Embed Size (px)
Citation preview

Integrated Metabolite and Transcript Profiling Identifya Biosynthetic Mechanism for Hispidol in Medicagotruncatula Cell Cultures1[C][W][OA]
Mohamed A. Farag, Bettina E. Deavours, Angelo de Fatima, Marina Naoumkina,Richard A. Dixon, and Lloyd W. Sumner*
Plant Biology Division, Samuel Roberts Noble Foundation, Ardmore, Oklahoma 73401 (M.A.F., B.E.D., M.N.,R.A.D., L.W.S.); Pharmacognosy Department, Faculty of Pharmacy, Cairo University, Cairo, Egypt 11562(M.A.F.); Department of Biology, Colorado State University, Fort Collins, Colorado 80523 (B.E.D.); andDepartamento de Quımica, Instituto de Ciencias Exatas, Universidade Federal de Minas Gerais, CampusPampulha, Belo Horizonte, Minas Gerais, 31270–901, Brazil (A.d.F.)
Metabolic profiling of elicited barrel medic (Medicago truncatula) cell cultures using high-performance liquid chromatographycoupled to photodiode and mass spectrometry detection revealed the accumulation of the aurone hispidol (6-hydroxy-2-[(4-hydroxyphenyl)methylidene]-1-benzofuran-3-one) as a major response to yeast elicitor. Parallel, large-scale transcriptomeprofiling indicated that three peroxidases, MtPRX1, MtPRX2, and MtPRX3, were coordinately induced with the accumulationof hispidol. MtPRX1 and MtPRX2 exhibited aurone synthase activity based upon in vitro substrate specificity and productprofiles of recombinant proteins expressed in Escherichia coli. Hispidol possessed significant antifungal activity relative to otherM. truncatula phenylpropanoids tested but has not been reported in this species before and was not found in differentiatedroots in which high levels of the peroxidase transcripts accumulated. We propose that hispidol is formed in cell cultures bymetabolic spillover when the pool of its precursor, isoliquiritigenin, builds up as a result of an imbalance between the upstreamand downstream segments of the phenylpropanoid pathway, reflecting the plasticity of plant secondary metabolism. Theresults illustrate that integration of metabolomics and transcriptomics in genetically reprogrammed plant cell cultures is apowerful approach for the discovery of novel bioactive secondary metabolites and the mechanisms underlying theirgeneration.
Phenylpropanoid metabolism encompasses a com-plex network of branching biochemical pathways thatcollectively provide plants with thousands of com-pounds that have diverse functions in plants, mostnotably in defense, such as cell wall strengthening andrepair (e.g. lignin and suberin), antimicrobial activity(e.g. furanocoumarin, pterocarpan, and isoflavonoidphytoalexins), and as signaling compounds such asluteolin (Peters et al., 1986) and apigenin (Peters andLong, 1988). In the core pathway, Phe is convertedinto p-coumaroyl-CoA, and the condensation ofthis molecule with three molecules of malonyl-CoA, via chalcone synthase activity, yields 4,2#,4#,6#-
tetrahydroxychalcone (naringenin chalcone), which isthe primary C15 flavanone skeleton fromwhich a largenumber of flavonoid subclasses diverge (Fig. 1). Inlegumes, the 6#-deoxy chalcone isoliquiritigenin isproduced through the concerted activity of chalconesynthase and chalcone reductase (Ralston et al., 2005).This trihydroxychalcone can be further metabolizedby chalcone isomerase to form liquiritigenin, a pre-cursor for a range of 5-deoxy flavones, flavonols, andisoflavones (Fig. 1). Exceptions to this biosyntheticroute are the aurones, a flavonoid subclass that can bedirectly synthesized from isoliquiritigenin (Strack,1997). Aurones (Fig. 1) serve significant roles in flowerpigmentation (Nakayama et al., 2001) and defenseresponses (Pare et al., 1991). In humans, aurones haverecently drawn much attention for their therapeuticpotential as anticancer, antidiabetic, antibacterial, an-tiparasitic, and antihormonal agents (Boumendjel,2003). While flavone, flavonol, and isoflavone biosyn-thetic pathways have been extensively studied usingdifferent molecular and biochemical approaches, onlya few studies on aurone biosynthesis have beenreported.
Aureusidin synthase, a polyphenol oxidase (PPO)homolog, has been identified in snapdragon (Antirrhi-num majus) as a key enzyme involved in aurone
1 This work was supported by the National Science FoundationPlant Genome Research Program (grant no. DBI–0109732).
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Lloyd W. Sumner ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
[W] The online version of this article contains Web-only data.[OA] Open Access articles can be viewed online without a sub-
scription.www.plantphysiol.org/cgi/doi/10.1104/pp.109.141481
1096 Plant Physiology�, November 2009, Vol. 151, pp. 1096–1113, www.plantphysiol.org � 2009 American Society of Plant Biologists
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
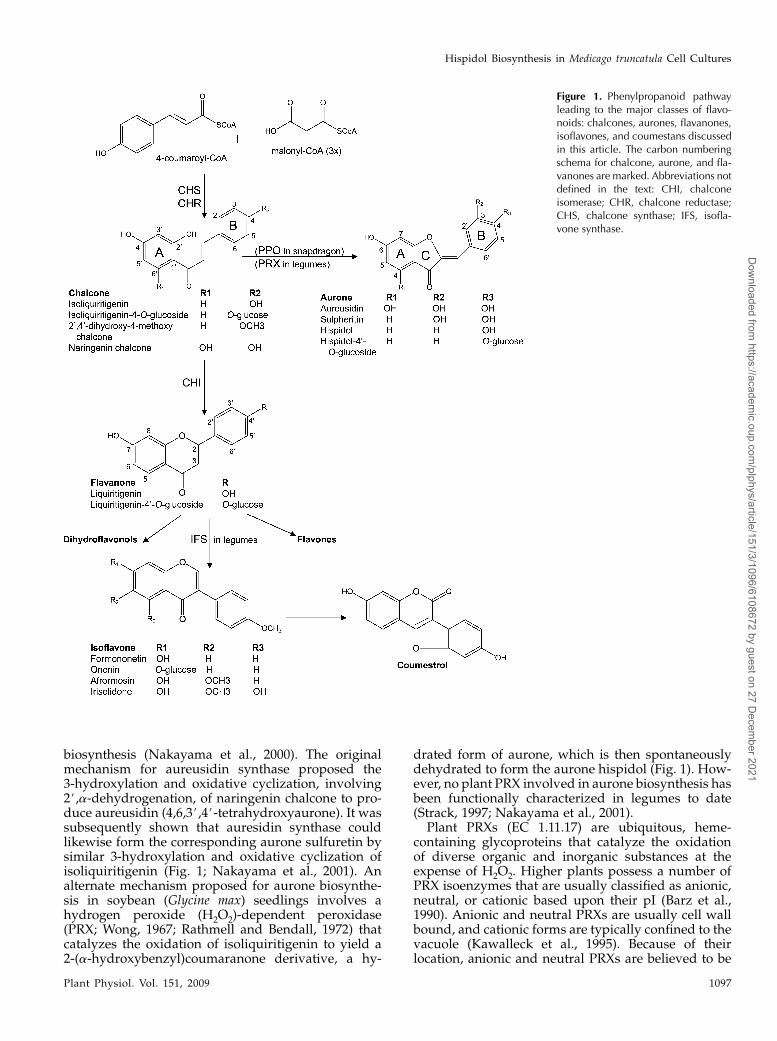
biosynthesis (Nakayama et al., 2000). The originalmechanism for aureusidin synthase proposed the3-hydroxylation and oxidative cyclization, involving2#,a-dehydrogenation, of naringenin chalcone to pro-duce aureusidin (4,6,3#,4#-tetrahydroxyaurone). It wassubsequently shown that auresidin synthase couldlikewise form the corresponding aurone sulfuretin bysimilar 3-hydroxylation and oxidative cyclization ofisoliquiritigenin (Fig. 1; Nakayama et al., 2001). Analternate mechanism proposed for aurone biosynthe-sis in soybean (Glycine max) seedlings involves ahydrogen peroxide (H2O2)-dependent peroxidase(PRX; Wong, 1967; Rathmell and Bendall, 1972) thatcatalyzes the oxidation of isoliquiritigenin to yield a2-(a-hydroxybenzyl)coumaranone derivative, a hy-
drated form of aurone, which is then spontaneouslydehydrated to form the aurone hispidol (Fig. 1). How-ever, no plant PRX involved in aurone biosynthesis hasbeen functionally characterized in legumes to date(Strack, 1997; Nakayama et al., 2001).
Plant PRXs (EC 1.11.17) are ubiquitous, heme-containing glycoproteins that catalyze the oxidationof diverse organic and inorganic substances at theexpense of H2O2. Higher plants possess a number ofPRX isoenzymes that are usually classified as anionic,neutral, or cationic based upon their pI (Barz et al.,1990). Anionic and neutral PRXs are usually cell wallbound, and cationic forms are typically confined to thevacuole (Kawalleck et al., 1995). Because of theirlocation, anionic and neutral PRXs are believed to be
Figure 1. Phenylpropanoid pathwayleading to the major classes of flavo-noids: chalcones, aurones, flavanones,isoflavones, and coumestans discussedin this article. The carbon numberingschema for chalcone, aurone, and fla-vanones are marked. Abbreviations notdefined in the text: CHI, chalconeisomerase; CHR, chalcone reductase;CHS, chalcone synthase; IFS, isofla-vone synthase.
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1097
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

mainly involved in plant defense. PRXs can oxidizevacuolar phenolic pools and also play key roles in thepolymerization steps involved in lignification andsuberization of plant cell walls (Chittoor et al., 1997).Functional characterizations of PRXs have been basedprimarily upon genomic sequence mining and in vitrobiochemical assays. However, the extremely widesubstrate specificities of PRXs, the high number ofPRX genes, and the diversity of PRX structures raiseimportant questions about their true in vivo substratesand functions.
Large-scale metabolite profiling (metabolomics) is apowerful tool for analyzing metabolism and genefunction (Sumner et al., 2003; Bino et al., 2004). Manyearly metabolomics studies were focused upon pri-mary metabolites for plant genotyping (Taylor et al.,2003), detection of silent phenotypes in transgenicpotato (Solanum tuberosum; Weckwerth et al., 2004),and the examination of stress responses (Broecklinget al., 2005). Parallel profiling of transcript and metab-olites has also been applied to study the effect of coldacclimation (Kaplan et al., 2004), phosphorus stress(Hernandez et al., 2007), and arbuscular mycorrhizalinteractions (Schaarschmidt et al., 2007) in plants.More recently, the profiling of natural products hasalso been incorporated into themetabolomics approachand integrated with transcriptome analysis to identifynovel gene functions associated with flavonoid bio-synthesis (Tohge et al., 2005; Naoumkina et al., 2007;Farag et al., 2008; Yonekura-Sakakibara et al., 2008).Such an integrated approach represents a powerfulplatform for the clarification of gene function in plantsecondary metabolism (Fridman and Pichersky, 2005).
Barrel medic (Medicago truncatula) is a rapidly de-veloping model for legume biology (Cook, 1999;Young et al., 2005) and an excellent species for study-ing the rich and unique secondary metabolism oflegumes (Dixon and Sumner, 2003). We have recentlyutilized elicited M. truncatula liquid suspension cellcultures to study biotic stress responses using an in-tegrated functional genomics approach that includedtranscriptomics (Suzuki et al., 2005), proteomics (Leiet al., 2005), and metabolomics (Broeckling et al., 2005;Farag et al., 2008). These studies involved independentapplications of the phytohormone methyl jasmonate(MeJA) and a yeast cell wall preparation (yeast elicitor[YE], a fungal pathogen mimic) to M. truncatula rootsuspension cell cultures. We report here that the anti-microbial aurone, hispidol, is a major YE-inducedsecondary metabolite in these cultures, and correla-tion analyses between metabolite and transcript pro-files implicated specific PRXs in aurone biosynthesis.The functionalities of these PRXs were subsequentlyconfirmed by biochemical analysis of recombinantproteins. The integration of metabolomics and tran-scriptomics data has thus led to the discovery of both anovel bioactive secondary metabolite and a novelmechanism for its biosynthesis. These results are dis-cussed in terms of biochemical and cellular responsesto biotic stress.
RESULTS
Hispidol and Hispidol-4#-O-b-D-Glucoside Are NovelPhenylpropanoid Compounds Induced in the Response
of M. truncatula to YE
A large-scale elicitation experiment was conductedusing liquid suspension cell cultures and two elicitors(YE and MeJA) to generate an integrated global dataset (transcriptome, proteome, and metabolome) and tofacilitate gene discovery and novel insight into bioticand abiotic stress responses associated with naturalproduct pathways in M. truncatula. Triplicate biolog-ical samples from control and elicited cell cultureswere harvested at 21 different time points between 0and 48 h postelicitation for each elicitor and eachreplicate from independent culture flasks (Broecklinget al., 2005). Global metabolite, protein, and transcriptprofiles were obtained for the sampled cells andspecifically queried as related to the nature and extentof the effect on phenylpropanoid biosynthesis. Theexpression levels of approximately 16,000 tentativeconsensus sequences (TCs) were monitored using acustom M. truncatula cDNA oligonucleotide micro-array. The accumulation patterns of approximately1,000 proteins were monitored with two-dimensionalgel electrophoresis (Lei et al., 2005). Approximately180 secondary metabolites were profiled using HPLCcoupled to photodiode and mass spectrometry detec-tion (HPLC-PDA-MS; Farag et al., 2007, 2008), andapproximately 500 primary metabolites were profiledby gas chromatography-mass spectrometry (GC-MS;Broeckling et al., 2005).
The effect on phenylpropanoid metabolite poolswas most dramatic in cell cultures treated with YE.M. truncatula and alfalfa (Medicago sativa) cell culturesare known to respond to YE by transcriptional in-duction of isoflavonoid biosynthetic genes and thedownstream accumulation of the isoflavonoid-derivedpterocarpan phytoalexin medicarpin (Suzuki et al.,2005). Increased levels of several isoflavonoids wereobserved after exposure to YE in our cultures, withmedicarpin notably induced 10-fold. However, twounknown compounds, P1 and P2, showed the largestfold inductions, up 45- and 15-fold, respectively, inresponse to YE, and their corresponding peaks werenot found in the HPLC-PDA-MS chromatograms ofcontrol cells (Fig. 2A). Neither P1 nor P2 was inducedin response to MeJA. In the negative ion electrosprayionization mode, the parent ion observed for P1 (re-tention time = 23.5 min) had mass-to-charge ratio (m/z)415, with a major fragment ion at m/z 253. The m/z 162difference between both ions suggests the loss of ahexose (Fig. 2B). P2 (retention time = 33.5 min) alsohad an ion peak atm/z 253, suggesting that P2 might bethe aglycone moiety observed as part of the hexoseconjugate in P1 (Fig. 2C). Enzymatic hydrolysis of P1with b-glucosidase followed by HPLC-PDA-MS anal-ysis yielded a single peak with the same retention timeand spectral characteristics as P2. GC-MS analysis of
Farag et al.
1098 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
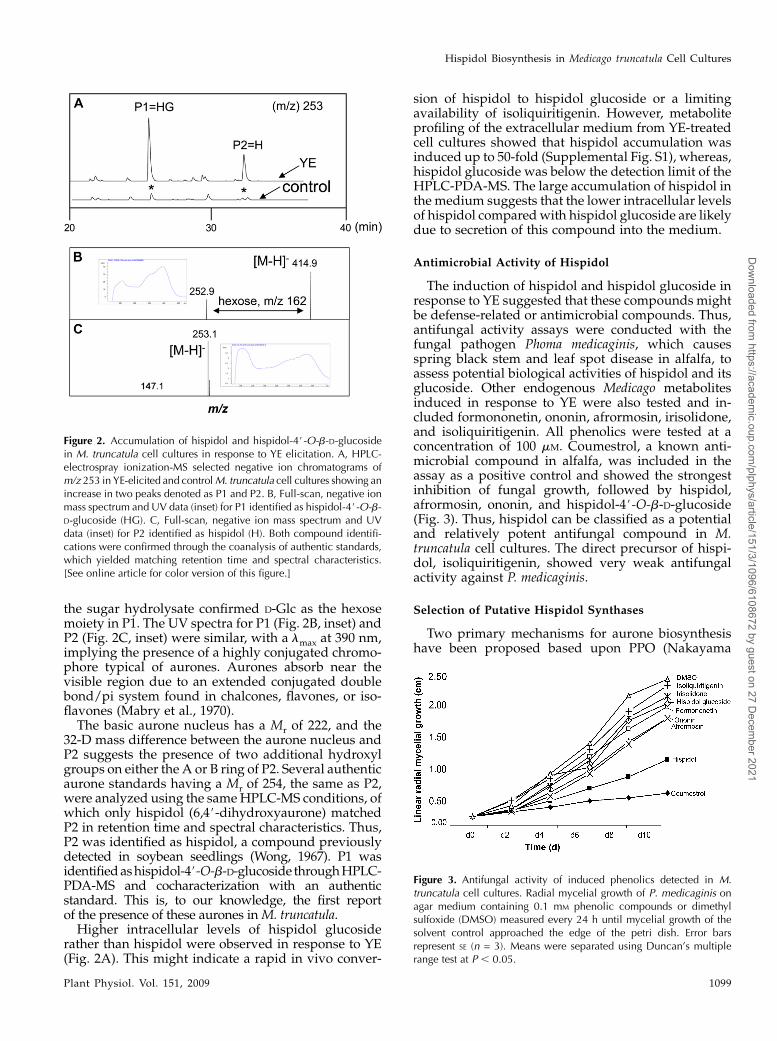
the sugar hydrolysate confirmed D-Glc as the hexosemoiety in P1. The UV spectra for P1 (Fig. 2B, inset) andP2 (Fig. 2C, inset) were similar, with a lmax at 390 nm,implying the presence of a highly conjugated chromo-phore typical of aurones. Aurones absorb near thevisible region due to an extended conjugated doublebond/pi system found in chalcones, flavones, or iso-flavones (Mabry et al., 1970).The basic aurone nucleus has a Mr of 222, and the
32-D mass difference between the aurone nucleus andP2 suggests the presence of two additional hydroxylgroups on either the A or B ring of P2. Several authenticaurone standards having a Mr of 254, the same as P2,were analyzed using the sameHPLC-MS conditions, ofwhich only hispidol (6,4#-dihydroxyaurone) matchedP2 in retention time and spectral characteristics. Thus,P2 was identified as hispidol, a compound previouslydetected in soybean seedlings (Wong, 1967). P1 wasidentifiedashispidol-4#-O-b-D-glucoside throughHPLC-PDA-MS and cocharacterization with an authenticstandard. This is, to our knowledge, the first reportof the presence of these aurones inM. truncatula.Higher intracellular levels of hispidol glucoside
rather than hispidol were observed in response to YE(Fig. 2A). This might indicate a rapid in vivo conver-
sion of hispidol to hispidol glucoside or a limitingavailability of isoliquiritigenin. However, metaboliteprofiling of the extracellular medium from YE-treatedcell cultures showed that hispidol accumulation wasinduced up to 50-fold (Supplemental Fig. S1), whereas,hispidol glucoside was below the detection limit of theHPLC-PDA-MS. The large accumulation of hispidol inthe medium suggests that the lower intracellular levelsof hispidol comparedwith hispidol glucoside are likelydue to secretion of this compound into the medium.
Antimicrobial Activity of Hispidol
The induction of hispidol and hispidol glucoside inresponse to YE suggested that these compounds mightbe defense-related or antimicrobial compounds. Thus,antifungal activity assays were conducted with thefungal pathogen Phoma medicaginis, which causesspring black stem and leaf spot disease in alfalfa, toassess potential biological activities of hispidol and itsglucoside. Other endogenous Medicago metabolitesinduced in response to YE were also tested and in-cluded formononetin, ononin, afrormosin, irisolidone,and isoliquiritigenin. All phenolics were tested at aconcentration of 100 mM. Coumestrol, a known anti-microbial compound in alfalfa, was included in theassay as a positive control and showed the strongestinhibition of fungal growth, followed by hispidol,afrormosin, ononin, and hispidol-4#-O-b-D-glucoside(Fig. 3). Thus, hispidol can be classified as a potentialand relatively potent antifungal compound in M.truncatula cell cultures. The direct precursor of hispi-dol, isoliquiritigenin, showed very weak antifungalactivity against P. medicaginis.
Selection of Putative Hispidol Synthases
Two primary mechanisms for aurone biosynthesishave been proposed based upon PPO (Nakayama
Figure 2. Accumulation of hispidol and hispidol-4#-O-b-D-glucosidein M. truncatula cell cultures in response to YE elicitation. A, HPLC-electrospray ionization-MS selected negative ion chromatograms ofm/z 253 in YE-elicited and controlM. truncatula cell cultures showing anincrease in two peaks denoted as P1 and P2. B, Full-scan, negative ionmass spectrum and UV data (inset) for P1 identified as hispidol-4#-O-b-D-glucoside (HG). C, Full-scan, negative ion mass spectrum and UVdata (inset) for P2 identified as hispidol (H). Both compound identifi-cations were confirmed through the coanalysis of authentic standards,which yielded matching retention time and spectral characteristics.[See online article for color version of this figure.]
Figure 3. Antifungal activity of induced phenolics detected in M.truncatula cell cultures. Radial mycelial growth of P. medicaginis onagar medium containing 0.1 mM phenolic compounds or dimethylsulfoxide (DMSO) measured every 24 h until mycelial growth of thesolvent control approached the edge of the petri dish. Error barsrepresent SE (n = 3). Means were separated using Duncan’s multiplerange test at P , 0.05.
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1099
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

et al., 2000, 2001) or PRX (Wong, 1967; Rathmell andBendall, 1972; Wilson and Wong, 1976) activity. Toidentify potential M. truncatula genes involved inhispidol biosynthesis, transcript profiles for the YE-elicited cell cultures were evaluated for correlatedexpression of putative PPOs and PRXs. Transcriptprofiling was performed using a custom M. truncatulamicroarray chip containing oligonucleotides repre-senting 16,086 TCs from theMedicago ESTand genomedatabases.
Potential involvement of PPOs in hispidol biosyn-thesis was investigated. BLASTn queries of the aur-eusidin synthase described previously (Nakayamaet al., 2000) against the M. truncatula gene indicesrevealed threeM. truncatula sequences with significantsimilarities (i.e. TC101697, P = 3.5e293; BG644378, P =2.7e212; and CX521674, P = 9.9e210). All three puta-tive PPOs were present on the M. truncatula micro-array, but the signals for all three were below thedetectable level in response to yeast elicitation. In addi-tion, the literature reports that alfalfa leaves and stemshave relatively little, if any, PPO activity (Sullivan et al.,2004; Sullivan and Hatfield, 2006). The above evidencediscounts the role of PPO-mediated hispidol biosyn-thesis in Medicago.
Putative PRXs were then evaluated as potentialhispidol synthases. The custom M. truncatula oligonu-cleotide microarray chip included 88 putative PRX TCsequences. Ten of the PRXs showed altered expressionin response to YE (Fig. 4A) and seven in response toMeJA (Fig. 4B; P , 0.05). Of the YE-modulated PRXTCs, six (TC106558, TC106564, TC106484, TC110836,TC102451, and TC107670) were up-regulated andthe others (TC102226, TC107346, TC102451, andTC101009) were down-regulated. The down-regulatedTCs were excluded as candidates (Fig. 4, A and B).Two of the YE-responding TCs (TC106564 andTC102451) were also induced in response to MeJAtreatment (Fig. 4, A and B) and, therefore, excluded,since metabolite profile data showed no induction ofhispidol or hispidol glucoside in response to MeJA(Fig. 4E). The induction of TC107670 was observedonly at single time point (i.e. after 2 h of YE treatment),which was inconsistent with the metabolite data.Therefore, this TC was also excluded, leaving threeputative candidate TCs (TC110836, TC106484, andTC106558). Replicated Affymetrix microarray analyses(Fig. 4C) were also performed at critical time points (2and 24 h postelicitation). The induction of all candi-date TCs was verified in the replicated Affymetrixmicroarray data and further confirmed by more rig-orous quantitative reverse transcription (qRT)-PCRanalysis of total RNA fromMeJA- and YE-treated cells(Fig. 4, D and F, respectively). In general, the qRT-PCRand Affymetrix microarray expression levels wereconsistent with the 16K custom oligonucleotide micro-array results. For example, the data showed that YEelicitor induced a significant transient increase from0.5 to 4 h in TC106558, whereas TC106484 andTC110836 showed a sustained increase from 2 to 24 h
(Fig. 4). At the metabolite level, both hispidol andhispidol-4#-O-b-D-glucoside showed similar inductionkinetics, with a sustained increase from 8 to 36 hpostelicitation (Fig. 4G). Both qRT-PCR and micro-array (Fig. 4) analyses showed no change or a milddecrease in the same three TCs in response to MeJA,which correlated with no change or moderate declinesin both hispidol and hispidol glucoside levels (Fig. 4E).In summary, the transcript profiles and qRT-PCR datacorrelated well with those of hispidol and hispidolglucoside accumulation, supporting the hypothesisthat TC110836, TC106484, and TC106558 might beinvolved in hispidol formation.
Nucleotide Sequence Analysis of PRX Genesin M. truncatula
TC110836, TC106484, and TC106558 (designatedMtPRX1, MtPRX2, and MtPRX3, respectively) werefully sequenced from both strands.MtPRX1,MtPRX2,and MtPRX3 encoded proteins containing 323, 355,and 357 amino acids with calculated pI values of 8.9,7.5, and 8.7, respectively. Strong amino acid similarity(79% identity at the amino acid level) was observedbetween MtPRX2 and MtPRX3, whereas MtPRX1shared only 54% identity with the other two PRXs.Deduced amino acid sequences showed all structuralmotifs required for functional class III PRX proteins.Highly conserved residues characteristic of heme-containing PRXs (Passardi et al., 2004) were identified,including two His residues interacting with the hemecenter (distal and proximal His residues) and eightCys residues forming disulfide bridges (Fig. 5A). Se-quence relatedness analysis of the three M. truncatulaPRXs and other functionally characterized members ofthe plant PRX classes indicated that the three MtPRXsdo not belong to class I ascorbate PRXs or class IIIanionic PRXs (Fig. 5B). MtPRX1 clustered with a groupof strongly cationic PRXs (pI. 8) andwasmost closelyrelated to a Nicotiana tabacum PRX (AB027752).MtPRX2 and MtPRX3 grouped with several otherlegume PRXs and exhibited the highest similaritiesto pathogen-induced PRXs from soybean (AF007211l)and alfalfa (X90693) at the amino acid level.
A C-terminal propeptide that targets PRXs for vac-uolar import (Welinder et al., 2002) was present inMtPRX2 and MtPRX3 but not in MtPRX1 (Fig. 5A).However, the possibility still exists that MtPRX1 istargeted to the vacuole due to its strong cationic nature(pI 8.9), as most cationic PRXs are vacuole localizeddue to the acidic nature of the vacuole (Ros Barceloet al., 2003). In addition, many vacuole-targeted pro-teins contain still undefined sorting signals (Carteret al., 2004).
In Silico Expression Analysis of MtPRX1, MtPRX2,and MtPRX3
In silico expression analysis was performed forMtPRX1,MtPRX2, andMtPRX3 to evaluate the spatial
Farag et al.
1100 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
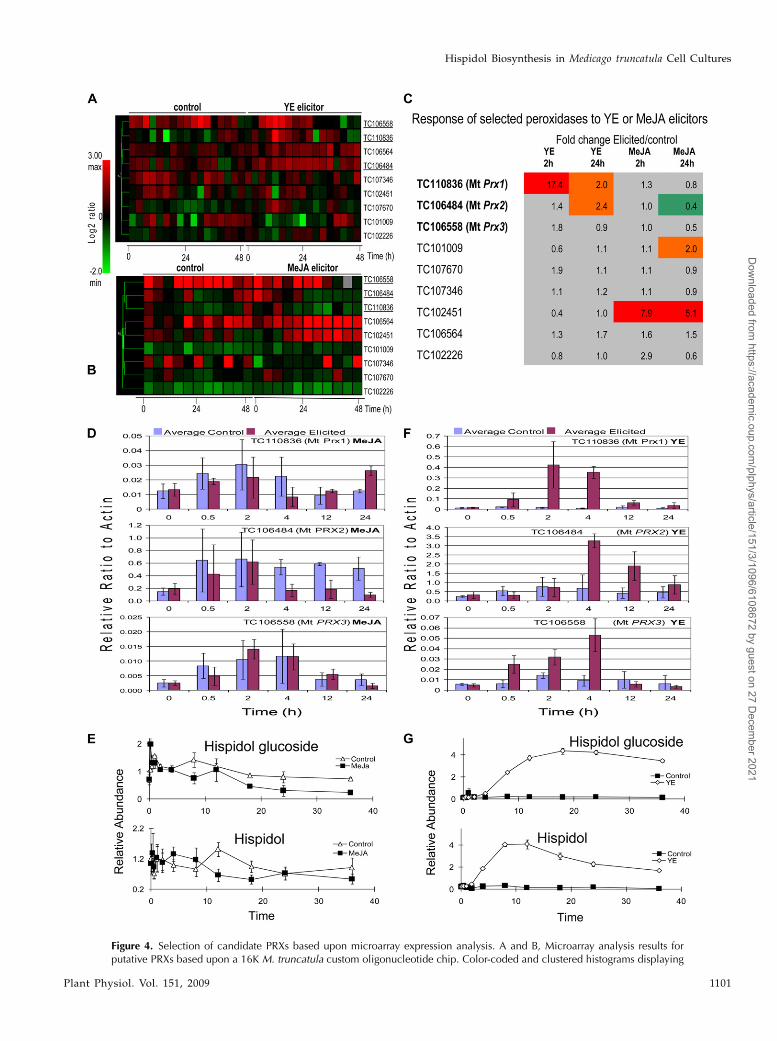
Figure 4. Selection of candidate PRXs based upon microarray expression analysis. A and B, Microarray analysis results forputative PRXs based upon a 16K M. truncatula custom oligonucleotide chip. Color-coded and clustered histograms displaying
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1101
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

expression of these PRXs (Fig. 6). Relative expressionlevel data were extracted from an expression atlasstudy where Affymetrix microarray data were col-lected for a series of M. truncatula tissues, includingleaf, petiole, stem, vegetative bud, flower, pods, andunnodulated root, as well as nodulated root develop-mental time course and a seed developmental timecourse (Benedito et al., 2008). MtPRX1 was mosthighly expressed in flower, pods, and roots. MtPRX2expression levels were approximately 200-fold higherin roots and nodules than in any other tissues.MtPRX3 expression levels were more evenly distrib-uted throughout all tissues.
Biochemical Characterization of Recombinant
PRX Enzymes
To determine the biochemical activities of each ofthe three candidate PRXs, their cDNAs were sub-cloned into a pET-28a expression vector and expressedin Escherichia coli BL21 (DE3) cells. Western-blot anal-ysis of the total protein extracts from isopropyl b-D-1-thiogalactopyranoside (IPTG)-induced cells harboringMtPRX1, MtPRX2, and MtPRX3 clones showed thepresence of a single protein band for each MtPRXbetween 34 and 39 kD and corresponding to thepredicted size of the recombinant proteins (Fig. 7).No protein bands were detected in cultures harboringan empty pET-28a vector (Fig. 7). Recombinant PRXproteins in cell lysates were tested for enzymaticactivity against guaicol, a phenolic substrate com-monly utilized in PRX enzymatic assays with equalamounts of total lysate protein used in each enzymaticreaction (see “Materials and Methods”). PRX enzy-matic activity was detected with all recombinant pro-teins: MtPRX1, 111 6 15 nmol mg21 min21; MtPRX2,105 6 21 nmol mg21 min21; and MtPRX3, 220 6 44nmol mg21 min21. It should be noted that variation inPRX enzymatic activity in cell lysates for MtPRX1, -2,and -3 could be attributed to minor differences inexpression levels of the recombinant proteins, as pu-rified recombinant proteins could not be isolated. NoPRX activity was detected in the cell lysate containingthe empty vector. To determine whether these PRXscould be involved in hispidol biosynthesis, isoliquir-itigenin was tested as a substrate and the productswere analyzed using HPLC-PDA-MS. In vitro assaysshowed the formation of hispidol from isoliquiritige-
nin using cell lysates expressing each of the threedifferent PRXs (Fig. 8, B–D). No aurone products wereobserved in the in vitro assays of control cells express-ing the empty vector (Fig. 8A). Although MtPRX2and MtPRX3 protein sequences are 79% identical (Fig.5A), their product profiles were quite different withisoliquiritigenin as a substrate. MtPRX2 yielded mainlyhispidol (Fig. 8C), whereas MtPRX3 formed pre-dominantly a quinol vinyl ether (QVE) and hispidolin lower amounts (Figs. 8D and 11). Furthermore,QVE was an observed product of horseradish perox-idase and not detected in elicited M. truncatula cellcultures.
Isoliquiritigenin-4-O-glucoside (Zhu et al., 2003) wasalso converted to hispidol glucoside by the recombi-nant PRXs (Fig. 11; Supplemental Fig. S2). A consid-erable proportion of the isoliquiritigenin glucosidewas isomerized in all in vitro assays to liquiritigeninglucoside, as flavanones are the thermodynamicallymore stable isomers of chalcones (Boland and Wong,1975). In the case of MtPRX3, a QVE glucoside was themajor product (Fig. 11).
To further assess MtPRX1 to -3 as putative vacuolarPRXs, enzymatic assays were performed that moni-tored the formation of hispidol from isoliquiritigeninby MtPRX1 to -3 as a function of pH over the range ofpH 3 to 9. Vacuolar enzymes tend to have loweroptimum pH, consistent with the acidic environmentof the vacuole, which is typically pH 5.5 (Taiz, 1992;Ros Barcelo et al., 2003), and is the case for snapdragonaureusidin synthase (Nakayama et al., 2000, 2001).Both MtPRX2 and -3 exhibited an activity optimum atpH 5, whereas MtPRX1 showed a slightly higheractivity optimum at pH 6 (Supplemental Fig. S3).The pH profiles of MtPRX2 and -3 were bimodal,with a small peak at pH 7.
Substrate Specificities and Mechanisms of MtPRXs
The ability of the recombinant MtPRXs to synthesizeaurones from a variety of chalcone substrates wasevaluated to determine the specificity of these enzy-matic reactions and the underlying mechanism(s) (Fig.11). Chalcones tested either lacked one of the three (4,2#, or 4#) hydroxyl groups present in isoliquiritigeninor contained an additional hydroxyl group on eitherthe A or B ring. Chalcones lacking the 2# or 4-hydroxylfunctions (i.e. compounds 6, 7, and 20) generated no
Figure 4. (Continued.)the changes in putative PRX expression levels in response to YE (A) or MeJA (B) elicitor from 0 to 48 h postelicitation comparedwith control unelicited cells and significant at P , 0.05. Red indicates up-regulated and green indicates down-regulated.Suspected PRXs related to hispidol biosynthesis are underlined. C, Replicated Affymetrix microarray analyses at key time points(2 and 24 h postelicitation) confirm the significant fold changes in MtPRX1 and MtPRX2 following YE. Other PRXs with foldchanges highlighted in gray were not significant. Correlation was seen betweenMtPRX transcripts and hispidol accumulation inM. truncatula cell cultures in response to YE or MeJA. D and F, qRT-PCR analysis of PRX transcripts in response to MeJA (D) or YE(F). E and G, Accumulation of hispidol and hispidol glucoside inM. truncatula cell suspensions in response to MeJA (E) or YE (G).The y axis values represent relative peak areas after normalization to the mean peak area for both compounds. Error barsrepresent SE (n = 3). Means are separated using Duncan’s multiple range test at P , 0.05.
Farag et al.
1102 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
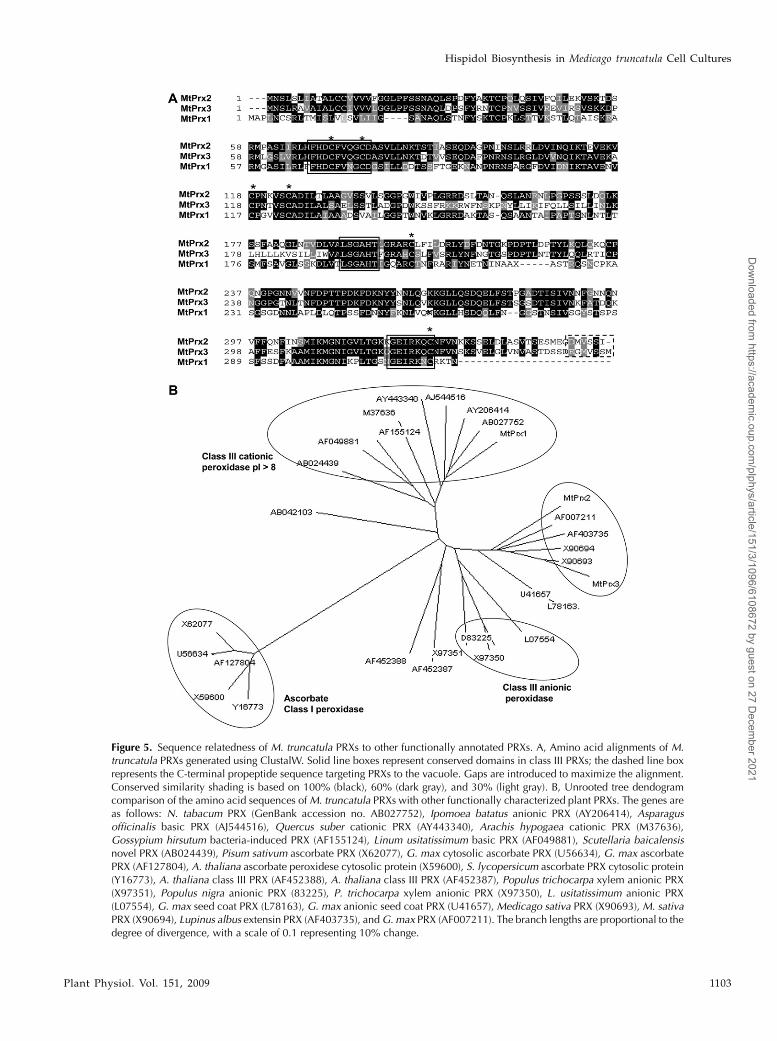
Figure 5. Sequence relatedness of M. truncatula PRXs to other functionally annotated PRXs. A, Amino acid alignments of M.truncatula PRXs generated using ClustalW. Solid line boxes represent conserved domains in class III PRXs; the dashed line boxrepresents the C-terminal propeptide sequence targeting PRXs to the vacuole. Gaps are introduced to maximize the alignment.Conserved similarity shading is based on 100% (black), 60% (dark gray), and 30% (light gray). B, Unrooted tree dendogramcomparison of the amino acid sequences ofM. truncatula PRXs with other functionally characterized plant PRXs. The genes areas follows: N. tabacum PRX (GenBank accession no. AB027752), Ipomoea batatus anionic PRX (AY206414), Asparagusofficinalis basic PRX (AJ544516), Quercus suber cationic PRX (AY443340), Arachis hypogaea cationic PRX (M37636),Gossypium hirsutum bacteria-induced PRX (AF155124), Linum usitatissimum basic PRX (AF049881), Scutellaria baicalensisnovel PRX (AB024439), Pisum sativum ascorbate PRX (X62077), G. max cytosolic ascorbate PRX (U56634), G. max ascorbatePRX (AF127804), A. thaliana ascorbate peroxidese cytosolic protein (X59600), S. lycopersicum ascorbate PRX cytosolic protein(Y16773), A. thaliana class III PRX (AF452388), A. thaliana class III PRX (AF452387), Populus trichocarpa xylem anionic PRX(X97351), Populus nigra anionic PRX (83225), P. trichocarpa xylem anionic PRX (X97350), L. usitatissimum anionic PRX(L07554), G. max seed coat PRX (L78163), G. max anionic seed coat PRX (U41657), Medicago sativa PRX (X90693), M. sativaPRX (X90694), Lupinus albus extensin PRX (AF403735), andG. max PRX (AF007211). The branch lengths are proportional to thedegree of divergence, with a scale of 0.1 representing 10% change.
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1103
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
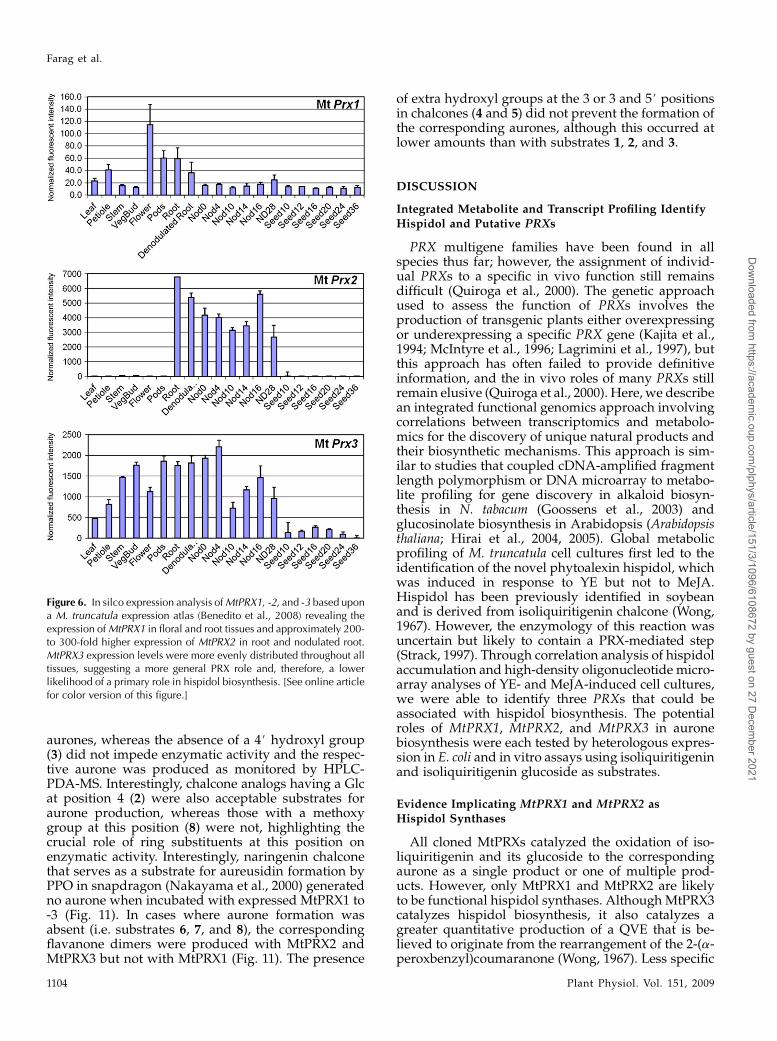
aurones, whereas the absence of a 4# hydroxyl group(3) did not impede enzymatic activity and the respec-tive aurone was produced as monitored by HPLC-PDA-MS. Interestingly, chalcone analogs having a Glcat position 4 (2) were also acceptable substrates foraurone production, whereas those with a methoxygroup at this position (8) were not, highlighting thecrucial role of ring substituents at this position onenzymatic activity. Interestingly, naringenin chalconethat serves as a substrate for aureusidin formation byPPO in snapdragon (Nakayama et al., 2000) generatedno aurone when incubated with expressed MtPRX1 to-3 (Fig. 11). In cases where aurone formation wasabsent (i.e. substrates 6, 7, and 8), the correspondingflavanone dimers were produced with MtPRX2 andMtPRX3 but not with MtPRX1 (Fig. 11). The presence
of extra hydroxyl groups at the 3 or 3 and 5# positionsin chalcones (4 and 5) did not prevent the formation ofthe corresponding aurones, although this occurred atlower amounts than with substrates 1, 2, and 3.
DISCUSSION
Integrated Metabolite and Transcript Profiling Identify
Hispidol and Putative PRXs
PRX multigene families have been found in allspecies thus far; however, the assignment of individ-ual PRXs to a specific in vivo function still remainsdifficult (Quiroga et al., 2000). The genetic approachused to assess the function of PRXs involves theproduction of transgenic plants either overexpressingor underexpressing a specific PRX gene (Kajita et al.,1994; McIntyre et al., 1996; Lagrimini et al., 1997), butthis approach has often failed to provide definitiveinformation, and the in vivo roles of many PRXs stillremain elusive (Quiroga et al., 2000). Here, we describean integrated functional genomics approach involvingcorrelations between transcriptomics and metabolo-mics for the discovery of unique natural products andtheir biosynthetic mechanisms. This approach is sim-ilar to studies that coupled cDNA-amplified fragmentlength polymorphism or DNA microarray to metabo-lite profiling for gene discovery in alkaloid biosyn-thesis in N. tabacum (Goossens et al., 2003) andglucosinolate biosynthesis in Arabidopsis (Arabidopsisthaliana; Hirai et al., 2004, 2005). Global metabolicprofiling of M. truncatula cell cultures first led to theidentification of the novel phytoalexin hispidol, whichwas induced in response to YE but not to MeJA.Hispidol has been previously identified in soybeanand is derived from isoliquiritigenin chalcone (Wong,1967). However, the enzymology of this reaction wasuncertain but likely to contain a PRX-mediated step(Strack, 1997). Through correlation analysis of hispidolaccumulation and high-density oligonucleotide micro-array analyses of YE- and MeJA-induced cell cultures,we were able to identify three PRXs that could beassociated with hispidol biosynthesis. The potentialroles of MtPRX1, MtPRX2, and MtPRX3 in auronebiosynthesis were each tested by heterologous expres-sion in E. coli and in vitro assays using isoliquiritigeninand isoliquiritigenin glucoside as substrates.
Evidence Implicating MtPRX1 and MtPRX2 asHispidol Synthases
All cloned MtPRXs catalyzed the oxidation of iso-liquiritigenin and its glucoside to the correspondingaurone as a single product or one of multiple prod-ucts. However, only MtPRX1 and MtPRX2 are likelyto be functional hispidol synthases. Although MtPRX3catalyzes hispidol biosynthesis, it also catalyzes agreater quantitative production of a QVE that is be-lieved to originate from the rearrangement of the 2-(a-peroxbenzyl)coumaranone (Wong, 1967). Less specific
Figure 6. In silco expression analysis ofMtPRX1, -2, and -3 based upona M. truncatula expression atlas (Benedito et al., 2008) revealing theexpression ofMtPRX1 in floral and root tissues and approximately 200-to 300-fold higher expression of MtPRX2 in root and nodulated root.MtPRX3 expression levels were more evenly distributed throughout alltissues, suggesting a more general PRX role and, therefore, a lowerlikelihood of a primary role in hispidol biosynthesis. [See online articlefor color version of this figure.]
Farag et al.
1104 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
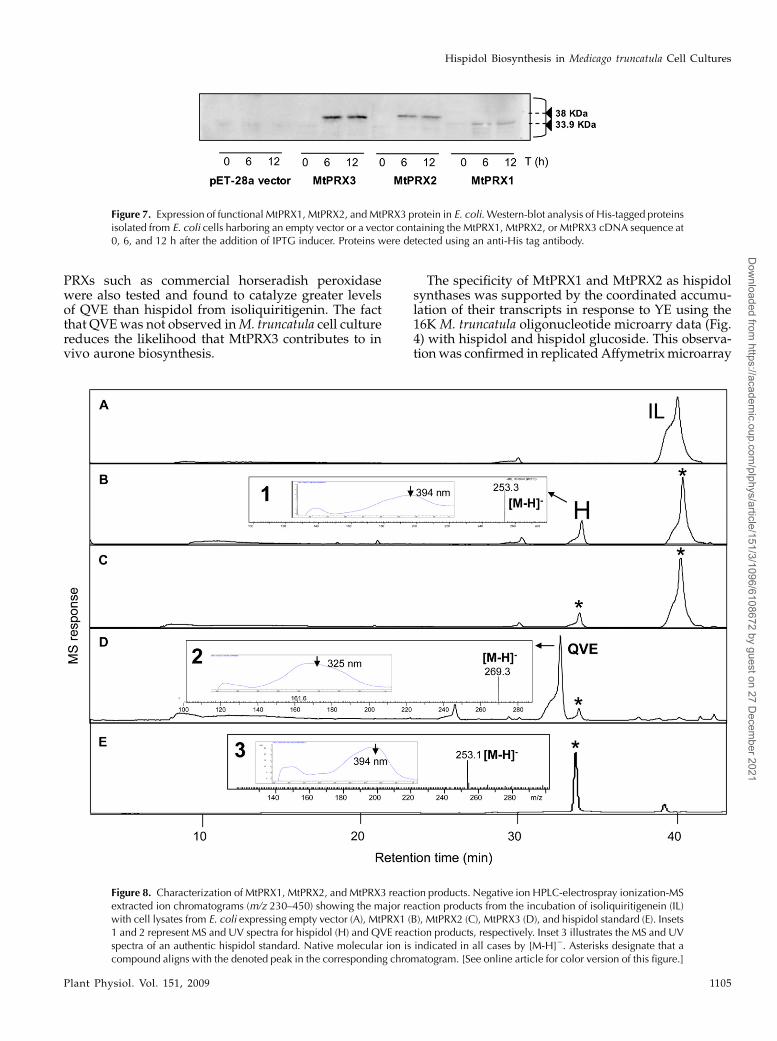
PRXs such as commercial horseradish peroxidasewere also tested and found to catalyze greater levelsof QVE than hispidol from isoliquiritigenin. The factthat QVEwas not observed inM. truncatula cell culturereduces the likelihood that MtPRX3 contributes to invivo aurone biosynthesis.
The specificity of MtPRX1 and MtPRX2 as hispidolsynthases was supported by the coordinated accumu-lation of their transcripts in response to YE using the16K M. truncatula oligonucleotide microarry data (Fig.4) with hispidol and hispidol glucoside. This observa-tionwas confirmed in replicatedAffymetrixmicroarray
Figure 8. Characterization of MtPRX1, MtPRX2, and MtPRX3 reaction products. Negative ion HPLC-electrospray ionization-MSextracted ion chromatograms (m/z 230–450) showing the major reaction products from the incubation of isoliquiritigenein (IL)with cell lysates from E. coli expressing empty vector (A), MtPRX1 (B), MtPRX2 (C), MtPRX3 (D), and hispidol standard (E). Insets1 and 2 represent MS and UV spectra for hispidol (H) and QVE reaction products, respectively. Inset 3 illustrates the MS and UVspectra of an authentic hispidol standard. Native molecular ion is indicated in all cases by [M-H]2. Asterisks designate that acompound aligns with the denoted peak in the corresponding chromatogram. [See online article for color version of this figure.]
Figure 7. Expression of functional MtPRX1, MtPRX2, andMtPRX3 protein in E. coli. Western-blot analysis of His-tagged proteinsisolated from E. coli cells harboring an empty vector or a vector containing the MtPRX1, MtPRX2, or MtPRX3 cDNA sequence at0, 6, and 12 h after the addition of IPTG inducer. Proteins were detected using an anti-His tag antibody.
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1105
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

analyses at 2 and 24 h postelicitation and by qRT-PCR.Other candidate M. truncatula PRXs were discountedbased upon the nonstatistically significant accumula-tion of their transcripts. Examples include TC101009,TC107670, TC107346, TC106564, and TC102226.
In silico expression analyses were also performedfor MtPRX1, MtPRX2, and MtPRX3 to evaluate thespatial expression of these PRXs (Fig. 6). MtPRX1 wasmost highly expressed in flower, pods, and roots.MtPRX2 expression levels were approximately 200-fold higher in roots and nodules than any othertissues. These data illustrate the high tissue specificityof MtPRX2, which would translate to the present cellcultures that were initiated from roots (Broecklinget al., 2005). Finally, MtPRX3 expression levels weremore evenly distributed throughout all tissues, sug-gesting amore general PRX role and, therefore, a lowerlikelihood of a primary role in hispidol biosynthesis.
To better assess the substrate specificities of each ofthe PRXs by kinetic analysis, multiple approacheswere exhaustively pursued to express and purify therecombinant MtPRX proteins. The proteins were het-erologously expressed in E. coli, yeast, and insect cell
systems. In all systems, only low levels of the PRXswere observed to accumulate, suggesting a potent celltoxicity of these enzymes in prokaryotic and eukaryoticsystems. Enrichment attempts using immobilizedmetalaffinity chromatography to purify the His-tagged pro-teins were also unsuccessful, and it is assumed that theHis tag was nonaccessible for affinity binding andpurification. Thus, quantitative kinetic data for thesePRXs could not be obtained.
Mechanism, Substrate Specificity, and Comparison ofPRX- and PPO-Mediated Aurone Biosynthesis
The mechanism for the PRX-mediated conversion ofisoliquiritigenin to hispidol proposed here is basedupon that originally described by Wong (1967) andWilson andWong (1976). This mechanism involves theformation of a phenoxy radical (II) that undergoesfurther self-rearrangement, leading to the formation ofa free radical (III; Fig. 9). The oxygenation of thea-carbon of III furnishes the peroxide IV, which, af-ter intramolecular Michael-like addition, provides adioxetane-type structure (V; Fig. 9). Intramolecular
Figure 9. A proposed biosyntheticpathway for isoliquiritigenin oxidationto hispidol and QVE as catalyzed byMtPRX1, MtPRX2, and MtPRX3 en-zymes in E. coli cell lysates (modifiedfrom Wong, 1967). The pathway isinitiated with the PRX-catalyzed gen-eration of a phenoxy radical (II), andthe subsequent steps most likely occurnonenzymatically. The additional stepsinclude radical rearrangement (III) fol-lowed by oxygenation of the a-carbon(IV) and epoxide cyclization with theb-carbon (V). The epoxide radical isthen utilized for the propagation ofan additional substrate radical (i.e.conversion of I to II), which initiates anew cycle of the reaction, and it isconverted to the neutral molecule.The neutral epoxide is then reducedand dehydrated to form VIII or theepoxide VII. These products thenreadily convert to hispidol or theQVE. The specific end products ob-served for each of the M. truncatulaPRXs are indicated in the final steps.
Farag et al.
1106 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021
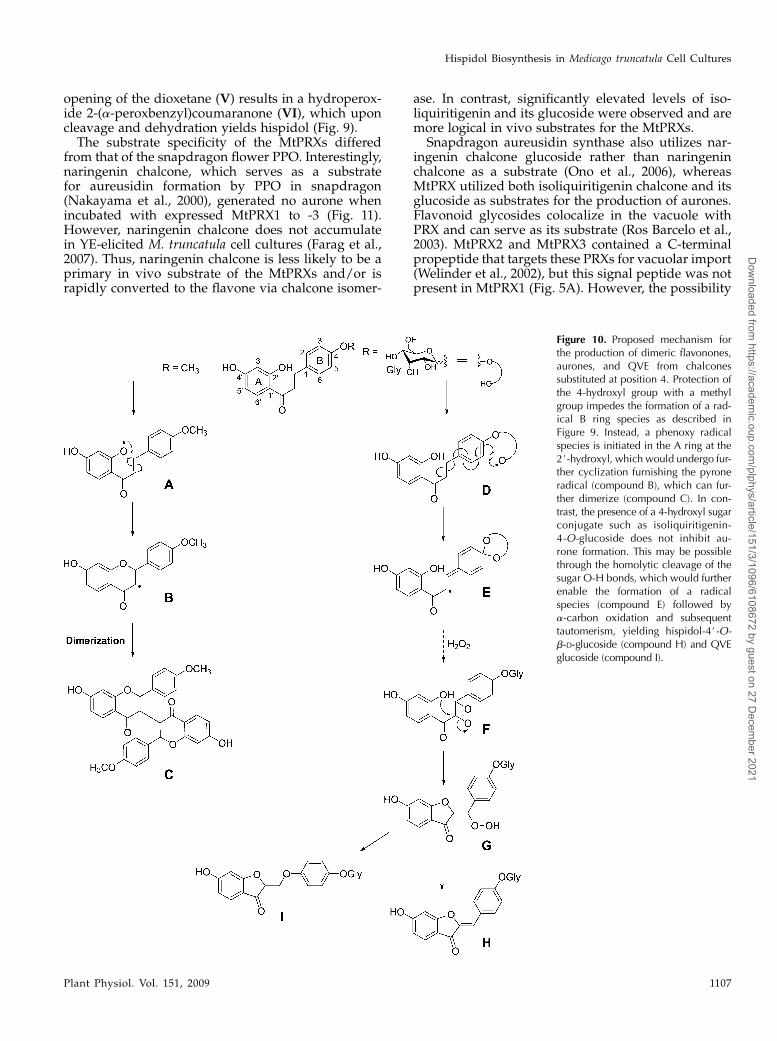
opening of the dioxetane (V) results in a hydroperox-ide 2-(a-peroxbenzyl)coumaranone (VI), which uponcleavage and dehydration yields hispidol (Fig. 9).The substrate specificity of the MtPRXs differed
from that of the snapdragon flower PPO. Interestingly,naringenin chalcone, which serves as a substratefor aureusidin formation by PPO in snapdragon(Nakayama et al., 2000), generated no aurone whenincubated with expressed MtPRX1 to -3 (Fig. 11).However, naringenin chalcone does not accumulatein YE-elicited M. truncatula cell cultures (Farag et al.,2007). Thus, naringenin chalcone is less likely to be aprimary in vivo substrate of the MtPRXs and/or israpidly converted to the flavone via chalcone isomer-
ase. In contrast, significantly elevated levels of iso-liquiritigenin and its glucoside were observed and aremore logical in vivo substrates for the MtPRXs.
Snapdragon aureusidin synthase also utilizes nar-ingenin chalcone glucoside rather than naringeninchalcone as a substrate (Ono et al., 2006), whereasMtPRX utilized both isoliquiritigenin chalcone and itsglucoside as substrates for the production of aurones.Flavonoid glycosides colocalize in the vacuole withPRX and can serve as its substrate (Ros Barcelo et al.,2003). MtPRX2 and MtPRX3 contained a C-terminalpropeptide that targets these PRXs for vacuolar import(Welinder et al., 2002), but this signal peptide was notpresent in MtPRX1 (Fig. 5A). However, the possibility
Figure 10. Proposed mechanism forthe production of dimeric flavonones,aurones, and QVE from chalconessubstituted at position 4. Protection ofthe 4-hydroxyl group with a methylgroup impedes the formation of a rad-ical B ring species as described inFigure 9. Instead, a phenoxy radicalspecies is initiated in the A ring at the2#-hydroxyl, which would undergo fur-ther cyclization furnishing the pyroneradical (compound B), which can fur-ther dimerize (compound C). In con-trast, the presence of a 4-hydroxyl sugarconjugate such as isoliquiritigenin-4-O-glucoside does not inhibit au-rone formation. This may be possiblethrough the homolytic cleavage of thesugar O-H bonds, which would furtherenable the formation of a radicalspecies (compound E) followed bya-carbon oxidation and subsequenttautomerism, yielding hispidol-4#-O-b-D-glucoside (compound H) and QVEglucoside (compound I).
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1107
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

still exists that MtPRX1 is targeted to the vacuole dueto its strong cationic nature (pI 8.9), as most cationicPRXs are vacuole localized due to the acidic nature ofthe vacuole (Ros Barcelo et al., 2003). In addition, theacidic pH optima for MtPRX1 to -3 are consistent withvacuolar localization. Vacuolar PRXs participate in theturnover and degradation of phenolic glycosides (RosBarcelo et al., 2003). The oxidative breakdown ofphenolics by PRXs begins with the enzymatic hydro-lysis of glycosides to release the aglycones, which arethen direct substrates for vacuolar PRXs. We hypoth-esize that M. truncatula PRXs utilize chalcone agly-cones as substrates produced via cleavage of itsglucoside in the vacuole under stress conditions toproduce the antifungal agent “hispidol.” In onion(Allium cepa), a similar deglucosidation of quercetinglucosides to quercetin yields the antifungal agent 3,4-dihydroxybenzoic acid as catalyzed by the action ofPRXs (Takahama and Hirota, 2000). The additional
hydroxyl group of chalcones containing the functionalmotif for aurone conversion increased their reactivityfor PPO but not for PRXs in M. truncatula. Both PPOand PRX mechanisms require the presence of 2#- and4-hydroxyl groups in the precursor chalcone.
Substitution of the 4-hydroxyl with a methoxy group(8) prevented conversion to aurone by any of the threeMtPRXs, whereas Glc attachment at this position (2)did not. Furthermore, the 4-methoxy-2#,4#-dihydroxy-chalcone substrate only yielded a flavanone dimer (16)with MtPRX2 and MtPRX3 (Fig. 11); trace levels weredetected with MtPRX1. Protection of the 4-hydroxylgroupwith a methoxy group inhibits the formation of aB ring radical species. Instead, a phenoxy radicalspecies is initiated in the A ring at the 2#-hydroxyl,which undergoes further cyclization, furnishing thepyrone radical (Fig. 10, compound B) and dimerizationto form compound C in Figure 10. Under our experi-mental conditions, the chalcones 6 and 7 (Fig. 11),
Figure 11. Substrates and related products formed during in vitro assays with recombinant M. truncatula PRXs. I, Aurone-producing chalcone substrates are listed with the amounts of aurones produced in each reaction expressed as a ratio relativeto that obtained with an empty vector in the following order (MtPRX1, MtPRX2, MtPRX3). 2 indicates absence, + indicatesratio, 10, ++ indicates ratio = 10 to 20, and +++ indicates ratio. 20. The relative ratio of each product formed by MtPRX1, -2,and -3 does not solely reflect difference in enzymatic activities but also differences in expression levels. II, Chalcones determinedto be inert as substrates for aurone or dimer formation. Names of the flavonoids are as follows: 1, isoliquiritigenin; 2,isoliquiritigenin-4-O-glucoside; 3, 2#,4-dihydroxychalcone; 4, 2#,4#,3,4-tetrahydroxychalcone; 5, 2#,4#,5#,3,4-pentahydroxy-chalcone; 6, 2#-hydroxy-4#-methoxychalcone; 7, 2#,4#-dihydroxychalcone; 8, 2#,4#-dihydroxy-4-methoxychalcone; 9, hispidol;10, hispidol-4#-O-b-D-glucoside; 11, 4#-hydroxyaurone; 12, 6,3#,4#-trihydroxyaurone (sulferitin); 13, 5,6,3#,4#-tetrahydroxyaur-one; 14, biflavanone A; 15, biflavanone B; 16, biflavanone C; 17, 6-hydroxy-2-(4-hydroxy-phenoxymethylene)-benzofuran-3-one(QVE); 18, 6-hydroxy-2-(4-glucosyl-phenoxymethylene)-benzofuran-3-one (QVEG); 19, 7, 2#,4#,6#-trimethoxy-2-hydroxychal-cone; 20, 4-methoxy-4#-hydroxychalcone.
Farag et al.
1108 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

which both contain 2#- and 4#-hydroxyl groups, alsoproduced the corresponding dimers. This suggeststhat the absence of a hydroxyl group at C4 or itsprotection with a methyl group provides the mecha-nistic diversion toward dimer formation. In contrast,the presence of the sugar hydroxyl groups in isoliquir-itigenin-4-O-glucoside may contribute to auroneformation. Homolytic cleavage of the O-H bonds ofthe sugar may also result in the formation of radicalspecies (Fig. 10, compound E) that then follow asimilar reaction mechanism, as proposed in Figure 9and the previous literature (Wong, 1967), yieldinghispidol-4#-O-b-D-glucoside (Fig. 10, compound H).Our data suggest that aurone biosynthesis is kineti-cally favored over the dimerization reaction, as dimerswere only produced as alternative oxidation productsby MtPRXs (Fig. 11) in situations where aurone for-mation was inhibited. Consistently, no dimers weredetected for the in vivo MtPRXs substrate isoliquir-itigenin and its respective glucoside. To the best of ourknowledge, these structure-activity relationships pro-vide the first mechanistic insights into aurone gluco-side biosynthesis in legumes.Hispidol biosynthesis in M. truncatula is unlikely to
occur via a PPO mechanism. This conclusion is basedupon the nondetectable levels of all three putativePPOs present on the M. truncatula microarray andbased upon our current mechanistic understanding.According to the reported PPO-catalyzed mechanismfor aurone synthesis (Nakayama et al., 2001), thesubstrate must contain hydroxyl groups at the 2# and4 positions, such that all products will then possess a3,4-dihydroxy-substituted B ring (Fig. 1). Hispidolcontains a 4-monohydroxy B ring and lacks the3-hydroxyl B ring group (Fig. 1); thus, the productionof hispidol via a PPO-catalyzedmechanism is unlikely.This argument is further supported by the fact that thepredicted substrate required for the PPO production ofhispidol would contain an unsubstituted B ring (i.e.2#,4#-dihydroxy chalcone or a 4-dehydroxy isoliquir-itigenin), which has not been identified in legumes todate. Nakayama et al. (2000, 2001) also suggest thathispidol biosynthesis occurs via a PRX-mediated asopposed to a PPO-mediated mechanism. Furthermore,alfalfa leaves and stems have been reported to haverelatively little if any PPO activity (Sullivan et al., 2004;Sullivan and Hatfield, 2006). The above evidence sup-ports the role of PRXs and discounts the role of PPO-mediated hispidol biosynthesis in legumes.
Is Hispidol a True Phytoalexin in Medicago?
Production of hispidol and hispidol glucoside islikely associated with the oxidative burst and produc-tion of H2O2, which are key components of plant dis-ease resistance (Lamb and Dixon, 1997). The presenceof an oxidative environment is supported by increasedH2O2 levels and PRX enzymatic activity detected in theM. truncatula culture medium in response to YE (Faraget al., 2008). Our data suggest that hispidol accu-
mulation requires the buildup of isoliquiritigenin,which is normally and rapidly shunted into the (iso)fla-vonoidpathwayby the activityof chalcone isomerase, inaddition to induction of a specific set of PRXs. Theaccumulated isoliquiritigenin and/or isoliquiritigeninglucoside serve as scavengers for removal of H2O2from the vacuole (Ros Barcelo et al., 2003) throughthe production of hispidol and hispidol glucoside byvacuole-targeted MtPRX1 and MtPRX2 activity. Manyflavonoids and lignans have powerful antioxidantactivity and can help remove H2O2 from acidic com-partments such as the vacuole by acting as substratesfor PRXs (Mehlhorn et al., 1996). Chalcones possesshigher antioxidant activity than flavanones and isofla-vones (Miranda et al., 2000). Thus, isoliquiritigenin andisoliquiritigenin glucoside are likely the best substratesfor PRX-mediated H2O2 removal in M. truncatula cellcultures.
On the basis of hispidol’s relative antifungal activityand its inducible synthesis from distant precursors,hispidol meets the criteria for classification as a phy-toalexin (VanEtten et al., 1994). The high antifungalactivity of hispidol relative to its biosynthetic precur-sor isoliquiritigenin illustrates how PRXs can effec-tively modify the cell’s potential antimicrobial activityprofile. Similar PRXs in YE-elicited cassava (Manihotesculenta) cell cultures were also found to enhance thefungal toxicity of the coumarins esculetin and scopo-letin up to 20-fold, but the oxidized products were notcharacterized (Gomez-Vasquez et al., 2004). However,the concept of hispidol as a true phytoalexin requiresthat the compound be detected in interactions ofMedicago with pathogens. To date, this has not beenobserved, but only a few interactions have been stud-ied. Neither can hispidol be induced in roots or leavesof M. truncatula following exposure to YE (data notshown). Furthermore, the presence of these MtPRXs intissues (e.g. roots) that do not appear to accumulatehispidol indicates that these enzymes have additionalfunctions and that hispidol formation may not even betheir true “function” but rather represents spillover ofmetabolite flux that cannot be rapidly incorporatedinto the main downstream branches of the phenyl-propanoid pathway. However, the genetic reprogram-ming associated with the establishment and elicitationof cell cultures can provide substrate pools and cata-lysts for the generation of novel metabolites. Suchflexibility of plant secondary metabolism provides abasis for the evolution of plant adaptation to changingenvironmental conditions, and the new knowledgegained from the observed plasticity of secondary me-tabolism can now be used for future metabolic engi-neering of this potential antifungal compound in planta.
MATERIALS AND METHODS
Cell Cultures and Elicitation
Cell cultures derived from barrel medic (Medicago truncatula ‘Jemalong’
line A17) roots were initiated, subcultured, elicited with YE and MeJA, and
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1109
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

harvested at 21 time points as described previously (Broeckling et al., 2005).
Triplicate biological replicates were collected for both control and elicited
samples at each time point, with each replicate collected from a separate
culture flask. Thus, each elicitation time course contained 126 independent
culture flasks and biological samples collected from each independent flask.
Cell culture medium was also sampled, but at a lower temporal resolution,
which included 0, 6, 12, 24, and 48 h postelicitation (Suzuki et al., 2005).
Chemicals and Reagents
Hispidol, coumestrol, afrormosin, and isoliquiritigenin-4-O-glucoside
were purchased from Apin Chemicals. Hispidol 4#-O-b-D-glucoside and
irisolidone were kindly provided by Dr. Tom Mabry (University of Texas,
Austin). Guaicol was purchased from Sigma-Aldrich. Other tested com-
pounds were purchased from Indofine. Solvents used were of HPLC-grade
purity. Naringenin chalcone was synthesized from naringenin according to
the method reported by Shimokoriyama (1957).
Extraction and HPLC-PDA-MS Analysis of
Phenolic Compounds
Phenolic compounds were extracted from lyophilized M. truncatula cells,
20 6 0.06 mg, with 1.8 mL of 80% methanol containing 2 mg of umbelliferone
(as an internal standard) for 10 h at room temperature using an orbital shaker
in the dark. Extract aliquots (1.4 mL) were centrifuged at 3,000g for 60 min,
and the supernatant was evaporated under a nitrogen stream until dry. The
residue was resuspended in 300 mL of 45% methanol, and samples were
analyzed by HPLC-PDA-MS. For analysis of phenolic compounds in the
culture medium, the medium was vacuum filtered from cell cultures and a
20-mL aliquot was extracted three times with 25 mL of ethyl acetate spiked
with 2 mg of umbelliferone (internal standard). The extracts were pooled,
evaporated to dryness using nitrogen, dissolved in 300 mL of methanol, and
analyzed by HPLC-PDA-MS.
An Agilent 1100 Series II HPLC system (Agilent Technologies) equipped
with a PDA detector was coupled to a Bruker Esquire ion-trap mass spectrom-
eter equipped with an electrospray ionization source. UV spectra were
obtained by scanning from 200 to 600 nm. A reverse-phase, C18, 5-mm, 4.6- 3250-mm column (J.T. Baker) was used for separations. Separations were
achieved using a linear gradient of 5% to 90% (v/v) B in 70 min. The mobile
phase consisted of eluentA (0.1% [v/v] acetic acid inwater) and eluent B (100%
acetonitrile). The flow rate was 0.8 mL min21, and the temperature of the
column was kept at 28�C. Both positive and negative ion mass spectra were
acquired. Positive ion electrospray ionization was performed using an ion
source voltage of 4.0 kV and a capillary offset voltage of 86.0 V. Nebulization
was aided with a coaxial nitrogen sheath gas provided at a pressure of 60 p.s.i.
Desolvationwas aided using a counter current nitrogenflow set at a pressure of
12 p.s.i. and a capillary temperature of 300�C.Mass spectra were recorded over
the range of 50 to 2,200 m/z. The Bruker ion-trap mass spectrometer was
operated under an ion current control of approximately 10,000 with a maxi-
mum acquire time of 100 ms. Chromatographic peak areas of cell culture
extracts were normalized based on the areas of recovered internal standard
peaks. Relativemetabolite abundances were calculated using a custom PEARL
script to extract peak areas of individual ions characteristic of each component
(Broeckling et al., 2006). Metabolites were identified through spectral and
retention time comparisonswith authentic compoundsprepared in an identical
manner. Peak areas were further normalized by dividing each peak area value
by the mean peak area value for that compound, with each time course treated
independently. ANOVAwas performed using JMP5 (SAS Institute) statistical
software.Meanswere separatedusingDuncan’smultiple range test atP, 0.05.
The cumulative metabolite profiling has been published previously (Farag
et al., 2008) and is compliant with the recentMetabolomics Standards Initiative
(Fiehn et al., 2007a, 2007b; Sumner et al., 2007).
Enzymatic Hydrolysis and Analysis of P1
Collected P1 fractions were evaporated, resuspended in 200 mL of water,
and subjected to b-glucosidase hydrolysis for 8 h at 37�C (Mabry et al., 1970).
The aglycone was then partitioned from the aqueous phase into ethyl acetate
(3 3 2 mL). The ethyl acetate fraction was evaporated to dryness under
nitrogen, and the residue was resuspended in 100 mL of methanol and
analyzed using HPLC-PDA-MS to characterize the aglycone. The aqueous
phase following liquid-liquid extraction with ethyl acetate and containing the
liberated sugar(s) was evaporated in a SpeedVac to complete dryness. The
sugars were derivatized as described (Farag et al., 2007) and analyzed by GC-
MS in parallel with authentic sugars (e.g. Glc, Gal, and malonic acid).
DNA Microarray Analysis
RNA Isolation, cDNA Labeling, and Hybridization
Total RNAwas isolated from 0.5 g ofM. truncatula suspension cells using 5
mL of Tri-Reagent (Molecular Research Center) following the manufacturer’s
protocol. The Amino Allyl cDNA Labeling Kit (Ambion) was used to label 25
mg of total RNA following the manufacturer’s protocol. The product was
resuspended in 70 mL of Microarray Hybridization Solution, version 2
(Amersham Biosciences), denatured at 95�C to 100�C for 2 min, pipetted
onto the slides, and covered with a coverslip (Corning) before sealing in a
hybridization chamber (Corning). The sealed chambers were wrapped in
aluminum foil and incubated at 42�C for 16 h. The arrays were subsequently
washed with 13 SSC, 0.1% (w/v) SDS, followed by a wash in 0.53 SSC, 0.01%
SDS, and a third wash with 0.053 SSC, at room temperature for 5 min each.
The slides were dried by centrifugation.
Custom M. truncatula OligonucleotideMicroarray Analysis
The 70-mer oligonucleotides representing 16,086 TCs from the Medicago
Genome (Qiagen Operon Oligo Set version 1.0) were printed on ArrayIt Brand
SuperAmine Substrate glass slides (Telechem) using an OmniGrid 100 (Gene
Machine). The oligonucleotides were normalized to melting temperature,
sequence, and concentration. Themelting temperatureswerewithin 78�C6 5�C.Oligonucleotides corresponded to the sequences of TCs located within 1,000
bases from the 3# ends, with less than 70% identity to all other genes and with
less than 20 contiguous bases common to any other gene. The oligonucleotides
also had no homopolymer tracts greater than eight bases and no hairpin stems
longer than nine bases. The array dimensions were 39 mm long3 18 mmwide,
with a spot size of approximately 100 to 110 mm in diameter and spot spacing of
185 3 185 mm. The array configuration used eight-by-four subarray grids. Each
subarray grid was composed of 23 3 23 spots. Slides were stored in a vacuum-
sealed box at room temperature and UV cross-linked at 200 mJ using a
Stratalinker (Stratagene) before use.
Each cDNA sample was hybridized against a reference cDNA sample that
was prepared from a common pool of reference cDNA, and a reference design
of the micorarray experiment was performed. Reference RNA was purified
from separate M. truncatula ‘Jemalong’ (line A17) cell suspension cultures
(maintained for approximately 3 years in subculture) and pooled for use. Cy3
dye (Amersham Biosciences) was used for labeling reference RNA, and Cy5
was used for experimental RNA. The oligonucleotide microarray data are
publicly available in a custom database for “omics” data (DOME; http://
medicago.vbi.vt.edu/data.html).
Affymetrix M. truncatula Microarray Analysis
The Affymetrix DNA microarray contained 61,200 probe sets. These were
composed of 32,167 M. truncatula EST/mRNA-based and chloroplast gene-
based probe sets (The Institute for Genomic Research [TIGR] Gene Index
version 8, January 2005, 36,878 unique sequences); 18,733 M. truncatula
International Medicago Genome Annotation Group and phase 2/3 bacterial
artificial chromosome prediction-based probe sets; 1,896 M. sativa EST/
mRNA-based probe sets; and 8,305 Sinorhizobium meliloti gene prediction-
based probe sets. For microarray experiments, total RNAwas extracted from
cell suspension cultures (Suzuki et al., 2005; Naoumkina et al., 2007) using TRI
Reagent (Molecular Research Center) and purified using the RNeasyMinElute
Cleanup Kit (Qiagen) according to the manufacturer’s instructions. For all
samples, 10 mg of total RNA was used for labeling reactions. Labeling,
hybridization, and data processing were performed according to the manu-
facturer’s instruction (Affymetrix). For both YE and MeJA treatments, tran-
script profiles were examined at early (2 h) and late (24 h) times after
elicitation. For each sample point, two biological replicates were included, and
the average signal intensities were used to calculate the ratio of treatment to its
corresponding control. The Affymetrix microarray data are publicly available
at ArrayExpresss (http://www.ebi.ac.uk/arrayexpress, ID = E-MEXP-1092).
Farag et al.
1110 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

Microarray Data Processing and Analysis
Arrays were imaged with a ScanArray 4000 scanner (Packard) at 10 mm
resolution and variable photomultiplier tube voltage settings to obtain the
maximal signal intensities. The fluorescence intensity for each label and each
element on the array was captured using GenePix Pro 4.1 (Axon). Normaliza-
tion of Cy3 and Cy5 signals was performed by adjusting the signal intensities of
the two images using a Lowess (subgrid) method and GeneTraffic software
(www.iobion.com). The local background was subtracted from the values of
each spot on the array. The statistical ANOVA of normalized data was
performed using GeneSpring software (www.silicongenetics.com).
qRT-PCR Analysis
Microarray results were confirmed by qRT-PCR using TC-specific primers
for tissue collected at 0 min, 30 min, 2 h, 4 h, 12 h, and 24 h postelicitation.
RNAwas isolated from triplicate control and elicited cells. Ten micrograms of
purified total RNA was treated with Turbo DNA-free DNase I (Ambion)
according to the manufacturer’s protocol, purified by RNeasy MinElute
columns (Qiagen), and checked for genomic DNA contamination by PCR
using the M. truncatula-specific actin primers MtActin-for (5#-TCAATG-
TGCCTGCCATGTATGT-3#) and MtActin-rev (5#-ACTCACACCGTCACCA-
GAATCC-3#). RNA integrity was evaluated with an Agilent 2100 Bioanalyzer
using RNA nanochips. RT reactions were performed using SuperScript
III reverse transcriptase (Invitrogen) according to the manufacturer’s
protocol. PCRs were performed on an optical 384-well plate with an ABI
PRISM 7900 HT sequence detection system (Applied Biosystems) using
SYBR Green to monitor double-stranded DNA synthesis. PCRs (10 mL final
volume) contained 5 mL of SYBR Green Master Mix reagent (Applied
Biosystems), 1 mL of cDNA, and 200 nM of each gene-specific primer:
MtPrx1for (5#-GTATGCCCCGGAGTTGTATC-3#), MtPrx1rev (5#-CCGTTTT-
AGCATCTGTTCTTCC-3#), MtPrx2for (5#-GTGCTTACTGGAAAGAAAGG-
AGA-3#), MtPrx2rev (5#-GAGGTCACACTAGCAAGATCCA-3#), MtPrx3for
(5#-GAAACAAGGAGAGATTAGAAAACA-3#), and MtPrx3rev (5#-CATTG-
AGCTAACCATACCCTCATC-3#). PCRs were performed as described else-
where (Czechowski et al., 2005). Data were analyzed using SDS 2.2.1 software
(Applied Biosystems). PCR efficiency was estimated using LinRegPCR soft-
ware as described (Ramakers et al., 2003), and the transcript levels were
determined by relative quantification (Pfaffl, 2001) using actin (TC107326) as a
reference gene (primers as above).
MtPRX Sequence Analyses
BLAST searches were performed using BLASTp version 2.2.2 (http://
www.ncbi.nih.gov/BLAST). The amino acid sequences were aligned using
ClustalW, and the similarities were calculated using Megalign (DNASTAR).
Publicly available programs (http://www.expacy.ch/tools/pi_tools.html/
and http://www.scripps.edu/~cdputnam/protcalc.html) were used to com-
pute pI and Mr for the deduced amino acid sequences. The subcellular
targeting of proteins was predicted using the programs PSORT (http://psort.
ims.u-tokyo.ac.jp/form.html), TargetP (http://www.cbs.dtu.dk/services/
TargetP/), and SignalP (http://www.cbs.dtu.dk/services/signalP/).
Functional Characterization of Recombinant PRXsExpressed in Escherichia coli
M. truncatula EST clones NF041G08NR (i.e. MtPRX1 corresponding to
TIGR release 8.0 TC110836), NF092A02EC (i.e. MtPRX2 corresponding
to TIGR release 8.0 TC106484), and NF089H06PL (i.e. MtPRX3 correspond-
ing to TIGR release 8.0 TC106558) were sequenced from both ends using M13
primers and found to contain full-length sequences. cDNA sequences
were PCR amplified from pBluescript II SK+ (Stratagene) using primers
with EcoRI and XhoI (MtPRX1 and MtPRX3) or SacI and HindIII (MtPRX2)
sites (underlined): MtPRX1F, 5#-AGGATTGAATTCATGGCTCCTTTAA-
ATTGT-3#; MtPRX1R, 5#-TTAACCTCGAGATTAGTTGGTTTTTCTACA-3#;MtPRX2F, 5#-CACATAGAGCTCAAAATGAATTCCCTTAGT-3#; MtPRX2R,
5#-ACCCACAAGCTTTTAGATTGAACTAACCAT-3#; MtPRX3F, 5#-TCAC-
AAGAATTCATGAACTCCCTTCGTGCT-3#; and MrPRX3R, 5#-CAATCA-
CTCGAGTTACATTGAGCTA-3#. Purified PCR products were digested with
appropriate restriction enzymes (Novagen) for 2 h and purified from an
agarose gel using the QIAquick gel extraction kit (Qiagen). The excised PCR
products were cloned into appropriately digested pET-28a (+) vector (Nova-
gen). The resulting plasmids were sequenced to confirm identity.
E. coli BL21 (DE3) cells harboring the pET-28a or MtPRX expression con-
structs were grown to an optical density of 600 nm of 0.8, and expression was
initiated by addition of IPTG to a final concentration of 0.5 mM, with further
incubation while shaking overnight at 20�C. Cultures were centrifuged, and
pellets were resuspended in a lysis buffer (50 mM Tris, pH 8.0, 500 mM NaCl, 20
mM imidazole, pH 8.0, 10% glycerol, 1% Tween 20, and 1 mM phenylmethyl-
sulfonyl fluoride). Cells were lysed by sonication (Biologics) and centrifuged,
and the resulting supernatant was removed and assayed for PRX activity
against guaicol. For PRX activity assays, cell lysates (50 mg of protein
equivalent) were added to 2 mL of 0.1% guaiacol and 0.03% H2O2 in 50 mM
potassium acetate, pH 6.0, and the increase in A470 over 3 min was measured
using a Beckman Coulter DU800 spectrophotometer. For all PRX enzymatic
reactions, equal amounts of protein were used as quantified using the
Bradford (1976) assay and using a commercial dye reagent (Bio-Rad) with
bovine serum albumin as the standard.
Product profiling of chalcone analogs incubated with recombinant PRXs
was assayed in 500 mL of 10 mM potassium phosphate buffer, pH 6.7,
containing 50 mg of protein equivalent of cell lysate, 8 mL of 5 mM substrate
dissolved in dimethyl sulfoxide, and 1 mL of 100 mM H2O2. Reactions were
carried out at 37�C for 2 h and stopped by adding 3 volumes of ethyl acetate.
After vortexing, the ethyl acetate phase was removed and evaporated to
dryness under nitrogen, and the residue was resuspended in methanol prior
to HPLC-PDA-MS analysis using the same instrumental conditions men-
tioned previously. All enzyme assays were performed in duplicate.
Protein Analysis
E. coli cell lysates were separated on a 12% Tris-HCl gel (30 mg of protein
loaded per lane) and electroblotted onto a nitrocellulose membrane (Transblot;
Bio-Rad) according to the procedure of Sambrook and Russell (2001). Blots
were incubated overnight at 4�C with the anti-His tag monoclonal antibody
(Novagen) diluted 1:1,000 in 3% bovine serum albumin, followed by incuba-
tion with anti-mouse horseradish peroxidase (Amersham). The serological
color reaction was developed using a color reagent (Amersham) according to
the manufacturer’s instructions. Western-blot analyses were performed in
duplicate with a representative blot presented.
Antifungal Agar Plate Bioassays
Antifungal assays were conducted essentially as described (Blount et al.,
1992). Stock plates of Phoma medicaginis were grown on potato dextrose agar
medium (Difco Laboratories) in sterile 100- 3 15-mm petri dishes until the
mycelia covered one-third of the plate. Compounds to be tested were
dissolved in dimethyl sulfoxide (Sigma) at stock concentrations of 20 mM.
The solvent (as a control) or compounds were then added to the melted agar
medium at 5 mL mL21 to give a final phenolic concentration of 0.1 mM. Five-
milliliter samples of medium were aseptically pipetted into sterile 35- 3 10-
mm petri dishes and allowed to cool. Using a sterile 4-mm cork borer, agar
discs with mycelia were cut from a stock fungal culture plate and placed on
the center of treatment and control plates. Radial mycelial growth was
measured from the edge of the disc to the outer edge of the mycelia, and the
results of treatments were calculated as the relative percentages of their
corresponding solvent controls.
Sequence data from this article can be found in the GenBank/EMBL data
libraries under the following accession numbers: MtPRX1, EF456703;
MtPRX2, EF456704; MtPRX3, EF456705.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Accumulation of hispidol in medium of YE-
elicited and control M. truncatula cell cultures.
Supplemental Figure S2. In vitro enzymatic assays using recombinant
MtPRX1, -2, and -3 with isoliquiritigenin-4-O-b-D-glucoside (ILG) re-
vealing the reaction product hispidol-4#-O-b-D-glucoside (HG).
Supplemental Figure S3. PRX activity of MtPRX1, -2, and -3 as a function
of assay pH.
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1111
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

ACKNOWLEDGMENTS
We thank Prof. T.J. Mabry for his contribution of authentic hispidol
glucoside and related compounds. We thank Drs. Michael Udvardi, Vagner
Benedito, Yuhong Tang, and Ivone Torres-Jerez for supplying us with the M.
truncatula microarray atlas data used to extract in silico expression data for
MtPRX1, MtPRX2, and MtPRX3. We also thank all those involved in the
large-scale culturing, elicitation, and rapid harvesting of M. truncatula sus-
pension cultures: Lahoucine Achnine, Courtney Allen, Stacy Allen, Victor
Asirvatham, Naveed Aziz, Jack W. Blount, Fang Chen, John Cooper, Anthony
Duran, Patrick Fennell, Xian Zhi He, Lisa Jackson, Parvathi Kota, Changjun
Liu, Srinu Reddy, Gail Shadle, Shashi Sharma, Hideyuki Suzuki, Ivone
Torres-Jerez, Bonnie Watson, and Deyu Xie.
Received May 18, 2009; accepted June 29, 2009; published July 1, 2009.
LITERATURE CITED
Barz W, Beimen A, Drager B, Jaques U, Otto C, Super E, Upmeier B (1990)
Turnover and storage of secondary products in cell cultures. In BV
Charlwood, MJC Rhodes, eds, Secondary Products from Plant Tissue
Culture. Kluwer, Dordrecht, The Netherlands, pp 79–101
Benedito VA, Torres-Jerez I, Murray JD, Andriankaja A, Allen S, Kakar
K, Wandrey M, Verdier J, Zuber H, Ott T, et al (2008) A gene expression
atlas of the model legume Medicago truncatula. Plant J 55: 504–513
Bino RJ, Hall RD, Fiehn O, Kopka J, Saito K, Draper J, Nikolau BJ,
Mendes P, Roessner-Tunali U, Beale MH, et al (2004) Potential of
metabolomics as a functional genomics tool. Trends Plant Sci 9: 418–425
Blount JW, Dixon RA, Paiva NL (1992) Stress responses in alfalfa (Medicago
sativa L.). XVI. Antifungal activity of medicarpin and its biosynthetic
precursors: implications for the genetic manipulation of stress metab-
olites. Physiol Mol Plant Pathol 41: 333–349
Boland M, Wong E (1975) Purification and kinetic properties of chalcone-
flavanone isomerase from soya bean. Eur J Biochem 50: 383–389
Boumendjel A (2003) Aurones: a subclass of flavones with promising
biological potential. Curr Med Chem 10: 2621–2630
Bradford MM (1976) A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding. Anal Biochem 72: 248–254
Broeckling CD, Huhman DV, Farag MA, Smith JT, May GD, Mendes P,
Dixon RA, Sumner LW (2005) Metabolic profiling ofMedicago truncatula
cell cultures reveals the effects of biotic and abiotic elicitors on metab-
olism. J Exp Bot 56: 323–336
Broeckling CD, Reddy IR, Duran AL, Zhao X, Sumner LW (2006) MET-
IDEA: data extraction tool for mass spectrometry-based metabolomics.
Anal Chem 78: 4334–4341
Carter C, Pan S, Zouhar J, Avila EL, Girke T, Raikhel NV (2004) The
vegetative vacuole proteome of Arabidopsis thaliana reveals predicted
and unexpected proteins. Plant Cell 16: 3285–3303
Chittoor J, Leach J, White F (1997) Differential induction of a peroxidase
gene family during infection of rice by Xanthomonas oryzae pv. oryzae.
Mol Plant Microbe Interact 10: 861–871
Cook DR (1999) Medicago truncatula: a model in the making! Curr Opin
Plant Biol 2: 301–304
Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible WR (2005)
Genome-wide identification and testing of superior reference genes for
transcript normalization in Arabidopsis. Plant Physiol 139: 5–17
Dixon RA, Sumner LW (2003) Legume natural products: understanding
and manipulating complex pathways for human and animal health.
Plant Physiol 131: 878–885
Farag MA, Huhman DV, Dixon RA, Sumner LW (2008) Metabolomics
reveals novel pathways and differential mechanistic and elicitor-
specific responses in phenylpropanoid and isoflavonoid biosynthesis
in Medicago truncatula cell cultures. Plant Physiol 146: 387–402
Farag MA, Huhman DV, Lei Z, Sumner LW (2007) Metabolic profiling and
systematic identification of flavonoids and isoflavonoids in roots and
cell suspension cultures of Medicago truncatula using HPLC-UV-ESI-MS
and GC-MS. Phytochemistry 68: 342–354
Fiehn O, Robertson D, Griffin J, van der Werf M, Nikolau B, Morrison N,
Sumner L, Goodacre R, Hardy N, Taylor C, et al (2007a) The metab-
olomics standards initiative (MSI). Metabolomics 3: 175–178
Fiehn O, Sumner L, Rhee S, Ward J, Dickerson J, Lange B, Lane G,
Roessner U, Last R, Nikolau B (2007b) Minimum reporting standards
for plant biology context information in metabolomic studies. Metab-
olomics 3: 195–201
Fridman E, Pichersky E (2005) Metabolomics, genomics, proteomics, and
the identification of enzymes and their substrates and products. Curr
Opin Plant Biol 8: 242–248
Gomez-Vasquez R, Day R, Buschmann H, Randles S, Beeching JR,
Cooper RM (2004) Phenylpropanoids, phenylalanine ammonia lyase
and peroxidases in elicitor-challenged cassava (Manihot esculenta)
suspension cells and leaves. Ann Bot (Lond) 94: 87–97
Goossens A, Hakkinen ST, Laakso I, Seppanen-Laakso T, Biondi S, De
Sutter V, Lammertyn F, Nuutila AM, Soderlund H, Zabeau M, et al
(2003) A functional genomics approach toward the understanding of
secondary metabolism in plant cells. Proc Natl Acad Sci USA 100:
8595–8600
Hernandez G, Ramirez M, Valdes-Lopez O, Tesfaye M, Graham MA,
Czechowski T, Schlereth A, Wandrey M, Erban A, Cheung F, et al
(2007) Phosphorus stress in common bean: root transcript and metabolic
responses. Plant Physiol 144: 752–767
Hirai MY, Klein M, Fujikawa Y, Yano M, Goodenowe DB, Yamazaki Y,
Kanaya S, Nakamura Y, KitayamaM, Suzuki H, et al (2005) Elucidation
of gene-to-gene and metabolite-to-gene networks in Arabidopsis by
integration of metabolomics and transcriptomics. J Biol Chem 280:
25590–25595
Hirai MY, Yano M, Goodenowe DB, Kanaya S, Kimura T, Awazuhara M,
Arita M, Fujiwara T, Saito K (2004) Integration of transcriptomics and
metabolomics for understanding of global responses to nutritional
stresses in Arabidopsis thaliana. Proc Natl Acad Sci USA 101: 10205–10210
Kajita S, Osakabe K, Katayama Y, Kawai S, Matsumoto Y, Hata K,
Morohoshi N (1994) Agrobacterium-mediated transformation of poplar
using a disarmed binary vector and the overexpression of a specific
member of a family of poplar peroxidase genes in transgenic poplar cell.
Plant Sci 103: 231–239
Kaplan F, Kopka J, Haskell DW, Zhao W, Schiller KC, Gatzke N, Sung
DY, Guy CL (2004) Exploring the temperature-stress metabolome of
Arabidopsis. Plant Physiol 136: 4159–4168
Kawalleck P, Schmelzer E, Hahlbrock K, Somssich I (1995) Two pathogen-
responsive genes in parsley encode a tyrosine-rich hydroxyproline-rich
glycoprotein (hrgp) and an anionic peroxidase. Mol Gen Genet 247:
444–452
Lagrimini LM, Gingas V, Finger F, Rothstein S, Liu T (1997) Character-
ization of antisense transformed plants deficient in the tobacco anionic
peroxidase. Plant Physiol 114: 1187–1196
Lamb C, Dixon RA (1997) The oxidative burst in plant disease resistance.
Annu Rev Plant Physiol Plant Mol Biol 48: 251–275
Lei Z, Elmer AM, Watson BS, Dixon RA, Mendes PJ, Sumner LW (2005) A
two-dimensional electrophoresis proteomic reference map and system-
atic identification of 1367 proteins from a cell suspension culture of the
model legume Medicago truncatula. Mol Cell Proteomics 4: 1812–1825
Mabry TJ, Harborne JB, Thomas MB (1970) The Systematic Identification
of Flavonoids. Springer-Verlag, New York
McIntyre C, Bettenay H, Manners J (1996) Strategies for the suppression of
peroxidase gene expression in tobacco. II. In vivo suppression of
peroxidase activity in transgenic tobacco using ribozyme and antisense
constructs. Transgenic Res 5: 263–270
Mehlhorn H, Lelandais M, Korth H, Foyer C (1996) Ascorbate is the
natural substrate for plant peroxidases. FEBS Lett 378: 203–206
Miranda C, Stevens J, Ivanov V, McCall M, Frei B, Deinzer M, Buhler D
(2000) Antioxidant and prooxidant actions of prenylated and non-
prenylated chalcones and flavanones in vitro. J Agric Food Chem 48:
3876–3884
Nakayama T, Sato T, Fukui Y, Yonekura-Sakakiba K, Hayashi H, Tanaka
Y, Kusumi T, Nishino T (2001) Specificity analysis and mechanism of
aurone synthesis catalyzed by aureusidin synthase, a polyphenol oxi-
dase homolog responsible for flower coloration. FEBS Lett 499: 107–111
Nakayama T, Yonekura-Sakakibara K, Sato T, Kikuchi S, Fukui Y,
Fukuchi-Mizutani M, Ueda T, Nakao M, Tanaka Y, Kusumi T, et al
(2000) Aureusidin synthase: a polyphenol oxidase homolog responsible
for flower coloration. Science 290: 1163–1166
Naoumkina M, Farag MA, Sumner LW, Tang Y, Liu CJ, Dixon RA (2007)
Different mechanisms for phytoalexin induction by pathogen and
wound signals in Medicago truncatula. Proc Natl Acad Sci USA 104:
17909–17915
Farag et al.
1112 Plant Physiol. Vol. 151, 2009
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021

Ono E, Hatayama M, Isono Y, Sato T, Watanabe R, Yonekura-Sakakibara
K, Fukuchi-Mizutani M, Tanaka Y, Kusumi T, Nishino T, et al (2006)
Localization of a flavonoid biosynthetic polyphenol oxidase in vacuoles.
Plant J 45: 133–143
Pare PW, Dmitrieva N, Mabry TJ (1991) Phytoalexin aurone induced in
Cephalocereus senilis liquid suspension culture. Phytochemistry 30:
1133–1135
Passardi F, Longet D, Penel C, Dunand C (2004) The class III peroxidase
multigenic family in rice and its evolution in land plants. Phytochem-
istry 65: 1879–1893
Peters N, Frost J, Long S (1986) A plant flavone, luteolin, induces expres-
sion of Rhizobium meliloti nodulation genes. Science 233: 977–980
Peters NK, Long SR (1988) Alfalfa root exudates and compounds which
promote or inhibit induction of Rhizobium meliloti nodulation genes.
Plant Physiol 88: 396–400
Pfaffl MW (2001) A new mathematical model for relative quantification in
real-time RT-PCR. Nucleic Acids Res 29: e45
Quiroga M, Guerrero C, Botella MA, Barcelo A, Amaya I, Medina MI,
Alonso FJ, de Forchetti SM, Tigier H, Valpuesta V (2000) A tomato
peroxidase involved in the synthesis of lignin and suberin. Plant Physiol
122: 1119–1128
Ralston L, Subramanian S, Matsuno M, Yu O (2005) Partial reconstruction
of flavonoid and isoflavonoid biosynthesis in yeast using soybean type I
and type II chalcone isomerases. Plant Physiol 137: 1375–1388
Ramakers C, Ruijter J, Deprez R, Moorman A (2003) Assumption-free
analysis of quantitative real-time polymerase chain reaction (PCR) data.
Neurosci Lett 339: 62–66
Rathmell W, Bendall D (1972) The peroxidase-catalysed oxidation of a
chalcone and its possible physiological significance. Biochem J 127:
125–132
Ros Barcelo A, Pomar F, Lopez-SerranoM, PedrenoMA (2003) Peroxidase:
a multifunctional enzyme in grapevines. Funct Plant Biol 30: 577–591
Sambrook J, Russell D (2001) Molecular Cloning: A Laboratory Manual.
Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Schaarschmidt S, Gonzalez MC, Roitsch T, Strack D, Sonnewald U,
Hause B (2007) Regulation of arbuscular mycorrhization by carbon: the
symbiotic interaction cannot be improved by increased carbon avail-
ability accomplished by root-specifically enhanced invertase activity.
Plant Physiol 143: 1827–1840
Shimokoriyama M (1957) Interconversion of chalcones and flavanones of a
phloroglucinol-type structure. J Am Chem Soc 79: 4199–4202
Strack D (1997) Phenolic Metabolism. In PM Dey, JB Harborne, eds, Plant
Biochemistry. Academic Press, San Diego, pp 387–416
Sullivan ML, Hatfield RD (2006) Polyphenol oxidase and o-diphenols inhibit
postharvest proteolysis in red clover and alfalfa. Crop Sci 46: 662–670
Sullivan ML, Hatfield RD, Thoma SL, Samac DA (2004) Cloning and
characterization of red clover polyphenol oxidase cDNAs and expres-
sion of active protein in Escherichia coli and transgenic alfalfa. Plant
Physiol 136: 3234–3244
Sumner L, Amberg A, Barrett D, Beale M, Beger R, Daykin C, Fan T, Fiehn
O, Goodacre R, Griffin J, et al (2007) Proposed minimum reporting
standards for chemical analysis. Metabolomics 3: 211–221
Sumner LW, Mendes P, Dixon RA (2003) Plant metabolomics: large-scale
phytochemistry in the functional genomics era. Phytochemistry 62: 817–
836
Suzuki H, Reddy MS, Naoumkina M, Aziz N, May GD, Huhman DV,
Sumner LW, Blount JW, Mendes P, Dixon RA (2005) Methyl jasmonate
and yeast elicitor induce differential transcriptional and metabolic re-
programming in cell suspension cultures of the model legume Medicago
truncatula. Planta 220: 696–707
Taiz L (1992) The plant vacuole. J Exp Biol 172: 113–122
Takahama U, Hirota S (2000) Deglucosidation of quercetin glucosides
to the aglycone and formation of antifungal agents by peroxidase-
dependent oxidation of quercetin on browning of onion scales. Plant
Cell Physiol 41: 1021–1029
Taylor CF, Paton NW, Garwood KL, Kirby PD, Steaf DA, Yin Z, Deutsch
EW, Selway L, Walker J, Riba-Garcia I, et al (2003) A systematic
approach to modeling, capturing and disseminating proteomics exper-
imental data. Nat Biotechnol 21: 247–254
Tohge T, Nishiyama Y, Hirai M, Yano M, Nakajima J, Awazuhara M,
Inoue E, Takahashi H, Goodenowe D, Kitayama M, et al (2005)
Functional genomics by integrated analysis of metabolome and tran-
scriptome of Arabidopsis plants over-expressing an MYB transcription
factor. Plant J 42: 218–235
VanEtten HD, Mansfield JW, Bailey JA, Farmer EE (1994) Two classes of
plant antibiotics: phytoalexins versus “phytoanticipins.” Plant Cell 6:
1191–1192
Weckwerth W, Loureiro ME, Wenzel K, Fiehn O (2004) Differential
metabolic networks unravel the effects of silent plant phenotypes.
Proc Natl Acad Sci USA 101: 7809–7814
Welinder KG, Justesen AF, Kjaersgard IVH, Jensen RB, Rasmussen SK,
Jespersen HM, Duroux L (2002) Structural diversity and transcription
of class III peroxidases from Arabidopsis thaliana. Eur J Biochem 269:
6063–6081
Wilson JM, Wong E (1976) Peroxidase catalysed oxygenation of 4,2#,4#-trihydroxychalcone. Phytochemistry 15: 1333–1341
Wong E (1967) Formation of 4#,6-dihydroxy-2-(alpha-hydroxybenzyl)cou-maranone from 2#,4,4#-trihydroxychalcone by cell-free extracts of soy-
bean. Phytochemistry 6: 1227–1233
Yonekura-Sakakibara K, Tohge T, Matsuda F, Nakabayashi R, Takayama
H, Niida R, Watanabe-Takahashi A, Inoue E, Saito K (2008) Compre-
hensive flavonol profiling and transcriptome coexpression analysis
leading to decoding gene-metabolite correlations in Arabidopsis. Plant
Cell 20: 2160–2176
Young ND, Cannon SB, Sato S, Kim D, Cook DR, Town CD, Roe BA,
Tabata S (2005) Sequencing the genes paces of Medicago truncatula and
Lotus japonicus. Plant Physiol 137: 1174–1181
Zhu H, BilginM, Snyder M (2003) Proteomics. Annu Rev Biochem 72: 783–812
Hispidol Biosynthesis in Medicago truncatula Cell Cultures
Plant Physiol. Vol. 151, 2009 1113
Dow
nloaded from https://academ
ic.oup.com/plphys/article/151/3/1096/6108672 by guest on 27 D
ecember 2021





























