayfor IFN-g
Bronchoalveolar lavage (BAL) were performed on IGT-ad-ministered and control mice, as described previously (Kumaret al., 1999). For spleen cell culture, single-cell suspensionswere prepared from the spleens of BALB/c mice and culturedin wells coated with anti-CD3 antibody (1 mg/ml, clone 17A2;PharMingen, San Diego, CA). IFN-g was assayed from BALfluid and 24-hr spleen cell culture supernatant, using an ELISAkit (R&D Systems, Minneapolis, MN).
ELISA for antibodies
Microtiter plates were coated overnight at 4°C with proteins(500 ng/well) from a purified RSV or HEp-2 cell protein prepa-ration. The plates were washed and then blocked at room tem-perature for 30 min with 100 ml of PBS containing 10% FBS.This solution was replaced with 2-fold serial dilutions of im-mune sera or of nasal washes prepared in PBS containing 10%
FBS and 0.2% (v/v) Tween 20. The plates were incubated for2 hr at room temperature and washed three times. For the eval-uation of IgA titers, 100 ml of a 1:1000 dilution of biotinylatedanti-mouse IgA antibody (556978; PharMingen) was added andthe plates were incubated for another 2 hr. After three washes,100 ml of avidin–peroxidase conjugate (1:10,000; Sigma) wasadded and the plates were incubated for another 1 hr. For theevaluation of IgG, anti-mouse IgG–peroxidase conjugate wasadded at a dilution of 1:10,000 (Boehringer, Mannheim, Ger-many). After three washes, substrate was added and absorbancewas read at 450 nm.
Virus neutralization assay
Different dilutions of serum obtained on day 21 were mixedwith 100 ml of RSV inoculum and incubated at 37°C for 1 hr.This was used to infect HEp-2 cell cultures growing in 48-wellculture plates. RSV titer was determined as described previ-ously.
Immunoblotting
Lung tissues from uninfected and mice infected with RSVwere homogenized in 1 ml of lysis buffer (0.5% Triton X-100,150 mM NaCl, 15 mM Tris, 1 mM CaCl2, 1 mM MgCl2 [pH7.4], 1 mM phenylmethylsulfonyl fluoride [PMSF], aprotinin[1 mg/ml], leupeptin [1 mg/ml], and pepstatin [1 mg/ml]). Thehomogenates were incubated on ice for 30 min and centrifugedat 3000 rpm for 10 min. Clear supernatants were collected andtotal protein was estimated with bicinchoninic acid (BCA)reagent (Pierce, Rockford, IL). Seventy-five micrograms of pro-tein extract was fractionated on a 4–20% gradient sodium do-decyl sulfate (SDS)-polyacrylamide gel, transferred to nitro-cellulose membrane, and processed as described previously(Behera et al., 1998). Briefly, the membrane was blocked andincubated overnight at 4°C with a 1:250 dilution of pooledserum from IGT-administered and control mice. The membrane
GENE THERAPY FOR RSV INFECTION 1417
TABLE 1. PRIMER SEQUENCES FOR RSV cDNAs
PCRcDNA Primer sequence product (bp)
NS1 Sense: 59-ctg acg gga tcc gaa ttc agg atg ggc agc aat tca ttg-39 419Antisense: 59-ggc att ctc gag tta tgg att aag atc aaa tcc aag taa-39
NS2 Sense: 59-ttg tcc gga tcc acc atg gac aca acc cac aa-39 374Antisense: 59-ggc att ctc gag tta tgg att gag atc ata ctt gt-39
M Sense: 59-gtc ggc gga tcc aat atg gaa aca tac gtg aac-39 770Antisense: 59-ggc att ctc gag tta atc ttc cat ggg ttt gat tgc a-39
SH Sense: 59-cac act gta ccc aca atg gaa aat aca tcc ata aca ata g-39 194Antisense: 59-gac aga ctg cag cta tgt gtt gac tcg agc tct tgg taa ctc-39
F Sense: 59-ggg ccg gga tcc aca atg gag ttg cta atc ctc aaa-39 1724Antisense: 59-cta tgt cga ctt agt tac taa atg caa tat tat tta-39
M2 Sense: 59-gtg tgc gga tcc aat atg tca cga agg aat cct tgc a-39 825Antisense: 59-ggc atg ctc gag tta tga cac taa tat ata tat tgt ata-39
N Sense: 59-gtg tgg gga tcc aag atg gct ctt agc aaa gtc-39 1175Antisense: 59-ggc att ctc gag tca aag ctc tac atc att atc t-39
G Sense: 59-gtg tgc gga tcc aac atg tcc aaa aac aag gac caa cgc-39 896Antisense: 59-gtt gtc gac taa cta ctg gcg tgg tgt gtt-39
P Sense: 59-ttg tgg gga tcc atc atg gaa aag ttt gct cct gaa-39 725Antisense: 59-ggc atg ctc gag tca gaa atc ttc aag tga tag atc at-39
was washed four times in washing buffer and incubated withanti-mouse IgG peroxidase conjugate for 1 hr at room temper-ature. After four more washes, the blot was developed and ex-posed to X-ray film.
Histology and scoring for airway inflammation
Histological staining and a semiquantitative analysis of air-way inflammation in the lungs of IGT and control groups ofmice were performed as described previously (Kumar et al.,1999). Lung inflammation was assessed after staining the sec-
tions with hematoxylin and eosin (H&E) and scoring for sever-ity on a scale of 0–3, indicating the degree of inflammation.The entire lung section was reviewed, and pathological changeswere evaluated for epithelial damage, peribronchovascular cellinfiltrate, and interstitial–alveolar cell infiltrate for mononuclearcells and polymorphs.
CTL studies
Splenocytes (2.5 3 106 cells/ml) from mice administeredIGT and from control groups were incubated in complete RPMI
KUMAR ET AL.1418
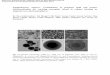
FIG. 1. (A) Expression of RSV cDNAs after IGT administration. BALB/c mice were intranasally administered a cocktail ofRSV cDNAs cloned in the plasmid vector pVAX (IGT) and complexed as chitosan nanospheres. Each mouse was instilled witha total of 25 mg of cocktail DNA. Animals were killed 3 days after IGT administration and RT-PCR was performed on total lungRNA. The products were electrophoresed. Marker lane, 1-kb DNA marker; lanes NS1, NS2, M, SH, F, M2, N, G, and P, PCRproducts corresponding to the RSV cDNAs. (B) Immunoblot analysis. Serum samples were collected from PBS- and IGT-ad-ministered mice, pooled, and used to detect RSV antigens from RSV-infected (lanes 2, 4, and 5) and uninfected (lanes 1 and 3)murine lungs on Western blots. Data from one of two experiments with similar results are shown.
containing interleukin 2 (IL-2, 10 U/ml) and persistently RSV-infected mitomycin (Sigma)-treated fibroblasts (BCH4 cells,2.5 3 106 cells/ml) (Fernie et al., 1981). Cultures were testedon day 6 for antigen-specific lysis by adding varying numbersof effector cells to 51Cr-labeled syngeneic fibroblasts, eitherpersistently RSV-infected (BCH4) or uninfected (BC) targetcells (1 3 104). After 5 hr of incubation at 37°C, cell super-natants were harvested for the determination of 51Cr in a g
counter. The percentage of specific lysis was calculated as [(ex-perimental cpm 2 spontaneous cpm)/(total cpm 2 spontaneouscpm)] 3 100. Spontaneous release and total release were de-termined from target cells incubated with medium alone or af-ter the addition of 2.5% Triton X-100, respectively.
Statistical analysis
Pairs of groups were compared by Student t test. Differencesbetween groups were considered significant at p , 0.05. Val-ues for all measurements are expressed as means 6 SD.
RESULTS
IGT induces expression of viral antigens
To examine whether IGT administration results in efficientexpression of the constituent RSV antigens, lung tissues of micewere examined for the presence of mRNA and proteins by RT-PCR and Western blot analysis, respectively. IGT-administeredmice show mRNA expression in the lung tissue within 3 days,as revealed by RT-PCR products from the corresponding ninedifferent cDNAs amplified with specific primers (Table 1 andFig. 1A). Evidence that these mRNAs are translated to producesufficient immunogens was obtained by testing pooled sera(n 5 4) of the IGT mice but not of the control (PBS-adminis-tered) mice; the pooled sera reacted with a number of RSV anti-gens present in RSV-infected murine lung homogenate in aWestern blot analysis (Fig. 1B). These results indicate that IGTinduces the production of RSV antigens, which elicit an anti-body response.
IGT is safe and effective
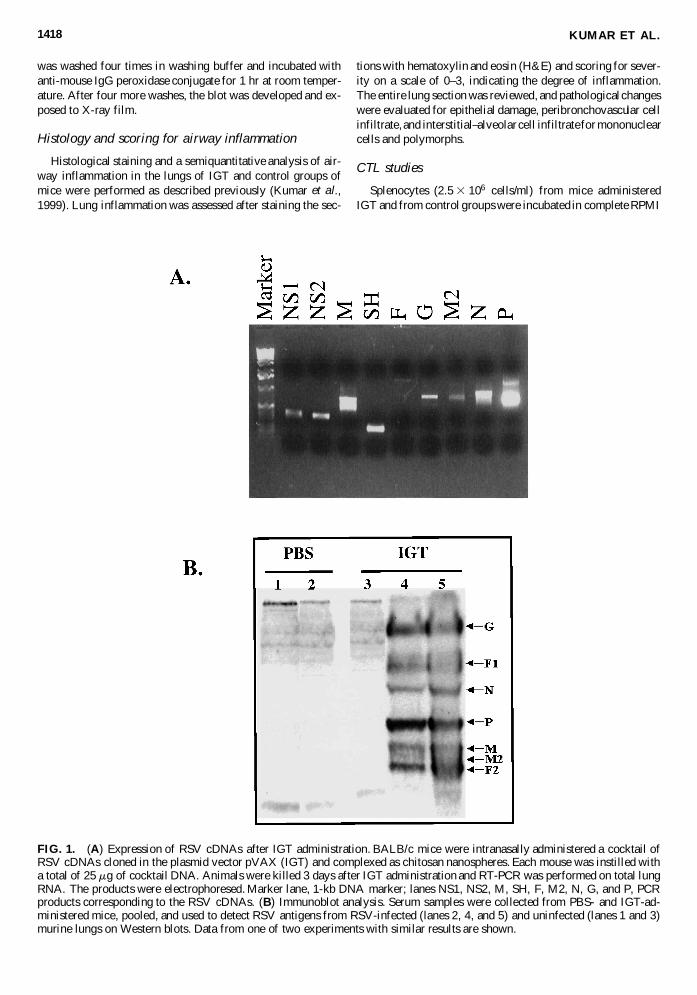
To test whether IGT administration induces airway hyperre-activity, the percent baseline enhanced pause (Penh) was mea-sured in different groups of mice. Mice receiving IGT exhibiteda similar response to methacholine challenge when comparedwith naked DNA, PBS alone (Fig. 2A), chitosan, and chitosanplus pVAX (data not shown) control groups. These results sug-gest that the IGT treatment by itself does not induce any sig-nificant change in airway hyperreactivity. To test the effective-ness of IGT, mice were administered a single dose of either IGT(a total of 25 mg of DNA), PBS, chitosan, empty plasmid vec-tor complexed as chitosan nanospheres (chitosan plus pVAX),or naked DNA in saline. Analysis of lung virus titers after acute,live RSV infection shows a significant (100-fold, p , 0.01) re-duction in RSV titers in IGT mice compared with controls (Fig.2B). Examination of total RSV antigen load in the immunizedand control groups of mice shows that immunized mice exhibita 77% decrease (p , 0.01) in antigen load when compared withPBS controls (Fig. 2C). These results indicate that IGT consti-tutes an effective prophylaxis against RSV infection.
GENE THERAPY FOR RSV INFECTION 1419
FIG. 2. IGT decreases viral titers and viral antigen load. (A)Determination of methacholine responsiveness of mice in-tranasally administered PBS, naked DNA, and IGT, as mea-sured by a whole-body plethysmograph. Methacholine respon-siveness was measured as percent baseline enhanced pause(Penh). The experiments were repeated with similar results. (Band C) Mice were administered IGT, naked DNA, chitosan pluspVAX, chitosan, or PBS as described previously, infected withRSV on day 16, and killed 4 days later (day 21). Lungs wereremoved and their homogenates were used for RSV plaque as-says and antigen loads. (B) RSV titers from lungs of mice in-tranasally administered IGT, naked DNA, chitosan plus pVAX,chitosan, or PBS. (C) Antigen load of RSV (measured byELISA) from lungs of mice intranasally administered IGT,naked DNA, chitosan plus pVAX, chitosan, or PBS. The ex-periment was repeated and similar results were obtained. Thevalues represent means 6 SD (n 5 4), compared with variouscontrol groups. **p , 0.01.
C.
KUMAR ET AL.1420
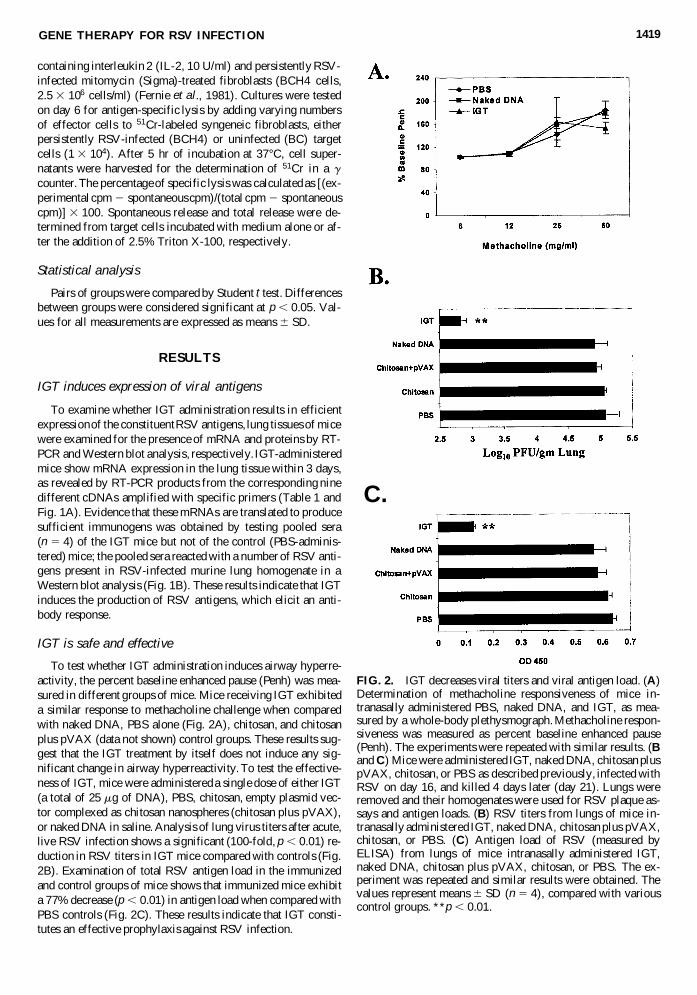
FIG. 3. Histological analysis of lung after gene transfer. Mice were treated as described previously, and infected with RSV onday 16. Four days later, these mice were killed, their lungs were removed, and histological sections were stained with hema-toxylin and eosin (H&E). IGT-administered mice (D) showed less epithelial damage and cellular infiltration than the chitosan(A), naked DNA (B), and chitosan plus pVAX (C) control groups. Arrows indicate epithelial damage and cellular infiltration.
IGT decreases RSV infection-induced pulmonary inflammation
Lung inflammation was examined in different groups ofmice. Mice treated with chitosan alone (Fig. 3A), chitosan pluspVAX (Fig. 3C), naked DNA (Fig. 3B), or PBS (data notshown) on acute RSV infection exhibit disruption of the ep-ithelium and cellular infiltration. Representative pathologicalfeatures reveal that groups of mice receiving IGT (Fig. 3D) ex-hibit less epithelial damage and reduced mononuclear cell(MNC) and polymorphonuclear cell (PMNC) infiltrates in theinterstitial and peribronchovascular regions, as compared withcontrols (Fig. 3A–C). These results suggest that IGT protectsmice from RSV infection-induced pulmonary inflammation. Asemiquantitative analysis using a scoring system for inflam-mation in the lung is shown in Table 2. Groups of mice that re-ceived IGT exhibit reduced epithelial damage (p , 0.01 com-pared with PBS; and p , 0.05, compared with other controlgroups) and pulmonary inflammation compared with controls.The group of mice that received IGT exhibit reduced (p , 0.001compared with PBS and p , 0.05 compared with chitosan and
chitosan plus pVAX) interstitial alveolar infiltrate and peri-bronchovascular infiltrate (p , 0.05 compared with PBS andchitosan plus pVAX). The scores for epithelial damage or in-filtration among the different control groups were not found tobe significant. These results suggest that IGT protects mice fromRSV infection-induced pulmonary inflammation.
IGT induces both serum and mucosal anti-RSVantibody response
Both serum and mucosal responses are important compo-nents of an effective gene transfer prophylaxis. Secreted IgAantibody is considered to be protective against mucosal patho-gens, as the nasal passage is the main site of entry for RSV(Godding et al., 2000). To examine whether IGT induces spe-cific antibodies in mice, RSV-specific antibody titers were mea-sured in IGT-administered and control mice. Animals adminis-tered IGT exhibit significantly higher serum IgG antibody titersthan controls (Fig. 4A). Incubation of RSV with the serum ob-tained from IGT mice reduces virus infection of HEp-2 cells,indicating the production of neutralizing antibodies after gene

transfer (Fig. 4B). IGT mice show significantly higher neutral-izing titers compared with control groups. The levels of IgAantibody in nasal wash were measured to verify whether thisclass of antibody was changed as a result of vaccination. Ani-mals administered IGT exhibit significantly higher RSV-spe-cific IgA antibody titers than controls (Fig. 4C). These resultsindicate that IGT induces increased production of neutralizingantibodies in serum and nasal IgA.
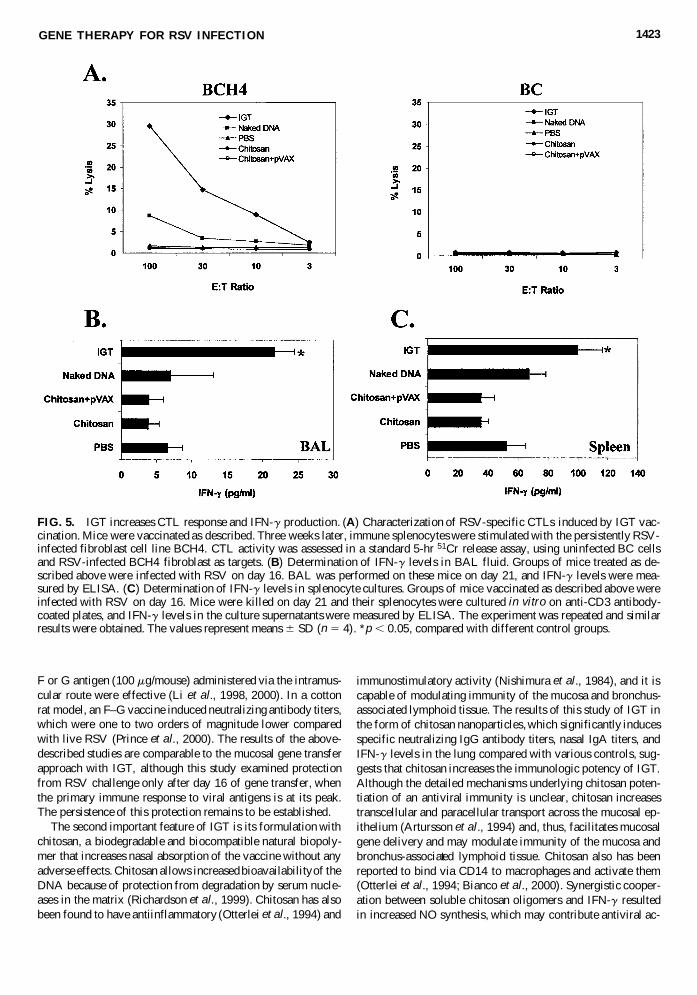
IGT generates RSV-specific CTL and IFN-g production
To test whether IGT also induces a CTL response, mice wereanalyzed for the presence of splenic, RSV-specific CTLs usingpersistently RSV-infected BCH4 cells as the target and RSV-negative BC cells as the control. PBS, naked DNA, chitosan,and empty vector complexed in chitosan nanosphere controlsdo not elicit a detectable CTL response. In contrast, mice ad-ministered IGT generate CTL responses (Fig. 5A), and theseCTLs are CD81 and MHC class I restricted (data not shown).IFN-g is considered to be the major antiviral cytokine. Thus,in order for a prophylaxis to be effective against RSV, it mustinduce IFN-g expression. IFN-g was assayed from the anti-CD3antibody-stimulated cultured spleen cells and the BAL fluid ofIGT-administered and control groups of mice. In both cases,IGT-administered mice exhibit significantly higher IFN-g pro-duction (Fig. 5B and C).
DISCUSSION
This study concerns the development of an effective and safeprophylaxis against RSV, utilizing a mucosal gene transfer ap-proach, which may provide for the protection of infants 2 to 6months of age, who are among the most susceptible to RSV in-fection. Furthermore, a mucosal gene transfer approach is con-sidered more appropriate for developing immunity in the lungsof these infants, who may have an immature immune system.The results of this study are deemed significant, because of thelack of a safe and effective vaccine against RSV. Currently,passive immunization with anti-RSV antibodies or with a hu-
manized antibody to the RSV-F antigen (Kneyber et al., 2000),at a monthly interval, is one of the options or often the onlyoption available to certain infants, who are at high risk of de-veloping RSV infection. These passive therapies are inconve-nient, expensive, and only partially effective.
The results demonstrate that the IGT utilized in this studyfor mucosal gene transfer is both safe and effective. The safetyis demonstrated by the lack of change in methacholine re-sponsiveness between vaccinated and control mice. IGT alsoinduced a significant decrease in overall lung inflammation ac-companying acute RSV infection, which presumably contrib-utes to the lung pathology in bronchiolitis and exacerbation ofasthma. This issue is important in view of the previous failureof the formalin-inactivated vaccine, which exacerbated the dis-ease (Chanock et al., 1992). The results of the semiquantitativeanalyses of epithelial damage and of perivascular, peri-bronchial, and interstitial infiltrating cells indicate that IGT sig-nificantly reduces cellular infiltration and epithelial damagecompared with naked DNA and unvaccinated mice. This dem-onstrates that IGT markedly attenuates inflammation while ren-dering protection against RSV infection. This is most likely dueto the effect of IGT on decreasing the virus titers, which lim-its the spread of infection and consequently results in less virus-induced inflammation.
IGT has two distinct components: a plasmid DNA cocktailconferring vaccine potency and chitosan conferring adjuvantactivity. Both of these components add intrinsic value to the ef-ficacy and safety of this vaccine. A core feature of IGT is thatit comprises a cocktail of pDNAs encoding all potentially im-munogenic RSV antigens, increasing the immune response and,thus, the efficacy of the vaccine. The mucosal gene transfer ap-proach has not previously been investigated for RSV. Differ-ences in composition, species, dosage, and route of adminis-tration preclude a direct comparison of results reported in thispaper with those published previously. Nonetheless, the resultsof this study show that a single dose of IGT, with a total of 25mg of DNA per mouse, given intranasally, induces a significantreduction (100-fold or 2 orders of magnitude) in viral titers af-ter an acute infection. Vaccines studied to date consist of a sub-unit, peptide, or DNA vaccine made up of the RSV-F, RSV-Gand/or RSV-M2 protein. In a mouse model, pDNAs encoding
GENE THERAPY FOR RSV INFECTION 1421
TABLE 2. SEMIQUANTITATIVE ANALYSIS OF LUNG PATHOLOGYa
Chitosan plus NakedLung pathology PBS Chitosan pVAX DNA IGT
Epithelial damage 2.53 6 0.17 2.24 6 0.12 2.39 6 0.16 2.25 6 0.30 1.4 6 0.52b–e
Interstitial–alveolar 2.66 6 0.21 2.45 6 0.35 2.47 6 0.19 2.36 6 0.33 1.76 6 0.35c,d,f
infiltratePeribronchovascular 2.01 6 0.20 1.89 6 0.25 2.16 6 0.11 1.81 6 0.57 1.46 6 0.23d,g
infiltrate
aEach value represents the mean 6 SD of five random fields from six individual lung sections from each mouse in a group(n 5 4).
bp , 0.01 compared with PBS control.cp , 0.05 compared with chitosan control.dp , 0.05 compared with chitosan plus pVax control.ep , 0.05 compared with naked DNA control.fp , 0.001 compared with PBS control.gp , 0.05 compared with PBS control.
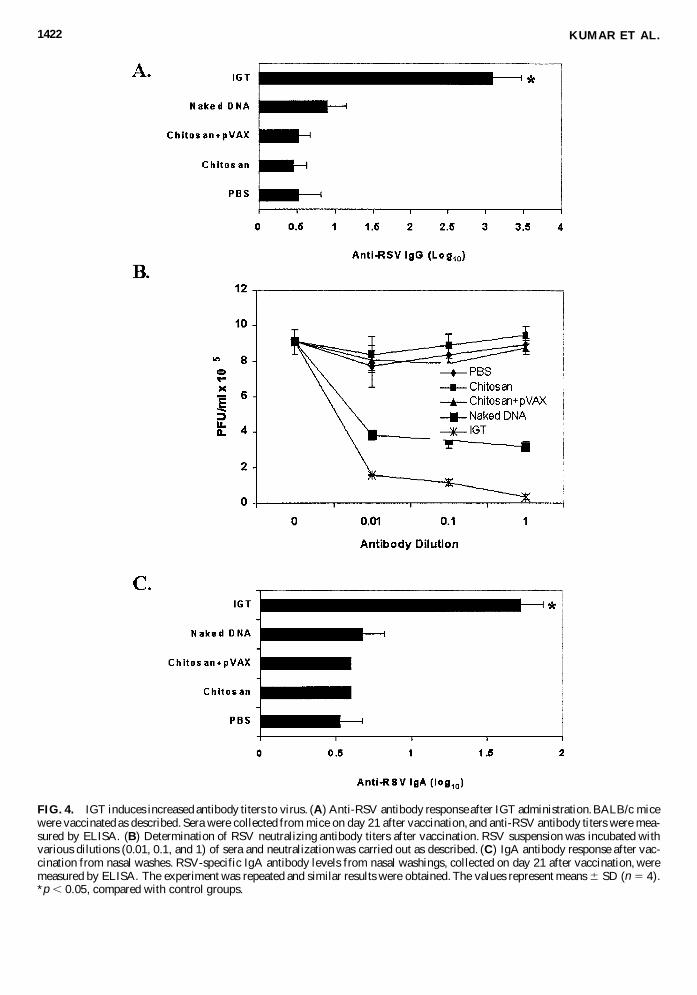
KUMAR ET AL.1422
FIG. 4. IGT induces increased antibody titers to virus. (A) Anti-RSV antibody response after IGT administration. BALB/c micewere vaccinated as described. Sera were collected from mice on day 21 after vaccination, and anti-RSV antibody titers were mea-sured by ELISA. (B) Determination of RSV neutralizing antibody titers after vaccination. RSV suspension was incubated withvarious dilutions (0.01, 0.1, and 1) of sera and neutralization was carried out as described. (C) IgA antibody response after vac-cination from nasal washes. RSV-specific IgA antibody levels from nasal washings, collected on day 21 after vaccination, weremeasured by ELISA. The experiment was repeated and similar results were obtained. The values represent means 6 SD (n 5 4).*p , 0.05, compared with control groups.
F or G antigen (100 mg/mouse) administered via the intramus-cular route were effective (Li et al., 1998, 2000). In a cottonrat model, an F–G vaccine induced neutralizing antibody titers,which were one to two orders of magnitude lower comparedwith live RSV (Prince et al., 2000). The results of the above-described studies are comparable to the mucosal gene transferapproach with IGT, although this study examined protectionfrom RSV challenge only after day 16 of gene transfer, whenthe primary immune response to viral antigens is at its peak.The persistence of this protection remains to be established.
The second important feature of IGT is its formulation withchitosan, a biodegradable and biocompatible natural biopoly-mer that increases nasal absorption of the vaccine without anyadverse effects. Chitosan allows increased bioavailability of theDNA because of protection from degradation by serum nucle-ases in the matrix (Richardson et al., 1999). Chitosan has alsobeen found to have antiinflammatory (Otterlei et al., 1994) and
immunostimulatory activity (Nishimura et al., 1984), and it iscapable of modulating immunity of the mucosa and bronchus-associated lymphoid tissue. The results of this study of IGT inthe form of chitosan nanoparticles, which significantly inducesspecific neutralizing IgG antibody titers, nasal IgA titers, andIFN-g levels in the lung compared with various controls, sug-gests that chitosan increases the immunologic potency of IGT.Although the detailed mechanisms underlying chitosan poten-tiation of an antiviral immunity is unclear, chitosan increasestranscellular and paracellular transport across the mucosal ep-ithelium (Artursson et al., 1994) and, thus, facilitates mucosalgene delivery and may modulate immunity of the mucosa andbronchus-associated lymphoid tissue. Chitosan also has beenreported to bind via CD14 to macrophages and activate them(Otterlei et al., 1994; Bianco et al., 2000). Synergistic cooper-ation between soluble chitosan oligomers and IFN-g resultedin increased NO synthesis, which may contribute antiviral ac-
GENE THERAPY FOR RSV INFECTION 1423
FIG. 5. IGT increases CTL response and IFN-g production. (A) Characterization of RSV-specific CTLs induced by IGT vac-cination. Mice were vaccinated as described. Three weeks later, immune splenocytes were stimulated with the persistently RSV-infected fibroblast cell line BCH4. CTL activity was assessed in a standard 5-hr 51Cr release assay, using uninfected BC cellsand RSV-infected BCH4 fibroblast as targets. (B) Determination of IFN-g levels in BAL fluid. Groups of mice treated as de-scribed above were infected with RSV on day 16. BAL was performed on these mice on day 21, and IFN-g levels were mea-sured by ELISA. (C) Determination of IFN-g levels in splenocyte cultures. Groups of mice vaccinated as described above wereinfected with RSV on day 16. Mice were killed on day 21 and their splenocytes were cultured in vitro on anti-CD3 antibody-coated plates, and IFN-g levels in the culture supernatants were measured by ELISA. The experiment was repeated and similarresults were obtained. The values represent means 6 SD (n 5 4). *p , 0.05, compared with different control groups.

tivity (Seo et al., 2000). The mechanism of chitosan-inducedimmune potentiation remains to be investigated.
In an effort to unravel the protective mechanisms underly-ing the efficacy of IGT, induced humoral and cellular immu-nity was investigated. A notable finding is that IGT signifi-cantly augmented levels of both neutralizing serum and mucosalIgA antibodies compared with naked DNA-vaccinated and un-vaccinated control groups. Previously, passive administrationof neutralizing serum antibodies was shown to decrease the riskof RSV disease in animal models and in humans (Groothuis etal., 1991; Hemming et al., 1995). Although the secreted IgAantibody provides protection against pathogens that enter viathe mucosal route (Godding et al., 2000), the role of secretoryIgA in protection against RSV is poorly understood. BecauseRSV is an obligatory intracellular mucosal pathogen affectingboth the upper and lower respiratory tract, it is likely that mu-cosal IgA might provide protection against severe RSV diseaseby precluding its entry into mucosa and/or inhibiting thecell–cell syncytial spread of RSV (Mazanec et al., 1992; Bom-sel et al., 1998). The protective role of IgA requires further in-vestigation.
Virus-specific CTLs play a major role in the clearance ofRSV infection (Graham et al., 1991). Passive transfer of RSV-specific T cells has been shown to effectively clear RSV (Can-non et al., 1987). The results reported in this paper demonstratethat IGT delivery generates a significantly stronger CTL re-sponse compared with naked DNA and other controls. Theseresults, which are in agreement with other experimental ap-proaches, support a role for vaccine-induced CTLs in virusclearance. Several studies indicate that the protective effect ofCTLs against cytopathic viruses is dependent on their produc-tion of cytokines such as IFN-g (Hsu et al., 1998; Okamoto etal., 1999). Indeed, IGT significantly enhanced the productionof IFN-g, which may be useful in fighting RSV infection. IFN-g has a direct antiviral effect and is particularly important instimulating the cytolytic activity of natural killer (NK) cells andCD81 CTLs, which play a critical role in the control of RSVinfection in a murine model (Hsu et al., 1998) and in humans(Aberle et al., 1999).
Collectively, these data demonstrate that IGT represents anovel gene transfer approach against RSV infection, which, ata single dose of about 1 mg/kg body weight, is capable of de-creasing viral titers by 2 orders of magnitude (100-fold) on pri-mary infection. The immunologic mechanisms for the effec-tiveness of this prophylaxis include the induction of high levelsof both serum IgG and mucosal IgA antibodies, the generationof an effective CTL response, and elevated lung-specific pro-duction of IFN-g with antiviral action. Although as a single-dose vaccine IGT is effective, it is possible that dose escalationand prime–booster strategies might further enhance its effec-tiveness. In addition, IGT significantly decreases pulmonary in-flammation and does not alter airway hyperresponsiveness, thusmaking it a safe vaccine.
ACKNOWLEDGMENTS
This study was supported by the grants from the VA MeritReview Award and the American Heart Association, FloridaAffiliate award to S.S.M. and by generous support from the Joy
McCann Culverhouse endowment to the Division of Allergyand Immunology.
REFERENCES
ABERLE, J.H., ABERLE, S.W., ALLISON, S.L., STIASNY, K.,ECKER, M., MANDL, C.W., BERGER, R., and HEINZ, F.X.(1999). A DNA immunization model study with constructs express-ing the tick-borne encephalitis virus envelope protein E in differentphysical forms. J. Immunol. 163, 6756–6761.
ARTURSSON, P., LINDMARK, T., DAVIS, S.S., and ILLUM, L.(1994). Effect of chitosan on the permeability of monolayers of in-testinal epithelial cells (Caco-2). Pharm. Res. 11, 1358–1361.
BEHERA, A.K., KUMAR, M., MATSUSE, H., LOCKEY, R.F., andMOHAPATRA, S.S. (1998). Respiratory syncytial virus induces theexpression of 5-lipoxygenase and endothelin-1 in bronchial epithe-lial cells. Biochem. Biophys. Res. Commun. 251, 704–709.
BEHERA, A.K., MATSUSE, H., KUMAR, M., KONG, X., LOCKEY,R.F., and MOHAPATRA, S.S. (2001). Blocking intercellular adhe-sion molecule-I on human epithelial cells decreases respiratory syn-cytial virus infection. Biochem. Biophys. Res. Commun. 280, 188–195.
BIANCO, I.D., BALSINDE, J., BELTRAMO, D.M., CASTAGNA,L.F., LANDA, C.A., and DENNIS, E.A. (2000). Chitosan-inducedphospholipase A2 activation and arachidonic acid mobilization inP388D1 macrophages. FEBS Lett. 466, 292–294.
BOMSEL, M., HEYMAN, M., HOCINI, H., LAGAYE, S., BELEC,L., DUPONT, C., and DESGRANGES, C. (1998). Intracellular neu-tralization of HIV transcytosis across tight epithelial barriers by anti-HIV envelope protein dIgA or IgM. Immunity 9, 277–287.
BRANDENBURG, A.H., NEIJENS, H.J., and OSTERHAUS, A.D.(2001). Pathogenesis of RSV lower respiratory tract infection: Im-plications for vaccine development. Vaccine 19, 2769–2782.
CANNON, M.J., STOTT, E.J., TAYLOR, G., and ASKONAS, B.A.(1987). Clearance of persistent respiratory syncytial virus infectionsin immunodeficient mice following transfer of primed T cells. Im-munology 62, 133–138.
CENTERS FOR DISEASE CONTROL AND PREVENTION (1999).Update: Respiratory syncytial virus activity—United States, 1998–1999 season. MMWR 48, 1104.
CHANOCK, R.M., PARROTT, R.H., CONNORS, M., COLLINS,P.L., and MURPHY, B.R. (1992). Serious respiratory tract diseasecaused by respiratory syncytial virus: Prospects for improved ther-apy and immunization. Pediatrics 90, 137–143.
COHEN, A.D., BOYER, J.D., and WEINER, D.B. (1998). Modulatingthe immune response to genetic immunization. FASEB J. 12, 1611–1626.
CONNORS, M., COLLINS, P.L., FIRESTONE, C.Y., and MURPHY,B.R. (1991). Respiratory syncytial virus (RSV) F, G, M2 (22K), andN proteins each induce resistance to RSV challenge, but resistanceinduced by M2 and N proteins is relatively short-lived. J. Virol. 65,1634–1637.
DOMACHOWSKE, J.B., and BONVILLE, C.A. (1998). Overnighttitration of human respiratory syncytial virus using quantitative shellvial amplification. Biotechniques 25, 644–647.
DOMACHOWSKE, J.B., and ROSENBERG, H.F. (1999). Respiratorysyncytial virus infection: Immune response, immunopathogenesis,and treatment. Clin. Microbiol. Rev. 12, 298–309.
DONNELLY, J.J., ULMER, J.B., and LIU, M.A. (1998). DNA vac-cines. Dev. Biol. Stand. 95, 43–53.
FERNIE, B.F., FORD, E.C., and GERIN, J.L. (1981). The develop-ment of BALB/c cells persistently infected with respiratory syncy-tial virus: Presence of ribonucleoprotein on the cell surface. Proc.Soc. Exp. Biol. Med. 167, 83–86.
KUMAR ET AL.1424

GODDING, V., VAERMAN, J.P., and SIBILLE, Y. (2000). Secretorymucosal immune mechanisms. Acta Otorhinolaryngol. Belg. 54,255–261.
GRAHAM, B.S., BUNTON, L.A., WRIGHT, P.F., and KARZON, D.T.(1991). Role of T lymphocyte subsets in the pathogenesis of primaryinfection and rechallenge with respiratory syncytial virus in mice. J.Clin. Invest. 88, 1026–1033.
GROOTHUIS, J.R., LEVIN, M.J., RODRIGUEZ, W., HALL, C.B.,LONG, C.E., KIM, H.W., LAUER, B.A., and HEMMING, V.G.(1991). Use of intravenous gamma globulin to passively immunizehigh-risk children against respiratory syncytial virus: Safety andpharmacokinetics. RSVIG Study Group. Antimicrob. Agents Chemo-ther. 35, 1469–1473.
GUY, B., PASCAL, N., FRANCON, A., BONNIN, A., GIMENEZ, S.,LAFAY-VIALON, E., TRANNOY, E., and HAENSLER, J. (2001).Design, characterization and preclinical efficacy of a cationic lipidadjuvant for influenza split vaccine. Vaccine 19, 1794–1805.
HALL, C.B. (1994). Prospects for a respiratory syncytial virus vaccine.Science 265, 1393–1394.
HEMMING, V.G., PRINCE, G.A., GROOTHUIS, J.R., and SIBER,G.R. (1995). Hyperimmune globulins in prevention and treatment ofrespiratory syncytial virus infections. Clin. Microbiol. Rev. 8, 22–33.
HSU, S.C., OBEID, O.E., COLLINS, M., IQBAL, M., CHARGELE-GUE, D., and STEWARD, M.W. (1998). Protective cytotoxic T lym-phocyte responses against paramyxoviruses induced by epitope-based DNA vaccines: Involvement of IFN-g. Int. Immunol. 10,1441–1447.
ILLUM, L., JABBAL-GILL, I., HINCHCLIFFE, M., FISHER, A.N.,and DAVIS, S.S. (2001). Chitosan as a novel nasal delivery systemfor vaccines. Adv. Drug Deliv. Rev. 51, 81–96.
KNEYBER, M.C., MOLL, H.A., and DE GROOT, R. (2000). Treat-ment and prevention of respiratory syncytial virus infection. Eur. J.Pediatr. 159, 399–411.
KUMAR, M., BEHERA, A.K., MATSUSE, H., LOCKEY, R.F., andMOHAPATRA, S.S. (1999). Intranasal IFN-g gene transfer protectsBALB/c mice against respiratory syncytial virus infection. Vaccine18, 558–567.
LI, X., SAMBHARA, S., LI, C.X., EWASYSHYN, M., PARRING-TON, M., CATERINI, J., JAMES, O., CATES, G., DU, R.P., andKLEIN, M. (1998). Protection against respiratory syncytial virus in-fection by DNA immunization. J. Exp. Med. 188, 681–688.
LI, X., SAMBHARA, S., LI, C.X., ETTORRE, L., SWITZER, I.,CATES, G., JAMES, O., PARRINGTON, M., OOMEN, R., DU,R.P., and KLEIN, M. (2000). Plasmid DNA encoding the respiratorysyncytial virus G protein is a promising vaccine candidate. Virology269, 54–65.
MATSUO, K., MACLAUGHLIN, F.C., MUMPER, R.J., WANG, J.,TAGLIAFERRI, J.M., GILL, I., HINCHCLIFFE, M., and ROL-LAND, A.P. (2000). Induction of innate immunity by nasal influenzavaccine administered in combination with an adjuvant (choleratoxin). Vaccine 18, 2713–2722.
MATSUSE, H., BEHERA, A.K., KUMAR, M., RABB, H., LOCKEY,R.F., and MOHAPATRA, S.S. (2000). Recurrent respiratory syncy-tial virus infections in allergen-sensitized mice lead to persistent air-way inflammation and hyperresponsiveness. J. Immunol. 164,6583–6592.
MAZANEC, M.B., KAETZEL, C.S., LAMM, M.E., FLETCHER, D.,and NEDRUD, J.G. (1992). Intracellular neutralization of virus by
immunoglobulin A anti-bodies. Proc. Natl. Acad. Sci. U.S.A. 89,6901–6905.
NICHOLAS, J.A., RUBINO, K.L., LEVELY, M.E., ADAMS, E.G.,and COLLINS, P.L. (1990). Cytolytic T-lymphocyte responses torespiratory syncytial virus: Effector cell phenotype and target pro-teins. J. Virol. 64, 4232–4241.
NISHIMURA, K., NISHIMURA, S., NISHI, N., SAIKI, I., TOKURA,S., and AZUMA, I. (1984). Immunological activity of chitin and itsderivatives. Vaccine 2, 93–99.
OKAMOTO, I., KOHNO, K., TANIMOTO, T., IKEGAMI, H., andKURIMOTO, M. (1999). Development of CD81 effector T cells isdifferentially regulated by IL-18 and IL-12. J. Immunol. 162, 3202–3211.
OTTERLEI, M., VARUM, K.M., RYAN, L., and ESPEVIK, T. (1994).Characterization of binding and TNF-a inducing ability of chitosanson monocytes: The involvement of CD14. Vaccine 12, 825–832.
PRINCE, G.A., CAPIAU, C., DESCHAMPS, M., FABRY, L., GAR-CON, N., GHEYSEN, D., PRIEELS, J.P., THIRY, G., VAN OP-STAL, O., and PORTER, D.D. (2000). Efficacy and safety studiesof a recombinant chimeric respiratory syncytial virus FG glycopro-tein vaccine in cotton rats. J. Virol. 74, 10287–10292.
RICHARDSON, S.C., KOLBE, H.V., and DUNCAN, R. (1999). Po-tential of low molecular mass chitosan as a DNA delivery system:Biocompatibility, body distribution and ability to complex and pro-tect DNA. Int. J. Pharm. 178, 231–243.
ROY, K., MAO, H.Q., HUANG, S.K., and LEONG, K.W. (1999). Oralgene delivery with chitosan–DNA nanoparticles generates immuno-logic protection in a murine model of peanut allergy. Nat. Med. 5,387–391.
SEO, W.G., PAE, H.O., KIM, N.Y., OH, G.S., PARK, I.S., KIM, Y.H.,KIM, Y.M., LEE, Y.H., JUN, C.D., and CHUNG, H.T. (2000). Syn-ergistic cooperation between water-soluble chitosan oligomers andinterferon-g for induction of nitric oxide synthesis and tumoricidalactivity in murine peritoneal macrophages. Cancer Lett. 159,189–195.
WYATT, L.S., WHITEHEAD, S.S., VENANZI, K.A., MURPHY,B.R., and MOSS, B. (1999). Priming and boosting immunity to res-piratory syncytial virus by recombinant replication-defective vac-cinia virus MVA. Vaccine 18, 392–397.
Address reprint requests to:Dr. Shyam S. Mohapatra
Joy McCann Culverhouse Airway Disease CenterDivision of Allergy and Immunology, Department of
Internal MedicineUniversity of South Florida and VA Hospital
12901 Bruce B. Downs Blvd.Tampa, FL 33612
E-mail: [email protected]
Received for publication February 1, 2002; accepted after re-vision June 17, 2002.
Published online: July 17, 2002.
GENE THERAPY FOR RSV INFECTION 1425

















![Slamf1 -/- [ BALB/c.129]](https://img.pdfslide.us/doc/110x75/56815051550346895dbe5296/slamf1-balbc129.jpg)