Embed Size (px)
Citation preview

Intermediate-Conductance Ca2�-Activated K� Channel,KCa3.1, as a Novel Therapeutic Target for Benign ProstaticHyperplasia□S
Susumu Ohya, Satomi Niwa, Yoshiyuki Kojima, Shoichi Sasaki, Motomu Sakuragi,Kenjiro Kohri, and Yuji ImaizumiDepartment of Molecular and Cellular Pharmacology, Graduate School of Pharmaceutical Sciences (S.O., S.N., Y.I.),Department of Nephro-Urology, Graduate School of Medical Sciences (Y.K., S.S., K.K.), Nagoya City University, Nagoya,Japan; and Hanno Research Center, Taiho Pharmaceutical Co., Ltd., Saitama, Japan (M.S.)
Received April 7, 2011; accepted May 19, 2011
ABSTRACTRecently, a new experimental stromal hyperplasia animalmodel corresponding to clinical benign prostatic hyperplasia(BPH) was established. The main objective of this study was toelucidate the roles of the intermediate-conductance Ca2�-ac-tivated K� channel (KCa3.1) in the implanted urogenital sinus(UGS) of stromal hyperplasia BPH model rats. Using DNA mi-croarray, real-time polymerase chain reaction, Western blot,and/or immunohistochemical analyses, we identified the ex-pression of KCa3.1 and its transcriptional regulators in im-planted UGS of BPH model rats and prostate needle-biopsysamples and surgical prostate specimens of BPH patients. Wealso examined the in vivo effects of a KCa3.1 blocker, 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole (TRAM-34), on theproliferation index of implanted UGS by measurement of UGSweights and proliferating cell nuclear antigen immunostaining.
KCa3.1 genes and proteins were highly expressed in implantedUGS rather than in the normal host prostate. In the implantedUGS, the gene expressions of two transcriptional regulators ofKCa3.1, repressor element 1-silencing transcription factor andc-Jun, were significantly down- and up-regulated, and the reg-ulations were correlated negatively or positively with KCa3.1expression, respectively. Positive signals of KCa3.1 proteinswere detected exclusively in stromal cells, whereas they werescarcely immunolocalized to basal cells of the epithelium inimplanted UGS. In vivo treatment with TRAM-34 significantlysuppressed the increase in implanted UGS weights comparedwith the decrease in stromal cell components. Moreover, sig-nificant levels of KCa3.1 expression were observed in humanBPH samples. KCa3.1 blockers may be a novel treatment optionfor patients suffering from BPH.
IntroductionBenign prostatic hyperplasia (BPH) is noncancerous en-
largement of the prostate gland with aging. BPH is demon-strated in 50% of men by the age of 60 years and is evident inup to 90% of men by 85 years (Berry et al., 1984). BPH causesincreased resistance to urine flow through the urethra andsometimes kidney damage, bladder stones, and urinary tract
infections and thereby affects the quality of life in one thirdof men older than 50 years (Djavan, 2003). �1-Adrenoceptorantagonists are used clinically to decrease the periurethraltone of prostatic smooth muscle (Andersson and Gratzke,2007) and can regulate prostatic growth by inducing apopto-sis of epithelial, stromal, and smooth muscle cells, but theirlong-term use can induce therapeutic tolerance (Kojima etal., 2009a,b). Antiandrogen therapy or surgery is used toresolve mechanical outlet obstructions (Pollard et al., 1989).
BPH is a proliferative process of both stromal and epithe-lial elements of the prostate arising in the periurethral andtransition zones of the gland and is hormonally dependent ontestosterone and dihydrotestosterone production (Berry etal., 1984). The major component of clinical BPH specimens isstromal extracellular matrix, which is organized by smoothmuscle, fibrous tissue element, and collagen (Shapiro et al.,
This work was supported by Grants-in Aid for Scientific Research from theJapan Society for the Promotion of Science [Grants 20390027, 21590098] (toY.I. and S.O., respectively); Takeda Science Foundation (to S.O.); and SaltScience Research Foundation [Grant 10C6] (to S.O.).
Article, publication date, and citation information can be found athttp://jpet.aspetjournals.org.
doi:10.1124/jpet.111.182782.□S The online version of this article (available at http://jpet.aspetjournals.org)
contains supplemental material.
ABBREVIATIONS: BPH, benign prostate hyperplasia; KCa3.1, intermediate-conductance Ca2�-activated K� channel; VSMC, vascular smoothmuscle cell; REST, repressor element 1-silencing transcription factor; UGS, urogenital sinus; AP-1, activator protein-1; PCR, polymerase chainreaction; bp, base pair; KCa, Ca2�-activated K� channel; IKCa, intermediate-conductance KCa channel; PCNA, proliferating cell nuclear antigen;TRAM-34, 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole; KCa1.1, large-conductance Ca2�-activated K� channel.
0022-3565/11/3382-528–536$25.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 338, No. 2Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics 182782/3705132JPET 338:528–536, 2011 Printed in U.S.A.
528
http://jpet.aspetjournals.org/content/suppl/2011/05/20/jpet.111.182782.DC1Supplemental material to this article can be found at:
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from

1992); however, this feature is histologically different fromthe testosterone-induced rodent BPH model (Ishigooka et al.,1996), and this animal BPH model is not adequate to eluci-date the pathological mechanisms of stromal hyperplasia.Mori et al. (2009) recently established a new experimentalrodent model characterized by stromal hyperplasia. Urogen-ital sinuses implanted into the ventral prostate, “implantedUGS,” contain a much higher ratio of stroma than the age-matched ventral prostate and the testosterone-induced BPHmodel and are similar to those in men with BPH.
Ca2�-activated K� channel (KCa) can communicate di-rectly from Ca2� signal pathways to changes in membranepotential required for various cellular processes. In excitablecells, KCa activation contributes to the posthyperpolarizationthat follows an action potential and generally suppressesmembrane excitability, whereas in nonexcitable cells hyper-polarization by KCa activation increases the driving force forCa2� entry, strongly associating with sustained Ca2� influx(Stocker, 2004). Intermediate-conductance KCa (IKCa) ispresent in undifferentiated vascular and urinary bladdersmooth muscles and participates in the control of cellularfunctions such as cell proliferation and maintenance of rest-ing tone (Ohya et al., 2000; Kohler et al., 2003). IKCa has beenmolecularly identified as KCa3.1 (Ishii et al., 1997) and is apossible molecular target for pharmacological intervention inrestenosis, urinary incontinence, prostate cancer, and auto-immune diseases (Kohler et al., 2003; Wulff et al., 2007;Ohya et al., 2009). In addition, at least two transcriptionalregulators of KCa3.1 have been identified in vascular smoothmuscle cells (VSMCs) and T lymphocytes: repressor ele-ment-1 silencing transcription factor (REST) and nucleartranscription factor activator protein-1 (AP-1) (Fos/Jun)(Ghanshani et al., 2000; Cheong et al., 2005).
The main objective of this study was to elucidate the pos-sibility that IKCa can be a potential therapeutic target inhuman BPH using a stromal hyperplasia BPH model. Ex-pressions of KCa3.1 genes and proteins in implanted UGS ofstromal hyperplasia BPH model rats were compared withthose of the normal host prostate by DNA microarray, real-time polymerase chain reaction (PCR), Western blot, andimmunohistochemical analyses. We also determined whetherimplanted UGS weights were suppressed by treatment with1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole (TRAM-34),a specific IKCa inhibitor. Similar examinations were per-formed using human prostate needle-biopsy samples andsurgical prostate specimens of BPH patients.
Materials and MethodsRNA Extraction, Reverse-Transcription PCR, and Real-
Time PCR. Experimental model rats for BPH with stromal hyper-plasia were prepared as reported previously (Mori et al., 2009).Implanted UGS and normal host prostate in stromal hyperplasiaBPH model rats were dissected 3 weeks after implantation. All of theexperiments were carried out in accordance with the guiding princi-ples for the care and use of laboratory animals (the Science andInternational Affairs Bureau of the Japanese Ministry of Education,Science, Sports and Culture) and also with the approval of the ethicscommittee of Nagoya City University and Taiho Pharmaceutical Co.Total RNA from human normal prostate was purchased from BDBiosciences (San Jose, CA) and BioChain (Hayward, CA) (21–50years old, three distinct lot number samples). Total RNA extractionand reverse transcription were performed as reported previously
(Ohya et al., 2009). We enrolled seven patients with BPH 54 to 86years old (mean age � 70.4 � 4.5 years). Prostate needle-biopsysamples and surgical prostate specimens of BPH patients were ob-tained with informed consent from all of the patients before thestudy after explaining the purpose and methods. The study wasapproved by the ethics committee of Nagoya City University. BPHwas diagnosed on the basis of the International Prostatic SymptomScore, the quality of life index, digital rectal examination, ultra-sonography, uroflowmetry, and prostate needle biopsy. The resultingcDNA product was amplified with gene-specific primers, designedusing Primer Express software (version 1.5; Applied Biosystems,Foster City, CA).
Quantitative real-time PCR performed with the use of SYBRGreen chemistry on an ABI 7700 sequence detector system (AppliedBiosystems) as reported previously (Ohya et al., 2009). The followingPCR primers for rat clones were used for real-time PCR: KCa3.1(NM_023021, 837–937), 101 base pair (bp); REST (NM_031788,2367–2489), 123 bp; c-Fos (NM_022197, 897-1025), 129 bp; FosB(NM_001013146, 1075–1209), 135 bp; Fra-1 (NM_012953, 848–968),121 bp; Fra-2 (NM_012954, 99–227), 129 bp; c-Jun (NM_021835,1195–1317), 123 bp; JunB (NM_021836, 357–492), 136 bp; JunD(NM_138875, 1036–1160), 125 bp; �-actin (NM_031144, 419–519),101 bp. The following PCR primers for human clones wereused: KCa3.1 (NM_002250, 172–293), amplicon � 122 bp; REST(NM_005612, 1415–1545), 131 bp; c-Fos (NM_005252, 533–658), 126bp; FosB (NM_006732, 1075–1209), 135 bp; Fra-1 (X16707, 680–781), 102 bp; Fra-2 (X16706, 102–226), 125 bp; c-Jun (NM_002228,1851–1999), 149 bp; JunB (NM_002229, 271–391), 121 bp; JunD(NM_005354, 1027–1134), 108 bp; �-actin (NM_001101, 411–511),amplicon � 101 bp. Regression analyses of the mean values of threemultiplex real-time PCRs for log10-diluted cDNA were used to gen-erate standard curves. Unknown quantities relative to the standardcurve for a particular set of primers were calculated, yielding tran-scriptional quantification of gene products relative to the endoge-nous standard �-actin. For PCR cloning of full-length KCa3.1, thefollowing PCR primers were used: KCa3.1 (NM_023021, 77–1459),1383 bp. To confirm the nucleotide sequences, amplified PCR prod-ucts and plasmid constructs were sequenced with an ABI PRISM3100 genetic analyzer (Applied Biosystems).
Genome-Wide DNA Microarray Analysis. To determine thequantity and purity of the RNA samples, purified DNase-free RNAfrom normal host prostate and implanted UGS was analyzed on aND-1000 spectrophotometer (NanoDrop Technologies, Wilmington,DE) and a 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA).We then used 12 Affymetrix (Santa Clara, CA) Rat Genome 230 2.0arrays. These microarrays contain �31,000 genes. Labeling, hybrid-ization (for 16 h), washing, and scanning of the microarray wereperformed by Bio Matrix Research (Chiba, Japan) following themanufacturer’s specifications (i.e., One-Cycle Target Labeling kit).The arrays were scanned on the GCS 3000 Affymetrix high-resolu-tion scanner and analyzed using GeneChip Operating Software ver-sion 1.4 (Affymetrix) and GeneSpring version 7.3.1 (Agilent Technol-ogies). Gene expression data were normalized by preparing“Per-Chip_Only” and were confirmed by Pearson correlation. Com-parative analysis among expression profiles of two samples wascarried out using GeneSpring software. Only gene expression levelswith statistical significance (p � 0.05) were recorded as being “pres-ent” above background levels, and genes with expression levels belowthis statistical threshold were considered as “absent,” as reportedpreviously (Kojima et al., 2009a).
Western Blot Analysis. The plasma membrane protein fractionwas prepared from rat tissues as reported previously (Ohya et al.,2005), and the nuclear protein fraction was prepared using aProteoJET Cytoplasmic and Nuclear Protein Extraction kit (Fer-mentas UAB, Vilnius, Lithuania). Equal amounts of proteins weresubjected to SDS-polyacrylamide gel electrophoresis (10%). Blotswere incubated with anti-KCa3.1 (Alomone Labs, Jerusalem, Israel),anti-REST (Millipore Corporation, Billerica, MA), and anti-c-Jun
KCa3.1 as a Novel Therapeutic Target for BPH 529
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from
antibodies (Cell Signaling Technology, Danvers, MA), and then in-cubated with anti-rabbit horseradish peroxidase-conjugated IgG(Millipore Corporation). An enhanced chemiluminescence detectionsystem (GE Healthcare Japan, Tokyo, Japan) was used for detectionof the bound antibody. The resulting images were analyzed by aLAS-1000 device (Fujifilm, Tokyo, Japan).
Immunohistochemical Analysis. The implanted UGS fromBPH model rats (4 weeks after implantation) and surgical prostatespecimens from BPH patients were fixed in 10% neutral bufferedformalin, embedded in paraffin, and thin-sectioned at 2 �m (Mori etal., 2009). Tissue sections were deparaffinized in xylene followed byrehydration in graded alcoholic solutions and phosphate-bufferedsaline. Endogenous peroxidase activity was blocked with 0.3% hy-drogen peroxide in methanol for 30 min at 4°C followed by washingwith phosphate-buffered saline. Three different anti-KCa3.1 antibod-ies [APC-064 (Alomone Labs), PAB7986 (Abnova, Taipei, Taiwan),and ab83740 (Abcam plc, Cambridge, UK)] were used for immuno-staining. All of the sections were developed by the avidin-biotincomplex method. The sections also were stained with hematoxylinand eosin staining and Masson’s trichrome. Microscopic images ofeach section were captured on a computer and analyzed with animage analysis system (Win ROOF; Tech-Jam, Osaka, Japan).
Measurement of Cell Proliferation by Proliferating CellNuclear Antigen Immunohistochemistry. To examine the pro-liferation of stromal and epithelial compartments of implanted UGSafter treatment with TRAM-34 (Sigma-Aldrich, St. Louis, MO), theexpression of proliferating cell nuclear antigen (PCNA) was analyzedas reported previously (Mori et al., 2009). After sectioning and stain-ing, PCNA positive and negative cells were counted in 10 separatemicroscopic fields for each graft. The percentage of PCNA-positivecells of total epithelium cells and stromal cells was calculated as thePCNA labeling index.
In Vivo Proliferation Assay. To examine the effect of a selectiveIKCa blocker TRAM-34 (150, 300, 600, and 1200 �g/kg) on prostaticgrowth in BPH model rats, the in vivo proliferation assay was per-formed as reported previously (Kojima et al., 2009a). The compoundwas administered subcutaneously 7 to 21 days after UGS implanta-tion (150, 300, 600, and 1200 �g/kg s.c.). Vehicle was given in thesame way. On day 21, the implanted UGS were collected andweighed and then fixed in formalin and embedded in paraffin.
Statistical Analysis. Statistical significance between two groupsor among multiple groups was evaluated using Welch’s t test orTukey’s test after the F test or analysis of variance. Data are pre-sented as the means � S.E.M.
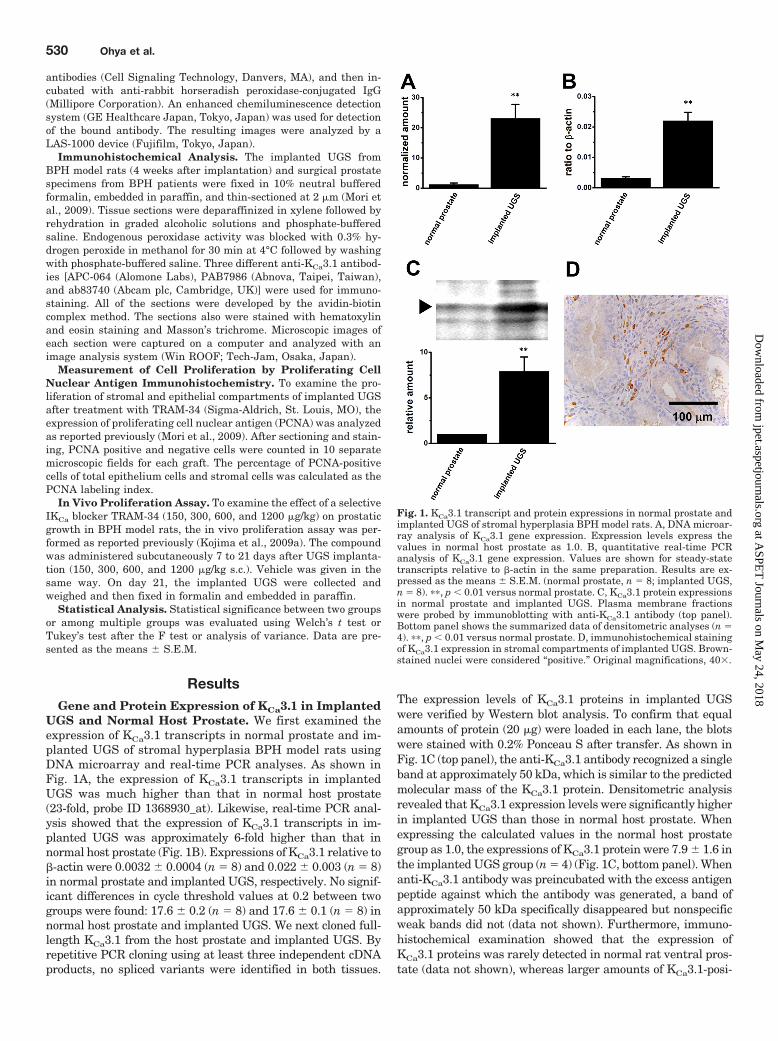
ResultsGene and Protein Expression of KCa3.1 in Implanted
UGS and Normal Host Prostate. We first examined theexpression of KCa3.1 transcripts in normal prostate and im-planted UGS of stromal hyperplasia BPH model rats usingDNA microarray and real-time PCR analyses. As shown inFig. 1A, the expression of KCa3.1 transcripts in implantedUGS was much higher than that in normal host prostate(23-fold, probe ID 1368930_at). Likewise, real-time PCR anal-ysis showed that the expression of KCa3.1 transcripts in im-planted UGS was approximately 6-fold higher than that innormal host prostate (Fig. 1B). Expressions of KCa3.1 relative to�-actin were 0.0032 � 0.0004 (n � 8) and 0.022 � 0.003 (n � 8)in normal prostate and implanted UGS, respectively. No signif-icant differences in cycle threshold values at 0.2 between twogroups were found: 17.6 � 0.2 (n � 8) and 17.6 � 0.1 (n � 8) innormal host prostate and implanted UGS. We next cloned full-length KCa3.1 from the host prostate and implanted UGS. Byrepetitive PCR cloning using at least three independent cDNAproducts, no spliced variants were identified in both tissues.
The expression levels of KCa3.1 proteins in implanted UGSwere verified by Western blot analysis. To confirm that equalamounts of protein (20 �g) were loaded in each lane, the blotswere stained with 0.2% Ponceau S after transfer. As shown inFig. 1C (top panel), the anti-KCa3.1 antibody recognized a singleband at approximately 50 kDa, which is similar to the predictedmolecular mass of the KCa3.1 protein. Densitometric analysisrevealed that KCa3.1 expression levels were significantly higherin implanted UGS than those in normal host prostate. Whenexpressing the calculated values in the normal host prostategroup as 1.0, the expressions of KCa3.1 protein were 7.9 � 1.6 inthe implanted UGS group (n � 4) (Fig. 1C, bottom panel). Whenanti-KCa3.1 antibody was preincubated with the excess antigenpeptide against which the antibody was generated, a band ofapproximately 50 kDa specifically disappeared but nonspecificweak bands did not (data not shown). Furthermore, immuno-histochemical examination showed that the expression ofKCa3.1 proteins was rarely detected in normal rat ventral pros-tate (data not shown), whereas larger amounts of KCa3.1-posi-
Fig. 1. KCa3.1 transcript and protein expressions in normal prostate andimplanted UGS of stromal hyperplasia BPH model rats. A, DNA microar-ray analysis of KCa3.1 gene expression. Expression levels express thevalues in normal host prostate as 1.0. B, quantitative real-time PCRanalysis of KCa3.1 gene expression. Values are shown for steady-statetranscripts relative to �-actin in the same preparation. Results are ex-pressed as the means � S.E.M. (normal prostate, n � 8; implanted UGS,n � 8). ��, p � 0.01 versus normal prostate. C, KCa3.1 protein expressionsin normal prostate and implanted UGS. Plasma membrane fractionswere probed by immunoblotting with anti-KCa3.1 antibody (top panel).Bottom panel shows the summarized data of densitometric analyses (n �4). ��, p � 0.01 versus normal prostate. D, immunohistochemical stainingof KCa3.1 expression in stromal compartments of implanted UGS. Brown-stained nuclei were considered “positive.” Original magnifications, 40.
530 Ohya et al.
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from
tive signals were detected in the stromal compartments of im-planted UGS (Fig. 1D).
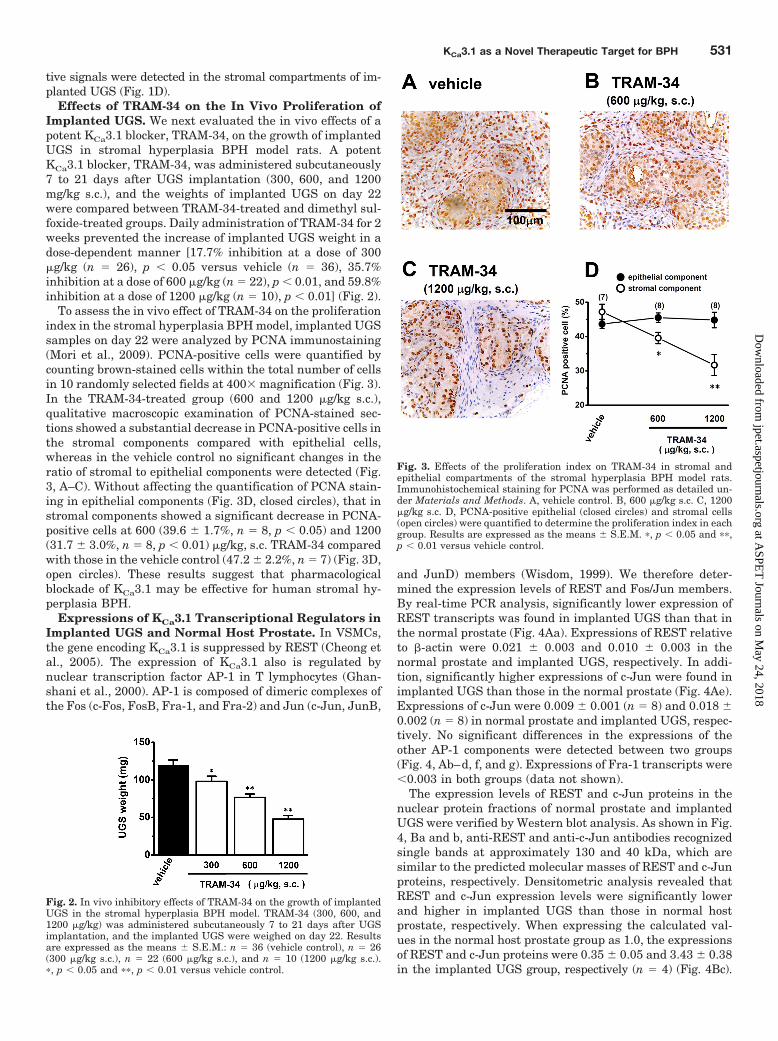
Effects of TRAM-34 on the In Vivo Proliferation ofImplanted UGS. We next evaluated the in vivo effects of apotent KCa3.1 blocker, TRAM-34, on the growth of implantedUGS in stromal hyperplasia BPH model rats. A potentKCa3.1 blocker, TRAM-34, was administered subcutaneously7 to 21 days after UGS implantation (300, 600, and 1200mg/kg s.c.), and the weights of implanted UGS on day 22were compared between TRAM-34-treated and dimethyl sul-foxide-treated groups. Daily administration of TRAM-34 for 2weeks prevented the increase of implanted UGS weight in adose-dependent manner [17.7% inhibition at a dose of 300�g/kg (n � 26), p � 0.05 versus vehicle (n � 36), 35.7%inhibition at a dose of 600 �g/kg (n � 22), p � 0.01, and 59.8%inhibition at a dose of 1200 �g/kg (n � 10), p � 0.01] (Fig. 2).
To assess the in vivo effect of TRAM-34 on the proliferationindex in the stromal hyperplasia BPH model, implanted UGSsamples on day 22 were analyzed by PCNA immunostaining(Mori et al., 2009). PCNA-positive cells were quantified bycounting brown-stained cells within the total number of cellsin 10 randomly selected fields at 400 magnification (Fig. 3).In the TRAM-34-treated group (600 and 1200 �g/kg s.c.),qualitative macroscopic examination of PCNA-stained sec-tions showed a substantial decrease in PCNA-positive cells inthe stromal components compared with epithelial cells,whereas in the vehicle control no significant changes in theratio of stromal to epithelial components were detected (Fig.3, A–C). Without affecting the quantification of PCNA stain-ing in epithelial components (Fig. 3D, closed circles), that instromal components showed a significant decrease in PCNA-positive cells at 600 (39.6 � 1.7%, n � 8, p � 0.05) and 1200(31.7 � 3.0%, n � 8, p � 0.01) �g/kg, s.c. TRAM-34 comparedwith those in the vehicle control (47.2 � 2.2%, n � 7) (Fig. 3D,open circles). These results suggest that pharmacologicalblockade of KCa3.1 may be effective for human stromal hy-perplasia BPH.
Expressions of KCa3.1 Transcriptional Regulators inImplanted UGS and Normal Host Prostate. In VSMCs,the gene encoding KCa3.1 is suppressed by REST (Cheong etal., 2005). The expression of KCa3.1 also is regulated bynuclear transcription factor AP-1 in T lymphocytes (Ghan-shani et al., 2000). AP-1 is composed of dimeric complexes ofthe Fos (c-Fos, FosB, Fra-1, and Fra-2) and Jun (c-Jun, JunB,
and JunD) members (Wisdom, 1999). We therefore deter-mined the expression levels of REST and Fos/Jun members.By real-time PCR analysis, significantly lower expression ofREST transcripts was found in implanted UGS than that inthe normal prostate (Fig. 4Aa). Expressions of REST relativeto �-actin were 0.021 � 0.003 and 0.010 � 0.003 in thenormal prostate and implanted UGS, respectively. In addi-tion, significantly higher expressions of c-Jun were found inimplanted UGS than those in the normal prostate (Fig. 4Ae).Expressions of c-Jun were 0.009 � 0.001 (n � 8) and 0.018 �0.002 (n � 8) in normal prostate and implanted UGS, respec-tively. No significant differences in the expressions of theother AP-1 components were detected between two groups(Fig. 4, Ab–d, f, and g). Expressions of Fra-1 transcripts were�0.003 in both groups (data not shown).
The expression levels of REST and c-Jun proteins in thenuclear protein fractions of normal prostate and implantedUGS were verified by Western blot analysis. As shown in Fig.4, Ba and b, anti-REST and anti-c-Jun antibodies recognizedsingle bands at approximately 130 and 40 kDa, which aresimilar to the predicted molecular masses of REST and c-Junproteins, respectively. Densitometric analysis revealed thatREST and c-Jun expression levels were significantly lowerand higher in implanted UGS than those in normal hostprostate, respectively. When expressing the calculated val-ues in the normal host prostate group as 1.0, the expressionsof REST and c-Jun proteins were 0.35 � 0.05 and 3.43 � 0.38in the implanted UGS group, respectively (n � 4) (Fig. 4Bc).
Fig. 2. In vivo inhibitory effects of TRAM-34 on the growth of implantedUGS in the stromal hyperplasia BPH model. TRAM-34 (300, 600, and1200 �g/kg) was administered subcutaneously 7 to 21 days after UGSimplantation, and the implanted UGS were weighed on day 22. Resultsare expressed as the means � S.E.M.: n � 36 (vehicle control), n � 26(300 �g/kg s.c.), n � 22 (600 �g/kg s.c.), and n � 10 (1200 �g/kg s.c.).�, p � 0.05 and ��, p � 0.01 versus vehicle control.
Fig. 3. Effects of the proliferation index on TRAM-34 in stromal andepithelial compartments of the stromal hyperplasia BPH model rats.Immunohistochemical staining for PCNA was performed as detailed un-der Materials and Methods. A, vehicle control. B, 600 �g/kg s.c. C, 1200�g/kg s.c. D, PCNA-positive epithelial (closed circles) and stromal cells(open circles) were quantified to determine the proliferation index in eachgroup. Results are expressed as the means � S.E.M. �, p � 0.05 and ��,p � 0.01 versus vehicle control.
KCa3.1 as a Novel Therapeutic Target for BPH 531
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from
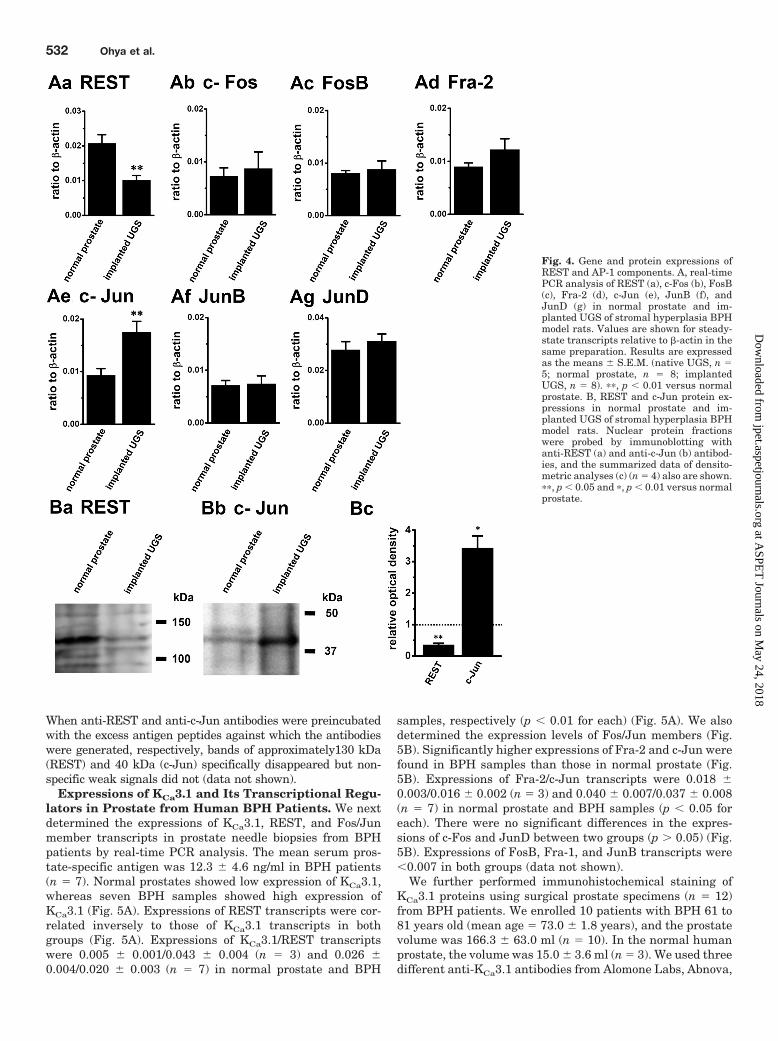
When anti-REST and anti-c-Jun antibodies were preincubatedwith the excess antigen peptides against which the antibodieswere generated, respectively, bands of approximately130 kDa(REST) and 40 kDa (c-Jun) specifically disappeared but non-specific weak signals did not (data not shown).
Expressions of KCa3.1 and Its Transcriptional Regu-lators in Prostate from Human BPH Patients. We nextdetermined the expressions of KCa3.1, REST, and Fos/Junmember transcripts in prostate needle biopsies from BPHpatients by real-time PCR analysis. The mean serum pros-tate-specific antigen was 12.3 � 4.6 ng/ml in BPH patients(n � 7). Normal prostates showed low expression of KCa3.1,whereas seven BPH samples showed high expression ofKCa3.1 (Fig. 5A). Expressions of REST transcripts were cor-related inversely to those of KCa3.1 transcripts in bothgroups (Fig. 5A). Expressions of KCa3.1/REST transcriptswere 0.005 � 0.001/0.043 � 0.004 (n � 3) and 0.026 �0.004/0.020 � 0.003 (n � 7) in normal prostate and BPH
samples, respectively (p � 0.01 for each) (Fig. 5A). We alsodetermined the expression levels of Fos/Jun members (Fig.5B). Significantly higher expressions of Fra-2 and c-Jun werefound in BPH samples than those in normal prostate (Fig.5B). Expressions of Fra-2/c-Jun transcripts were 0.018 �0.003/0.016 � 0.002 (n � 3) and 0.040 � 0.007/0.037 � 0.008(n � 7) in normal prostate and BPH samples (p � 0.05 foreach). There were no significant differences in the expres-sions of c-Fos and JunD between two groups (p � 0.05) (Fig.5B). Expressions of FosB, Fra-1, and JunB transcripts were�0.007 in both groups (data not shown).
We further performed immunohistochemical staining ofKCa3.1 proteins using surgical prostate specimens (n � 12)from BPH patients. We enrolled 10 patients with BPH 61 to81 years old (mean age � 73.0 � 1.8 years), and the prostatevolume was 166.3 � 63.0 ml (n � 10). In the normal humanprostate, the volume was 15.0 � 3.6 ml (n � 3). We used threedifferent anti-KCa3.1 antibodies from Alomone Labs, Abnova,
Fig. 4. Gene and protein expressions ofREST and AP-1 components. A, real-timePCR analysis of REST (a), c-Fos (b), FosB(c), Fra-2 (d), c-Jun (e), JunB (f), andJunD (g) in normal prostate and im-planted UGS of stromal hyperplasia BPHmodel rats. Values are shown for steady-state transcripts relative to �-actin in thesame preparation. Results are expressedas the means � S.E.M. (native UGS, n �5; normal prostate, n � 8; implantedUGS, n � 8). ��, p � 0.01 versus normalprostate. B, REST and c-Jun protein ex-pressions in normal prostate and im-planted UGS of stromal hyperplasia BPHmodel rats. Nuclear protein fractionswere probed by immunoblotting withanti-REST (a) and anti-c-Jun (b) antibod-ies, and the summarized data of densito-metric analyses (c) (n � 4) also are shown.��, p � 0.05 and �, p � 0.01 versus normalprostate.
532 Ohya et al.
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from
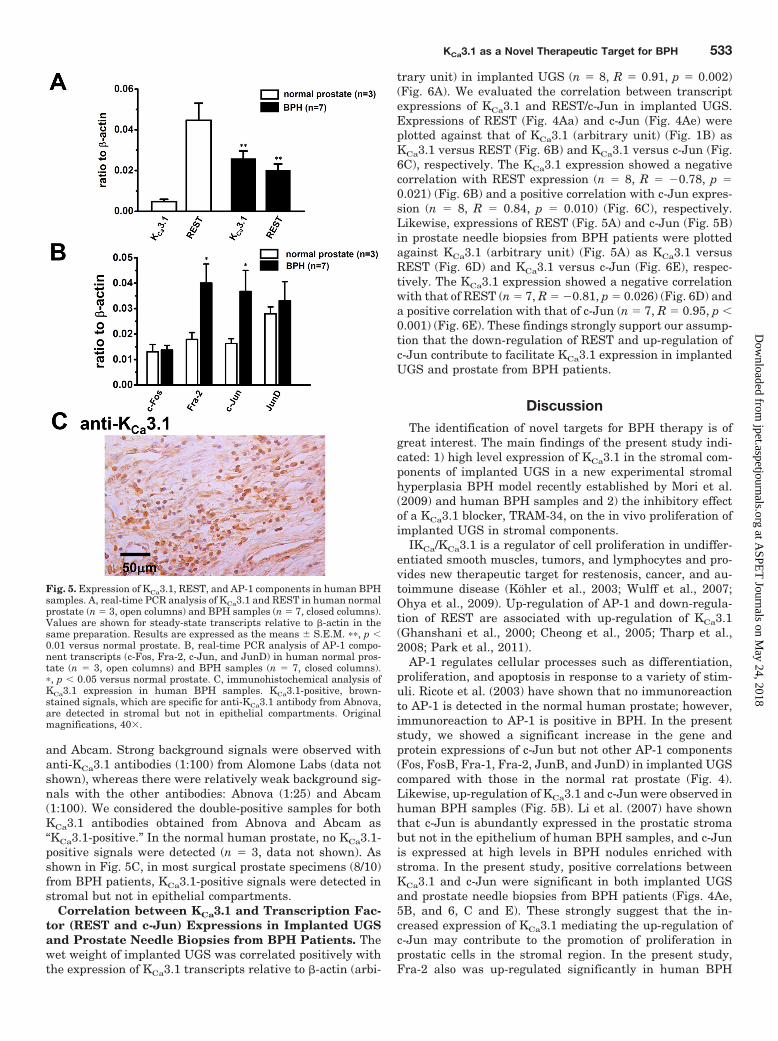
and Abcam. Strong background signals were observed withanti-KCa3.1 antibodies (1:100) from Alomone Labs (data notshown), whereas there were relatively weak background sig-nals with the other antibodies: Abnova (1:25) and Abcam(1:100). We considered the double-positive samples for bothKCa3.1 antibodies obtained from Abnova and Abcam as“KCa3.1-positive.” In the normal human prostate, no KCa3.1-positive signals were detected (n � 3, data not shown). Asshown in Fig. 5C, in most surgical prostate specimens (8/10)from BPH patients, KCa3.1-positive signals were detected instromal but not in epithelial compartments.
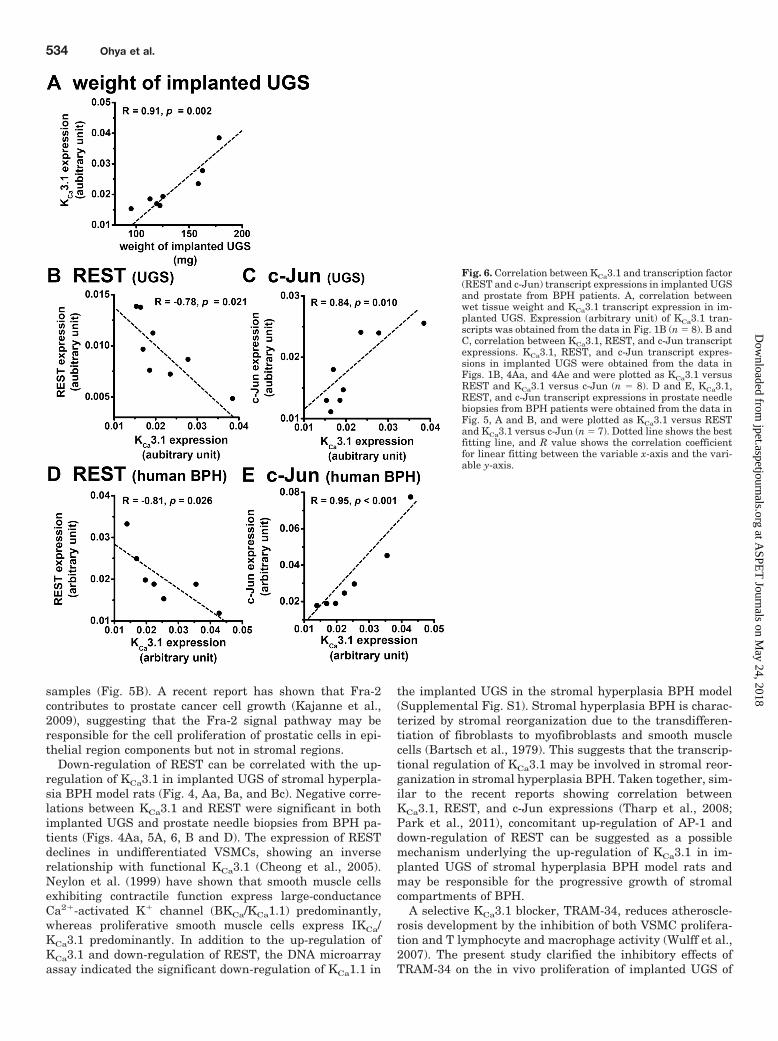
Correlation between KCa3.1 and Transcription Fac-tor (REST and c-Jun) Expressions in Implanted UGSand Prostate Needle Biopsies from BPH Patients. Thewet weight of implanted UGS was correlated positively withthe expression of KCa3.1 transcripts relative to �-actin (arbi-
trary unit) in implanted UGS (n � 8, R � 0.91, p � 0.002)(Fig. 6A). We evaluated the correlation between transcriptexpressions of KCa3.1 and REST/c-Jun in implanted UGS.Expressions of REST (Fig. 4Aa) and c-Jun (Fig. 4Ae) wereplotted against that of KCa3.1 (arbitrary unit) (Fig. 1B) asKCa3.1 versus REST (Fig. 6B) and KCa3.1 versus c-Jun (Fig.6C), respectively. The KCa3.1 expression showed a negativecorrelation with REST expression (n � 8, R � 0.78, p �0.021) (Fig. 6B) and a positive correlation with c-Jun expres-sion (n � 8, R � 0.84, p � 0.010) (Fig. 6C), respectively.Likewise, expressions of REST (Fig. 5A) and c-Jun (Fig. 5B)in prostate needle biopsies from BPH patients were plottedagainst KCa3.1 (arbitrary unit) (Fig. 5A) as KCa3.1 versusREST (Fig. 6D) and KCa3.1 versus c-Jun (Fig. 6E), respec-tively. The KCa3.1 expression showed a negative correlationwith that of REST (n � 7, R � 0.81, p � 0.026) (Fig. 6D) anda positive correlation with that of c-Jun (n � 7, R � 0.95, p �0.001) (Fig. 6E). These findings strongly support our assump-tion that the down-regulation of REST and up-regulation ofc-Jun contribute to facilitate KCa3.1 expression in implantedUGS and prostate from BPH patients.
DiscussionThe identification of novel targets for BPH therapy is of
great interest. The main findings of the present study indi-cated: 1) high level expression of KCa3.1 in the stromal com-ponents of implanted UGS in a new experimental stromalhyperplasia BPH model recently established by Mori et al.(2009) and human BPH samples and 2) the inhibitory effectof a KCa3.1 blocker, TRAM-34, on the in vivo proliferation ofimplanted UGS in stromal components.
IKCa/KCa3.1 is a regulator of cell proliferation in undiffer-entiated smooth muscles, tumors, and lymphocytes and pro-vides new therapeutic target for restenosis, cancer, and au-toimmune disease (Kohler et al., 2003; Wulff et al., 2007;Ohya et al., 2009). Up-regulation of AP-1 and down-regula-tion of REST are associated with up-regulation of KCa3.1(Ghanshani et al., 2000; Cheong et al., 2005; Tharp et al.,2008; Park et al., 2011).
AP-1 regulates cellular processes such as differentiation,proliferation, and apoptosis in response to a variety of stim-uli. Ricote et al. (2003) have shown that no immunoreactionto AP-1 is detected in the normal human prostate; however,immunoreaction to AP-1 is positive in BPH. In the presentstudy, we showed a significant increase in the gene andprotein expressions of c-Jun but not other AP-1 components(Fos, FosB, Fra-1, Fra-2, JunB, and JunD) in implanted UGScompared with those in the normal rat prostate (Fig. 4).Likewise, up-regulation of KCa3.1 and c-Jun were observed inhuman BPH samples (Fig. 5B). Li et al. (2007) have shownthat c-Jun is abundantly expressed in the prostatic stromabut not in the epithelium of human BPH samples, and c-Junis expressed at high levels in BPH nodules enriched withstroma. In the present study, positive correlations betweenKCa3.1 and c-Jun were significant in both implanted UGSand prostate needle biopsies from BPH patients (Figs. 4Ae,5B, and 6, C and E). These strongly suggest that the in-creased expression of KCa3.1 mediating the up-regulation ofc-Jun may contribute to the promotion of proliferation inprostatic cells in the stromal region. In the present study,Fra-2 also was up-regulated significantly in human BPH
Fig. 5. Expression of KCa3.1, REST, and AP-1 components in human BPHsamples. A, real-time PCR analysis of KCa3.1 and REST in human normalprostate (n � 3, open columns) and BPH samples (n � 7, closed columns).Values are shown for steady-state transcripts relative to �-actin in thesame preparation. Results are expressed as the means � S.E.M. ��, p �0.01 versus normal prostate. B, real-time PCR analysis of AP-1 compo-nent transcripts (c-Fos, Fra-2, c-Jun, and JunD) in human normal pros-tate (n � 3, open columns) and BPH samples (n � 7, closed columns).�, p � 0.05 versus normal prostate. C, immunohistochemical analysis ofKCa3.1 expression in human BPH samples. KCa3.1-positive, brown-stained signals, which are specific for anti-KCa3.1 antibody from Abnova,are detected in stromal but not in epithelial compartments. Originalmagnifications, 40.
KCa3.1 as a Novel Therapeutic Target for BPH 533
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from
samples (Fig. 5B). A recent report has shown that Fra-2contributes to prostate cancer cell growth (Kajanne et al.,2009), suggesting that the Fra-2 signal pathway may beresponsible for the cell proliferation of prostatic cells in epi-thelial region components but not in stromal regions.
Down-regulation of REST can be correlated with the up-regulation of KCa3.1 in implanted UGS of stromal hyperpla-sia BPH model rats (Fig. 4, Aa, Ba, and Bc). Negative corre-lations between KCa3.1 and REST were significant in bothimplanted UGS and prostate needle biopsies from BPH pa-tients (Figs. 4Aa, 5A, 6, B and D). The expression of RESTdeclines in undifferentiated VSMCs, showing an inverserelationship with functional KCa3.1 (Cheong et al., 2005).Neylon et al. (1999) have shown that smooth muscle cellsexhibiting contractile function express large-conductanceCa2�-activated K� channel (BKCa/KCa1.1) predominantly,whereas proliferative smooth muscle cells express IKCa/KCa3.1 predominantly. In addition to the up-regulation ofKCa3.1 and down-regulation of REST, the DNA microarrayassay indicated the significant down-regulation of KCa1.1 in
the implanted UGS in the stromal hyperplasia BPH model(Supplemental Fig. S1). Stromal hyperplasia BPH is charac-terized by stromal reorganization due to the transdifferen-tiation of fibroblasts to myofibroblasts and smooth musclecells (Bartsch et al., 1979). This suggests that the transcrip-tional regulation of KCa3.1 may be involved in stromal reor-ganization in stromal hyperplasia BPH. Taken together, sim-ilar to the recent reports showing correlation betweenKCa3.1, REST, and c-Jun expressions (Tharp et al., 2008;Park et al., 2011), concomitant up-regulation of AP-1 anddown-regulation of REST can be suggested as a possiblemechanism underlying the up-regulation of KCa3.1 in im-planted UGS of stromal hyperplasia BPH model rats andmay be responsible for the progressive growth of stromalcompartments of BPH.
A selective KCa3.1 blocker, TRAM-34, reduces atheroscle-rosis development by the inhibition of both VSMC prolifera-tion and T lymphocyte and macrophage activity (Wulff et al.,2007). The present study clarified the inhibitory effects ofTRAM-34 on the in vivo proliferation of implanted UGS of
Fig. 6. Correlation between KCa3.1 and transcription factor(REST and c-Jun) transcript expressions in implanted UGSand prostate from BPH patients. A, correlation betweenwet tissue weight and KCa3.1 transcript expression in im-planted UGS. Expression (arbitrary unit) of KCa3.1 tran-scripts was obtained from the data in Fig. 1B (n � 8). B andC, correlation between KCa3.1, REST, and c-Jun transcriptexpressions. KCa3.1, REST, and c-Jun transcript expres-sions in implanted UGS were obtained from the data inFigs. 1B, 4Aa, and 4Ae and were plotted as KCa3.1 versusREST and KCa3.1 versus c-Jun (n � 8). D and E, KCa3.1,REST, and c-Jun transcript expressions in prostate needlebiopsies from BPH patients were obtained from the data inFig. 5, A and B, and were plotted as KCa3.1 versus RESTand KCa3.1 versus c-Jun (n � 7). Dotted line shows the bestfitting line, and R value shows the correlation coefficientfor linear fitting between the variable x-axis and the vari-able y-axis.
534 Ohya et al.
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from

the stromal hyperplasia BPH model in a dose-dependentmanner (Fig. 2). Moreover, in the TRAM-34-treated group,qualitative macroscopic examination of PCNA-stained sec-tions showed a correlative decrease in PCNA-positive cells instromal compartments including smooth muscle cells with-out any changes in those in epithelial compartments (Fig. 3).Recent studies have shown that KCa such as KCa3.1 andsmall-conductance Ca2�-activated K� channel (SKCa2/KCa2.2) participate in the modulation of cell proliferation inseveral types of cells and that their pharmacological blockadeand down-regulation inhibit cell proliferation by G0/G1 arrest(Morimoto et al., 2007; Wang et al., 2007; Grgic et al., 2009).This suggests that the inhibitory effects of TRAM-34 on invivo cell proliferation in the implanted UGS of stromal BPHmodel rats may be responsible for the termination of cellcycle progression from G0/G1 by TRAM-34 treatment.
Although �1-adrenoceptor antagonists are considered anappropriate treatment for all BPH patients, irrespective ofprostate size, combination therapy with �1-adrenoceptor an-tagonists and 5�-reductase inhibitors is a recently acceptedtreatment for patients with clinically enlarged prostates (Mc-Connell et al., 2003). 5�-Reductase inhibitors are well toler-ated, but the most common adverse effects are sexual dys-function, including a reduced libido, erectile dysfunction,and, less frequently, ejaculation disorders (Gravas andOelke, 2010); therefore, other options are required to reduceprostate volume and the risk of clinical progression of BPH.Daily administration of TRAM-34 at relatively high concen-tration did not induce any changes in blood chemistry orhematology or at necropsy of the major organs of rodents (Siet al., 2006). Pharmacological blockade of KCa3.1 may berelatively safe and well tolerated as a therapeutic target ofBPH. In addition, pharmacological blockade of KCa3.1 pre-vents up-regulation of KCa3.1 and down-regulation of RESTin coronary arteries of the postangioplasty restenosis model(Tharp et al., 2008). This suggests that pharmacologicalblockade of KCa3.1 prevents phenotypic modulation of pros-tatic stromal cells in stromal hyperplasia BPH. Moreover,Tharp et al. (2006, 2008) have shown that in proliferative,dedifferentiated VSMCs cells up-regulation of KCa3.1 is fol-lowed by KCa3.1 promoter histone acetylation. This suggeststhat TRAM-34 also prevents KCa3.1 promoter activation atthe epigenetic level. Further study will provide more detailedmechanisms underlying TRAM-34-induced antiproliferativeeffects in prostatic stromal cells from BPH patients.
In VSMCs with a dedifferentiated, proliferating pheno-type, hyperpolarization by K� channel activation increasesthe driving force for Ca2� entry via voltage-independentCa2� channels such as store-operated Ca2� channel, recep-tor-operated Ca2� channel and Ca2�-release activating Ca2�
channel, which are encoded by mammalian homologues oftransient receptor potential genes and Orai/stromal interac-tion module genes (Inoue et al., 2006; Albert et al., 2007;Guibert et al., 2008; House et al., 2008). However, molecularcomponents directly involving VSMC proliferation remain tobe determined. Dedifferentiated VSMCs are characterized byloss of voltage-gated Ca2� channel, which is a major Ca2�
channel component in smooth muscle cells with a differenti-ated, contractile phenotype. In implanted UGS of stromalhyperplasia BPH model rats, voltage-gated Ca2� channelwas rarely expressed (data not shown). In the present study,the molecular identification and functional characterization
of voltage-independent Ca2� channels remain to be deter-mined in either implanted UGS or prostatic tissues fromBPH patients. Several researchers have reported that down-regulation of KCa1.1 and voltage-gated K� channel is ob-served in lower urinary tracts in BPH and prostatitis pa-tients (Liang et al., 2006; Chang et al., 2010). DNAmicroarray analysis showed that KCa1.1 transcript expres-sion (probe ID 1372929_at) in implanted UGS was markedlylower than that in the normal host prostate. No significantdifference in voltage-gated K� channel transcript expression(probe ID 1369802_at) was detected between them (Supple-mental Fig. S1); however, the relative expression level ofvoltage-gated K� channel was much lower than that ofKCa3.1in implanted UGS and prostate from BPH patients(data not shown). The possibilities of voltage-independentCa2� channels and other K� channel subtypes as therapeutictargets for BPH are also pressing questions.
In summary, our present study suggests that the activa-tion of Ca2�-release activating Ca2� channels by up-regula-tion of KCa3.1 may be implicated in the development of BPH,which represents a field of great interest for scientific andclinical research communities. We further showed that bothc-Jun and REST are candidates for the transcriptional reg-ulation of KCa3.1 in stromal compartments of BPH. In vivotreatment with TRAM-34 significantly suppressed cell pro-liferation in stromal compartments, supporting the notionthat KCa3.1 blockers may be a novel treatment option forpatients suffering from stromal hyperplasia BPH.
Acknowledgments
We thank to Dr. Oda (Taiho Pharmaceutical Co., Ltd., Japan) forgiving us valuable advice and suggestions.
Authorship Contributions
Participated in research design: Ohya, Sasaki, Kohri, andImaizumi.
Conducted experiments: Ohya, Niwa, Kojima, and Sakuragi.Performed data analysis: Ohya, Niwa, Kojima, and Sakuragi.Wrote or contributed to the writing of the manuscript: Ohya, Ko-
jima, Sasaki, Kohri, and Imaizumi.
ReferencesAlbert AP, Saleh SN, Peppiatt-Wildman CM, and Large WA (2007) Multiple activa-
tion mechanisms of store-operated TRPC channels in smooth muscle cells.J Physiol 583:25–36.
Andersson KE and Gratzke C (2007) Pharmacology of alpha1-adrenoceptor antago-nists in the lower urinary tract and central nervous system. Nat Clin Pract Urol4:368–378.
Bartsch G, Muller HR, Oberholzer M, and Rohr HP (1979) Light microscopic stereo-logical analysis of the normal human prostate and of benign prostatic hyperplasia.J Urol 122:487–491.
Berry SJ, Coffey DS, Walsh PC, and Ewing LL (1984) The development of humanbenign prostatic hyperplasia with age. J Urol 132:474–479.
Chang S, Gomes CM, Hypolite JA, Marx J, Alanzi J, Zderic SA, Malkowicz B, WeinAJ, and Chacko S (2010) Detrusor overactivity is associated with downregulationof large-conductance calcium- and voltage-activated potassium channel protein.Am J Physiol Renal Physiol 298:F1416–F1423.
Cheong A, Bingham AJ, Li J, Kumar B, Sukumar P, Munsch C, Buckley NJ, NeylonCB, Porter KE, Beech DJ, et al.(2005) Downregulated REST transcription factor isa switch enabling critical potassium channel expression and cell proliferation. MolCell 20:45–52.
Djavan B (2003) Lower urinary tract symptoms/benign prostatic hyperplasia: fastcontrol of the patient’s quality of life. Urology 62:6–14.
Ghanshani S, Wulff H, Miller MJ, Rohm H, Neben A, Gutman GA, Cahalan MD, andChandy KG (2000) Up-regulation of the IKCa1 potassium channel during T-cellactivation. Molecular mechanism and functional consequences. J Biol Chem 275:37137–37149.
Gravas S and Oelke M (2010) Current status of 5alpha-reductase inhibitors in themanagement of lower urinary tract symptoms and BPH. World J Urol 28:9–15.
Grgic I, Kiss E, Kaistha BP, Busch C, Kloss M, Sautter J, Muller A, Kaistha A,Schmidt C, Raman G, et al. (2009) Renal fibrosis is attenuated by targeted
KCa3.1 as a Novel Therapeutic Target for BPH 535
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from

disruption of KCa3.1 potassium channels. Proc Natl Acad Sci USA 106:14518–14523.
Guibert C, Ducret T, and Savineau JP (2008) Voltage-independent calcium influx insmooth muscle. Prog Biophys Mol Biol 98:10–23.
House SJ, Potier M, Bisaillon J, Singer HA, and Trebak M (2008) The non-excitablesmooth muscle: calcium signaling and phenotypic switching during vascular dis-ease. Pflugers Arch 456:769–785.
Inoue R, Jensen LJ, Shi J, Morita H, Nishida M, Honda A, and Ito Y (2006) Transientreceptor potential channels in cardiovascular function and disease. Circ Res 99:119–131.
Ishigooka M, Hayami S, Hashimoto T, Suzuki Y, Katoh T, and Nakada T (1996)Relative and total volume of histological components in benign prostatic hyper-plasia: relationships between histological components and clinical findings. Pros-tate 29:77–82.
Ishii TM, Silvia C, Hirschberg B, Bond CT, Adelman JP, and Maylie J (1997) Ahuman intermediate conductance calcium-activated potassium channel. Proc NatlAcad Sci USA 94:11651–11656.
Kajanne R, Miettinen P, Tenhunen M, and Leppa S (2009) Transcription factor AP-1promotes growth and radioresistance in prostate cancer cells. Int J Oncol 35:1175–1182.
Kojima Y, Sasaki S, Hayashi Y, Tsujimoto G, and Kohri K (2009a) Subtypes ofalpha1-adrenoceptors in BPH: future prospects for personalized medicine. NatClin Pract Urol 6:44–53.
Kojima Y, Sasaki S, Oda N, Koshimizu TA, Hayashi Y, Kiniwa M, Tsujimoto G, andKohri K (2009b) Prostate growth inhibition by subtype-selective alpha(1)-adrenoceptor antagonist naftopidil in benign prostatic hyperplasia. Prostate 69:1521–1528.
Kohler R, Wulff H, Eichler I, Kneifel M, Neumann D, Knorr A, Grgic I, Kampfe D, SiH, Wibawa J, et al. (2003) Blockade of the intermediate-conductance calcium-activated potassium channel as a new therapeutic strategy for restenosis. Circu-lation 108:1119–1125.
Li W, Wu CL, Febbo PG, and Olumi AF (2007) Stromally expressed c-Jun regulatesproliferation of prostate epithelial cells. Am J Pathol 171:1189–1198.
Liang CZ, Guo QK, Hao ZY, Yang S, Wang DB, Wu LX, Liu C, Wang KX, and ZhangXJ (2006) K channel expression in prostate epithelium and its implications in menwith chronic prostatitis. BJU Int 97:190–192.
McConnell JD, Roehrborn CG, Bautista OM, Andriole GL Jr, Dixon CM, Kusek JW,Lepor H, McVary KT, Nyberg LM Jr, Clarke HS, et al. (2003) The long-term effectof doxazosin, finasteride, and combination therapy on the clinical progression ofbenign prostatic hyperplasia. N Engl J Med 349:2387–2398.
Mori F, Oda N, Sakuragi M, Sakakibara F, Kiniwa M, and Miyoshi K (2009) Newhistopathological experimental model for benign prostatic hyperplasia: stromalhyperplasia in rats. J Urol 181:890–898.
Morimoto T, Ohya S, Hayashi H, Onozaki K, and Imaizumi Y (2007) Cell-cycle-dependent regulation of Ca2�-activated K� channel in Jurkat T-lymphocyte.J Pharmacol Sci 104:94–98.
Neylon CB, Lang RJ, Fu Y, Bobik A, and Reinhart PH (1999) Molecular cloning andcharacterization of the intermediate-conductance Ca(2�)-activated K(�) channelin vascular smooth muscle: relationship between K(Ca) channel diversity andsmooth muscle cell function. Circ Res 85:e33–e43.
Ohya S, Kimura K, Niwa S, Ohno A, Kojima Y, Sasaki S, Kohri K, and Imaizumi Y
(2009) Malignancy grade-dependent expression of K�-channel subtypes in humanprostate cancer. J Pharmacol Sci 109:148–151.
Ohya S, Kimura S, Kitsukawa M, Muraki K, Watanabe M, and Imaizumi Y (2000)SK4 encodes intermediate conductance Ca2�-activated K� channels in mouseurinary bladder smooth muscle cells. Jpn J Pharmacol 84:97–100.
Ohya S, Kuwata Y, Sakamoto K, Muraki K, and Imaizumi Y (2005) Cardioprotectiveeffects of estradiol include the activation of large-conductance Ca(2�)-activatedK(�) channels in cardiac mitochondria. Am J Physiol Heart Circ Physiol 289:H1635–H1642.
Park S, Kim JA, Joo KY, Choi S, Choi EN, Shin JA, Han KH, Jung SC, and Suh SH(2011) Globotriaosylceramide leads to K(Ca)3.1 channel dysfunction: a new insightinto endothelial dysfunction in Fabry disease. Cardiovasc Res 89:290–299.
Pollard M, Luckert PH, and Snyder D (1989) Prevention and treatment of experi-mental prostate cancer in Lobund-Wistar rats. I. Effects of estradiol, dihydrotes-tosterone, and castration. Prostate 15:95–103.
Ricote M, Royuela M, García-Tunon I, Bethencourt FR, Paniagua R, and Fraile B(2003) Pro-apoptotic tumor necrosis factor-alpha transduction pathway in normalprostate, benign prostatic hyperplasia and prostatic carcinoma. J Urol 170:787–790.
Shapiro E, Becich MJ, Hartanto V, and Lepor H (1992) The relative proportion ofstromal and epithelial hyperplasia is related to the development of symptomaticbenign prostate hyperplasia. J Urol 147:1293–1297.
Si H, Heyken WT, Wolfle SE, Tysiac M, Schubert R, Grgic I, Vilianovich L, GiebingG, Maier T, Gross V, et al. (2006) Impaired endothelium-derived hyperpolarizingfactor-mediated dilations and increased blood pressure in mice deficient of theintermediate-conductance Ca2�-activated K� channel. Circ Res 99:537–544.
Stocker M (2004) Ca(2�)-activated K� channels: molecular determinants and func-tion of the SK family. Nat Rev Neurosci 5:758–770.
Tharp DL, Wamhoff BR, Turk JR, and Bowles DK (2006) Upregulation of interme-diate-conductance Ca2�-activated K� channel (IKCa1) mediates phenotypic mod-ulation of coronary smooth muscle. Am J Physiol Heart Circ Physiol 291:H2493–H2503.
Tharp DL, Wamhoff BR, Wulff H, Raman G, Cheong A, and Bowles DK (2008) Localdelivery of the KCa3.1 blocker, TRAM-34, prevents acute angioplasty-inducedcoronary smooth muscle phenotypic modulation and limits stenosis. ArteriosclerThromb Vasc Biol 28:1084–1089.
Wang ZH, Shen B, Yao HL, Jia YC, Ren J, Feng YJ, and Wang YZ (2007) Blockageof intermediate-conductance Ca(2�)-activated K(�) channels inhibits progressionof human endometrial cancer. Oncogene 26:5107–5114.
Wisdom R (1999) AP-1: one switch for many signals. Exp Cell Res 253:180–185.Wulff H, Kolski-Andreaco A, Sankaranarayanan A, Sabatier JM, and Shakkottai V
(2007) Modulators of small- and intermediate-conductance calcium-activated po-tassium channels and their therapeutic indications. Curr Med Chem 14:1437–1457.
Address correspondence to: Yuji Imaizumi, Department of Molecular andCellular Pharmacology, Graduate School of Pharmaceutical Sciences, NagoyaCity University, 3-1 Tanabe-dori, Muzuhoku, Nagoya 467-8603, Japan.E-mail: [email protected]
536 Ohya et al.
at ASPE
T Journals on M
ay 24, 2018jpet.aspetjournals.org
Dow
nloaded from


![New INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/187/3/local/back... · 2005. 12. 3. · hycanthone effects onspermatogonial cells, de-oxyribonucleic acidsynthesis](https://img.pdfslide.us/doc/110x75/6067c6518625ed3f66076f25/new-index-jpet-jpet-2005-12-3-hycanthone-effects-onspermatogonial-cells.jpg)















![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/234/3/local/back...effect, 708 Blockade, reticuloendothelial, enzyme-al-bumin conjugates, chronic adininis-tration](https://img.pdfslide.us/doc/110x75/60757ab7f966210d5e51d2f2/index-jpet-jpet-effect-708-blockade-reticuloendothelial-enzyme-al-bumin.jpg)






![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/178/3/local/back... · 2005. 11. 25. · INDEX 651. chemoreceptor, demonstration by inhibition ofcarbonic anhydrase,](https://img.pdfslide.us/doc/110x75/60791f369cbd2b1cd042ecd2/index-jpet-jpet-2005-11-25-index-651-chemoreceptor-demonstration-by-inhibition.jpg)

![INDEX [jpet.aspetjournals.org]jpet.aspetjournals.org/content/jpet/230/3/local/back-matter.pdf · histrionicotoxin effects (frogs), 619 ... distribution kinetics ana- ... and myocardium](https://img.pdfslide.us/doc/110x75/5b7ac0067f8b9ae1328d73ab/index-jpet-jpet-histrionicotoxin-effects-frogs-619-distribution-kinetics.jpg)

