Embed Size (px)
Citation preview

Ei
IEa
b
c
a
ARRA
KAHESAM
1
iwMsftM(1lSeh2mtpv
0d
Industrial Crops and Products 32 (2010) 639–649
Contents lists available at ScienceDirect
Industrial Crops and Products
journa l homepage: www.e lsev ier .com/ locate / indcrop
ssential oils, genetic relationships and in vitro establishment of Helichrysumtalicum (Roth) G. Don ssp. italicum from wild Mediterranean germplasm
. Morone-Fortunatoa, C. Montemurrob, C. Rutaa,∗, R. Perrinia, W. Sabettab, A. Blancob,. Lorussoc, P. Avatoc
Dipartimento di Scienze delle Produzioni vegetali, Università “Aldo Moro”, Via Amendola 165/A, I-70125 Bari, ItalyDipartimento di Biologia e Chimica Agro-Forestale ed Ambientale, sezione di Genetica, Università “Aldo, Moro”, Via Amendola 165/A, I-70125 Bari, ItalyDipartimento Farmaco-Chimico, Università “Aldo Moro”, Via Orabona 4, I-70125 Bari, Italy
r t i c l e i n f o
rticle history:eceived 6 May 2010eceived in revised form 26 July 2010ccepted 30 July 2010
a b s t r a c t
In this paper we describe the morphological, histological and chemical characters of 20 native Helichry-sum italicum (Roth) G. Don ssp. italicum genotypes collected from different locations in Italy and Corsica(France) and grown under similar edaphic and climatic conditions. An AFLP technique was used to exam-ine the level of genetic variability among the genotypes with the aim to disclose a possible correlationbetween the genetic and chemical data. The chemical analysis recognizes at least three different chemo-
eywords:steraceaeelichrysum italicum ssp. italicumssential oilecretory glands
types based on the major constituents in the essential oils. This was confirmed by the AFLP analysisresulting in a dendrogram divided into three main clades. Because of the polymorphic trait of Helichry-sum species, an efficient micropropagation protocol was established for the 20 H. italicum (Roth) G. Donssp. italicum genotypes to guarantee the availability of stable genetic material with defined chemicalprofiles for industrial applications. The genotypes were found to influence the in vitro performance and
of th
FLP markersicropropagationthe chemical composition
. Introduction
The genus Helichrysum (Miller) belongs to the Asteraceae fam-ly and is a very large genus including approximately 600 species
idespread all over the world. The genus is represented in theediterranean area by nearly 25 native species. Among them, eight
pecies are present in Italy: Helichrysum italicum (Roth) G. Don; H.rigidum (Labill.) Willd. (endemic of Sardinia and Corsica); H. mon-elinasanum E. Schmid and H. saxatile Moris (restricted to Sardinia,
onte Linas); H. nebrodense Heldr. and H. siculum (Sprengel) Boiss.endemic of Sicily); H. rupestre (Rafin.) DC.; H. stoechas (Pignatti,984). The strong trend of Helichrysum species to show a high
evel of anatomical and morphological polymorphism (Jahn andchönfelder, 1995), creates a diversity of several varieties and/orcotypes (Pignatti, 1984). A high chemical and genetic diversityas also been reported (Harborne and Turner, 1984; Angioni et al.,003; Tundis et al., 2005). H. italicum (syn. H. angustifolium DC) is the
ost widespread species in the Italian environment and includeswo subspecies, H. italicum ssp. italicum and H. italicum ssp. micro-hyllum (Willd.) Nyman (Pignatti, 1984). Besides its ornamentalalue, the success of this species is also due to the several activ-
∗ Corresponding author. Tel.: +39 0805443045; fax: +39 0805442976.E-mail address: [email protected] (C. Ruta).
926-6690/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.indcrop.2010.07.023
e essential oils, but not the phenotypic traits of the plants.© 2010 Elsevier B.V. All rights reserved.
ities related to the essential oils produced by the glandular hairspresent on the leaves and flower heads of the plant.
Metabolites isolated from H. italicum and especially its volatileoil have been found to display many biological properties, such asthe anti-inflammatory (Sala et al., 2001, 2002, 2003; Appendinoet al., 2007; Voinchet and Giraud-Robert, 2007), anti-allergic andantimicrobial (Chinou et al., 1997; Nostro et al., 2001, 2002, 2004;Voinchet and Giraud-Robert, 2007), antioxidant (Sala et al., 2002)and anti-viral (Nostro et al., 2003; Appendino et al., 2007). Eventhough this large genus has been extensively investigated, thechemical variations and the genetic relationships within the samespecies still remain unclear. This is especially true for the essentialoil composition and the genetic relationship of genotypes belong-ing to H. italicum (Roth) G. Don species. Compared to other plants,the use of molecular markers to exploit its genetic variability hasalso been limited. Angioni et al. (2003) carried out a molecularcharacterization of two H. italicum chemotypes with RAPD (ran-dom amplified polymorphism DNA) technique and Smissen et al.(2007) used AFLP (amplified fragment length polymorphism) fin-gerprinting to investigate intergeneric hybridization between the
endemic New Zealand species of H. lanceolatum and Anaphalioidesbellidioides.Following our previous investigation on H. italicum ssp. micro-phyllum (Perrini et al., 2009b), we describe the morphological,histological and chemical characters of 20 wild H. italicum (Roth)

640 I. Morone-Fortunato et al. / Industrial Crop
Table 1Genotypes collection and relative code number of H. italicum (Roth) G. Don ssp.italicum.
Code number Genotype
2 Brindisi di Montagna (Potenza)3 Località Centopozzi (Foggia)5 Località Baia di Crovani (Corsica)6 Marina di Pulsano (Taranto)7 Monopoli (Bari)8 Noci (Bari)9 Orto Botanico (Bari)
10 Policoro (Matera)11 Località Pollino, Sanseverino Lucano (Potenza)13 Putignano (Bari)14 Ruggiano, Manfredonia (Foggia)15 Sambuchello (Foggia)16 Sammichele di Bari (Bari)17 Località Torre Gattarella, Vieste (Foggia)18 Località Valle del Sinni (Basilicata)19 Località Valle dell’Inferno (Foggia)20 Locorotondo (Bari)23 Località Punta Polveraia (Isola D’Elba)
Giee2b
satm2s
2
2
FCfnomNtDaab
t
2
(lbe
in a final volume of 50 �l. PCR thermal conditions were 21 cycles
26 Orto Botanico (Siena)27 Termini Imerese (Palermo)
. Don ssp. italicum genotypes collected from different locationsn Italy and from Corsica (France) and then grown under the samedaphic and climatic conditions at Bari, Italy. Moreover, AFLP mark-rs were used to examine the level of genetic variability among the0 genotypes with the objective to disclose a possible correlationetween genetic and chemical data.
The polymorphic traits of Helichrysum species cannot guaranteetable genetic material with defined chemical profiles for industrialpplications. To overcome this, an efficient micropropagation pro-ocol to reproduce and stabilize selected clones of H. italicum ssp.ichrophyllum was recently established by our group (Perrini et al.,
009b). The usefulness of this protocol was verified in the currenttudy on H. italicum ssp. italicum genotypes.
. Materials and methods
.1. Plant material
Twenty H. italicum (Roth) G. Don ssp. italicum (Table 1 andig. 1) genotypes collected from different areas of Italy andorsica (France) were maintained in an open-field (“La Pietra”
arm—Monopoli countryside, Bari, South Italy) and irrigated whenecessary through micro-irrigation applying water at a volumef 300 m3/ha. The soil was characterized as red in colour, with aedium depth, good structure and medium fertility (1.336 g kg−1
; 127.78 mg kg−1 P2O5; 268.23 mg kg−1 K2O; 2.08% organic mat-er; pH = 7.7). The climate of the area is typically Mediterranean.uring mild, rainy winters, the average monthly temperature isround 9.5 ◦C and during hot, dry summers 23.2 ◦C. The meannnual precipitation is 573 mm of which 36% from April to Septem-er with a relative humidity of 68%.
Genotypes are identified by a code number referring to the loca-ion of collection of the plants (Table 1).
.2. Taxonomic identification
The Italian National Flora was used for taxonomic identification
Pignatti, 1984). The morphological parameters used were basaleaves length, basal leaves width, trichomes diameter, trichomesasal cell diameter, external bracts glandular hairs, number of flow-rs/head, number of heads/corymb, and head diameters.s and Products 32 (2010) 639–649
2.3. Histology
Closed flower section and sections (40–50 �m thick) obtainedby sliding microtome of fresh leaf tissues from plants grown in thefield and in vitro propagated were stained with Sudan IV (Gomori,1952). Single flowers and leaf sections were mounted on slidesto examine the secretory glands directly under a light microscope(Zeiss).
2.4. Analysis of the essential oils
Plants of the different H. italicum (Roth) G. Don ssp. italicumgenotypes were harvested at the flowering stage and air-driedbefore the extraction of the essential oils. Dried flower heads weresteam-distilled to recover the essential oils (2 h 30 min; with aSpring type apparatus, Albrigi, Italy). Oils were kept in the refriger-ator until analyzed.
A Trace GC Ultra Thermo Finnigan gas chromatograph equippedwith a FID detector was used for the compositional analysis of theessential oils. Samples (1 �l) were injected in the cold on-columnmode in a DB-5 (J&W Scientific) fused silica capillary columnof 30 m × 0.25 mm; 0.25 �m film thickness. The chromatographicconditions were as follows: detector temperature 300 ◦C; columntemperature was programmed from 60 ◦C (5 min isothermal) to270 ◦C (30 min isothermal) at 4 ◦C/min. Arithmetic indices (AI)were measured against a series of straight-chain alkanes (C7–C27).Hydrogen was the carrier gas (35 kP; 2.0 ml/min). Data were pro-cessed using the Chrom-Card 32-bit computing software.
Gas chromatography–mass spectrometry (GC–MS) analyseswere performed with a Hewlett Packard 6890-5973 mass spec-trometer interfaced with a HP Chemstation according to Avato etal. (2005). Samples (1 �l) were injected using the splitless sam-pling technique. Identification of the oil constituents was based oncomparison of their GC retention times in combination with arith-metic indices (AI) and by means of reference mass spectra fromstandard compounds and/or from library files (Joulain and König,1998; Avato et al., 2005; Adams, 2007).
2.5. Genetic analysis
DNA was extracted from 50 mg of fresh leaves with the GeneElute Plant kit (Sigma) following the manufacture’s instructions.DNA concentration was determined using a spectrophotometer at260 nm, and by means of electrophoresis on 0.8% agarose gel with�DNA standard.
AFLP analysis was conducted essentially as described by Voset al. (1995) with some modifications according to the indica-tions reported in Montemurro et al. (2005, 2008). Genomic DNA(250 ng) was double digested for 1 h at 37 ◦C in a final volume of40 �l with 10 units of PstI and 10 units of MseI (New England Bio-lab), and 1× R/L restriction/ligation buffer (10 mM Tris–HCl, pH 7.5,10 mM magnesium acetate, 50 mM potassium acetate). To this mix-ture 10 �l of a ligation mix containing 50 pmol of double-strandedadapters for MseI and 5 pmol adapters for PstI, 3.5 units T4 ligase(Invitrogen) and 1× R/L restriction/ligation buffer, were added andlet to react overnight at 15 ◦C. Thirty microliters of the resultingdigestion–ligation mixture were used, without dilution, for PCRpre-amplification by adding 1× PCR buffer Euroclone (160 mM(NH4)2SO4, 670 mM Tris–HCl, pH 8.8, 0.1% Tween 20), 50 mMMgCl2, 75 ng of primer MseI (+1N) and 375 ng of primer PstI (+1N),0.2 mM of each dNTP, 2 units of Taq DNA polymerase (Euroclone),
of 30 s at 94 ◦C, 1 min at 56 ◦C, and 1 min at 72 ◦C. A I-Cycler (Bio-rad) thermal cycler was used. Five microliters of pre-amplificationproducts were used as a template for selective amplification byadding 1× PCR buffer Euroclone (160 mM (NH4)2SO4, 670 mM

I. Morone-Fortunato et al. / Industrial Crops and Products 32 (2010) 639–649 641
e anal
TMADwta0ttpAptve
aaucUw
2
2
etw((aTo
Fig. 1. Four selected genotypes (5-7-9-26) among th
ris–HCl, pH 8.8, 0.1% Tween 20), 50 mM MgCl2, 96 ng primerseI (5′-GATGAGTCCTGAGTAA-3′ + 3N) and 3.2 ng primer PstI (5′-GACTGCGTACATGCAG-3′ + 3N), 0.2 mM dNTP each, 0.5 units TaqNA polymerase (Euroclone), in a final volume of 10 �l. PstI primersere radiolabelled with �-[33P]-ATP. The following PCR condi-
ions were used: 94 ◦C for 3 min, 12 cycles of 30 s at 94 ◦C, 30 st 65 ◦C (the annealing temperature was reduced every cycle by.7 ◦C) and 1 min at 72 ◦C. Twenty-three additional cycles werehen done at: 30 s at 94 ◦C, 30 s at 56 ◦C and 1 min at 72 ◦C. Selec-ive PCR was performed in a MJR-PTC200 thermal cycler. Threerimer combinations with three selective nucleotides were used.mplified fragments were separated using a denaturing 5% (w/v)olyacrylamide gel electrophoresis (PAGE) (OWL Separation Sys-em), with a 100 bp DNA ladder (MBI-fermentas). Bands wereisualised by autoradiography and scored manually for their pres-nce or absence.
AFLP polymorphic bands were scored as either present (1) orbsent (0) to create a binary matrix. All the molecular data werenalyzed using the NTSYS program. The SIMQUAL function, wassed to compute a similarity analysis of qualitative data by the Jac-ard index. The resulting matrix was subsequently analyzed by thenweighted Pair Group Method (UPGMA), and the similarity treeas obtained with the SAHN clustering program.
.6. Micropropagation
.6.1. Establishment and multiplication stageApical and axillary buds, 4–5 mm length, were used as the
xplant source. Buds were sterilized in a 0.1% (w/v) HgCl2 solu-ion for 15 min and successively rinsed with distilled-sterileater. For in vitro cultures, we used the basic culture medium
BM), containing the macronutrients, the micronutrients, FeEDTA25 mg l−1), thiamine HCl (0.4 mg l−1), myoinositol (100 mg l−1),gar (7 g l−1) according to Morone-Fortunato and Avato (2008).he pH of the medium was adjusted to 5.6–5.8. Sterilizationf culture media was performed in an autoclave at 121 ◦C for
yzed clones of H. italicum (Roth) G. Don ssp. italicum.
20 min. Primary explants were cultured on BM enriched with6-benzylaminopurine + indole-3-butyric acid (BAP 1 mg l−1 + IBA0.2 mg l−1), with sucrose (20 g l−1) added as the carbon source(Perrini et al., 2009b). A total of 30 primary explants per geno-type were put into 70 ml culture tubes (Sigma) containing 20 mlmedium. Microcuttings were established approximately after 4weeks of culture, when shoots had reached 3–4 cm length anddeveloped 5–6 nodes. The efficiency of the culture medium wasdetermined by recording the percentage of primary explants devel-oping shoots, the number of shoots/explant and the shoot length.Shoots were subcultured three times and then transferred ontofresh multiplication media for proliferation.
2.6.2. Root formation stageEach regenerated shoot was rooted in 500 ml jars containing
100 ml of BM with sucrose (30 g l−1) but without growth regula-tors or supplemented with indole-3-butyrrc acid (IBA 0.2 mg l−1)(Perrini et al., 2009b). Each jar contained five shoots; for each treat-ment a total of 25 shoots were used. The percentage of rootingshoots and the number of roots/shoot was scored after 15 days. Allcultures were incubated in a growth chamber at 21 ± 1 ◦C with aphotoperiod of 16 h light, under a light intensity of 50 �E m−2 s−1.
2.6.3. Acclimatization stageThe in vitro rooted young plants were rinsed with tap water
to remove adhering medium and individually transferred to thegreenhouse (constant 50% humidity level, and at 21 ± 3 ◦C temper-ature), in Jiffy-Pots or in pots (8 cm diameter) with a peat mixture(organic carbon 46%, organic nitrogen 1–2%, organic matter 80%)mixed with perlite or sand at 1:1 (v/v) ratio. The percentage ofplants surviving was recorded after 30 days.
2.7. Cytology
Root tips were treated for 4 h with a 0.05% aqueous solution ofcolchicine (Sigma) and then fixed in ethanol–acetic acid 3:1 (v/v).

6 al Crop
So1
2
bvv(ef
3
3
aDm1hdghr2r67mwod
3
Himaimgottbttt
3
ttpa
app
42 I. Morone-Fortunato et al. / Industri
tandard Feulgen-staining and squashing procedures were used tobtain the preparations for the cytological analysis (Blanco et al.,996).
.8. Statistical analysis
The experiments were conducted under a complete randomizedlock design with 3 replications. Data were processed by analysis ofariance (ANOVA) and comparisons within and between the meanalues of treatments were made by the Student–Newman–KeulsSNK) test calculated at the confidence level of P ≤ 0.01. Dataxpressed as percentages were transformed using angular trans-ormation.
. Results
.1. Taxonomic identification
The morphological characters used in this study identifiedll the genotypes as belonging to the H. italicum ssp. italicum.istinctive traits of each analyzed genotypes were in good agree-ent with those reported in the literature (Table 2; Pignatti,
984), except for the number of flowers/head and the number ofeads/corymb. Observed differences for these characters reflect theistinct behaviour of the various genotypes even though they wererown under the same conditions. Only genotypes 10, 11 and 14ave in fact the number of heads/corymb comparable to valueseported in the literature (Pignatti, 1984). Genotypes 2, 3, 23 and7 developed the number of heads/corymb of 42, 48, 38 and 43,espectively, that is slightly higher than reported, while genotypes, 8, 15, 16 and 26 showed much higher values, 80, 78, 72, 80, and7, respectively (Table 2). The remaining genotypes showed inter-ediate values in the range of 58–61 heads/corymb. Another traitorth noting is the low density or total absence of secretory glands
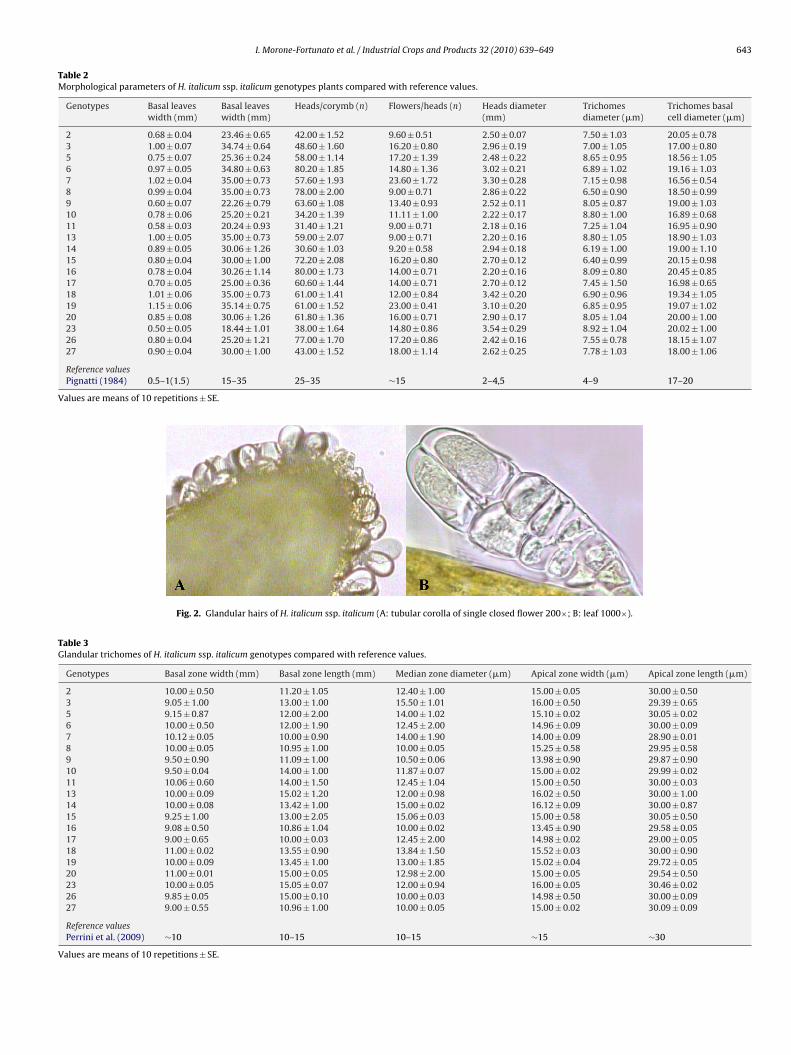
n the external bracts of the flower heads, while there is a very highensity of glands on each tubular flower (Fig. 2A).
.2. Histology
Histological studies of the flower heads, and the fresh leaves of. italicum ssp. italicum indicated that glandular trichomes secret-
ng essential oils are present in both organs and are of one singleorphological type and for all genotypes formed by 12 cells (Fig. 2A
nd B), divided into three zones (Table 3). These observations aren agreement with those previously reported for H. italicum ssp.
icrophyllum (Perrini et al., 2009b). The inspection of the secretorylands of H. italicum ssp. italicum genotypes was generally easiern the tubular corolla of single closed flowers than on leaves dueo the presence of thick hairs on leaves. Therefore, the counting ofhe gland number was not possible due to their very dense distri-ution on the flowers and to the presence of many non-glandularrichomes on the leaves and stems. The diameter of non-glandularrichomes and their basal cell were of 4–9 and 17–20 �m, respec-ively (Table 2) agreeing with the literature (Pignatti, 1984).
.3. Chemistry
The amount of essential oils recovered from the flower heads ofhe different genotypes of H. italicum ssp. italicum under investiga-ion was in general quite low, ranging from 0.1 to 0.2%. The highestroduction of essential oils was found in the two genotypes 5 (0.4%)
nd 16 (0.5%).The chemical profiling by means of GC and GC/MS analysesllowed to group the genotypes according to the major constituentsresent in their essential oils. Some of the genotypes (3, 5, 7 and 23)roduce an essential oils with a consistent or predominant amount
s and Products 32 (2010) 639–649
of monoterpenoids, in total ranging from 26.4% in genotype 23 to53.6% in genotype 5, with nerol and its esters, neryl acetate andneryl propanoate as the main constituents (Table 4).
The other genotypes are characterized by essential oils very richin sesquiterpenoids (from 61.6%, in genotype 15 to 91.3%, in geno-type 20). Among these, essential oils of genotypes 2, 6, 8, 9, 13,16, 18 and 20 can be distinguished by the high content of both�- and �-selinene accounting from 35.8% (genotype 13) to 64.2%(genotype 8) of the total. In contrast, some genotypes were char-acterized by a high amount of either �-selinene (10, 11 and 19) or�-selinene (14, 15), and in general those genotypes are peculiar inthat a decrease in the amount of selinene compounds is associatedwith a relative increase in the content of �-curcumene (Table 4).Genotype 17 appears unique in the collection in that produces32.3% of �-bisabolene together with �-curcumene (27.7%).
�-Diketones represent another chemical class present to adifferent extent in almost all genotypes, and were particu-larly abundant in genotypes 3, 5 and 7 (12.9, 24.2, and 11.2%respectively). Dominant �-diketones are 4, 6, 9-trimethyldec-8-en-3, 5-dione (5.5–11.0%) and 5, 7, 10-trimethylundec-9-en-4,6-dione (2.6–6.9%). The �-diketone 4, 6-dimethyloctan-3, 5-dione(0.2–2.4%) appears instead as a common constituent in almost allthe oils (Table 4).
Among the minor monoterpene and sesquiterpene con-stituents, carvacrol (1.6–14.8%), �-caryophyllene (1.2–18.6%) andar-curcumene (1.1–8.3%) were found in discrete and variableamounts in almost all the extracted essential oils (Table 4).
Finally, in genotypes 5 (2.2%), 8 (1.8%), 11 (3.8%), 15 (8.9%), 19(7.9%), 23 (2.1%) and 27 (5.2%), rosifoliol represents a character-istic component, not detected in the essential oils from the othergenotypes.
3.4. Genetic analysis
The AFLP analysis (Fig. 3) was conducted with three primer com-binations and revealed a total of 357 amplified DNA fragmentsranging from 80 to 800 bp (Table 5). A total of 195 polymor-phic bands were observed, but only 110 of these were consideredfor the cluster analysis, excluding the highest and lowest molec-ular weight bands that could create ambiguity in the analysis.The polymorphism percentage ranged from 26% (P-AGG/M-ACG)to 37% (P-AGC/M-ACT). Among the three tested primer combina-tions, just one (P-AGG/M-ACG) was sufficient to distinguish the 20genotypes.
The Diversity Index (DI) was calculated as: DI = 1 − Pi2, where Piis the phenotypic frequency for each assay unit (Russell et al., 1997).This index expresses the ability of a marker system to discrimi-nate between the samples used. It is equal to 0 for monomorphicmarkers and approximate to 1 for a heterogeneous population. Theefficiency of AFLPs in providing a large number of bands with justone electrophoretic analysis is reflected in the high value of themean of Diversity Index (92.1%), and in the number of separategenotypes per assay unit (each AFLPs primer combination) thatranged from 17 to 20.
Based on results from the 110 AFLP scored bands, a dendrogramrepresenting the genetic similarity of the 20 examined H. italicumssp. italicum genotypes was obtained (Fig. 4).
3.5. Micropropagation
In the current study, the micropropagation protocol used forH. italicum ssp. microphyllum (Perrini et al., 2009b) was appliedto all the H. italicum ssp. italicum genotypes providing successfulregeneration.
I. Morone-Fortunato et al. / Industrial Crops and Products 32 (2010) 639–649 643
Table 2Morphological parameters of H. italicum ssp. italicum genotypes plants compared with reference values.
Genotypes Basal leaveswidth (mm)
Basal leaveswidth (mm)
Heads/corymb (n) Flowers/heads (n) Heads diameter(mm)
Trichomesdiameter (�m)
Trichomes basalcell diameter (�m)
2 0.68 ± 0.04 23.46 ± 0.65 42.00 ± 1.52 9.60 ± 0.51 2.50 ± 0.07 7.50 ± 1.03 20.05 ± 0.783 1.00 ± 0.07 34.74 ± 0.64 48.60 ± 1.60 16.20 ± 0.80 2.96 ± 0.19 7.00 ± 1.05 17.00 ± 0.805 0.75 ± 0.07 25.36 ± 0.24 58.00 ± 1.14 17.20 ± 1.39 2.48 ± 0.22 8.65 ± 0.95 18.56 ± 1.056 0.97 ± 0.05 34.80 ± 0.63 80.20 ± 1.85 14.80 ± 1.36 3.02 ± 0.21 6.89 ± 1.02 19.16 ± 1.037 1.02 ± 0.04 35.00 ± 0.73 57.60 ± 1.93 23.60 ± 1.72 3.30 ± 0.28 7.15 ± 0.98 16.56 ± 0.548 0.99 ± 0.04 35.00 ± 0.73 78.00 ± 2.00 9.00 ± 0.71 2.86 ± 0.22 6.50 ± 0.90 18.50 ± 0.999 0.60 ± 0.07 22.26 ± 0.79 63.60 ± 1.08 13.40 ± 0.93 2.52 ± 0.11 8.05 ± 0.87 19.00 ± 1.0310 0.78 ± 0.06 25.20 ± 0.21 34.20 ± 1.39 11.11 ± 1.00 2.22 ± 0.17 8.80 ± 1.00 16.89 ± 0.6811 0.58 ± 0.03 20.24 ± 0.93 31.40 ± 1.21 9.00 ± 0.71 2.18 ± 0.16 7.25 ± 1.04 16.95 ± 0.9013 1.00 ± 0.05 35.00 ± 0.73 59.00 ± 2.07 9.00 ± 0.71 2.20 ± 0.16 8.80 ± 1.05 18.90 ± 1.0314 0.89 ± 0.05 30.06 ± 1.26 30.60 ± 1.03 9.20 ± 0.58 2.94 ± 0.18 6.19 ± 1.00 19.00 ± 1.1015 0.80 ± 0.04 30.00 ± 1.00 72.20 ± 2.08 16.20 ± 0.80 2.70 ± 0.12 6.40 ± 0.99 20.15 ± 0.9816 0.78 ± 0.04 30.26 ± 1.14 80.00 ± 1.73 14.00 ± 0.71 2.20 ± 0.16 8.09 ± 0.80 20.45 ± 0.8517 0.70 ± 0.05 25.00 ± 0.36 60.60 ± 1.44 14.00 ± 0.71 2.70 ± 0.12 7.45 ± 1.50 16.98 ± 0.6518 1.01 ± 0.06 35.00 ± 0.73 61.00 ± 1.41 12.00 ± 0.84 3.42 ± 0.20 6.90 ± 0.96 19.34 ± 1.0519 1.15 ± 0.06 35.14 ± 0.75 61.00 ± 1.52 23.00 ± 0.41 3.10 ± 0.20 6.85 ± 0.95 19.07 ± 1.0220 0.85 ± 0.08 30.06 ± 1.26 61.80 ± 1.36 16.00 ± 0.71 2.90 ± 0.17 8.05 ± 1.04 20.00 ± 1.0023 0.50 ± 0.05 18.44 ± 1.01 38.00 ± 1.64 14.80 ± 0.86 3.54 ± 0.29 8.92 ± 1.04 20.02 ± 1.0026 0.80 ± 0.04 25.20 ± 1.21 77.00 ± 1.70 17.20 ± 0.86 2.42 ± 0.16 7.55 ± 0.78 18.15 ± 1.0727 0.90 ± 0.04 30.00 ± 1.00 43.00 ± 1.52 18.00 ± 1.14 2.62 ± 0.25 7.78 ± 1.03 18.00 ± 1.06
Reference valuesPignatti (1984) 0.5–1(1.5) 15–35 25–35 ∼15 2–4,5 4–9 17–20
Values are means of 10 repetitions ± SE.
Fig. 2. Glandular hairs of H. italicum ssp. italicum (A: tubular corolla of single closed flower 200×; B: leaf 1000×).
Table 3Glandular trichomes of H. italicum ssp. italicum genotypes compared with reference values.
Genotypes Basal zone width (mm) Basal zone length (mm) Median zone diameter (�m) Apical zone width (�m) Apical zone length (�m)
2 10.00 ± 0.50 11.20 ± 1.05 12.40 ± 1.00 15.00 ± 0.05 30.00 ± 0.503 9.05 ± 1.00 13.00 ± 1.00 15.50 ± 1.01 16.00 ± 0.50 29.39 ± 0.655 9.15 ± 0.87 12.00 ± 2.00 14.00 ± 1.02 15.10 ± 0.02 30.05 ± 0.026 10.00 ± 0.50 12.00 ± 1.90 12.45 ± 2.00 14.96 ± 0.09 30.00 ± 0.097 10.12 ± 0.05 10.00 ± 0.90 14.00 ± 1.90 14.00 ± 0.09 28.90 ± 0.018 10.00 ± 0.05 10.95 ± 1.00 10.00 ± 0.05 15.25 ± 0.58 29.95 ± 0.589 9.50 ± 0.90 11.09 ± 1.00 10.50 ± 0.06 13.98 ± 0.90 29.87 ± 0.9010 9.50 ± 0.04 14.00 ± 1.00 11.87 ± 0.07 15.00 ± 0.02 29.99 ± 0.0211 10.06 ± 0.60 14.00 ± 1.50 12.45 ± 1.04 15.00 ± 0.50 30.00 ± 0.0313 10.00 ± 0.09 15.02 ± 1.20 12.00 ± 0.98 16.02 ± 0.50 30.00 ± 1.0014 10.00 ± 0.08 13.42 ± 1.00 15.00 ± 0.02 16.12 ± 0.09 30.00 ± 0.8715 9.25 ± 1.00 13.00 ± 2.05 15.06 ± 0.03 15.00 ± 0.58 30.05 ± 0.5016 9.08 ± 0.50 10.86 ± 1.04 10.00 ± 0.02 13.45 ± 0.90 29.58 ± 0.0517 9.00 ± 0.65 10.00 ± 0.03 12.45 ± 2.00 14.98 ± 0.02 29.00 ± 0.0518 11.00 ± 0.02 13.55 ± 0.90 13.84 ± 1.50 15.52 ± 0.03 30.00 ± 0.9019 10.00 ± 0.09 13.45 ± 1.00 13.00 ± 1.85 15.02 ± 0.04 29.72 ± 0.0520 11.00 ± 0.01 15.00 ± 0.05 12.98 ± 2.00 15.00 ± 0.05 29.54 ± 0.5023 10.00 ± 0.05 15.05 ± 0.07 12.00 ± 0.94 16.00 ± 0.05 30.46 ± 0.0226 9.85 ± 0.05 15.00 ± 0.10 10.00 ± 0.03 14.98 ± 0.50 30.00 ± 0.0927 9.00 ± 0.55 10.96 ± 1.00 10.00 ± 0.05 15.00 ± 0.02 30.09 ± 0.09
V
Reference valuesPerrini et al. (2009) ∼10 10–15 10–15
alues are means of 10 repetitions ± SE.
∼15 ∼30

644I.M
orone-Fortunatoet
al./IndustrialCropsand
Products32 (2010) 639–649
Table 4Composition of the essential oils from H. italicum ssp. italicum genotypes.
Compounds aAI % Identification
2 3 5 6 7 8 9 10 11 13 14 15 16 17 18 19 20 23 26 27 GC/MS
3,4-Hexanedione 800 – – – 6.8 11.3 – – – – – – – 2.6 – – – – – – – GC/MSTricyclene 921 – – – 0.7 – – – – – – – – – – – – – – – – GC/MSArtemisiatriene 923 – – – 2.1 – – – – – – – – – – – – – – – – GC, GC/MS�-Pinene 930 0.9 3.6 – – – – – – – – 1.2 tr – – 0.8 – 0.5 – 1.1 – GC, GC/MS3-Octanone 979 – – – 0.7 2.8 – – – – – – – – – – – – – – – GC, GC/MSp-Cymene 1018 tr 1.2 0.5 0.5 1.7 tr tr – – 0.6 0.4 0.8 tr 0.5 0.5 1.8 tr – 1.6 0.4 GC, GC/MS1,8- Cineole 1024 – – – – – – – – – – – 3.3 – – – 4.1 – – – 0.3 GC, GC/MS�- Terpinene 1055 – – – – – – – – – – – 0.3 – – – 0.7 – – – 0.7 GC, GC/MS2,4-Dimethyleptan-3,5-dione 1070 – – 0.5 tr tr – – – – – – – – – – – – – – – GC/MSLinalool 1095 1.5 0.4 3.9 0.9 0.5 tr 1.0 – 1.4 0.4 tr 1.5 0.5 – 1.5 1.9 – 5.1 1.2 – GC, GC/MSTagetone 1140 – 0.4 – 0.4 – tr 0.9 – 0.3 – 0.4 – 0.4 0.4 0.4 – – – – – GC, GC/MSTerpinen-4-ol 1174 – – tr – 0.2 – – – – – – 1.8 – – – 2.2 – 1.2 – 4.2 GC, GC/MS4,6-Dimethyloctan-3,5-dione 1180 0.9 0.8 2. 4 1.2 0.6 0.7 0.9 1.5 0.8 – – 0.2 0.3 0.2 1.2 – – 0.7 1.6 0.9 GC/MS�-Terpineol 1186 0.8 0.8 1.0 0.5 2.3 tr 1.5 1.0 0.8 0.9 0.5 2.6 0.4 1.0 0.9 2.9 – 1.2 0.4 0.6 GC, GC/MSNerol 1126 0.5 2.4 3.9 0.6 18.8 1.5 1.7 0.8 0.5 0.9 1.0 2.0 1.3 0.9 2.7 1.9 0.4 1.7 0.5 0.4 GC, GC/MSLavandulyl acetate 1290 0.7 0.7 0.9 – 0.4 – 1.0 – – – – 0.2 tr – 0.8 – – 0.5 – – GC, GC/MSCarvacrol 1300 3.8 6.2 1.9 8.4 9.8 6.7 6.8 14.8 8.7 3.8 1.6 9.5 7.4 9.8 5.4 2.6 5.8 3.3 3.2 3.6 GC, GC/MS�-Terpinylacetate 1346 – – 0.4 tr – – – – – – – – – – – – – – – – GC, GC/MSNeryl acetate 1360 1.7 15.1 32.0 0.4 8.5 0.7 3.2 1.0 1.1 0.8 1.8 4.5 1.4 1.0 2.8 4.7 1.4 11.4 8.1 1.2 GC, GC/MS�-Ylangene 1375 – 3.3 – – 1.3 – – – 0.4 0.2 4.9 – – – 0.5 – – – 2.1 0.7 GC/MSiso-Italicene 1403 – – 0.4 tr 0.5 – – – – – – 0.6 tr 0.7 – 1.2 0.4 – – – GC/MSItalicene 1407 0.7 2.7 0.9 1.4 – – 1.7 0.8 1.3 0.4 1.1 1.9 0.6 2.5 0.7 1.9 1.5 – 6.5 1.6 GC/MS�-cis-Bergamotene 1415 – 2.6 0.6 – – – – – – 0.7 – 1.0 – – – 1.3 – – – – GC/MS�-Caryophyllene 1420 5.1 1.2 – 3.0 2.7 tr – 10.1 12.7 6.2 8.6 6.5 14.4 10.9 4.5 7.3 18.6 7.8 2.6 3.0 GC, GC/MS�-trans-Bergamotene 1430 – 2.9 0.3 – – – – – – 1.1 – 1.1 – 0.5 – 1.4 – – – – GC/MS4,6,9-Trimethyldec-8-en-3,5-dione 1438 – 5.5 11.0 – 6.2 – – – – – – – – – tr – – – – – GC/MS�-Humulene 1452 – 0.7 – tr – – 1.2 0.9 0.7 0.7 0.3 0.6 0.7 0.3 – 0.7 0.6 3.2 1.7 0.8 GC, GC/MSNeryl propanoate 1452 0.7 0.5 3.6 tr 6.8 – – – 2.3 2.7 – 0.2 tr – 1.5 0.2 – 0.9 – – GC, GC/MS�-Acoradiene 1464 – 0.3 – tr – – – 6.7 0.2 – – 0.2 – – – – – – – – GC/MS�-Acoradiene 1470 – 0.6 0.3 tr – – – – – – – 0.2 – – – – – – – – GC/MS4,5- di-epi-Aristolochene 1471 0.8 – – 0.7 – 0.7 – – 0.2 0.4 0.3 0.3 – – 0.7 0.3 0.4 0.5 5.1 – GC/MSSelinen-4,11-diene 1478 8.1 – – 6.0 – 7.4 4.8 3.3 3.1 4.2 7.1 2.9 5.1 1.2 6.9 3.3 6.0 1.4 – 2.0 GC/MS�-Curcumene 1479 4.2 23.3 5.0 2.3 – – 6.5 3.3 8.4 1.6 15.0 14.3 3.4 27.7 – 17.1 14.7 7.7 41.0 14.5 GC/MSar-Curcumene 1480 – 4.9 6.4 8.3 6.4 – 2.9 1.1 3.1 – – – – 2.7 – – – 6.1 – 2.2 GC/MS�-Amorphene 1483 – – – – 3.2 – – – – – – – – – 4.0 – – – – – GC/MS2,4,6,9-Tetramethylundec-8-en-3,5-dione 1486 – – – – 1.8 – – – – – – – – – – – – – – – GC/MS5,7,10-Trimethylundec-9-en-4,6-dione 1487 – 6.1 6.9 – 2.6 – – – – – – – – – – – – – – – GC/MS�-Selinene 1489 33.3 3.6 – 19.7 – 38.0 25.7 15.9 11.6 20.0 2.8 3.8 24.5 2.3 28.9 15.3 26.7 6.1 22.0 4.7 GC/MS�-Selinene 1498 26.5 – – 22.7 – 26.2 14.7 9.1 9.5 15.8 19.3 13.2 19.4 3.6 22.2 9.5 18.8 0.8 – 4.9 GC/MS�-Muurolene 1500 – 2.9 – – – – – 1.2 – 2.0 19.8 – – – – – – – – 0.7 GC/MS�-Bisabolene 1505 – 1.1 – – – – – – – – – – – 32.3 – – – – – – GC/MS7-epi-�-Selinene 1525 1.0 – 0.6 0.4 – 0.6 – – – – 0.6 0.3 0.3 – – – – – – – GC/MS�-Amorphene 1513 – – – – 1.5 – – – 1.6 0.5 – – – – – – – 0.6 – 2.7 GC/MS�-Cadinene 1530 1.0 1.7 – – 0.6 – 2.6 1.3 1.4 2.5 – – – 0.8 – – 1.3 – 2.0 GC, GC/MS�-Cadinene 1540 – 0.3 – – 0.2 – – – 0.2 0.4 0.3 – – – – – – – – – GC/MS�-Calacorene 1544 – 0.7 – – 0.4 – – – – – – – – – 0.8 – – – – – GC/MSNerolidol 1561 – 1.3 0.3 – 0.3 – – – – – – 0.2 – – 1.5 – – – – – GC, GC/MSGeranyl butanoate 1562 – 0.5 – – – – – – – – 2.0 – – – – – – – – – GC, GC/MS3,5,7,10-Tetramethylundec-9-en-4,6-dione 1570 – 0.5 3.4 – – – – – – – – – – – – – – – – – GC/MSCaryophyllene alchool 1573 – – – 0.6 – – – – – – – 0.4 1.1 – – – – – – – GC/MS�-Caryophyllene oxide 1581 0.6 – – 6.9 0.8 4.3 2.5 1.1 1.5 2.0 – – 14.9 0.7 1.0 – 1.3 1.1 0.3 – GC, GC/MSNeryl isovalerate 1582 0.5 0.6 2.5 – 1.6 – 1.1 0.8 0.4 – 0.8 – – 0.5 – – 1.1 – – GC/MS

I. Morone-Fortunato et al. / Industrial Crops and Products 32 (2010) 639–649 645
Tabl
e4
(Con
tinu
ed)
Com
pou
nd
saA
I%
Iden
tifi
cati
on
23
56
78
910
1113
1415
1617
1819
2023
2627
GC
/MS
Gu
aiol
1594
––
1.7
–0.
40.
7–
1.1
2.1
1.0
2.2
––
––
0.7
–1.
8–
3.3
GC
,GC
/MS
Ros
ifol
iol
1606
––
2.2
––
1.8
––
3.8
–8.
9–
––
7.9
–2.
10.
35.
2G
C/M
SN
eril
tigl
ate
1615
––
––
3.0
––
––
––
––
––
––
––
–G
C/M
SC
ube
nol
-1,1
0d
iep
i16
20–
––
–1.
8–
––
––
––
––
6.8
0.6
––
––
GC
/MS
�-E
ud
esm
ol16
306.
7–
1.0
––
––
–11
.19.
73.
00.
4–
0.5
–0.
42.
38.
6–
20.4
GC
/MS
�-E
ud
esm
ol16
48–
–1.
6–
–1.
1–
13.7
1.6
2.9
–1.
5–
––
1.5
–2.
4–
2.3
GC
,GC
/MS
�-E
ud
esm
ol16
52–
–1.
5–
–0.
6–
–2.
21.
0–
3.3
––
–3.
1–
2.8
–4.
1G
C/M
SSe
lin
-11-
en-4
-�-o
l16
68–
––
3.7
–7.
02.
76.
14.
7–
––
1.0
––
––
––
–G
C/M
S7(
11)-
en-4
-Eu
des
mol
1685
––
2.4
––
––
––
––
––
––
––
––
–G
C/M
SG
uai
olac
etat
e17
27–
––
––
––
2.0
––
––
––
––
––
––
GC
,GC
/MS
Yie
ld%
100
98.7
100
98.9
99.0
98.0
80.8
98.9
97.6
82.3
96.7
89.3
99.7
99.7
98.3
96.5
99.4
81.4
99.3
87.4
aA
rith
met
icin
dex
es(A
I)re
lati
veto
an-
alka
ne
seri
es(C
7–C
27)
calc
ula
ted
ona
DB
-5co
lum
n.
Fig. 3. AFLP electrophoretic patterns of 20 H. italicum genotypes (2-27) obtainedwith the primer combination Pst-AGG/Mse-ACG.
3.5.1. Establishment and multiplication stageThe ANOVA analyses of percentage of proliferation, number of
shoots/explant, and growth showed significant statistical differ-ences among the investigated genotypes with the results reportedusing the SNK test (Table 6). More than half of the H. italicum ssp.italicum genotypes responded to hormonal stimulation with prolif-eration values greater than 50% (even 86–100% for some genotypes)(Table 6). Only four genotypes (7, 9, 11, and 13) showed a lowerproliferation percentage from 33 to 47% (Table 6).
The number of shoots/explant varied from a maximum valueof 6.53 (genotype 16) to a minimum of 1.47 (genotype 13). Allthe neo-formed shoots always developed from axillary buds, neverfrom basal calli. The elongation of the shoots was also variable,ranging from 0.77 to 2.87 cm (for genotypes 6 and 3, respectively),with no relationship between the number of shoots and theirelongation.
3.5.2. Root formation stageAnalysis of variance (ANOVA) of the rooted shoots percentage
and number of new roots showed highly significant genotypeseffect with the results of the SNK test presented in Table 7.
All the genotypes responded to rooting with values ranging from50 to 100% of the shoots with roots. The only exception was geno-type 17, which naturally showed a lower rate of rooting (16.67%),further decreasing with the addition of IBA (8.33%). Indeed, the cul-ture medium enriched with IBA, increased rooting percentages inonly three genotypes (2, 10, and 19).
The different response of the genotypes may be referred to threedifferent conditions of endogenous hormones. The first, coveringthe majority of our genotypes, characterized by an optimal hor-monal balance, so that the addition of an exogenous hormone,competitively, determines a decrease of rooting. In the second con-

646 I. Morone-Fortunato et al. / Industrial Crops and Products 32 (2010) 639–649
Table 5Characteristics of the banding patterns of the 3 AFLP primer combinations used among the 20 H. italicum ssp. italicum genotypes examined.
Primer combination Total bands (N) Polymorphic bands (N) Scored bands (N) Polymorphism (%) Genotypes per assay unit (N) Diversity index (%)
dcfist
TPa
CMl
e
Fw
Pst-AGG/Mse-ACG 143 70 37Pst-AGC/Mse-AGT 120 70 38Pst-AGC/Mse-ACT 94 55 35
ition, involving only three genotypes, an increase of the hormoneoncentration increases the percentage of rooting. Finally, the lastve genotypes, which do not respond to exogenous hormones pos-ibly have an optimal quantity of endogenous hormones, so thathey show 100% rooting and the further supply of growth regulators
able 6roliferation, shoot number and shoot length of H. italicum ssp. italicum genotypes,fter 30 days of in vitro culture.
Genotypes Proliferationa (%) Shoots/explantb (n) Shoot lengthb (cm)
2 93.35ab 5.67ab 2.54ab3 73.30abcde 2.93cdef 2.87a5 53.34defg 4.67abed 2.50abc6 66.67bcdef 2.80cdef 0.77i7 40.00fg 1.47f 1.27efghi8 93.33ab 3.27cdef 1.97bed9 46.66efg 1.87ef 1.69cdefg
10 60.00cdefg 1.93ef 1.69cdefg11 33.30g 1.73ef 0.94ghi13 33.35g 1.47f 0.95ghi14 86.67abc 4.20bede 1.50defgh15 100.00a 5.13abc 1.90bede16 100.00a 6.53a 1.97bed17 66.67bcdef 2.60def 1.20fghi18 100.00a 3.00cdef 1.90bede19 80.00abed 2.80cdef 2.00bed20 53.34defg 2.47def 1.44defghi23 53.34defg 1.80ef 0.88hi26 100.00a 3.40cdef 0.87hi27 100.00a 3.27cdef 1.75cdef
ulture medium: BM + BAP 1 mg l−1 + IBA 0.2 mg l−1 + sucrose 20 g l−1.eans followed by the same letters are not significantly different at the P ≤ 0.01
evel (SNK’ test).a Percentage (%) is mean of 3 experiments. Each experiment comprises 30 primary
xplants per genotype.b Values are means of 50 repetitions. Each explant was a repetition.
ig. 4. UPGMA dendrogram of 20 H. italicum genotypes obtained with data coming from thhile letters “a” and “b” are referred to sub-groups of cluster 1.
26 20 96.332 17 88.237 19 95.2
does not influence the endogenous hormonal balance determininga decrease of rooting.
The number of roots obtained, although variable, allowed thegrowth of plantlets suitable for acclimatization stage of H. italicumssp. italicum.
3.5.3. Acclimatization stageThe response to acclimatization of the 20 genotypes under study
is described in Fig. 5. Plants growth was always satisfactory show-ing very high percentages (80% and 100%) of plants acclimatizedregardless of the use of Jiffy-Pots or conventional pots.
3.6. Cytology
Caryological analysis confirmed that micropropagated clones ofH. italicum ssp. italicum had a chromosome number of 2n = 28, thesame as the mother plant, agreeing with a report by Bacchetta etal. (2003) for the genus Helichrysum.
4. Discussion
The histological analysis showed the presence of glandular tri-chomes on flowers, leaves and young stems. The glands of allH. italicum ssp. italicum genotypes were consistent with thosereported by Perrini et al. (2009b) for the H. italicum ssp. micro-phyllum, both in structure and size, but, they differ from the flatglandular hairs (glands) of Arctium lappa, otherwise considered
representative of the Asteraceae family (Fahn, 1979). Observationsmade on leaves sections of in vitro propagated plants during themultiplication stage, showed the presence of glandular hairs mor-phologically identical to those observed on fresh plant materialfrom the field. Density of the glandular hairs is very high on theree AFLP primer combinations. Brackets on the right indicate the two main clusters,
I. Morone-Fortunato et al. / Industrial Crops and Products 32 (2010) 639–649 647
Table 7Root formation and root number of H. italicum ssp. italicum genotypes, after 15 days of in vitro culture.
Genotypes Root formation (%) Roots (n)
Hormone free IBA (0.2 mg l−1) Hormone free IBA (0.2 mg l−1)
2 66.67bc 91.67a 6.00bc 5.82bc3 100.00a 91.68a 8.92bc 8.82bc5 100.00a 91.69a 11.17bc 16.00b6 100.00a 83.33ab 8.33bc 14.90b7 100.00a 25.00e 7.25bc 3.67bc8 100.00a 83.33ab 10.75bc 13.00bc9 100.00a 91.67a 10.50bc 10.90bc
10 83.33ab 91.68a 8.80bc 9.36bc11 100.00a 100.00a 9.42bc 12.67bc13 100.00a 100.00a 8.00bc 13.00bc14 100.00a 100.00a 10.25bc 13.00bc15 58.33cd 41.67d 5.71bc 7.00bc16 100.00a 50.00a 30.42a 26.83a17 16.67ef 8.33f 2.00bc 4.00bc18 66.67bc 50.00cd 5.25bc 3.33bc19 83.33ab 100.00a 7.90bc 8.75bc20 100.00a 100.00a 7.17bc 7.50bc23 50,00cd 41.67d 1.60c 4.00bc
V by the
tmsm2
Hit2ip�e(aoa
ibop
26 91.67a 83.33ab27 100.00a 100.00a
alues are means of 50 repetitions. Each explant was a repetition. Means followed
ubular corolla of each flower, while their density decreases veryuch on the bracts covering the capitula, in contrast to H. italicum
sp. microphyllum where secretory glands were observed with aore or less constant density on all plant organs (Perrini et al.,
009b).Essential oils produced by the secretory glands of different
elichrysum species have been studied and the analysis of H.talicum ssp. italicum showed essentially three different chemo-ypes for the essential oils (Bianchini et al., 2001; Paolini et al.,006; Satta et al., 1999; Tucker et al., 1997). Essential oils from H.
talicum ssp. italicum characterized by high amounts of monoter-enoid constituents such as neryl acetate, neryl propanoate and-pinene have been described by Bianchini et al. (2001) and Paolinit al. (2006). A typical oil rich in geraniol (36%) and geranyl acetate15%) has been reported for plants collected in Greece (Bianchini etl., 2001; Chinou et al., 1996). In contrast, other investigations rec-gnized plant populations of H. italicum ssp. italicum with dominantmounts of sesquiterpenes (Bianchini et al., 2001).
Despite the fact that fresh or dry flower heads of H. italicum arencluded in several European Pharmacopoeias for official use, to theest of our knowledge, previous chemical studies on the essentialils from this species have been substantially carried out on thelant aerial parts, including flowers, leaves and stems (Charles and
Fig. 5. In vitro establishment of H. italicum ssp. italicum
8.64bc 10.80bc12.17bc 12.42bc
same letters are not significantly different at the P ≤ 0.01 level (SNK’ test).
Simon, 1991; Bianchini et al., 2001; Paolini et al., 2006; Satta etal., 1999; Tucker et al., 1997). Data reported here are the chemi-cal characterization of the essential oils produced only by the plantflowers, which are especially rich in glandular structures as con-firmed by our histological observations (Fig. 2). This work followsour recent investigation (Perrini et al., 2009b) for the chemical char-acterization of the essential oil from flower heads of H. italicum ssp.microphyllum.
According to the results reported in Table 4, genotypes of H.italicum ssp. italicum from different locations in Italy represent dif-ferent chemotypes confirming the compositional variability of theessential oils of this species in relation to its geographical originand vegetation cycle (Charles and Simon, 1991; Lawrence, 1998;Bianchini et al., 2001; Angioni et al., 2003; Paolini et al., 2006). Atleast three chemical compositional profiles have been evidencedfrom this study: (a) genotypes rich in nerol and its esters (I); (b)genotypes with a dominance of �- and �-selinene (II); (c) geno-types with high amounts of �-curcumene (III). Chemical profile I of
H. italicum ssp. italicum plants resembles that previously reportedfor the essential oils from the aerial parts of plants collected inother locations in Italy or in Corsica, with the difference that, inthose studies, nerol and its esters were found to be often asso-ciated with good quantities of �-curcumene (Charles and Simon,after 30 days from transplant in the greenhouse.

6 al Crop
1P
piaaaqdtp
titc(Lebc
eMoswaps
iaiesw
adtlgmafa
1t(tAlioi12a
lsit
48 I. Morone-Fortunato et al. / Industri
991; Lawrence, 1998; Bianchini et al., 2001; Angioni et al., 2003;aolini et al., 2006).
Nerol and its esters have also been found as characteristic com-onents of the essential oil of flowers from some genotypes of H.
talicum ssp. microphyllum (Satta et al., 1999; Perrini et al., 2009b)veraging 40–50% of the total oil components. Based on the avail-ble chemical data in the literature (Bianchini et al., 2001; Satta etl., 1999; Paolini et al., 2006; Perrini et al., 2009), identification anduantification of the botanical origin of the essential oils or theirerived products from one or the other subspecies based only onhe chemistry might not be appropriate in the case of Helichrysumopulations sharing chemotype I.
Chemotypes II and III appear new for H. italicum ssp. italicum. Thewo sesquiterpenes �- and �-selinene (chemotype II) have not beendentified previously as typical dominant constituents of the essen-ial oils from this subspecies. �-Curcumene, identifies genotypes ofhemotype III, and is associated with high amounts of �-selineneTable 4). In contrast to previous reports (Charles and Simon, 1991;awrence, 1998; Bianchini et al., 2001; Angioni et al., 2003; Paolinit al., 2006), the content of nerol and its esters in genotypes ofoth chemotypes II and III, compared to genotypes belonging tohemotype I, appears instead greatly reduced (Table 4).
Within the subspecies of H. italicum, �-diketones are consid-red especially characteristic of H. italicum ssp. italicum (Tira and Diodica, 1967; Manitto et al., 1972). This is also confirmed by results
f the current study (Table 4) and previous findings on H. italicumsp. microphyllum (Perrini et al., 2009b). Interestingly, genotypesith the highest content and the larger number of �-diketones
re those belonging to chemotype I. In contrast, �-diketones areresent in very low amounts in those genotypes accumulatingesquiterpenes in their oils (Table 4).
Presence of the epi-eudesmol rosifoliol is reported as variablen H. italicum ssp. michrophyllum (Satta et al., 1999; Bianchini etl., 2001; Perrini et al., 2009b), but has never been detected in H.talicum ssp. italicum, probably due to a misidentification with otherudesmol compounds. Discrete amounts of rosifoliol were found inome of the genotypes (5, 8, 11, 15, 19, 23 and 27), always associatedith other eudesmol derivatives (Table 4).
The finding of different chemotypes of H. italicum ssp. italicums described here should also be regarded as interesting for aiversified industrial utilization of the plant, beside its applica-ion in phytotherapy. Nerol and its derivatives, for example, areargely employed as cosmetic ingredient for their sweet rose fra-rance, while curcumene derivatives or drugs containing them areainly used in the food industry (Ansari and Curtis, 1974). Alfa-
nd �-selinene have, respectively, a sweet woody and herbaceousragrance and they have acquired importance in chemical ecologys pheromones (Harborne, 1993; Breitmaier, 2006).
All 20 genotypes subjected to genetic characterization with10 AFLP polymorphic bands disclosed a clear genetic distinc-ion, besides their chemical diversity. The phenetic dendrogramFig. 4) showed a clustering among the 20 genotypes that reflectshe chemical characterization suggesting that the analysis usingFLP markers might provide the evidence for some genomic regions
inked to loci involved in the expression of genes coding for chem-cal compounds that were identified in the investigated essentialils. In the literature are present several works regarding the chem-cal analysis and antimicrobial activity of Helichrysum (Satta et al.,999; Barberán et al., 1990; Süzgeca et al., 2005; Perrini et al.,009b), but very few that combine the phenotype and genotypenalysis (Scialabba et al., 2008; Perrini et al., 2009a).
The dendrogram includes three main clades, two of them areinked at a genetic distance of 0.50 in one main branch (cluster 1hared in “a” and “b”), while the other one appears clearly separatedn the phenetic tree at the same distance (cluster 2). Furthermore,he main branch, indicated with the number “1” is divided up into
s and Products 32 (2010) 639–649
two minor clades: one including genotypes 2, 5, 6, 3, 7, and 8; theother including genotypes 9, 10, 13, 16, 19, 20, 11, 18, and 27. Thesecond cluster labelled as “2” includes the remaining genotypes14-15-23-17-26. The mean genetic similarity among the genotypeswas 0.75. No ambiguous cases of homonymy were found.
As described above, the chemical analysis has differentiated theessential oils from the 20 genotypes of H. italicun ssp. italicum intothree principal chemotypes, that are also reflected in the geneticstudy. Among genotypes classified as extremely rich in nerol andits esters (Table 4), the genotypes 3, 5 and 7 are grouped togetherin the first subgroup of cluster “1”, while genotype 23 is separatedat a distance of 0.58 and located in cluster “2”. In agreement withthe chemical analysis, genotypes characterized by a prevalence ofboth �- and �- selinene are all grouped in the cluster number “1”.Very interestingly, genotypes distinguished by a high amount ofeither �-selinene or �-selinene appear separated in the cluster,the first group of genotypes (10, 11, 19) included in one of thebranches inside clade “1”, the others (14, 15) in clade “2”. More-over, the unique genotype 17, with a high amount of �-bisabolenetogether with �-curcumene, forms an independent branch insidegroup “2”. Similarly, genotype 26 characterized by a chemical pro-file of the essential oils, difficult to include in the above definedchemotypes, forms another independent branch inside clade “2”,close to genotype 17 at a distance of 0.58.
Finally, data described in this paper show that the micro-propagation protocol previously established for H. italicum ssp.microphyllum (Perrini et al., 2009b) is also suitable for the micro-propagation of H. italicum ssp. italicum. The growth conditionsadopted in this study allowed us to obtain in vitro clones of H.italicum ssp. italicum corresponding to the mother plant in termsof histological, caryological and morphological characteristics.Adaptability of the 20 genotypes, although always positive, pro-duced some variable responses in the in vitro growing conditions(Tables 6 and 7), likely determined by genetic differences betweenthe genotypes (Hartmann et al., 2002; Pecaut and Martin, 1992).
5. Conclusion
Our observations demonstrate that even if the genetic char-acters influence the essential oil composition, they are not adeterminant in changing the phenotypic traits. In this work, an effi-cient micropropagation protocol was established for H. italicum ssp.italicum. The protocol assures that the genetic clones are identicalto the mother plant (a selected genotype).
The constitution of micropropagated clones ensures uniformityof the regenerated material and the essential oils yield and quality(Noguiera and Romano, 2002; Kalemba and Thiem, 2002; Bertoli etal., 2004; Perrini et al., 2009b; Avato et al., 2005; Morone-Fortunatoand Avato, 2008) and allows the possibility to use H. italicum ssp.italicum in the industrial sector.
References
Adams, R.P., 2007. Identification of Essential Oil Components by Gas Chromatogra-phy/Mass Spectrometry. Allured Publishing Corporation.
Angioni, A., Barra, A., Arlorio, M., Coisson, J.D., Russo, M.T., Pirisi, F.M., Satta, M.,Cabras, P., 2003. Chemical composition, plant genetic differences, and antifungalactivity of the essential oil of Helichrysum italicum G. Don ssp. michrophyllum(Willd) Nym. J. Agric. Food Chem. 51, 1030, 1034.
Ansari, H.R., Curtis, A.J., 1974. Sesquiterpenes in the perfumery industry. J. Soc.Cosmet. Chem. 25, 203–231.
Appendino, G., Ottino, M., Marquez, N., Bianchi, F., Giana, A., Ballero, M., Sterner,O., Fiebich, B.L., Munoz, E., 2007. Arzanol, an anti-inflammatory and anti-HIV-
1 phloroglucinol �-pyrone from Helichrysum italicum ssp. microphyllum. J. Nat.Prod. 70, 608–612.Avato, P., Morone-Fortunato, I., Ruta, C., D’Elia, R., 2005. Glandular hairs and essentialoils in micropropagated plants of Salvia officinalis L. Plant Sci. 169, 29–36.
Bacchetta, G., Brullo, S., Mossa, L., 2003. Note tassonomiche sul genere HelichrysumMiller (Asteraceae) in Sardegna. Informatore Botanico Italiano 35 (1), 217–225.

al Crop
B
B
B
B
B
C
C
C
FG
HHH
J
J
K
LM
M
M
M
N
N
N
N
Voinchet, V., Giraud-Robert, A.M., 2007. Utilisation de l’huile essentielle d’helichryseitalienne et de l’huile végétale de rose musquée après intervention de chirurgie
I. Morone-Fortunato et al. / Industri
arberán, F.T., Sanmartín, E.I., Lorente, F.T., Rumberoa, A., 1990. Antimicrobial phe-nolic compounds from three Spanish Helichrysum species. Phytochemistry 29(4), 1093–1095.
ianchini, A., Tomi, P., Costa, J., Bernardini, A.F., 2001. Composition of Helichrysumitalicum G. Don fil. subsp. italicum essential oils from Corsica (France). Flav. Fragr.J. 16, 30–34.
ertoli, A., Morelli, I., Fraternale, D., Giampieri, L., Ricci, D., 2004. Volatile costituentof micropropagated plants of Bupleurum fruticosum L. Plant Sci. 167, 807–810.
lanco, A., Simeone, R., Resta, P., De Pace, C., Delre, V., Caccia, R., Scarascia Mug-nozza, G.T., Frediani, M., Cremonini, R., Cionini, P.G., 1996. Genomic relationshipsbetween Dasypyrum villosum (L.) Candargy and D. hordeaceum (Cosson etDurieu) Candargy. Genome 39, 83–92.
reitmaier, E., 2006. Terpenes: Flavors, Fragrances, Pharmaca, Pheromones. Wiley-VCH, London.
harles, D.J., Simon, J.E., 1991. Volatile compounds of the curry plant. Hort. Sci. 26,69–70.
hinou, I.B., Roussis, V., Perdetzolou, D., Loukis, A., 1996. Chemical and biologicalstudies on two Helichrysum species of greek origin. Planta Med. 62, pp. 377–339.
hinou, I.B., Roussis, V., Perdetzolou, D., Tzakou, O., Loukis, A., 1997. Chemical andantibacterial studies of two Helichrysum species of Greek origin. Planta Med. 63,181–183.
ahn, A., 1979. Secretory Tissues in Plants. Academic Press, London.omori, G., 1952. Microscopic Histochemistry Principles and Practice. University of
Chicago Press, Chicago.arborne, J.B., Turner, B.L., 1984. Plant Chemosystematics. Academic Press, London.arborne, J.B., 1993. Introduction to Ecological Chemistry. Elsevier Ltd., London.artmann, H.T., Kester, D.E., Davies F.T.Jr., Geneve, R.L., 2002. Hartmann and Kester’s
Plant Propagation: Principles and Practices, 7th ed. Prentice-Hall, Inc., UpperSaddle River, NJ, p. 644.
ahn, R., Schönfelder, P., 1995. Exkursionflora fur Kreta. Verlag Eugen Ulmer,Stuttgart, Germany.
oulain, D., König, A., 1998. The Atlas of Spectral Data of Sesquiterpenes Hydrocar-bons. E.B. Verlag, Hamburg.
alemba, D., Thiem, B., 2002. Costituents of essential oils of four micropropagatedSolidago species. Flav. Fragr. J. 19, 40–43.
awrence, B.M., 1998. Helichrysum oil and extract. Perf. Flav. 23, 55–59.anitto, P., Monti, D., Colombo, E., 1972. Two new �-diketones from Helichrysum
italicum. Phytochemistry 11, 2112–2114.ontemurro, C., Pasqualone, A., Simeone, R., Sabetta, W., Blanco, A., 2008. AFLP
molecular markers to identify virgin olive oils from single Italian cultivars. Eur.Res. Food Technol. 226, 1439–1444.
ontemurro, C., Simeone, R., Pasqualone, A., Ferrara, E., Blanco, A., 2005. Geneticrelationships and cultivar identification among sixty olive accessions using AFLPand SSR markers. J. Hortic. Sci. Biotechnol. 80, 105–110.
orone-Fortunato, I., Avato, P., 2008. Plant development and synthesis of essentialoils in micropropagated and mycorrhiza inoculated plants of Origanum vulgareL. ssp. hirtum (Link) Ietswaart. Plant Cell Tiss. Org. Cult. 93, 139–149.
oguiera, J.M.F., Romano, A., 2002. Essential oils form micropropagated plants ofLavandula viridis. Phytochem. Anal. 13, 4–7.
ostro, A., Bisignano, G., Cannatelli, M.A., Crisafi, G., Germano, M.P., Alonzo, V.,2001. Effects of Helichrysum italicum extract on growth and enzymatic activityof Staphylococcus aureus. Int. J. Antimicrob. Agents 17, 517–520.
ostro, A., Cannatelli, M.A., Crisafi, G., Musolino, A.D., Procopio, F., Alonzo, V., 2004.
Modification of hydrophobicity, in vitro adherence and cellular aggregation ofStreptococcus mutans by Helichrysum italicum extract. Lett. Appl. Microbiol. 38,423–427.ostro, A., Cannatelli, M.A., Marino, A., Picerno, I., Pizzimenti, F.C., Scoglio, M.E.,Spataro, P., 2003. Evaluation of antiherpesvirus,1 and genotoxic activities ofHelichrysum italicum extract. New Microbiol. 26 (1), 125–128.
s and Products 32 (2010) 639–649 649
Nostro, A., Cannatelli, M.A., Musolino, A.D., Procopio, F., Alonzo, V., 2002. Helichrysumitalicum extract interferes with the production of enterotoxins by Staphylococcusaureus. Lett. Appl. Microbiol. 35, 181–184.
Paolini, J., Desjober, J.M., Costa, J., Bernardini, A.F., Buti Castellini, C., Cioni, P.L.,Flamini, G., Morelli, I., 2006. Composition of essential oils of Helichrysum italicumG. Don fil. subsp. italicum from Tuscan Achipelago islands. Flav. Fragr. J. 21,805–808.
Pecaut, P., Martin, F., 1992. Non-conformity of in vitro propagated plants of earlyMediterranean varieties of globe artichoke (Cynara scolymus L.). Acta Hortic.300, 363–366.
Perrini, R., Alba, V., Ruta, C., Morone-Fortunato, I., Blanco, A., Montemurro, C., 2009a.An evaluation of a new approach to the regeneration of Helichrysum italicum(Roth) G. Don, and the molecular characterization of the variation among setsof differently derived regenerants. Cell. Mol Biol. Lett. 14, 377–394.
Perrini, R., Morone-Fortunato, I., Lorusso, E., Avato, P., 2009b. Glands, essential oilsand in vitro establishment of Helichrysum italicum (Roth) G. Don ssp. italicum.Ind. Crops Prod. 29, 395–403.
Pignatti, S., 1984. Flora d’Italia. Edizione Edagricole, Bologna.Russell, J.R., Fuller, J.D., Macaulay, M., Hatz, B.G., Jahoor, A., Powell, W., Waugh, R.,
1997. Direct comparison of level of genetic variation among barley accessionsdetected by RFLPs, AFLPs. Don ssp. italicum. TAG 95, 714–722.
Sala, A., Recio, M., Giner, R.M., Manez, S., Tournier, H., Schinella, G., Rios, J.L., 2002.Anti-inflammatory and antioxidant properties of Helichrysum italicum. J. Pharm.Pharmacol. 54 (3), 365–371.
Sala, A., Recio, M.C., Giner, R.M., Manez, S., Rios, J.L., 2001. New acetophenone glu-cosides isolated from extracts of Helichrysum italicum with antiinflammatoryactivity. J. Nat. Prod. 64 (10), 1360–1362.
Sala, A., Recio, M.C., Schinella, G.R., Mànez, S., Giner, R.M., Cerdà, N.M., Ríos, J.L., 2003.Assessment of the anti inflammatory activity and free radical scavenger activityof tiliroside. Eur. J. Pharmacol. 461, 53–61.
Satta, M., Tuberoso, C.L.G., Angioni, A., Pirisi, F.M., Cabras, P., 1999. Analysis of theessential oil of Helichrysum italicum G. Don. JEOR 11, 711–715.
Scialabba, A., Agrimonti, C., Abbate, G.M., Marmiroli, N., 2008. Assessment of geneticvariation in Sicilian Helichrysum (Asteraceae) and implication to germplasm con-servation. Plant Biol. 142 (2), 287–297.
Smissen, R.D., Breitwieser, I., Ward, J.M., 2007. Hybridization between New ZealandEverlastings Helichrysum lanceolatum and Anaphalioides bellidioides (Asteraceae:Gnaphalieae). Bot. J. Linnean Soc. 154, 89–98.
Süzgeca, S., Mericlia, A.H., Houghtonb, P.J., Cubukcua, B., 2005. Flavonoids ofHelichrysum compactum and their antioxidant and antibacterial activity. Fitoter-apia 76, 269–272.
Tira, S., Di Modica, G., 1967. New �-diketones from Helichrysum italicum G. Don.Tetrahedron Lett. 2, 143–148.
Tucker, A.O., Maciarello, M.J., Charles, D.J., Simon, J.E., 1997. Volatile leaf oil of thecurry plant (Helichrysum italicum (Roth) G. Don subsp. italicum) and dwarf curryplant (subsp. michrophyllum (Willd) Nym) in the North America herb trade.JEOR 9, 583–585.
Tundis, R., Statti, G.A., Conforti, F., Bianchi, A., Agrimonti, C., Sacchetti, G., Muzzoli,M., Ballero, M., Manichini, F., Poli, F., 2005. Influence of enviromental factorson composition of volatile constituents and biological activity of Helichrysumitalicum (Roth) Don (Asteraceae). Nat. Prod. Res. 19, 379–387.
plastique réparatrice et esthétique. Phytothérapie 2, 67–72.Vos, P., Hogers, R., Bleeker, M., Reijans, M., Van De Lee, T., Hornes, M., Frijters, A.,
Pot, J., Peleman, J., Kuiper, M., Zabeau, M., 1995. AFLP: a new technique for DNAfingerprinting. Nucleic Acid Res. 23, 4407–4414.





























