Embed Size (px)
Citation preview

Brief Overview of Potential Ecosystem Impacts of Marine Phosphate
Mining in the Western Cape, South Africa
Prepared for WWF South Africa
by
Jock Currie

1
Introduction
In this report, the potential effects of marine phosphate (or any similar bulk sediment) mining operations on
marine ecosystems in the Western Cape province of South Africa are outlined. The report was prompted by
the application of Green Flash Trading (Pty) Ltd for a licence to prospect for phosphates and other minerals
in extensive areas off the west and south coasts of the Western Cape.
The prospecting phase as laid out by Green Flash Trading (GFT) in their Environmental Management Plans
(EMPs; Green Flash Trading 2012a, 2012b) lacks necessary detail to assess the potential extent and severity
of their impacts on the marine ecosystem. This report does not attempt to address the impacts of those
prospecting activities, but rather focuses on the scenario of full-scale mining of offshore sediments, which
could follow prospecting stages. However to provide a geographical context, which is important in
consideration of the types of habitats and priority conservation areas that could be impacted, the prospecting
licence areas as indicated in the GFT EMPs are employed. This report is restricted to the potential impacts of
the mining process offshore and does not include the impacts resulting from the transfer of dredged material
onshore and their processing or beneficiation. The onshore processing will likely result in large volumes of
waste products, which could contain toxic chemicals and concentrated contaminants (Gecko 2011). During
consideration of any proposed marine mining projects, the ecological and socio-economic implications of
these processes and waste products would require rigorous assessments, in addition to those of the offshore
impacts outlined below.
Phosphate and other bulk sediments have not been mined on a commercial scale from the deep sea floor
anywhere in the world. As a result, potential ecosystem impacts can at best be inferred from other types of
mining operations that have taken place in similar biogeographic regions, at comparable depths and using
similar tools, together with our understanding of the mining strategy and the local oceanography and
ecology. The most comparable existing mining operations would be the deep water diamond mining
activities offshore of the west coast of Namibia (Penney et al. 2007). During its relatively short history, this
vessel-based remote mining strategy has concentrated on locating and recovering localised sediments
containing concentrated diamond deposits. However new methods and technologies have led to exponential
increases in the achievable mining rate of the sea floor (Figure 5.2b in Penney et al. 2007), culminating in
technologies such as the Trailing Suction Hopper-Dredge (TSHD), which allows large volumes of sediments
to be dredged, stored in hopper compartments on board, and transferred to another vessel or onshore for
further processing. TSHD vessels have the ability to dredge sea floor sediments at a rate of > 100 000 m 2/day
(Penney et al. 2007) or 4500 m3/hour (NMP 2012) from depths exceeding 130 m. Such new technologies are
overcoming the economical restraints that have prevented exploitation of deep sea bulk minerals and if these
are to be mined off the Western Cape within the near future, the method used will very likely involve a
TSHD or similar high-volume dredging vessel.
The 223-m TSHD, Cristobal Colon, which Namibian Marine Phosphate (Pty) Ltd proposes to use for their

2
planned phosphate mining off the Namibian coast, has a dredging speed of 1-2 knots and drags an 11-m
wide* dredge head over the sea floor, cutting a trench that is up to 0.75 m deep (NMP 2012). The dredge
heads typically have cutting 'teeth' and powerful water jets to break up hard or consolidated sediments before
they enter the suction tube (Penney et al. 2007). Excess water and fines are released back into the water
column at 10-15 m depth, from a hopper overflow funnel. The 46 000 m3 hopper capacity of the Cristobal
Colon will allow transfer of 64 175 tonnes of sediment to shore per 36-hour dredging cycle in the Namibian
project (NMP 2012). The company plans to eventually mine 5.5 million tonnes of sediment during 47 weeks
at sea annually, removing sediments up to 3 m in depth and over areas of ~2.4 km 2. For greater detail on the
various dredging, trenching and drilling mechanisms employed in the marine diamond mining industry, see
Penney et al. (2007).
As bulk mining of the sea floor and large-scale dredging at these substantial depths has not taken place
before, there is an urgent requirement to understand the impacts of such proposed activities on the
surrounding ecosystems, which encompass unique biodiversity and sustain valuable natural resources. The
following sections outline the potential threats to these ecosystems, as identified from literature covering the
effects of deep water diamond mining and dredging.
Impacts on Demersal and Pelagic Organisms
Flight from Disturbance, Noise and/or Degraded Water Quality
Heemstra (1994) recorded some fish being sucked into large dredges. However, it is assumed that the
majority of mobile organisms that can out-swim the approaching dredge head will escape. Most larger
organisms that are able to actively propel themselves will likely move out of the immediate vicinity, while
those sensitive to sound would avoid a larger area (potentially on the order of several km for certain fish and
mammals; Engås et al. 1996, Southall et al. 2008, Slabbekoorn et al. 2010) around the noise pollution caused
by the surface vessel, the dredging tool on the sea floor and the underwater positioning (sonar) system.
Assuming bulk mining plans would be similar to those of the NMP project in Namibia (NMP 2012), the
dredging vessel will focus on a target mining area for several years and will be at sea for the majority of that
time, creating a semi-permanent source of acoustic disturbance. As large volumes of dredged sediment have
to be transferred ashore frequently (every 37 hours in the Namibian example), a significant increase in
shipping traffic will develop between the mining site and the selected port, creating persistent disturbance
along the transfer route for species sensitive to sound. The scale and implications of acoustic impacts would
require careful investigation for certain species, taking into account their sensitivity, the duration and
intensity of noise disturbances and its locality in relation to the animal's habitat use and migration patterns.
Pulfrich (2010) provides a useful overview of the literature pertaining to acoustic trauma on marine fauna,
with a focus on deep water seismic exploration on the west coast of Namibia.
*This width will likely be reduced to increase the operational depth.

3
Raised Turbidity and Reduction of Light Levels
Both within and below the fines plume, light penetration will be reduced, lowering photosynthetic rates and
visibility. Greatly enhanced concentrations of sediment particles in the water column can also affect
biological processes such as hatching, larval survival and foraging behaviour (Westerberg et al. 1996, Clarke
et al. 2000). Penney et al. (2007) judged the tailings plumes in offshore diamond mining areas of Namibia
not to pose a significant threat to pelagic fauna, arguing that the sediment loads are below lethal
concentrations and that the effects are localised around the mining vessel. Environmental Evaluation Unit
(1996) conclude that the photosynthetic inhibition will be limited to an area < 1 km2 around the vessel and is
of little consequence. As dense phytoplankton blooms frequently reduce visibility in the Benguela region, the
loss of light will likely be of little consequence to other marine organisms.
Biogeochemical Impacts
The chemical composition of sediments and the water-column, as well as bottom water and stratified surface
waters, differ substantially due to differences in biogeochemical rates and processes within them. Therefore
the disturbed benthic plume and the release of fines and bottom water from the surface vessel will alter
water-column chemistry, impacting pelagic, demersal and benthic organisms. Sediments underlying
upwelling ecosystems such as the Benguela are typically enriched with organic matter that has settled from
the productive waters above (Chapman and Shannon 1985). High re-mineralization rates within and on the
surface of the sediments will commonly result in elevated inorganic nutrient concentrations, in many
instances well beyond the concentrations seen in upwelling South Atlantic Central Water (Bailey and
Chapman 1991). The re-mineralization consumes oxygen, frequently causing hypoxic (or anoxic) sediments
and depleted oxygen levels in overlying bottom waters (Brüchert et al. 2009).
Thus a disturbed benthic plume would be expected to impact surrounding oxygen content: Depleted oxygen
concentrations from disturbed interstitial (sediment) water will mix with surrounding bottom water, reducing
oxygen levels in the latter. In addition, re-suspended anoxic sediments would strip oxygen from surrounding
bottom water (Brüchert et al. 2003), while the re-mineralization of re-suspended organic material would
provide a further sink for oxygen. Environmental Evaluation Unit (1996) estimate that lowered oxygen
impacts due to the benthic plume could potentially extend for several km during continuous diamond mining,
although they point out that the severity and resultant ecosystem implications are difficult to predict.
The fines overspill released from the surface vessel will equally contain an elevated organic matter load,
originating from the dredged sediments and from destroyed benthic fauna that is sucked up by the dredge
(Newell et al. 1999). The decomposition of this organic load will increase oxygen demand, potentially
decreasing oxygen concentrations within the water column and on the sediment surfaces where it settles.
Over prolonged time-periods, this continual enrichment could lead to reduced oxygen concentrations below
the thermocline and might aggravate naturally-occurring bottom-water hypoxia. Penney et al. (2007) cite a
study by CSIR (2006) that did not find significantly lowered oxygen concentrations within surveyed

4
sediment plumes in the Atlantic 1 Mining Licence Area offshore of southern Namibia, and argue that organic
enrichment from the overspill of diamond mining vessels is likely negligible compared to the scale of natural
organic loads and bottom-water hypoxia in such highly productive regions. Environmental Evaluation Unit
(1996) estimate that the high sediment load releases from diamond mining vessels would have a low
intensity impact on a spatial scale of about five km 2. The volume, persistence (over time) and relative
concentration of organic enrichment in the discarded fines/water, compared to background particulate
organic matter loads should be carefully considered before this impact is considered negligible to the
surrounding ecosystem.
High inorganic nutrient loads from sediments and bottom water will be released near the surface in the fines
overspill. The current designs of TSHDs release these at a depth of 10-15 m, which would typically be within
the surface mixed layer in offshore Benguela waters (Shannon 1985). The high nutrient loads could provide a
point source of eutrophication, fuelling elevated phytoplankton blooms and microbial productivity
downstream of the mining vessel. The severity of this impact will depend on the volume of waters released
and the scale of mixing within surface waters before the more-dense bottom water sinks down to a level of
neutral buoyancy (Environmental Evaluation Unit 1996). Within the productive upwelled waters of the
Benguela, this point source of eutrophication is likely to have little impact on surrounding ecosystems. In
nutrient-impoverished offshore waters, the ecosystem impact of eutrophication might be more significant.
Re-suspension of Metals
Early studies suggested that Benguela shelf sediments could be associated with harmful concentrations of
heavy metals (Calvert and Price 1970, Chapman and Shannon 1985) and elevated levels of several metals
have been found in sediment samples from the Atlantic 1 Mining area in southern Namibia (Environmental
Evaluation Unit 1996, CSIR 2006). Metal uptake by marine organisms can take place by direct absorption or
via food intake and the effects may include acute toxicity from short-term elevated exposures, or via long-
term bioaccumulation (Penney et al. 2007). Investigating the concentrations of trace elements and pesticides
in sediments and overspill plumes in the Atlantic 1 Mining area, Environmental Evaluation Unit (1996)
estimated that in some areas, plume concentrations of cadmium (Cd), chromium (Cr), copper (Cu), and zinc
(Zn) could exceed the maximum concentrations recommended for the coastal zone by (Lusher 1984). Penney
et al. (2007) argue that the dilution by surrounding waters, scale of the transient diamond mining activities
and dynamic nature of plumes would make chemical contamination of the ecosystem unlikely. Considering
the larger scale and greater persistence at a site of foreseen phosphate mining operations (Coles et al. 2002,
NMP 2012), together with the potential threat of bio-magnification leading to contamination of fishery
resources, site-specific investigations of trace metal concentrations and other contaminants should be
investigated prior to bulk mining activity. If the sediments are found to contain elevated levels of toxic
contaminants, a detailed study should assess the severity, scale and bio-availability of the re-suspended
contaminants in benthic plumes and the fines overspill.

5
Impacts on Benthic Organisms
Physical Destruction of Benthic Organisms and Habitat
From the scale of the heavy machinery involved, the violence of a mechanical 'cutter' and the depth of
sediment removal (on the order of 0.5 m), it is clear that bulk dredging will result in effective destruction and
removal of the benthic ecosystem from within a mined area (Desprez et al. 2000, Penney et al. 2007). Having
reviewed the literature, Penney et al. (2007) conclude that the recovery time of sea bed communities
increases with depth and provide a schematic indicating that areas at 200 m depth can take roughly 20 years
to reach a 'recovered' state. Their definition of a 'recovered state', however, does not necessarily imply a
community statistically similar to pre-disturbance, but one which has reached a mature 'equilibrium' with
similar biomass and diversity indices. Whether such a 'recovered', but altered, community would provide the
same ecosystem functions as the original assemblage, is very hard to assess.
The removal of benthic habitat has implications for the surrounding ecosystem beyond the destruction of
benthic (and slow-moving demersal) organisms from the dredged area. The physical structure created by
organisms such as sponges, gorgonians and deep water corals, as well as the food source provided by the
benthic community, play an essential role in various life-stages of demersal fish and macro-invertebrates
(Auster et al. 1997, Rogers et al. 2008, Baillon et al. 2012, Baker et al. 2012). The animals which were
directly dependent on the destroyed habitat will perish, unless they are able to relocate into similar habitat
nearby that is able to absorb their abundances (i.e. is below carrying capacity).
Changes in Benthic Substrate/Sediment Size
The physical sediment composition is an important determinant of the biological assemblage that inhabits it
(Hall 1994, Rees et al. 1999, Schratzberger et al. 2004). If the remaining sediment is identical to that which
was removed, community composition and structure should have the potential to recover to match non-
dredged reference areas (e.g. Simonini et al. 2007). Frequently however, the depth or particle size of
remaining sediment is different from the pre-disturbed sea floor and the benthic community that re-
establishes afterwards is therefore different to that which occurred prior to dredging (Parkins and Field 1998,
Boyd et al. 2003, Penney et al. 2007). Sediments surrounding the dredged area can also be altered by settling
fines, tailings and benthic plumes, thereby driving a different community structure once dredging ceases and
the fauna recover (Boyd and Rees 2003). In the Atlantic 1 Mining Licence Area in southern Namibia, Parkins
and Field (1998) attributed differences in the sediment composition and its organic content as the drivers of
observed differences in community assemblages between mined and un-mined sites.
Local hydrodynamics and sediment particle size play a large role in the recovery potential of dredged areas
(Boyd et al. 2005), making it difficult to provide blanket estimates of recovery rates. In areas that are
naturally subjected to high deposition rates or high energy environments, the recovery of soft sediments
towards a pre-disturbed state is relatively rapid (Rees et al. 1999, Boyd et al. 2005). Penney et al. (2007)
point out that benthic communities dredged in the Atlantic 1 Mining Licence Area were significantly

6
different to un-dredged areas 9-10 years after dredging stopped, whereas sites further south, perhaps because
they were subject to higher deposition rates from the Orange river, had developed benthic communities
similar to un-mined sites within 5-6 years. Boyd et al. (2005) show that sites dredged in a depth of 22 m and
in a 'moderate' energy environment had communities that were significantly different from non-dredged
reference sites after six years of recovery and conclude that re-establishment of benthic assemblages in low-
energy environments would likely take many years, possibly decades. The depth/location of the GFT licence
areas precludes substantial wave influence or river depositional input and strong currents are not common at
these greater depths.
Considering that remaining sediments will frequently have a different structure to their pre-disturbed state
(Parkins and Field 1998, Boyd et al. 2003, Penney et al. 2007), habitat and resultant ecosystem changes in
such mined areas could be considered permanent, as recovery to pre-disturbed sediment structures (and
hence similar habitat) would occur on geological time scales. If the local hydrodynamics and/or source of
those sediments has changed since they were formed, they may never recover to the pre-disturbed physical
structure (and hence ecological assemblage). This argument would be especially relevant to consolidated or
hard grounds if they were to be mined, unless the remaining post-mined sea floor were to provide the same
physical structure than in its pre-mined state.
Burial and Smothering
There are two different mechanisms by which surrounding (non-dredged) habitats could be impacted by
burial and smothering disturbance. Disturbance of sediments that are not sucked up by the dredge will create
a benthic mining plume, causing a certain amount of burial and smothering adjacent to the dredged
trenches/lanes. The spatial scale of such affected areas would partly depend on the dredging method
(apparatus) and the dredging pattern, besides the local hydrodynamics and sediment characteristics (Boyd
and Rees 2003). Environmental Evaluation Unit (1996) assumed a surrounding area equal to the dredged
area in their estimates of benthic communities impacted by the benthic plume.
The second mechanism would be a result of the fines plume released at or near the surface. There are many
variables which potentially affect the intensity and the geographical scale of such an impact, including the
grain size and density of fines released, water depth, and the vertical water structure and currents (at different
depths). Relatively intricate and site-specific ocean circulation models would have to be employed to gain a
realistic estimate of the intensity and extent of the fines plume and its settling rate on the benthos. Studying
plumes from deep water diamond mining vessels off Namibia, Carter and Midgley (2000) showed that plume
lengths varied between 700 – 5 500 m, with widths of 700 – 3 500 m and that average sediment
concentrations neared background levels about 2 000 m downstream from the vessel. Penney et al. (2007)
rate the impact of diamond mining tailings deposition on benthic organisms as a regional-scale effect of high
environmental significance, especially in deep water habitats that are not naturally subject to high
sedimentation rates. The proportion of discarded sediments in phosphate (or other bulk) mining operations

7
may be lower than that for diamond mining, as the latter is likely more selective of sediment size. However
the overall scale and temporal persistence of dredging activity is likely to be greater for the bulk mining
operations (Coles et al. 2002). Investigating the impacts of marine aggregate extraction on the south coast of
the UK, where no on-board screening takes place and release of fines is minimal, Boyd and Rees (2003)
showed a gradient in macrofaunal community composition with increasing distance (beyond 2 km) from the
point of dredging activity.
Increased Organic Fallout
Both the benthic plume and the fines plume will likely be enriched in organic particles from disturbed or
dredged sediments respectively, which could provide an increased food source for organisms that filter these
particles from the water-column (Boyd et al. 2000, Boyd and Rees 2003). The population or community
impacts within the water-column would likely be negligible, as the majority of filter feeders are planktonic
and would be surrounded by the heightened food concentrations only briefly before it is diluted by
surrounding waters downstream. However benthic filter feeders could gain increased amounts of settling
particulate organic matter (POM) in the area surrounding the mining operation. Boyd and Rees (2003) did
find increased densities of filter-feeders between 500 and 1000 m from a site of intense dredging. If the same
deposit is mined consistently for a period of months to years, a persistent enrichment of settling POM might
alter the benthic community composition.
Biogeochemical Impacts
The biogeochemical impacts for benthic organisms will consist mainly of the reduced oxygen levels in
surface sediments and bottom waters, as outlined in the 'Impacts on Demersal and Pelagic Organisms'
section, hence their description is not repeated here.
Microbial Impacts
Microbial life, including bacteria, fungi and viruses, plays a dominant role in the regulation of
biogeochemical cycles throughout the biosphere, including the deep ocean (e.g. Raghukumar et al. 2001,
Brüchert et al. 2003, Damare et al. 2006, Danovaro et al. 2008). It is unknown how the microbial
communities will recover and what the implications are to the broader ecosystem of their removal or
disruption in mined areas. This question requires urgent attention in future field and impact studies.
Spatial Considerations of Habitat, Conservation Priority and Fisheries
To provide a brief synopsis of the geographical context of the potential impacts discussed above, the GFT
prospecting licence areas have been overlaid onto maps of benthic habitats, conservation focus areas and
fishery footprints.
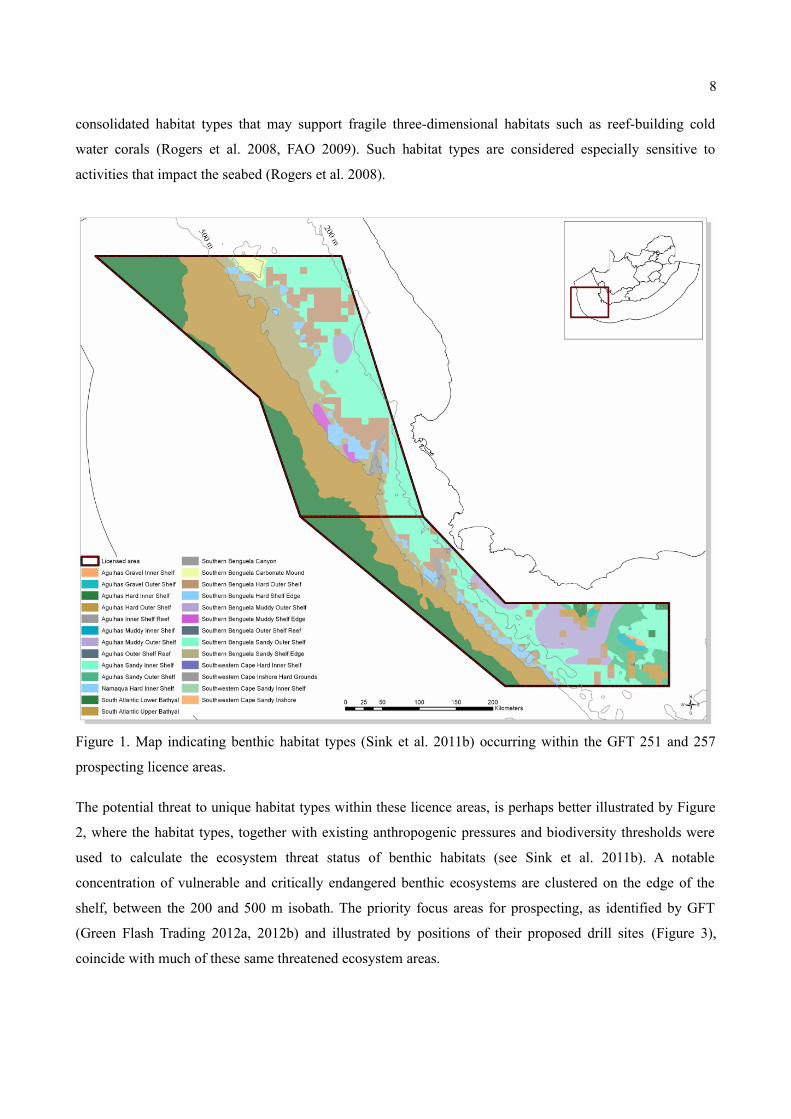
The GFT prospecting licence areas cover 26 recognized benthic habitat types, which fall within several
broader ecozones (Sink et al. 2011b). Some of these habitats fall predominantly within or are restricted
entirely to within the licence area and exist nowhere else. Several of these habitat types are rocky or
8
consolidated habitat types that may support fragile three-dimensional habitats such as reef-building cold
water corals (Rogers et al. 2008, FAO 2009). Such habitat types are considered especially sensitive to
activities that impact the seabed (Rogers et al. 2008).
Figure 1. Map indicating benthic habitat types (Sink et al. 2011b) occurring within the GFT 251 and 257
prospecting licence areas.
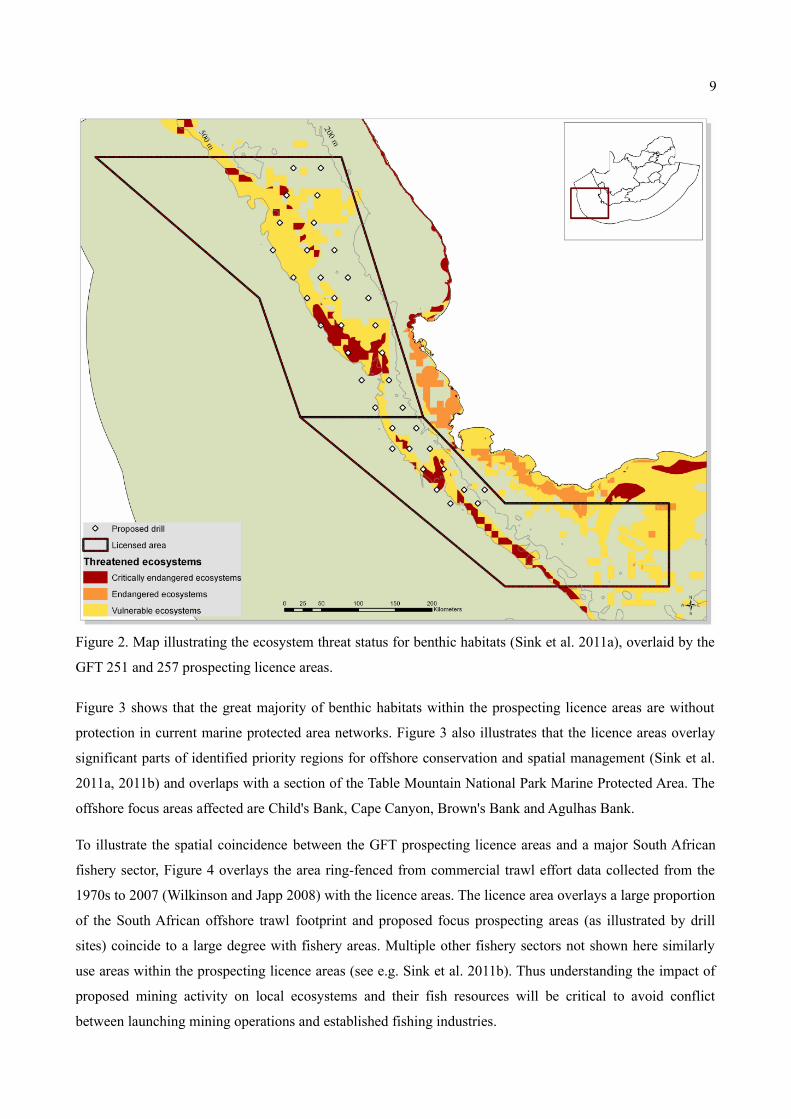
The potential threat to unique habitat types within these licence areas, is perhaps better illustrated by Figure
2, where the habitat types, together with existing anthropogenic pressures and biodiversity thresholds were
used to calculate the ecosystem threat status of benthic habitats (see Sink et al. 2011b). A notable
concentration of vulnerable and critically endangered benthic ecosystems are clustered on the edge of the
shelf, between the 200 and 500 m isobath. The priority focus areas for prospecting, as identified by GFT
(Green Flash Trading 2012a, 2012b) and illustrated by positions of their proposed drill sites (Figure 3),
coincide with much of these same threatened ecosystem areas.
9
Figure 2. Map illustrating the ecosystem threat status for benthic habitats (Sink et al. 2011a), overlaid by the
GFT 251 and 257 prospecting licence areas.
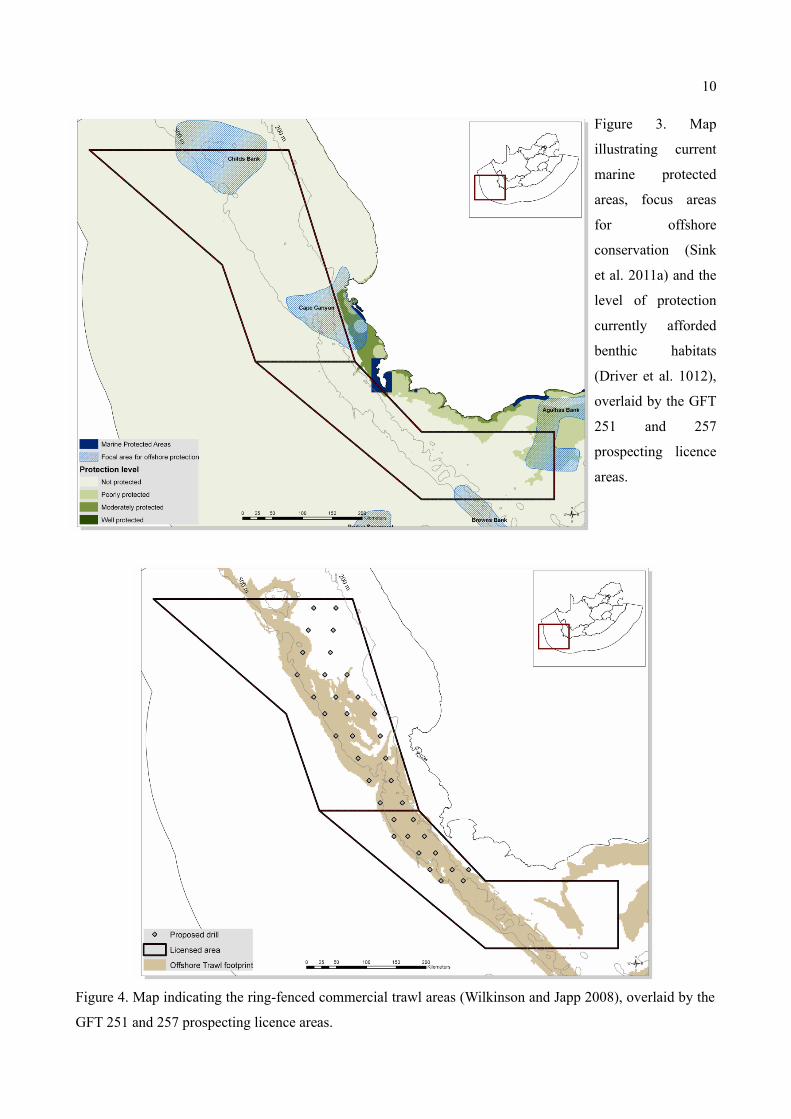
Figure 3 shows that the great majority of benthic habitats within the prospecting licence areas are without
protection in current marine protected area networks. Figure 3 also illustrates that the licence areas overlay
significant parts of identified priority regions for offshore conservation and spatial management (Sink et al.
2011a, 2011b) and overlaps with a section of the Table Mountain National Park Marine Protected Area. The
offshore focus areas affected are Child's Bank, Cape Canyon, Brown's Bank and Agulhas Bank.
To illustrate the spatial coincidence between the GFT prospecting licence areas and a major South African
fishery sector, Figure 4 overlays the area ring-fenced from commercial trawl effort data collected from the
1970s to 2007 (Wilkinson and Japp 2008) with the licence areas. The licence area overlays a large proportion
of the South African offshore trawl footprint and proposed focus prospecting areas (as illustrated by drill
sites) coincide to a large degree with fishery areas. Multiple other fishery sectors not shown here similarly
use areas within the prospecting licence areas (see e.g. Sink et al. 2011b). Thus understanding the impact of
proposed mining activity on local ecosystems and their fish resources will be critical to avoid conflict
between launching mining operations and established fishing industries.
10
Figure 3. Map
illustrating current
marine protected
areas, focus areas
for offshore
conservation (Sink
et al. 2011a) and the
level of protection
currently afforded
benthic habitats
(Driver et al. 1012),
overlaid by the GFT
251 and 257
prospecting licence
areas.
Figure 4. Map indicating the ring-fenced commercial trawl areas (Wilkinson and Japp 2008), overlaid by the
GFT 251 and 257 prospecting licence areas.

11
Conclusions and Recommendations
Drawing on the literature discussed above, bulk sediment mining of the deep sea floor in the GFT
prospecting licence areas 251 and 257 will have severe impacts on the benthic habitats that are dredged.
Benthic and demersal organisms in adjacent areas will suffer direct and knock-on effects, the extent, intensity
and duration of which will depend on many factors related to the mining strategy and site-specific sediment,
biogeochemical and ecosystem properties. While pelagic waters will be affected locally, dilution with
downstream waters will likely reduce the direct ecological impact to a large degree. The most likely
identified impacts of foreseen mining methods in these deep ecosystems are briefly touched on above.
Whether there are synergistic effects, what the cumulative and knock-on effects might be, and the overall
implications to biodiversity, ecosystem functioning and fishery resources, will require far more attention. In-
depth and site-specific studies with detailed input on the mining methods and tools, resource target, local
geology, oceanography, biogeochemistry, ecosystems and life-cycles of fishery resources, will be required to
estimate the ecological and economic impacts from specific mining projects. Certain of these impacts could
be partly mitigated, although the destruction, and in many instances, permanent alteration of sea floor habitat
seems inevitable with dredging of deep sea environments.
The implications of such mining activities for biodiversity conservation will depend to a large degree on the
extent of the habitat type in question. Certain unique habitats are extremely restricted in their spatial extent
and may already be threatened by other ocean uses. A bulk sediment mining operation in or near such a
habitat could provide a real threat to its survival. Other benthic habitats may be more ubiquitous and wide-
spread and hence mining in such habitats might threaten only a small fraction of the total area covered by
similar assemblages of species. It is imperative to carefully consider the location of mining relative to
threatened ecosystems and priority conservation targets. As the mining can result in a permanently altered
community, conservation of unique habitat types and their assemblage of biodiversity is not compatible with
bulk sediment mining of the same area.
Due to extraction of a renewable resource, fisheries are by their nature sustainable if managed correctly.
Mining of valuable minerals or other geological materials is generally not sustainable, as the formation rate
of target minerals (and surrounding disturbed sediments) is far outpaced by the rate of resource extraction.
South Africa is a signatory to the Reykjavik Declaration, prescribing an Ecosystem Approach to Fisheries
management, which requires the holistic approach of maintaining a healthy ecosystem in support of
managing fishery resources sustainably. The damage to benthic habitats and surrounding ecosystems has the
potential to impact on fishery resources, hence the location of proposed mining activity should be assessed
with careful consideration of existing fishery footprints, as well as habitats or ecosystems that are critical to
various life-stages of the fishery species. Baillon et al. (2012) have recently showed close association
between the larvae of commercially harvested redfish (Sebastes spp) and cold-water corals in the north
Atlantic. Such relationships, many of which are yet to be discovered, are the reason researchers and fishery

12
managers support the ecosystem approach to fisheries.
If bulk mining of the sea floor is to be considered, the financial gains and job creation for local communities
need to be weighed up against the economic, social and ethical implications of permanent alteration of mined
habitats, loss of biodiversity, and potential ecological knock-on effects and disturbance to fishery resources.
There is growing recognition globally that the loss of ecosystem services and costs to future resource users
need to be taken into consideration, and compensated by the proponents, prior to development of industries
that impact on the 'natural capital' of biodiversity and ecosystems (Costanza et al. 1997, Turner et al. 2001,
Sukhdev 2011).
References
Auster PJ, Malatesta RJ, Donaldson CLS. 1997. Distributional responses to small-scale habitat variability by
early juvenile silver hake, Merluccius bilinearis. Environmental Biology of Fishes 50: 195–200.
Bailey GW, Chapman P. 1991. Short-term variability during an anchor station study in the southern Benguela
upwelling system: Chemical and physical oceanography. Progress In Oceanography 28: 9–37.
Baillon S, Hamel J-F, Wareham VE, Mercier A. 2012. Deep cold-water corals as nurseries for fish larvae .
Frontiers in Ecology and the Environment in press.
Baker KD, Haedrich RL, Snelgrove PVR, Wareham VE, Edinger EN, Gilkinson KD. 2012. Small-scale
patterns of deep-sea fish distributions and assemblages of the Grand Banks, Newfoundland continental
slope. Deep Sea Research Part I: Oceanographic Research Papers 65: 171–188.
Boyd SE, Limpenny DS, Rees HL, Cooper KM. 2005. The effects of marine sand and gravel extraction on
the macrobenthos at a commercial dredging site (results 6 years post-dredging). ICES Journal of Marine
Science 62: 145–162.
Boyd SE, Limpenny DS, Rees HL, Cooper KM, Campbell S. 2003. Preliminary observations of the effects
of dredging intensity on the re-colonisation of dredged sediments off the southeast coast of England (Area
222). Estuarine, Coastal and Shelf Science 57: 209–223.
Boyd SE, Rees HL, Richardson CA. 2000. Nematodes as Sensitive Indicators of Change at Dredged Material
Disposal Sites. Estuarine, Coastal and Shelf Science 51: 805–819.
Boyd S., Rees H. 2003. An examination of the spatial scale of impact on the marine benthos arising from
marine aggregate extraction in the central English Channel. Estuarine, Coastal and Shelf Science 57: 1–16.
Brüchert V, Currie B, Peard KR. 2009. Hydrogen sulphide and methane emissions on the central Namibian
shelf. Progress in Oceanography 83: 169–179.

13
Brüchert V, Jorgensen BB, Neumann K, Riechmann D, Schlösser M, Schulz H. 2003. Regulation of bacterial
sulfate reduction and hydrogen sulfide fluxes in the central Namibian coastal upwelling zone . Geochimica et
Cosmochimica Acta 67: 4505–4518.
Calvert SE, Price NB. 1970. Minor Metal Contents of Recent Organic-rich Sediments off South West Africa
227: 593–595.
Carter RA, Midgley J. 2000. Characteristics of fine tailings plumes generated by ship-based marine diamond
mining off Lüderitz: Dimensions and effects on water quality. In: Proceedings of the Symposium on Co-
Management of Resources off the southwestern coast of Africa, 20-24 June 2000. Lüderitz, Namibia.
Chapman P, Shannon LV. 1985. The Benguela Ecosystem. Part II. Chemistry and related processes .
Oceanography and Marine Biology: An Annual Review 23: 183–251.
Clarke D, Engler RM, Wilber DH. 2000. Assessment of Potential Impacts of Dredging Operations Due to
Sediment Resuspension., DOER Technical Notes Collection (ERDC TN-DOER-E9), U.S. Army Engineer
Research and Development Center. Vicksburg.
Coles SKP, Wright CI, Sinclair DA, Bossche PV den. 2002. The Potential for Environmentally Sound
Development of Marine Deposits of Potassic and Phosphatic Minerals Offshore, Southern Africa . Marine
Georesources and Geotechnology 20: 87–110.
Costanza R, d’ Arge R, Groot R de, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O’Neill RV,
Paruelo J, Raskin RG, Sutton P, Belt M van den. 1997. The value of the world’s ecosystem services and
natural capital. Nature 387: 253–260.
CSIR. 2006. Physical Effects of Sediment Discharged from Marine Dredging and Plant Operations in the
Atlantic 1 and Uubvlei Regions. CSIR Report No. CSIR/NRE/ECO/ER/2006/0203/C.
Damare S, Raghukumar C, Raghukumar S. 2006. Fungi in deep-sea sediments of the Central Indian Basin .
Deep Sea Research Part I: Oceanographic Research Papers 53: 14–27.
Danovaro R, Dell’Anno A, Corinaldesi C, Magagnini M, Noble R, Tamburini C, Weinbauer M. 2008. Major
viral impact on the functioning of benthic deep-sea ecosystems. Nature 454: 1084–1087.
Desprez M. 2000. Physical and biological impact of marine aggregate extraction along the French coast of
the Eastern English Channel: short- and long-term post-dredging restoration. ICES Journal of Marine
Science 57: 1428–1438.
Driver A, Sink KJ, Nel JN, Holness S, van Niekerk L, Daniels F, Jonas Z, Majiedt PA, Harris L, Maze K.
2012. National Biodiversity Assessment 2011: An assessment of South Africa’s biodiversity and ecosystems.
Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs,
Pretoria.

14
Engås A, Løkkeborg S, Ona E, Soldal AV. 1996. Effects of seismic shooting on local abundance and catch
rates of cod (Gadus morhua) and haddock (Melanogrammus aeglefinus). Canadian Journal of Fisheries and
Aquatic Sciences 53: 2238–2249.
Environmental Evaluation Unit. 1996. Impacts of deep sea diamond mining, in the Atlantic 1 Mining
Licence Area in Namibia, on the natural systems of the marine environment. Environmental Evaluation Unit
Report No. 11/96/158. University of Cape Town. Prepared for De Beers Marine (Pty) Ltd. pp. 370.
FAO. 2009. International guidelines for the management of deep-sea fisheries in the high seas. FAO, Rome.
Gecko. 2011. Proposed Vision Industrial Park: Comparative marine environmental risk assessment of two
proposed locations for the Vision Industrial Park. Prepared for Gecko Industries (Pty) Ltd, Namibia. CSIR,
PISCES Environmental Services, Steffani Marine Environmental Consultant.
Green Flash Trading. 2012a. Environmental Management Plan in the ocean off Cape Columbine and Cape
Infanta, Western Cape Province. Green Flash Trading 257 (Pty) Ltd.
Green Flash Trading. 2012b. Environmental Management Plan in the ocean off Adam Se Baai and the area to
the south towards Table Bay, Western Cape Province. Green Flash Trading 251 (Pty) Ltd.
Hall SJ. 1994. Physical disturbance and marine benthic communities: life in unconsolidated sediments.
Oceanography and Marine Biology: An Annual Review 32: 179–239.
Heemstra PC. 1994. Assessment of deep-sea mining effects in the fish fauna off the south coast of Namibia
and west coast of South Africa from Lüderitz, Namibia to the mouth of the Olifants River at depths of 85 to
200 metres, Report by the J. L. B. Smith Institutes of Ichthyology.
Lusher JA. 1984. Water quality criteria for the South African coastal zone. South African National Scientific
Programmes Report No. 94. CSIR, Pretoria.
Newell R., Hitchcock D., Seiderer L. 1999. Organic enrichment associated with outwash from marine
aggregates dredging: a probable explanation for surface sheens and enhanced benthic production in the
vicinity of dredging operations. Marine Pollution Bulletin 38: 809–818.
NMP. 2012. Environmental impact assessment report for the marine component: Proposed recovery of
phosphate enriched sediments from the Marine Licence Area No. 170 off Walvis Bay Namibia. Prepared for
Namibian Marine Phosphate (Pty) Ltd. J Midgley and Associates, Enviro Dynamics, CSIR.
Parkins CA, Field JG. 1998. The effects of deep sea diamond mining on the benthic community structure of
the Atlantic 1 Mining Licence Area. Annual Monitoring Report – 1997, Unpublished report to De Beers
Marine, April 1988.
Penney AJ, Pulfrich A, Rogers J, Steffani N, Mabille V. 2007. Data Gathering and Gap Analysis for

15
Assessment of Cumulative Effects of Marine Diamond Mining Activities on the BCLME Region. Project:
BEHP/CEA/03/02. Final Report to the BCLME mining and petroleum activities task group.
Pulfrich A. 2010. Environmental Impact Assessment for proposed seismic surveys in the central Walvis
Basin and southern Orange Basin areas, Namibia: Marine faunal assessment . Prepared for HRT Oil and Gas
and Universal Power Corp.
Raghukumar C, Bharathi PAL, Ansari Z., Nair S, Ingole B, Sheelu G, Mohandass C, Nagender Nath B,
Rodrigues N. 2001. Bacterial standing stock, meiofauna and sediment–nutrient characteristics: indicators of
benthic disturbance in the Central Indian Basin. Deep Sea Research Part II: Topical Studies in
Oceanography 48: 3381–3399.
Rees HL, Pendle MA, Waldock R, Limpenny DS, Boyd SE. 1999. A comparison of benthic biodiversity in
the North Sea, English Channel, and Celtic Seas. ICES Journal of Marine Science 56: 228–246.
Rogers AD, Clark MR, Hall-Spencer JM, Gjerde KM. 2008. The Science behind the Guidelines: A Scientific
Guide to the FAO Draft International Guidelines (December 2007) For the Management of Deep-Sea Fisher-
ies in the High Seas and Examples of How the Guidelines May Be Practically Implemented. IUCN,
Switzerland.
Schratzberger M, Whomersley P, Kilbride R, Rees H l. 2004. Structure and taxonomic composition of
subtidal nematode and macrofauna assemblages at four stations around the UK coast. Journal of the Marine
Biological Association of the United Kingdom 84: 315–322.
Shannon LV. 1985. The Benguela Ecosystem, Part 1. Evolution of the Benguela, physical features and
processes. Oceanography and Marine Biology: An Annual Review 23: 105–182.
Simonini R, Ansaloni I, Bonini P, Grandi V, Graziosi F, Iotti M, Massamba-N’Siala G, Mauri M, Montanari
G, Preti M, De Nigris N, Prevedelli D. 2007. Recolonization and recovery dynamics of the macrozoobenthos
after sand extraction in relict sand bottoms of the Northern Adriatic Sea . Marine Environmental Research 64:
574–589.
Sink KJ, Attwood CG, Lombard AT, Grantham HS, Leslie R, Samaai T, Kerwath SE, Majiedt P, Fairweather
TP, Hutchings L, Van der Lingen CD, Atkinson LJ, Wilkinson S, Holness S, Wolf T. 2011a. Systematic
planning to identify focus areas for offshore biodiversity protection in South Africa , Final report for the
Offshore Marine Protected Area project. South African National Biodiversity Institute, Cape Town.
Sink KJ, Holness S, Harris L, Majiedt P, Atkinson LJ, Robinson T, Kirkman S, Hutchings L, Leslie R,
Lamberth S, Kerwath SE, von der Heyden S, Lombard AT, Attwood CG, Branch GM, Fairweather TP,
Taljaard S, Weerts S, Cowley PD, Awad A, Halpern BS, Grantham HS, Wolf T. 2011b. National Biodiversity
Assessment 2011: Technical Report. Volume 4: Marine and Coastal Component. South African National
Biodiversity Institute, Pretoria.

16
Slabbekoorn H, Bouton N, van Opzeeland I, Coers A, ten Cate C, Popper AN. 2010. A noisy spring: the
impact of globally rising underwater sound levels on fish. Trends in Ecology & Evolution 25: 419–427.
Southall BL, Bowles AE, Ellison WT, Finneran JJ, Gentry RL, Greene CR, Kastak D, Ketten DR, Miller JH,
Nachtigall PE, Richardson WJ, Thomas JA, Tyack PL. 2008. Marine mammal noise exposure criteria: Initial
scientific recommendations. Bioacoustics 17: 273–275.
Sukhdev P. 2011. Putting a price on nature: the economics of ecosystems and biodiversity . Solutions 1: 34–
43.
Turner RK, Bateman IJ, Adger WN. 2001. Ecological economics and coastal zone ecosystems’ values: an
overview. In: Turner RK, Bateman IJ, Adger WN (eds), Economics of Coastal and Water resources: Valuing
Environmental Functions. Kluwer Academic Publishers, Dordrecht. pp 1–43.
Westerberg H, Rönnbäck P, Frimansson H. 1996. Effects on suspended sediments on cod egg and larvae and
on the behaviour of adult herring and cod. In: ICES Council Meeting Papers. p 13.
Wilkinson S, Japp D. 2008. Spatial boundaries of the South African hake-directed trawling industry: Trawl
Footprint Estimation prepared for the South African Deepsea Trawling Industry Association. Capricorn
Fisheries Monitoring cc, Cape Town.





























