Embed Size (px)
Citation preview

Analytical Biochemistry 304, 174–179 (2002)
Homogenous Assays for Escherichia coli DnaB-StimulatedDnaG Primase and DnaB Helicase and Their Usein Screening for Chemical Inhibitors
Yi Zhang, Fude Yang, Yeh-Chih Kao,1 Michael G. Kurilla,2 David L. Pompliano,3 and Ira B. Dicker4
Bristol-Myers Squibb Company, Pharmaceutical Research Institute, Wilmington, Delaware 19880
Escherichia coli DnaG primase is a single-strandedDNA-dependent RNA polymerase. Primase catalyzesthe synthesis of a short RNA primer to initiate DNAreplication at the origin and to initiate Okazaki frag-ment synthesis for synthesis of the lagging strand.Primase activity is greatly stimulated through its in-teraction with DnaB helicase. Here we report a 96-wellhomogeneous scintillation proximity assay (SPA) forthe study of DnaB-stimulated E. coli primase activityand the identification of E. coli primase inhibitors. Theassay uses an adaptation of the general priming reac-tion by employing DnaG primase, DnaB helicase, andribonucleotidetriphosphates (incorporation of [3H]CTP)for in vitro primer synthesis on single-stranded oligo-nucleotide and M13mp18 DNA templates. The primaseproduct is captured by polyvinyl toluene–polyethyl-eneimine-coated SPA beads and quantified by count-ing by beta-scintography. In the absence of helicase asa cofactor, primer synthesis is reduced by 85%. Theprimase assay was used for screening libraries of com-pounds previously identified as possessing antimicro-bial activities. Primase inhibitory compounds werethen classified as direct primase inhibitors or mixedprimase/helicase inhibitors by further evaluation in aspecific assay for DnaB helicase activity. By this ap-proach, specific primase inhibitors could be identified.© 2002 Elsevier Science (USA)
Key Words: scintillation proximity assay; primase;DnaG; helicase; DnaB.
1 This paper is dedicated in memoriam to Dr. Yeh-Chih Kao.2 Current address: [email protected] Current address: GlaxoSmithKline, King of Prussia, PA 19406.4 To whom correspondence should be addressed at Bristol-Myers
Squibb Company, Infectious Diseases, 5 Research Parkway, Depart-ment 106, Rm. 350M, Wallingford, CT 06492-1996. Fax: (203) 677-
6088. E-mail: [email protected].174
The global antimicrobial market is dominated bycompounds of only a few structural classes such asbroad-spectrum beta-lactams, macrolides, fluoroquino-lones, tetracyclines, aminoglycosides, and glycopep-tides. Although these compounds are generally effec-tive antibiotics, their utility has become limited insome situations due to the increasing prevalence ofresistant organisms. Thus, the development of antibi-otics with novel mechanisms of action may provide asolution to this problem.
DNA replication is a known target of the clinicallyimportant fluoroquinolone class of antibiotics whichinterfere with DNA gyrase and topoisomerase IV andare bactericidal. The quinolones fulfill the three essen-tial criteria for a good antibiotic; i.e., the activities oftheir targets are essential, they have a broad spectrum,and they are selective for their prokaryotic versus eu-karyotic counterparts. However, there are currently nomarketed antibacterial drugs that target other compo-nents of the replication machinery, though novel inhib-itors of PolC, the main replicative polymerase of gram-positive bacteria, have been reported (1). Targetingsuch components may lead to novel antibiotics andthereby overcome the problem of resistance.
The priming of DNA synthesis in Escherichia coli isthought to require the interaction of the primase(DnaG)5 and helicase (DnaB) enzymes at the replica-tion fork (2, 3), but additional interactions betweenthese proteins and other components of the replisomehave been suggested (4, 5). Because of the centrality ofthe priming activity of DnaG, and thus its attractive-ness as a drug target, our objective has been to identifynovel antibiotics which target this activity. To simplify
5 Abbreviations used: SPA, scintillation proximity assay; PVT-PEI, polyvinyl toluene–polyethyleneimine; primase, DnaG; helicase,DnaB; S/N, signal to noise; DTT, dithiothreitol; BSA, bovine serum
Received October 2, 2001; published online April 11, 2002
doi:10.1006/abio.2002.5627, available online at http://www.idealibra
om on ry.calbumin; DMSO, dimethyl sulfoxide.
0003-2697/02 $35.00© 2002 Elsevier Science (USA)
All rights reserved.

matters, we used the general priming method (6, 7) forprimer synthesis which requires only the DnaG andDnaB enzymes. Herein, we describe a novel homoge-neous assay which is useful for studying these enzymesand which also allows for the detection of inhibitors ofE. coli DnaG and DnaB. Also described is a specificDnaB helicase assay, useful in the assignment of mech-anism of action for those compounds which inhibitDnaB-stimulated DnaG activity.
MATERIAL AND METHODS
Materials
Unlabeled oligodeoxynucleotides were purchasedfrom Sigma-Genosys. PVT-PEI SPA beads were fromAmersham. White opaque 96-well Optiplates, for SPApurposes, and black 96-well HTRF-9 Optiplates, forfluorescence assays, were purchased from Packard In-strument Co. (Meriden, CT). [3H]CTP was purchasedfrom NEN (specific activity 20 �Ci/pmol). M13mp18ssDNA was purchased from New England Biolabs (No.4040S). RNA guard was purchased from AmershamPharmacia (No. 27-0861-01; 26 U/�l). Adenosine 5�-O-(3-thiotriphosphate) (ATP-�-S) and Novobiocin werepurchased from Sigma. The Texas Red-labeled oligode-oxynucleotide used in the helicase assay was pur-chased from Synthegen, Inc. (Houston, TX). Overex-pression clones for E. coli DnaG and DnaB were kindlyprovided by Dr. Kenneth Marians, Memorial Sloan-Kettering Cancer Center. DnaG and DnaB were puri-fied as previously described (8).
SPA Primase Assay
Primase activity was monitored by a SPA method asfollows in buffer A (50 mM Tris–HCl (pH 7.5), 4%wt/vol sucrose, 8 mM DTT, 5 mM MgCl2, 40 �g/ml BSA,0.1 �g/�l Rifampicin, 25 unit/ml RNA guard, 100 �MGTP, 100 �M UTP, 3 �M CTP, 1 mM ATP). To eachwell of a 96-well Packard white opaque Optiplate mi-crotiterplate were added sequentially 1 �l DMSO ortest compound, 20 �l of 2.5X DnaG, and 20 �l of 2.5XDnaB. Unless specifically noted, reactions contained208 nM DnaG, 8 nM DnaB, and 3.3 nM mp18 ssDNA or0.1 �g single-stranded oligonucleotide template andwere conducted at 30°C. Reactions were initiated byadding 10 �l of buffer A containing a mixture of 5XDNA and 0.4 �Ci [3H]CTP (final specific activity[3H]CTP 2.7 Ci/mmol). Reactions were incubated for 30min (or the indicated times) at which time a suspensionof 50 �l of 2.5 mg/ml PVT-PEI SPA beads (Amersham;prepared in 0.3 M Na–Citrate, pH 3.0) were added.Plates were read after 1 h on a Topcount instrument(Packard). For inhibition studies, residual DnaG activ-ity was defined as activity which was above that re-corded for control reactions which were without both
the DnaG and the DnaB enzymes. Inhibition studieswere conducted at 30°C for 30 min and the concentra-tions or amounts of cold CTP, [3H]CTP, DnaG, DnaB,and M13mp18 ssDNA were 3 �M, 0.4 �Ci, 208 nM, 8nM, and 3.3 nM, respectively. The counting efficiencyby the primase SPA method using the Topcount instru-ment was assessed by comparison to a filter-bindingmethod in which 10 �l of the reaction mixture wasapplied to a 2.5-cm DE81 filter paper, washed threetimes with both 0.5 M Na2HPO4 and water, dried byvacuum filtration, and counted in 5 ml scintillationcocktail (9). The counting efficiency (25%) was used todetermine specific activities for the primase substratesof Table 1.
Homogeneous Fluorescence Energy Transfer (FRET)DnaB Helicase Assay
DnaB helicase activity was monitored by a FRETmethod in which the fluorescence of a double-stranded,forked DNA substrate, labeled on the 5� end with thefluorochrome Texas Red, is internally quenched by aDabcyl moiety located on the complementary strand.Upon unwinding, the Texas Red-labeled oliogo is cap-tured by a complementary strand lacking a quenchermoiety, thus allowing continuous monitoring of helicaseactivity.
The substrate was prepared by combining solutionsof the following oligodeoxynucleotides at room temper-ature in a 1/1.8 ratio in buffer A (10 mM Tris–HCl, pH7.6, 25 mM NaCl, 1 mM EDTA): oligo A (5�-TTT TTTTTT TTT TTT TTT TTT TTT TTT TTT CCG TCT CACTGG TGA AAA GAA AAA CCA CCC-3�; tagged on the3� end with Texas Red) and oligo B (5�-GGG TGG TTTTTC TTT TCA CCA GTG AGA CGG TTT TTT TTTTTT TTT TTT TTT TTT TTT TTT-3�; tagged on the 5�end with Dabcyl).
TABLE 1
Effect of Oligodeoxynucleotide Chain Lengthon the Specific DnaG Activity
Length Specific activity
87 3.3 � 0.445 3.3 � 0.535 4.1 � 0.325 0.15 � 0.02
Note. The substrate sequences were 5� CCCACCAAAAAGAA-25 35 45’ ’ ’
AAGTGGTCACTCTGCCCGTTGTCGACTAACGGGAAGTGGCGG-87’
ACCGGACTCTCTCAACGTCGTTCGCCAGGTG 3� . Activity is de-fined as pmol CTP incorporated/nmol DnaG/min for reactions con-taining 8 nM DnaB and 0.1 �g template. Assay conditions were asdescribed under Material and Methods except that M13mp18 ssDNAwas replaced by the designated oligos. The data represent themean � range/2 for two experiments.
175DnaG PRIMASE SCINTILLATION PROXIMITY ASSAY
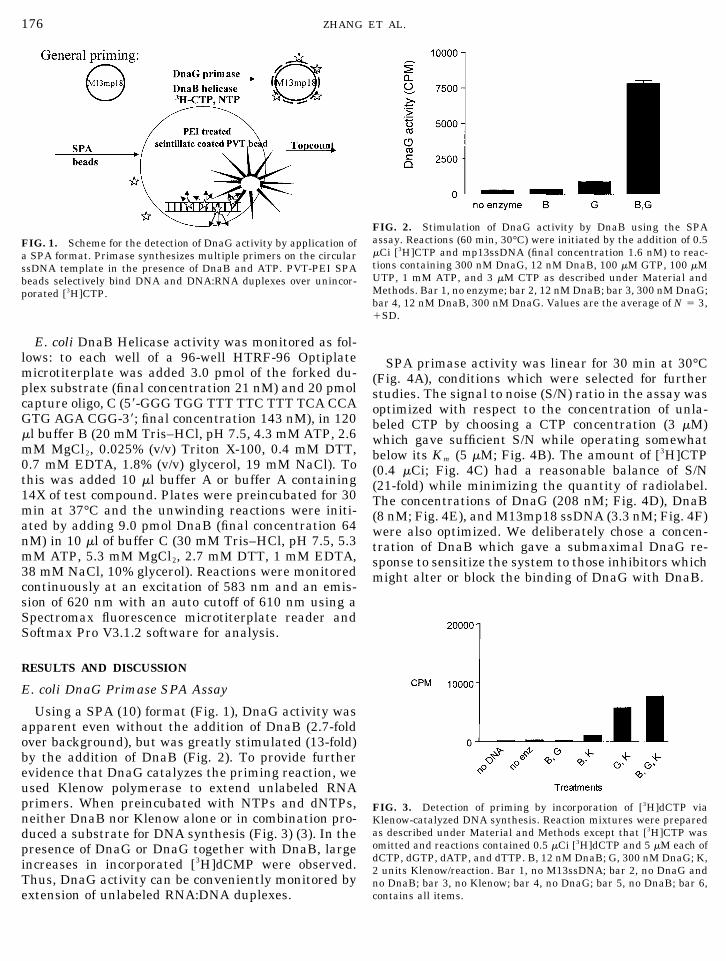
E. coli DnaB Helicase activity was monitored as fol-lows: to each well of a 96-well HTRF-96 Optiplatemicrotiterplate was added 3.0 pmol of the forked du-plex substrate (final concentration 21 nM) and 20 pmolcapture oligo, C (5�-GGG TGG TTT TTC TTT TCA CCAGTG AGA CGG-3�; final concentration 143 nM), in 120�l buffer B (20 mM Tris–HCl, pH 7.5, 4.3 mM ATP, 2.6mM MgCl2, 0.025% (v/v) Triton X-100, 0.4 mM DTT,0.7 mM EDTA, 1.8% (v/v) glycerol, 19 mM NaCl). Tothis was added 10 �l buffer A or buffer A containing14X of test compound. Plates were preincubated for 30min at 37°C and the unwinding reactions were initi-ated by adding 9.0 pmol DnaB (final concentration 64nM) in 10 �l of buffer C (30 mM Tris–HCl, pH 7.5, 5.3mM ATP, 5.3 mM MgCl2, 2.7 mM DTT, 1 mM EDTA,38 mM NaCl, 10% glycerol). Reactions were monitoredcontinuously at an excitation of 583 nm and an emis-sion of 620 nm with an auto cutoff of 610 nm using aSpectromax fluorescence microtiterplate reader andSoftmax Pro V3.1.2 software for analysis.
RESULTS AND DISCUSSION
E. coli DnaG Primase SPA Assay
Using a SPA (10) format (Fig. 1), DnaG activity wasapparent even without the addition of DnaB (2.7-foldover background), but was greatly stimulated (13-fold)by the addition of DnaB (Fig. 2). To provide furtherevidence that DnaG catalyzes the priming reaction, weused Klenow polymerase to extend unlabeled RNAprimers. When preincubated with NTPs and dNTPs,neither DnaB nor Klenow alone or in combination pro-duced a substrate for DNA synthesis (Fig. 3) (3). In thepresence of DnaG or DnaG together with DnaB, largeincreases in incorporated [3H]dCMP were observed.Thus, DnaG activity can be conveniently monitored byextension of unlabeled RNA:DNA duplexes.
SPA primase activity was linear for 30 min at 30°C(Fig. 4A), conditions which were selected for furtherstudies. The signal to noise (S/N) ratio in the assay wasoptimized with respect to the concentration of unla-beled CTP by choosing a CTP concentration (3 �M)which gave sufficient S/N while operating somewhatbelow its Km (5 �M; Fig. 4B). The amount of [3H]CTP(0.4 �Ci; Fig. 4C) had a reasonable balance of S/N(21-fold) while minimizing the quantity of radiolabel.The concentrations of DnaG (208 nM; Fig. 4D), DnaB(8 nM; Fig. 4E), and M13mp18 ssDNA (3.3 nM; Fig. 4F)were also optimized. We deliberately chose a concen-tration of DnaB which gave a submaximal DnaG re-sponse to sensitize the system to those inhibitors whichmight alter or block the binding of DnaG with DnaB.
FIG. 1. Scheme for the detection of DnaG activity by application ofa SPA format. Primase synthesizes multiple primers on the circularssDNA template in the presence of DnaB and ATP. PVT-PEI SPAbeads selectively bind DNA and DNA:RNA duplexes over unincor-porated [3H]CTP.
FIG. 2. Stimulation of DnaG activity by DnaB using the SPAassay. Reactions (60 min, 30°C) were initiated by the addition of 0.5�Ci [3H]CTP and mp13ssDNA (final concentration 1.6 nM) to reac-tions containing 300 nM DnaG, 12 nM DnaB, 100 �M GTP, 100 �MUTP, 1 mM ATP, and 3 �M CTP as described under Material andMethods. Bar 1, no enzyme; bar 2, 12 nM DnaB; bar 3, 300 nM DnaG;bar 4, 12 nM DnaB, 300 nM DnaG. Values are the average of N � 3,�SD.
FIG. 3. Detection of priming by incorporation of [3H]dCTP viaKlenow-catalyzed DNA synthesis. Reaction mixtures were preparedas described under Material and Methods except that [3H]CTP wasomitted and reactions contained 0.5 �Ci [3H]dCTP and 5 �M each ofdCTP, dGTP, dATP, and dTTP. B, 12 nM DnaB; G, 300 nM DnaG; K,2 units Klenow/reaction. Bar 1, no M13ssDNA; bar 2, no DnaG andno DnaB; bar 3, no Klenow; bar 4, no DnaG; bar 5, no DnaB; bar 6,contains all items.
176 ZHANG ET AL.

Use of Oligodeoxynucleotides as Substrates
Four oligodeoxynucleotides (Table 1), with lengthsfrom 25 to 87 bp (random sequence from M13mp18ssDNA), were evaluated as DnaG substrates (11). Thespecific activities of the 35-, 45-, and 87-mers werecomparable. The 25-mer was not a good substrate inthe assay, possibly because this is too short an oligode-oxynucleotide for the assembly of the DnaG:DNA com-plex.
Screening of Compound Libraries Using the PrimaseSPA Assay
Compounds with demonstrated antibacterial activ-ity (library 1 or 2) were tested in the DnaG/DnaB SPA.Library 1 contained 162/976 compounds (18%) whichinhibited activity by �70% (Fig. 5A) while library 2contained 37/626 compounds (5.9%) which inhibitedactivity by �70% (Fig. 5B). As a positive control, eachplate contained a serial dilution of aurintricarboxylic
FIG. 4. The signal to noise in the assay was optimized with respect to time (30 min) and temperature (A) and the concentrations ofunlabeled CTP (B), [3H]CTP (C), DnaG (D), DnaB (E), and M13ssDNA (F). Unless varied, reactions were conducted at 30°C for 30 min andthe concentrations or amounts of cold CTP, [3H]CTP, DnaG, DnaB, and M13mp18 ssDNA were 3 �M, 0.4 �Ci, 208 nM, 8 nM, and 3.3 nM,respectively. Values are average of N � 3 �SD.
177DnaG PRIMASE SCINTILLATION PROXIMITY ASSAY
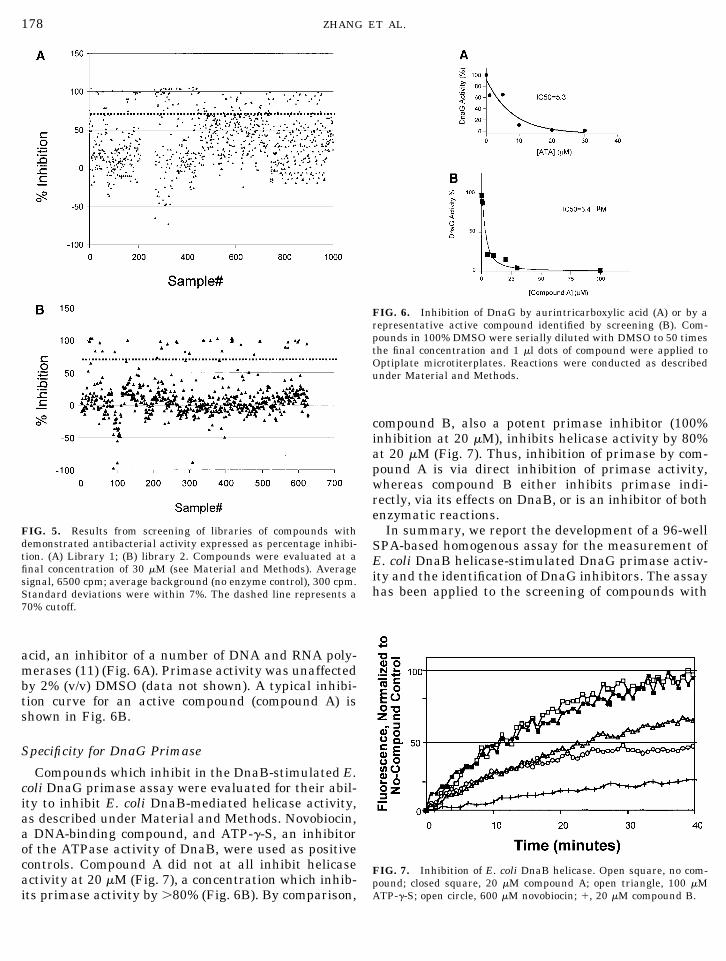
acid, an inhibitor of a number of DNA and RNA poly-merases (11) (Fig. 6A). Primase activity was unaffectedby 2% (v/v) DMSO (data not shown). A typical inhibi-tion curve for an active compound (compound A) isshown in Fig. 6B.
Specificity for DnaG Primase
Compounds which inhibit in the DnaB-stimulated E.coli DnaG primase assay were evaluated for their abil-ity to inhibit E. coli DnaB-mediated helicase activity,as described under Material and Methods. Novobiocin,a DNA-binding compound, and ATP-�-S, an inhibitorof the ATPase activity of DnaB, were used as positivecontrols. Compound A did not at all inhibit helicaseactivity at 20 �M (Fig. 7), a concentration which inhib-its primase activity by �80% (Fig. 6B). By comparison,
compound B, also a potent primase inhibitor (100%inhibition at 20 �M), inhibits helicase activity by 80%at 20 �M (Fig. 7). Thus, inhibition of primase by com-pound A is via direct inhibition of primase activity,whereas compound B either inhibits primase indi-rectly, via its effects on DnaB, or is an inhibitor of bothenzymatic reactions.
In summary, we report the development of a 96-wellSPA-based homogenous assay for the measurement ofE. coli DnaB helicase-stimulated DnaG primase activ-ity and the identification of DnaG inhibitors. The assayhas been applied to the screening of compounds with
FIG. 5. Results from screening of libraries of compounds withdemonstrated antibacterial activity expressed as percentage inhibi-tion. (A) Library 1; (B) library 2. Compounds were evaluated at afinal concentration of 30 �M (see Material and Methods). Averagesignal, 6500 cpm; average background (no enzyme control), 300 cpm.Standard deviations were within 7%. The dashed line represents a70% cutoff.
FIG. 6. Inhibition of DnaG by aurintricarboxylic acid (A) or by arepresentative active compound identified by screening (B). Com-pounds in 100% DMSO were serially diluted with DMSO to 50 timesthe final concentration and 1 �l dots of compound were applied toOptiplate microtiterplates. Reactions were conducted as describedunder Material and Methods.
FIG. 7. Inhibition of E. coli DnaB helicase. Open square, no com-pound; closed square, 20 �M compound A; open triangle, 100 �MATP-�-S; open circle, 600 �M novobiocin; �, 20 �M compound B.
178 ZHANG ET AL.

demonstrated antibacterial activity and has identifiedcandidates which inhibit DnaG activity in vitro. Inprinciple, inhibitors may block or alter the interactionof DnaG with DnaB. Thus, further characterization ofinhibitors by secondary assays which monitor DnaG orDnaB activities independently of one another is re-quired to establish mechanism of action. In the case ofDnaG this is conveniently monitored by performingreactions in the absence of DnaB (Fig. 2) with or with-out extension by Klenow polymerase and/or by evalu-ating the effect of such inhibitors on the helicase activ-ity of E. coli DnaB. The latter approach has been usedto differentiate primase inhibitors from dual primase/helicase inhibitors (Figs. 6B and 7) and serves as thestarting point for more detailed characterization ofsuch inhibitors.
Because the product of DnaG activity is a heterodu-plex RNA:DNA hybrid, the opportunity exists for mea-suring the combined activities of DnaG, DnaB, and aDNA-dependent DNA polymerase. This has been dem-onstrated by the use of Klenow in a linked assay inwhich the radiolabel is provided as a [3H]dNTP. Otheraccessory replication proteins involved in the process ofboth leading- and lagging-strand DNA synthesis mayalso be incorporated into this system, thus furtherincreasing its usefulness for the identification of antag-onists of the DNA replication machine. Last, most scin-tillation proximity assays monitoring polymerase ac-tivity use biotinylated nucleotide substrates (13, 14),followed by capture onto streptavidin-coated SPAbeads. In this assay unmodified substrates may beused, eliminating any confounding factors associatedwith using nonnatural substrates.
ACKNOWLEDGMENTS
We gratefully acknowledge the assistance of Ms. Jennifer Masonfor suggestions for assay development and Drs. Mark Harpel andRobert Copeland for critical reading of the manuscript.
REFERENCES
1. Daly, J. S., Giehl, T. J., Brown, N. C., Zhi, C., Wright, G. E., andEllison, R. T. 3rd. (2000) Antimicrob. Agents Chemother. 44(8),2217–2222.
2. Lu, Y.-B., Ratnakar, P. V. A. L., Mohanty, B. K., and Bastia, D.(1996) Direct physical interaction between DnaG primase andDnaB helicase of E. coli is necessary for optimal synthesis ofprimer RNA. Proc. Natl. Acad. Sci. USA 93, 12902–12907.
3. Tougu, K., Peng, H., and Marians, K. J. (1994) Identification of adomain of Escherichia coli primase required for functional inter-action with the DnaB helicase at the replication fork. J. Biol.Chem. 269, 4675–4682.
4. Wu, C. A., Zechner, E. L., Reems, J. A., McHenry, C. S., andMarians, K. J. (1992) Coordinated leading- and lagging-strandsynthesis at the Escherichia coli DNA replication fork. V. Pri-mase action regulates the cycle of Okazaki fragment synthesis.J. Biol. Chem. 267, 4074–4083.
5. Kim, S., Dallmann, H. G., McHenry, C. S., and Marians, K. J.(1996) Tau couples the leading- and lagging-strand polymerasesat the Escherichia coli DNA replication fork. J. Biol. Chem. 271,21406–21412.
6. Arai, K., and Kornberg, A. (1981) Mechanism of dnaB proteinaction. IV. General priming of DNA replication by dnaB proteinand primase compared with RNA polymerase. J. Biol. Chem.256, 5267–5272.
7. Arai, K., and Kornberg, A. (1979) A general priming systememploying only dnaB protein and primase for DNA replication.Proc. Natl. Acad. Sci. USA 76, 4308–4312.
8. Marians, K. J. (1995) PhiX174-type primosomal proteins: Puri-fication and assay. Methods Enzymol. 262, 512–516.
9. Sambrook, Fritsch, and Maniatis. (1989) Molecular Cloning: ALaboratory Manual, 2nd ed., Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
10. Hart, H. E., and Greenwald, E. B. (1979) Scintillation Proximityassay (SPA)—A new method of immunoassay: Direct and inhi-bition mode detection with human albumin and rabbit antihu-man alabumin. Mol. Immunol. 16, 265–267.
11. Swart, J. R., and Griep, M. A. (1993) Primase from Escherichiacoli primes single-stranded templates in the absence of single-stranded DNA-binding protein or other auxiliary proteins: Tem-plate sequence requirements based on the bacteriophage G4complementary strand origin and Okazaki fragment initiationsites. J. Biol. Chem. 268, 12970–12976.
12. Nakane, H., Balzarini, J., De Clercq, E., and Ono, K. (1988)Differential inhibition of various deoxyribonucleic acid poly-merases by Evans blue and aurintricarboxylic acid. Eur. J. Bio-chem. 177, 91–96.
13. Zheng, W., Carroll, S. S., Inglese, J., Graves, R., Howells, L., andStrulovici, B. (2001) Miniaturization of a Hepatitis C Virus RNApolymerase assay using a �102. degree Colled CCD Camera-Based Imaging System. Anal. Biochem. 290, 214–220.
14. Liu, J., Feldman, P. A., Lippy, J. S., Bobkova, E., Kurilla, M. G.,and Chung, T. D. (2001) A scintillation proximity assay for RNAdetection. Anal. Biochem. 289(2), 239–245.
179DnaG PRIMASE SCINTILLATION PROXIMITY ASSAY
















![Replicación del DNA - [DePa] Departamento de Programas ...depa.fquim.unam.mx/amyd/archivero/Replicacion_9218.pdf · 1) Nucleasa + DNA Ori 2) pp dna A + DNA Ori 3) pp dnaB + DNA Ori](https://img.pdfslide.us/doc/110x75/5c16883109d3f29f108cbfaf/replicacion-del-dna-depa-departamento-de-programas-depafquimunammxamydarchiveroreplicacion9218pdf.jpg)









![Rif1 Supports the Function of the CST Complex in Yeast ... · cell cycle arrest [19–21]. Interestingly, Stn1 interacts with Pol12 [22], a subunit of the DNA polymerase a (pola)-primase](https://img.pdfslide.us/doc/110x75/608d5e2b94e36f65cb565cd0/rif1-supports-the-function-of-the-cst-complex-in-yeast-cell-cycle-arrest-19a21.jpg)


