Embed Size (px)
Citation preview

Original Article
Histone chaperone ASF1 is involved in gene transcriptionactivation in response to heat stress in Arabidopsis thaliana
Minjie Weng1,†, Yue Yang1,†, Haiyang Feng1, Zongde Pan1, Wen-Hui Shen1,2, Yan Zhu1 & Aiwu Dong1
1State Key Laboratory of Genetic Engineering, International Associated Laboratory of CNRS-Fudan-HUNAU on PlantEpigenome Research, Department of Biochemistry, Institute of Plant Biology, School of Life Sciences, Fudan University,Shanghai 20043, China and 2Institut de Biologie Moléculaire des Plantes du CNRS, Université de Strasbourg, 12 rue du GénéralZimmer, 67084 Strasbourg Cédex, France
ABSTRACT
ANTI-SILENCING FUNCTION 1 (ASF1) is an evolution-arily conserved histone chaperone involved in diversechromatin-based processes in eukaryotes. Yet, its role in tran-scription and the underlying molecular mechanisms remainlargely elusive, particularly in plants. Here, we show that theArabidopsis thaliana ASF1 homologous genes, AtASF1Aand AtASF1B, are involved in gene transcription activationin response to heat stress. The Atasf1ab mutant displaysdefective basal as well as acquired thermotolerance pheno-types. Heat-induced expression of several key genes, includ-ing the HEAT SHOCK PROTEIN (HSP) genes Hsp101,Hsp70, Hsa32, Hsp17.6A and Hsp17.6B-CI, and the HEATSHOCK FACTOR (HSF) gene HsfA2 but not HsfB1 is dras-tically impaired in Atasf1ab as compared with that in wildtype. We found that AtASF1A/B proteins are recruited ontochromatin, and their enrichment is correlated withnucleosome removal and RNA polymerase II accumulationat the promoter and coding regions of HsfA2 and Hsa32 butnot HsfB1. Moreover, AtASF1A/B facilitate H3K56acetylation (H3K56ac), which is associated with HsfA2 andHsa32 activation. Taken together, our study unravels animportant function of AtASF1A/B in plant heat stressresponse and suggests that AtASF1A/B participate in tran-scription activation of some but not all HSF and HSP genesvia nucleosome removal and H3K56ac stimulation.
Key-words: ANTI-SILENCING FUNCTION 1; transcrip-tion regulation.
INTRODUCTION
Nucleosome, the basic unit of chromatin, is composed ofroughly 147 base pairs (bp) of DNA wrapped around theglobular histone octamer comprising one centre-positioned(H3-H4)2 tetramer and two side-positioned H2A-H2Bdimers. Histone chaperones facilitate histone–DNA interac-tions during nucleosome assembly and/or disassembly, and
play crucial roles in chromatin dynamics relevant to DNAreplication, repair, recombination and transcription(Avvakumov et al. 2011).ANTI-SILENCING FUNCTION 1(ASF1) is a highly conserved histone chaperone specific forH3-H4, and its monomeric state is detected in the form ofASF1-H3-H4 ternary complex, which is mutually exclusivefrom that of (H3-H4)2 tetramer (English et al. 2006; Natsumeet al. 2007). This suggests that loading of ASF1 likely has an(H3-H4)2 tetramer-disrupting activity in the context ofnucleosomes. ASF1 also interacts with other chromatinfactors, such as CHROMATIN ASSEMBLY FACTOR-1(CAF-1) and HISTONE REGULATORY HOMOLOGA (HIRA), which mediate replication-dependent andreplication-independent H3 incorporation into chromatin,respectively (Tagami et al. 2004).
Several studies have also revealed critical importance ofASF1 in establishment of H3K56 acetylation (H3K56ac;reviewed in Avvakumov et al. 2011). K56 is located withinN-terminal α-helix of H3, and its acetylation requires ASF1for catalysis by histone acetyltransferase Rtt109 and CBP/p300 in yeast and animal cells, respectively (Schneider et al.2006; Das et al. 2009; Avvakumov et al. 2011). Adding acetylinterrupts histone–DNA contact and increases rate ofnucleosomal DNA ends unwrapping from histone octamer(Neumann et al. 2009), and promotes nucleosome disassem-bly during transcriptional activation in yeast (Rufiange et al.2007; Williams et al. 2008; Lin & Schultz 2011).
The Arabidopsis thaliana genome comprises two ASF1homologous genes, AtASF1A and AtASF1B, which playredundant roles and participate in S-phase replication-dependent chromatin assembly (Zhu et al. 2011).The loss-of-function Atasf1ab double mutant displays chromatininstability and DNA damage as well as activation of expres-sion of cell cycle checkpoint and DNA repair genes (Zhuet al. 2011). More recently, it was shown that AtASF1A andAtASF1B are involved in UV-induced DNA damage repair,likely through interaction with the TIP60-like histoneacetyltransferases HAM1 and HAM2 (Lario et al. 2013). Inthe present study, we report a crucial function of AtASF1Aand AtASF1B in plant heat stress tolerance.
Heat stress is expecting to occur more frequently as aconsequence of warming climate, which seriously challengesecological plant species demography and agricultural crop
Correspondence: Y. Zhu. Fax: +86 21 55665673; e-mail: [email protected]; A. Dong. Fax: +86 21 55665673; e-mail: [email protected]
†These authors contributed equally to this work.
Plant, Cell and Environment (2014) 37, 2128–2138 doi: 10.1111/pce.12299
bs_bs_banner
© 2014 John Wiley & Sons Ltd2128

yield (Saidi et al. 2011; Qu et al. 2013). During evolution,plants have evolved various molecular and physiologicalmechanisms to cope with heat stress. Among others, HEATSHOCK FACTOR (HSF) and HEAT SHOCK PROTEIN(HSP) play a central role in the onset of cellularthermotolerance (Saidi et al. 2011; Qu et al. 2013). TheArabidopsis HSF family comprises over 20 members, whichcan be grouped into A, B and C classes based on proteinstructural domain organization (Nover et al. 2001). The Aclass members contain a C-terminal AHA domain and mostact as transcription activators. Although HsfA1a/b are notheat induced and function as immediate activators in heatresponse (Lohmann et al. 2004; Busch et al. 2005), HsfA2 isthe most potently heat-induced gene and is responsible foramplification and long-term maintenance of heat response(Charng et al. 2007). The B class members, for example,HsfB1 and HsfB2b, possess a tetrapeptide LFGV motif andgenerally function as transcriptional repressors (Ikeda et al.2011). Function of the unique C class member HsfC1 remainsunknown so far. At downstream of HSFs, HSPs function asmolecular chaperones involved in maintenance and/or resto-ration of protein homeostasis under heat stress, and aregrouped into Hsp100, Hsp90, Hsp70, Hsp60 and small Hspsubgroups (Swindell et al. 2007). Intriguingly, some Hsps (e.g.Hsp90, Hsp70) also participate in the feedback control ofsubcellular compartmentalization and activity of plant HSFs(reviewed in Scharf et al. 2012). Detailed genetic and physio-logical analyses have implicated HSFs and HSPs in the regu-lation of plant basal thermotolerance upon direct exposureto a heat stress. Meanwhile, exposure of plants to a moderatelevel of high temperature (e.g. 37 °C for Arabidopsis) canefficiently activate the HSF-HSP gene networks, which sig-nificantly improves plant thermotolerance to a second moresevere heat stress (e.g. 45 °C for Arabidopsis; reviewed inSaidi et al. 2011; Mittler et al. 2012). This process is generallyreferred as acclimation or ‘priming’, and the enhanced plantability is referred as acquired thermotolerance.
Chromatin remodelling is believed to be advantageousin regulation of a large set of genes and in gene primingin plant response to stresses (Berr et al. 2012; Zhu et al.2012). The histone H2A variant, H2A.Z, was proposedas a thermosensor and was shown to be evicted fromnucleosomes at Hsp70 and other genes following exposure ofArabidopsis plants to a moderately warm temperature(27 °C; Kumar & Wigge 2010). The arp6 mutant defective inH2A.Z incorporation showed constitutive up-regulation ofHsp70 and a genome-wide expression pattern mimickingwild-type (WT) plants grown at warm temperatures(Kumar & Wigge 2010). In Brachypodium distachyon,ARP6-mediated H2A.Z incorporation was reported to play animportant function in thermal stress response on grain yield(Boden et al. 2013). Long heat stress, 15–30 h at 37 °C or 48 hat 42 °C, was shown to release silencing of a reportertransgene as well as some endogenous heterochromatin lociin Arabidopsis (Lang-Mladek et al. 2010; Pecinka et al. 2010;Tittel-Elmer et al. 2010). The silencing release was not asso-ciated with any detectable changes of DNA methylationor histone methylation and mutants of several chromatin
regulators, for example, CMT3, DDM1, DRM2, MOM1 andKYP, did not show any detectable perturbation in the heatstress response (Pecinka et al. 2010; Tittel-Elmer et al. 2010).
In this study, we investigate the function of AtASF1A andAtASF1B in heat stress response. We show that the loss-of-function Atasf1ab double mutant exhibits both basal andacquired thermotolerance defects associated with impairedtranscription activation of some HSF and HSP genes. Weprovide evidence highlighting a crucial role of AtASF1A/Bin heat stress gene activation via nucleosome removal andH3K56ac stimulation.
MATERIALS AND METHODS
Plant material and growth conditions
Atasf1a, Atasf1b and Atasf1ab mutants have been describedpreviously (Zhu et al. 2011). Seeds were produced in growthchamber at 22 °C in a 16 h light/8 h dark photoperiod.
Transcriptome analysis
Twelve-day-old seedlings of Atasf1a, Atasf1b, Atasf1ab andWT grown on agar-solidified MS medium M0255 (Duchefa;http://www.duchefa.com) at 22 °C under a light intensityof about 100 μEm−2 s−1 in a 16 h light/8 h dark photoperiodwere used in transcriptome analysis. RNA was extractedusing the TRIzol kit according to the manufacturer’s instruc-tions (Invitrogen, Carlsbad, CA, USA). Two biological repli-cates were analysed for gene expression using AgilentArabidopsis4*44K (Palo Alto, CA, USA) oligonucleotidearray containing 43603 probes (Shanghai BiotechnologyCorporation, Pudong, Shanghai, China). The raw data ofmicroarray have been deposited in public database NCBI-GEO (GSE48854).Data analysis was performed by the onlineinformation analysis platform based on the statistic package R(http://sas.ebioservice.com). Genes with expression changedmore than twofold between WT, and the mutants are consid-ered as differentially expressed.Gene ontology (GO) analysiswas performed as previously described (Liu et al. 2009b).
Heat and oxidative stress treatments
For basal thermotolerance assay, 30 WT and 30 Atasf1abseeds were germinated on a same plate containing agar-solidified MS medium. Each determination was performedwith three replicate plates. The plates were incubated at22 °C for 30 h, placed at 45 °C for 2 h and returned to 22 °Cfor 10 d before examination of heat effects. For acquiredthermotolerance assay, the seed plates were incubated at22 °C for 10 d, treated at 37 °C for 1.5 h, returned to 22 °Cfor 2 h (2H) or 2 d (2D) for recovery, treated at 45 °C for 2 hand finally returned to 22 °C for 6 d before examination ofheat effects.
Heat effects on H2O2 production was detected in the roots(from 30 plants per sample) by fluorescent dye 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA) stainingand visualization under fluorescence microscopy as described
AtASF1 in heat stress response 2129
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

previously (Kim et al. 2012). For oxidative stress treatment,over 200 12-day-old seedlings per sample were transferredto the liquid MS medium M0255 without (control) or with20 μm Rose Bengal (RB) (R3877; Sigma, St Louis, MO,USA), and grown at 22 °C under 100 μEm−2 s−1 light.Samples were collected at described time points and ana-lysed for gene expression.
Chlorophylls and ion leakage measurements
Total chlorophylls were measured by non-macerationmethods. Seedlings (100 mg per sample) were incubated in5 mL dimethyl sulfoxide (DMSO) at 65 °C for 4 h in thedark. Absorbance was recorded at 645, 663, 470 nm in aSmartSpecTM Plus Spectrophotometer (170–2525; Bio-Rad,Hercules, CA, USA). Chlorophyll and carotenoid contentswere calculated as described previously (Chauhan et al.2012). Ten-day-old seedlings (15 plants per sample) wereused for the ion leakage measurement after different heattreatments according to Hong et al. (2003). Briefly, seedlingswere incubated in deionized water for 1 d, and the resultingsolution was measured for conductivity. Values were normal-ized to the total conductivity measured on dead seedlingsafter autoclaving, and the resulted percentage represents therelative ion leakage activity of living seedlings.
RT-PCR and Western blot analysis
RNA isolation and RT-PCR were performed as describedpreviously (Liu et al. 2009a), and the primers for RT-PCRwere listed in the Supporting Information Table S1. Totalproteins were extracted from the 12-day-old seedlings beforeand after heat treatment at 37 °C for 1.5 h and analysed byWestern blot using anti-Hsp101 (ab80123; Abcam, Cam-bridge, UK) and anti-Hsp17.6 (ab80183; Abcam) antibodies.Histones were prepared as described previously (Gao et al.2012) and analysed in Western blot using anti-H3 (ab1791;Abcam) and anti-H3K56ac (07–677; Millipore, Billerica, MA,USA) antibodies.
Chromatin immunoprecipitation (ChIP) analysis
About 2 g per sample of 12-day-old seedlings untreated ortreated by incubation at 37 °C for 1.5 h was used for ChIPanalysis. ChIP experiments were performed as described pre-viously (Johnson et al. 2002). The anti-AtASF1 rabbitpolyclonal antiserum was generated against AtASF1Apeptide (N-terminal 159 aa) through a commercial service byAbmart, Inc (http://www.abmart.cn). Other antibodies usedin ChIP are anti-H3 (ab1791; Abcam), anti-H3K56ac (07–677; Millipore) and anti-Pol II (sc-900; Santa Cruz Biotech-nology, Santa Cruz, CA, USA). UBC28 was used as aninternal reference gene (Pecinka et al. 2009). The gene-specific primers used in ChIP are listed in the SupportingInformation Table S1.
In vitro histone acetyltransferase (HAT) assay
The open reading frame (ORF) of yeast Rtt109 wascloned into the expression vector pGEX-4T1 (Amersham,
Piscataway, NJ, USA) using BamHI and SalI sites. The ORFof AtASF1A was cloned into the expression vector pET-14b(Novagen, Madison, WI, USA) using BglII and XhoI sites.Recombinant proteins were expressed in Escherichia coliand purified according to a previously described procedure(Dong et al. 2005). In vitro HAT assay was performed asdescribed previously (Driscoll et al. 2007) using mixturehistones (223565; Roche, Indianapolis, IN, USA) as substratesand anti-H3K56ac antibody (07–677; Millipore) in Westernblot detection.
RESULTS
Transcriptome analysis reveals a large numberof misregulated genes in the Atasf1ab mutant
To investigate AtASF1A/B function in genome transcriptionregulation, we performed transcriptome analysis usingAgilent Arabidopsis 44K oligonucleotide microarray on12-day-old seedlings of WT and the double-mutant Atasf1abas well as the single mutants Atasf1a and Atasf1b (Zhuet al. 2011). Compared with WT, Atasf1ab showed 695 up-regulated genes and 1204 down-regulated genes whereasAtasf1a and Atasf1b only showed small numbers of per-turbed genes (Fig. 1a; Supporting Information Tables S2–S7),which is in agreement with the redundant function ofAtASF1A and AtASF1B. As expected, AtASF1A andAtASF1B were among the down-regulated genes found inthe respective mutants (Supporting Information Tables S5–S7).The perturbed genes in Atasf1ab are distributed along allthe five Arabidopsis chromosomes without any obvious pref-erential regions (Supporting Information Fig. S1). GO analy-sis revealed that genes involved in response to variousstimuli, including heat and oxidative stress, are over-represented in the down-regulation category of genes inAtasf1ab (Fig. 1b). RT-PCR analysis confirmed that severalheat stress genes, for example, HsfA2, Hsp101, Hsp70, Hsa32,Hsp17.6A and Hsp17.6B-CI, are down-regulated in Atasf1ab(Supporting Information Fig. S2). Hereinafter, we focus onplant heat stress response to reveal AtASF1A/B functions.
Atasf1ab mutant plants show defects in bothbasal and acquired thermotolerances
Thermotolerances of Atasf1ab mutant were assessed viawell-established heat stress phenotypic assays (Yeh et al.2012). We germinated the mutant and WT seeds on the sameplate at standard growth chamber temperature (22 °C) for30 h, heat treated at 45 °C for 2 h and then incubated at 22 °Cfor 10 d. As shown in Fig. 2a, about 100% of WT seedlingssurvived from the heat treatment whereas only about 40%of Atasf1ab seedlings could be recovered from the sameheat treatment (n > 200). To assess Atasf1ab in acquiredthermotolerance, 10-day-old mutant and WT seedlings wereacclimated by incubation at 37 °C for 1.5 h followed by arecovery at 22 °C for 2 h (R-2H) or 2 d (R-2D), and thentreated by heat stress at 45 °C for 2 h followed by additional6 d of growth. We found that both WT and Atasf1ab plants
2130 M. Weng et al.
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

grow better with the R-2H than with the R-2D recoverytreatment condition (Fig. 2b). Nonetheless, Atasf1ab com-pared with WT showed more severe growth inhibition ateither R-2H or R-2D recovery condition, and the mutant butnot WT seedlings displayed an obvious chlorotic phenotype(Fig. 2b). Chlorophyll content analysis confirmed that chlo-rophyll a/b and carotenoid levels are drastically reduced inAtasf1ab compared with WT (Fig. 2c).
It is known that heat stress can change cell membranepermeability by affecting membrane transporter activitiesand membrane integrity (Hong et al. 2003). We investigatedcell membrane permeability by ion leakage measurementusing 10-day-old seedlings. Untreated (control) or acclima-tion temperature-treated (37 °C, 1.5 h) seedlings showed lowvalues of ion leakage. Nevertheless, a statistically significantincrease of ion leakage was detected in Atasf1ab comparedwith WT (Fig. 2d). The severe heat stress treatment (45 °C,2 h) drastically induced ion leakage, which likely exceeds thenormal membrane function capacity and in this case nodifference between Atasf1ab and WT could be detected.
Remarkably, acclimation greatly ameliorated plant toleranceand lower values of ion leakage were detected after thesevere heat stress treatment. The R-2H recovery conditionwas superior over the R-2D condition in ion leakage reduc-tion (Fig. 2d), which is consistent with the observed betterrecovery plant growth phenotype (Fig. 2b). At both R-2Hand R-2D conditions, Atasf1ab showed higher ion leakagethan WT did (Fig. 2d).
Collectively, our plant growth and physiological analysisdata clearly establish that both basal and acquiredthermotolerances are impaired in the Atasf1ab mutant.
Activation of heat-induced genes is impaired inAtasf1ab mutant plants
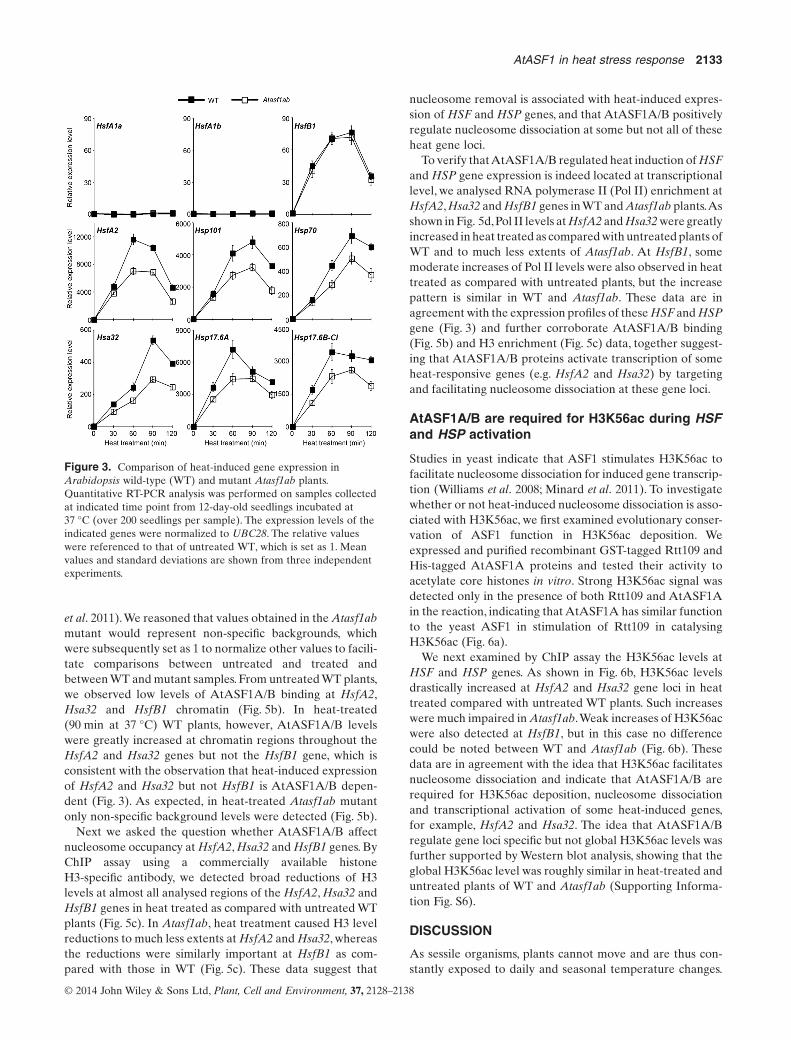
Most of HSF and HSP genes are expressed at relatively lowlevels in plants grown at 22 °C. Perturbed expression of HSFand HSP genes in Atasf1ab identified by our transcriptomeanalysis using plants grown in the standard growth chambermay not be necessarily temperature related. To test whetherAtASF1A/B have a role in heat-related gene expression, weanalysed expression levels of HSF and HSP genes in Atasf1aband WT during a time course of 120 min incubation of plants at37 °C (Fig. 3).We found that expression of HsfA1a/b remainedlargely unchanged, whereas HsfB1 expression was induced upto 75 folds,peaked at 60 and 90 min after the heat treatment.Forthese three genes,no significant differences of expression couldbe detected between Atasf1ab and WT. In contrast, the otherexamined genes, for example, HsfA2, Hsp101, Hsp70, Hsa32,Hsp17.6A and Hsp17.6B-CI, showed varied greater folds(several hundred to more than 10 000 at peak values) of heatinduction, and their expression levels were overall drasticallylower in Atasf1ab than in WT following the heat treatment(Fig. 3).The expression level of AtASF1A/B was not affected byheat treatment in WT (Supporting Information Fig. S3). OurWestern blot analyses further confirmed that the Hsp101 andHsp17.6 protein levels are drastically induced by the heat treat-ment and that they were lower in Atasf1ab than in WT(Supporting Information Fig. S4). Taken together, these dataindicate that AtASF1A/B are required for a maximal heat-induced expression of HsfA2,Hsp101,Hsp70,Hsa32,Hsp17.6Aand Hsp17.6B-CI, but not HsfB1.
AtASF1A/B are also required for maximalactivation of HSF and HSP genes in response tooxidative stress
In agreement with previous reports (Volkov et al. 2006;Konigshofer et al. 2008), heat treatment induced hydrogenperoxide (H2O2) production (Fig. 4a),which was visualized byfluorescent dye H2DCFDA staining of roots (Kim et al. 2012).In this assay, we did not notice any significant differencebetween Atasf1ab and WT. We next asked the questionwhether Atasf1ab affects HSF and HSP gene expression inresponse to oxidative stress.We tested this by treating 12-day-oldWT and Atasf1ab plants grown under light with the chemi-cal RB, a photosensitizer-producing oxidative stress (Fischeret al. 2004). We found that RB treatments efficiently induced
(a)
(b)
Figure 1. Comparison of the differentially expressed genes in theArabidopsis Atasf1a, Atasf1b and Atasf1ab mutants. (a) Venndiagrams show the number and overlap of significantly up- anddown-regulated genes in the Atasf1a, Atasf1b and Atasf1abmutants compared with wild type. (b) BiNGO analysis ondown-regulated genes in the Atasf1ab mutant. As indicated bycolour scale bar, the yellow to orange colour of the circlescorresponds to the level of significance of over-represented geneontology category according to a multiple t-test with falsediscovery rate-corrected P-value under 0.05. Circle size isproportional to number of genes in the category.
AtASF1 in heat stress response 2131
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

the expression of Hsp17.6A, HsfA2, Hsp17.6B-CI, Hsp101,Hsp70 and to a less extent Hsa32 (Fig. 4b). Compared withWT, Atasf1ab showed much reduced induction of these genesat most of the examined time points following the RB treat-ment (Fig. 4b). These data suggest that heat may induce HSFand HSP gene expression partly through signalling of oxida-tive stress and that AtASF1A/B are involved at the signaltransduction terminal transcription.
AtASF1 binding and nucleosome occupancy atHSF and HSP genes
To investigate molecular basis of AtASF1A/B func-tion in transcription, we first asked the question whether
AtASF1A/B proteins bind chromatin at HSF and HSP genes.We generated polyclonal antibodies against the N-terminal159 amino acids peptide of AtASF1A. In Western blotassay, the antibody specifically recognized the YFP-fusedAtASF1A (Zhu et al. 2011) but not NRP1, another histonechaperone (Zhu et al. 2006), from total protein extracts oftransgenic plants (Supporting Information Fig. S5). Becauseof highly conserved protein sequences between AtASF1Aand AtASF1B (Zhu et al. 2011), this antibody likely can alsorecognize the AtASF1B protein. By ChIP assay using thisantibody, we analysed AtASF1A/B protein enrichment atdiverse regions of HsfA2, Hsa32 and HsfB1 (Fig. 5a). Thesethree genes are all necessary for acclimation-inducedacquired thermotolerance (Charng et al. 2006, 2007; Ikeda
(a) (b)
(c) (d)
Figure 2. Comparison of basal and acquired thermotolerance between Arabidopsis wild-type (WT) and mutant Atasf1ab plants.(a) Comparison of plant basal thermotolerance. For heat treatment, 30-hour-old seedlings were incubated at 45 °C for 2 h, recovered andgrown at 22 °C for 10 d before photographing. Untreated plants grown at 22 °C were shown as controls. (b) Comparison of plant acquiredthermotolerance. For heat treatment, 10-day-old seedlings were first treated at 37 °C for 1.5 h and recovered at 22 °C for 2 h (R-2H) or 2 d(R-2D), and then treated at 45 °C for 2 h, recovered and grown at 22 °C for 6 d before photographing. Untreated plants grown at 22 °C wereshown as controls. (c) Heat bleaching effect as measured by reduction levels of three major chlorophylls, chl a, chl b and carotenoids.Extracts from heat acclimated (R-2H) and untreated control seedlings described in (b) were analysed. The percentage of treated comparedwith untreated samples was shown as relative levels. The mean values and standard deviations from three independent experiments areshown. Significant difference at P < 0.05 between WT and Atasf1ab mutant is indicated by asterisk (*). (d) Heat effects measured by ionleakage. The mean value and standard deviation represent the results of three independent experiments. Significant difference at P < 0.05between WT and Atasf1ab mutant is indicated by asterisk (*).
2132 M. Weng et al.
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138
et al. 2011).We reasoned that values obtained in the Atasf1abmutant would represent non-specific backgrounds, whichwere subsequently set as 1 to normalize other values to facili-tate comparisons between untreated and treated andbetween WT and mutant samples. From untreated WT plants,we observed low levels of AtASF1A/B binding at HsfA2,Hsa32 and HsfB1 chromatin (Fig. 5b). In heat-treated(90 min at 37 °C) WT plants, however, AtASF1A/B levelswere greatly increased at chromatin regions throughout theHsfA2 and Hsa32 genes but not the HsfB1 gene, which isconsistent with the observation that heat-induced expressionof HsfA2 and Hsa32 but not HsfB1 is AtASF1A/B depen-dent (Fig. 3). As expected, in heat-treated Atasf1ab mutantonly non-specific background levels were detected (Fig. 5b).
Next we asked the question whether AtASF1A/B affectnucleosome occupancy at HsfA2, Hsa32 and HsfB1 genes. ByChIP assay using a commercially available histoneH3-specific antibody, we detected broad reductions of H3levels at almost all analysed regions of the HsfA2, Hsa32 andHsfB1 genes in heat treated as compared with untreated WTplants (Fig. 5c). In Atasf1ab, heat treatment caused H3 levelreductions to much less extents at HsfA2 and Hsa32, whereasthe reductions were similarly important at HsfB1 as com-pared with those in WT (Fig. 5c). These data suggest that
nucleosome removal is associated with heat-induced expres-sion of HSF and HSP genes, and that AtASF1A/B positivelyregulate nucleosome dissociation at some but not all of theseheat gene loci.
To verify thatAtASF1A/B regulated heat induction of HSFand HSP gene expression is indeed located at transcriptionallevel, we analysed RNA polymerase II (Pol II) enrichment atHsfA2, Hsa32 and HsfB1 genes in WT and Atasf1ab plants.Asshown in Fig. 5d,Pol II levels at HsfA2 and Hsa32 were greatlyincreased in heat treated as compared with untreated plants ofWT and to much less extents of Atasf1ab. At HsfB1, somemoderate increases of Pol II levels were also observed in heattreated as compared with untreated plants, but the increasepattern is similar in WT and Atasf1ab. These data are inagreement with the expression profiles of these HSF and HSPgene (Fig. 3) and further corroborate AtASF1A/B binding(Fig. 5b) and H3 enrichment (Fig. 5c) data, together suggest-ing that AtASF1A/B proteins activate transcription of someheat-responsive genes (e.g. HsfA2 and Hsa32) by targetingand facilitating nucleosome dissociation at these gene loci.
AtASF1A/B are required for H3K56ac during HSFand HSP activation
Studies in yeast indicate that ASF1 stimulates H3K56ac tofacilitate nucleosome dissociation for induced gene transcrip-tion (Williams et al. 2008; Minard et al. 2011). To investigatewhether or not heat-induced nucleosome dissociation is asso-ciated with H3K56ac, we first examined evolutionary conser-vation of ASF1 function in H3K56ac deposition. Weexpressed and purified recombinant GST-tagged Rtt109 andHis-tagged AtASF1A proteins and tested their activity toacetylate core histones in vitro. Strong H3K56ac signal wasdetected only in the presence of both Rtt109 and AtASF1Ain the reaction, indicating that AtASF1A has similar functionto the yeast ASF1 in stimulation of Rtt109 in catalysingH3K56ac (Fig. 6a).
We next examined by ChIP assay the H3K56ac levels atHSF and HSP genes. As shown in Fig. 6b, H3K56ac levelsdrastically increased at HsfA2 and Hsa32 gene loci in heattreated compared with untreated WT plants. Such increaseswere much impaired in Atasf1ab.Weak increases of H3K56acwere also detected at HsfB1, but in this case no differencecould be noted between WT and Atasf1ab (Fig. 6b). Thesedata are in agreement with the idea that H3K56ac facilitatesnucleosome dissociation and indicate that AtASF1A/B arerequired for H3K56ac deposition, nucleosome dissociationand transcriptional activation of some heat-induced genes,for example, HsfA2 and Hsa32. The idea that AtASF1A/Bregulate gene loci specific but not global H3K56ac levels wasfurther supported by Western blot analysis, showing that theglobal H3K56ac level was roughly similar in heat-treated anduntreated plants of WT and Atasf1ab (Supporting Informa-tion Fig. S6).
DISCUSSION
As sessile organisms, plants cannot move and are thus con-stantly exposed to daily and seasonal temperature changes.
Figure 3. Comparison of heat-induced gene expression inArabidopsis wild-type (WT) and mutant Atasf1ab plants.Quantitative RT-PCR analysis was performed on samples collectedat indicated time point from 12-day-old seedlings incubated at37 °C (over 200 seedlings per sample). The expression levels of theindicated genes were normalized to UBC28. The relative valueswere referenced to that of untreated WT, which is set as 1. Meanvalues and standard deviations are shown from three independentexperiments.
AtASF1 in heat stress response 2133
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

Heat stress can affect nucleosome composition and stability,and greatly impacts plant genome function (Kumar & Wigge2010; Lang-Mladek et al. 2010; Pecinka et al. 2010; Tittel-Elmer et al. 2010; Boden et al. 2013).Therefore, identificationof chromatin regulators involved in heat stress response hascrucial importance in understanding plant adaptationto environments. In this study, we found that the histonechaperone AtASF1 plays important roles in plant thermotol-erance, in HSF and HSP gene transcription, and in nucleo-some removal and H3K56ac enrichment at some HSF andHSP genes.
Our phenotypic and physiological analyses clearly showedthat the Atasf1ab mutant plants have reduced basal andacquired thermotolerance. This differs from the absence ofheat stress growth phenotype of the mutants deprived ofregulators involved in DNA methylation (CMT3, DRM2)or chromatin remodelling (DDM1,MOM1) or histone methy-lation (KYP, SUVH2) (Pecinka et al. 2010; Tittel-Elmer et al.2010;Popova et al. 2013).Reduced basal thermotolerance wasreported in the more recent study for several mutants relatedto RNA-directed DNA methylation (RdDM) pathway, forexample,nrpd2,hda6,ago4,dcl3 and rdr2 (Popova et al. 2013).Although its thermotolerance to high temperature has not yetbeen analysed, the H2A.Z incorporation defective mutantarp6 exhibits warm temperature (27 °C), responsive growthand developmental phenotypes, including plant architectureand flowering time (Kumar & Wigge 2010). Our studytogether with these previous studies indicates that multiple
epigenetic pathways are involved in various aspects of plantgrowth response to heat. Yet future studies are required todetermine whether some of these epigenetic factors mightwork together to regulate same aspect of plant heat response.
In nature heat is frequently accompanied by strong sun-light. AtASF1 has been reported as required for repair ofDNA damage induced by UV-B (Lario et al. 2013) as well asby other types of genotoxic stress (Zhu et al. 2011). Heatinduces H2O2 accumulation, which is a potent agent causingdamage to DNA, proteins and other molecules (Gill & Tuteja2010). Our study shows that the H2O2 production is normal,but oxidative response genes are down-regulated in Atasf1ab.Down-regulation of heat and oxidative stress response genescontrasts the unchanged expression or up-regulation of DNAdamage repair genes previously reported in Atasf1ab (Zhuet al. 2011; Lario et al. 2013). Our focused study on severalgenes, including HsfA2, Hsp101, Hsp70, Hsa32, Hsp17.6A andHsp17.6B-CI, revealed that AtASF1A/B positively regulatebasal as well as heat and oxidative stress-responsive expres-sion of these genes. The strong heat-induced expressionprovides an attractive system to address mechanisms ofAtASF1A/B function in gene expression control.
Our gene expression and ChIP data show that heat inducesHsfA2 and Hsa32 transcription, which is associated with PolII and AtASF1A/B accumulation at these gene loci. Position-ing of nucleosome in the path of transcriptionally engagedPol II is generally detrimental (Petesch & Lis 2012).For transcription initiation and elongation to proceed, Pol
(a) (b)
Figure 4. Comparison of oxidative stress response of Arabidopsis wild-type (WT) and mutant Atasf1ab plants. (a) Comparison of H2O2
production. Roots of 5-day-old WT (indicated by red arrowhead) and Atasf1ab mutant (indicated by yellow arrowhead) seedlings before(control) and after heat treated at 45 °C (heat) for indicated time were stained with fluorescent dye 2′,7′-dichlorodihydrofluorescein diacetate(H2DCFDA) and imaged. (b) Comparison of expression levels of heat stress genes. Quantitative RT-PCR analysis was performed on samplescollected at indicated time point from 12-day-old seedlings treated with the photosensitizer Rose Bengal (RB). The expression levels of theindicated genes were normalized to UBC28. The relative values were referenced to that of the mock-treated WT control, which is set as 1.Mean values and standard deviations are shown from three independent experiments.
2134 M. Weng et al.
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

II-containing transcriptional machinery needs to overcomethe polar barrier of nucleosomes at the transcription start siteand throughout the gene body. Our data reveal that HsfA2and Hsa32 activation is associated with AtASF1A/B-dependent nucleosome removal, suggesting a positive role ofAtASF1A/B in nucleosome dissociation. In line with thislatter proposal, AtASF1A/B specifically bind H3 (Zhu et al.2011) and the yeast and human ASF1-H3-H4 ternarycomplex is mutually exclusive from the nucleosomal core(H3-H4)2 tetramer (English et al. 2006; Natsume et al. 2007).The heat response is a universally conserved reaction, andnucleosome loss is also associated with yeast and animal HSFand HSP genes (Erkina et al. 2010; Teves & Henikoff 2011).
Therefore, it is reasonable to speculate that our newlydiscovered role of ASF1 may extend to heat response inother organisms.
Yeast ASF1 stimulates Rtt109 in catalysing H3K56ac(Schneider et al. 2006; Avvakumov et al. 2011). Our in vitroassay revealed that AtASF1A can also stimulate H3K56acproduction by Rtt109. Nevertheless, Western blot analysisrevealed roughly similar levels of H3K56ac in WT andAtasf1ab, indicating that AtASF1A/B are not major deter-mining factors of global H3K56ac in Arabidopsis. In animals,both CBP/p300 and GCN5 acetyltransferases are involved incatalysing H3K56ac (Das et al. 2009; Kong et al. 2011).Arabidopsis possesses five CBP/p300-type acetyltransferases
(a)
(b)
(c)
(d)
Figure 5. Analysis of AtASF1, H3 and polymerase II (Pol II) protein levels at gene-specific chromatin regions in Arabidopsis wild-type(WT) and mutant Atasf1ab plants and in response to heat stress. (a) A schematic representation of gene structure indicating the regionsexamined by chromatin immunoprecipitation (ChIP). Black boxes represent exons; white boxes represent untranslated region; lines representpromoter and introns; number-labelled bars represent regions amplified by corresponding primer pairs. (b–d) ChIP analysis for the relativeoccupancy of AtASF1 (b), H3 (c) and Pol II (d) at the indicated gene chromatin regions. WT and Atasf1ab plants untreated or treated byincubation at 37 °C for 1.5 h were analysed. Values were referenced to those of untreated Atasf1ab, which were set as 1. Mean values andstandard deviations are shown from two independent experiments. The symbols * and # indicate statistically significant differences (P < 0.05)observed for heat effect in either WT or Atasf1ab and for Atasf1ab effect on the heat induction compared with WT, respectively.
AtASF1 in heat stress response 2135
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

(AtHAC1/2/4/5/12; Pandey et al. 2002), and AtHAC1/5/12have been shown to possess broad substrate specificities invitro, including H3K9/14 and H4K5/8/12/16 sites (Earleyet al. 2007). The Arabidopsis ortholog AtGCN5 can alsoacetylate H3K14 in vitro (Earley et al. 2007), and its depriva-tion affects H3K9/27 acetylation levels in planta (Benhamedet al. 2006). It is yet unknown whether AtHAC1/2/4/5/12 andAtGCN5 are also involved in H3K56ac deposition. Our ChIPanalysis has uncovered a substantial role of AtASF1A/B inH3K56ac deposition at HsfA2 and Hsa32 loci. We found thatAtASF1A/B are required for maximal levels of H3K56acenrichment, which occurs at HsfA2 and Hsa32 loci in plantresponse to heat treatment. It has been reported thatAtASF1A/B bind HAM1/2 (Lario et al. 2013), and thatHAM1/2 are required for H4K5ac deposition at Arabidopsis-flowering regulatory genes FLC and MAF3/4 (Xiao et al.2013). It is currently unclear whether HAM1/2 have H3K56ac-catalysing activity or whether another (yet unknown identity)enzyme is responsible for H3K56ac deposition at HsfA2and Hsa32. In any case, our observed H3K56ac increase inresponse to heat is positively correlated with nucleosomeremoval in HsfA2 and Hsa32 activation. This is in agreementwith the function of H3K56ac in facilitating nucleosomedissociation.
A recent study in yeast found that H3K56ac promotesH2A.Z removal from the nucleosome through changing thechromatin remodelling factor SWR-C substrate specificity,providing a link between H3K56ac and H2A.Z in theregulation of nucleosome turnover (Watanabe et al. 2013).
Warm temperature (27 °C) treatment of Arabidopsisplants affects H2A.Z/H2A exchange activity at many genesincluding HSP70, and the arp6 mutant defective in H2A.Zincorporation displays a constitutive heat response pheno-type (Kumar & Wigge 2010). Future characterization ofArabidopsis enzymes involved in H3K56ac deposition andchromatin remodelling factors involved in H2A.Z exchangein relation with AtASF1 function will shed further insightinto mechanisms of nucleosome dynamics in plant responseto heat stress.
ACKNOWLEDGMENTS
This work was supported by the Chinese Ministry ofScience and Technology (grant no. 2012CB910500 and2011CB944600) and the National Natural Science Founda-tion of China (31271374 and 30971443). The research wasconducted within the context of the International AssociatedLaboratory Plant Epigenome Research, LIA PER.
REFERENCES
Avvakumov N., Nourani A. & Cote J. (2011) Histone chaperones: modulatorsof chromatin marks. Molecular Cell 41, 502–514.
Benhamed M., Bertrand C., Servet C. & Zhou D.X. (2006) Arabidopsis GCN5,HD1, and TAF1/HAF2 interact to regulate histone acetylation required forlight-responsive gene expression. The Plant Cell 18, 2893–2903.
Berr A., Menard R., Heitz T. & Shen W.H. (2012) Chromatin modification andremodelling: a regulatory landscape for the control of Arabidopsis defenceresponses upon pathogen attack. Cellular Microbiology 14, 829–839.
(a) (b)
Figure 6. Analysis of H3K56ac levels at gene-specific chromatin regions in Arabidopsis wild-type (WT) and mutant Atasf1ab plants and inresponse to heat stress. (a) In vitro assay for H3K56 acetylation. Recombinant proteins GST-Rtt109 (indicated by black arrowhead) andHis-AtASF1A (indicated by black arrow) were produced and tested in the enzyme activity assay. The histone mixture (indicated by openarrowhead) was used as substrates. The upper panel shows protein gel staining by Coomassie Brilliant Blue (CBB) for loading control, andthe lower panel shows Western blot (WB) detection by anti-H3K56ac antibody. (b) Relative H3K56ac levels at indicated gene chromatinregions. WT and Atasf1ab plants untreated or treated by incubation at 37 °C for 1.5 h were analysed using anti-H3 and anti-H3K56ac.H3K56ac-to-H3 ratio values were referenced to those of untreated Atasf1ab, which were set as 1. Mean values and standard deviations areshown from two independent experiments. The symbols * and # indicate statistically significant differences (P < 0.05) observed for heat effectin either WT or Atasf1ab and for Atasf1ab effect on the heat induction compared with WT, respectively.
2136 M. Weng et al.
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

Boden S.A., Kavanova M., Finnegan E.J. & Wigge P.A. (2013) Thermal stresseffects on grain yield in Brachypodium distachyon occur via H2A.Z-nucleosomes. Genome Biology 14, R65.
Busch W., Wunderlich M. & Schoffl F. (2005) Identification of novel heat shockfactor-dependent genes and biochemical pathways in Arabidopsis thaliana.The Plant Journal 41, 1–14.
Charng Y.Y., Liu H.C., Liu N.Y., Hsu F.C. & Ko S.S. (2006) Arabidopsis Hsa32,a novel heat shock protein, is essential for acquired thermotolerance duringlong recovery after acclimation. Plant Physiology 140, 1297–1305.
Charng Y.Y., Liu H.C., Liu N.Y., Chi W.T., Wang C.N., Chang S.H. & Wang T.T.(2007) A heat-inducible transcription factor, HsfA2, is required for exten-sion of acquired thermotolerance in Arabidopsis. Plant Physiology 143,251–262.
Chauhan H., Khurana N., Nijhavan A., Khurana J.P. & Khurana P. (2012) Thewheat chloroplastic small heat shock protein (sHSP26) is involved in seedmaturation and germination and imparts tolerance to heat stress. Plant, Cell& Environment 35, 1912–1931.
Das C., Lucia M.S., Hansen K.C. & Tyler J.K. (2009) CBP/p300-mediatedacetylation of histone H3 on lysine 56. Nature 459, 113–117.
Dong A., Liu Z., Zhu Y., Yu F., Li Z., Cao K. & Shen W.H. (2005) Interactingproteins and differences in nuclear transport reveal specific functions for theNAP1 family proteins in plants. Plant Physiology 138, 1446–1456.
Driscoll R., Hudson A. & Jackson S.P. (2007) Yeast Rtt109 promotes genomestability by acetylating histone H3 on lysine 56. Science 315, 649–652.
Earley K.W., Shook M.S., Brower-Toland B., Hicks L. & Pikaard C.S. (2007) Invitro specificities of Arabidopsis co-activator histone acetyltransferases:implications for histone hyperacetylation in gene activation. The PlantJournal 52, 615–626.
English C.M., Adkins M.W., Carson J.J., Churchill M.E. & Tyler J.K.(2006) Structural basis for the histone chaperone activity of Asf1. Cell 127,495–508.
Erkina T.Y., Zou Y., Freeling S., Vorobyev V.I. & Erkine A.M. (2010) Func-tional interplay between chromatin remodeling complexes RSC, SWI/SNFand ISWI in regulation of yeast heat shock genes. Nucleic Acids Research 38,1441–1449.
Fischer B.B., Krieger-Liszkay A. & Eggen R.L. (2004) Photosensitizers neutralred (type I) and rose bengal (type II) cause light-dependent toxicity inChlamydomonas reinhardtii and induce the Gpxh gene via increased singletoxygen formation. Environmental Science & Technology 38, 6307–6313.
Gao J., Zhu Y., Zhou W., Molinier J., Dong A. & Shen W.H. (2012) NAP1family histone chaperones are required for somatic homologous recombi-nation in Arabidopsis. The Plant Cell 24, 1437–1447.
Gill S.S. & Tuteja N. (2010) Reactive oxygen species and antioxidant machin-ery in abiotic stress tolerance in crop plants. Plant Physiology and Biochem-istry 48, 909–930.
Hong S.W., Lee U. & Vierling E. (2003) Arabidopsis hot mutants define multi-ple functions required for acclimation to high temperatures. Plant Physiol-ogy 132, 757–767.
Ikeda M., Mitsuda N. & Ohme-Takagi M. (2011) Arabidopsis HsfB1 andHsfB2b act as repressors of the expression of heat-inducible Hsfs but posi-tively regulate the acquired thermotolerance. Plant Physiology 157, 1243–1254.
Johnson L., Cao X. & Jacobsen S. (2002) Interplay between two epigeneticmarks: DNA methylation and histone H3 lysine 9 methylation. CurrentBiology 12, 1360–1367.
Kim M., Lee U., Small I., des Francs-Small C.C. & Vierling E. (2012) Mutationsin an Arabidopsis mitochondrial transcription termination factor-relatedprotein enhance thermotolerance in the absence of the major molecularchaperone HSP101. The Plant Cell 24, 3349–3365.
Kong S., Kim S.J., Sandal B., Lee S.M., Gao B., Zhang D.D. & Fang D. (2011)The type III histone deacetylase Sirt1 protein suppresses p300-mediatedhistone H3 lysine 56 acetylation at Bclaf1 promoter to inhibit T cell activa-tion. The Journal of Biological Chemistry 286, 16967–16975.
Konigshofer H., Tromballa H.W. & Loppert H.G. (2008) Early events in sig-nalling high-temperature stress in tobacco BY2 cells involve alterations inmembrane fluidity and enhanced hydrogen peroxide production. Plant, Cell& Environment 31, 1771–1780.
Kumar S.V. & Wigge P.A. (2010) H2A.Z-containing nucleosomes mediate thethermosensory response in Arabidopsis. Cell 140, 136–147.
Lang-Mladek C., Popova O., Kiok K., Berlinger M., Rakic B., Aufsatz W., . . .Luschnig C. (2010) Transgenerational inheritance and resetting of stress-induced loss of epigenetic gene silencing in Arabidopsis. Molecular Plant 3,594–602.
Lario L.D., Ramirez-Parra E., Gutierrez C., Spampinato C.P. & Casati P. (2013)ANTI-SILENCING FUNCTION1 proteins are involved in ultraviolet-induced DNA damage repair and are cell cycle regulated by E2F transcrip-tion factors in Arabidopsis. Plant Physiology 162, 1164–1177.
Lin L.J. & Schultz M.C. (2011) Promoter regulation by distinct mechanisms offunctional interplay between lysine acetylase Rtt109 and histone chaperoneAsf1. Proceedings of the National Academy of Sciences of the United States ofAmerica 108, 19599–19604.
Liu Z.Q., Zhu Y., Gao J., Yu F., Dong A.W. & Shen W.H. (2009a) Molecularand reverse genetic characterization of NUCLEOSOME ASSEMBLYPROTEIN1 (NAP1) genes unravels their function in transcription andnucleotide excision repair in Arabidopsis thaliana. The Plant Journal 59,27–38.
Liu Z.Q., Gao J., Dong A.W. & Shen W.H. (2009b) A truncated ArabidopsisNUCLEOSOME ASSEMBLY PROTEIN 1, AtNAP1;3T, alters plantgrowth responses to abscisic acid and salt in the Atnap1;3-2 mutant. Molecu-lar Plant 2, 688–699.
Lohmann C., Eggers-Schumacher G., Wunderlich M. & Schoffl F. (2004) Twodifferent heat shock transcription factors regulate immediate early expres-sion of stress genes in Arabidopsis. Molecular Genetics and Genomics 271,11–21.
Minard L.V., Williams J.S., Walker A.C. & Schultz M.C. (2011) Transcriptionalregulation by Asf1: new mechanistic insights from studies of the DNAdamage response to replication stress. The Journal of Biological Chemistry286, 7082–7092.
Mittler R., Finka A. & Goloubinoff P. (2012) How do plants feel the heat?Trends in Biochemical Sciences 37, 118–125.
Natsume R., Eitoku M., Akai Y., Sano N., Horikoshi M. & Senda T. (2007)Structure and function of the histone chaperone CIA/ASF1 complexed withhistones H3 and H4. Nature 446, 338–341.
Neumann H., Hancock S.M., Buning R., Routh A., Chapman L., Somers J., . . .Chin J.W. (2009) A method for genetically installing site-specific acetylationin recombinant histones defines the effects of H3 K56 acetylation. MolecularCell 36, 153–163.
Nover L., Bharti K., Doring P., Mishra S.K., Ganguli A. & Scharf K.D. (2001)Arabidopsis and the heat stress transcription factor world: how many heatstress transcription factors do we need? Cell Stress and Chaperones 6, 177–189.
Pandey R., Muller A., Napoli C.A., Selinger D.A., Pikaard C.S., Richards E.J.,. . . Jorgensen R.A. (2002) Analysis of histone acetyltransferase and histonedeacetylase families of Arabidopsis thaliana suggests functional diversifica-tion of chromatin modification among multicellular eukaryotes. NucleicAcids Research 30, 5036–5055.
Pecinka A., Rosa M., Schikora A., Berlinger M., Hirt H., Luschnig C. &Mittelsten Scheid O. (2009) Transgenerational stress memory is not ageneral response in Arabidopsis. PLoS ONE 4, e5202.
Pecinka A., Dinh H.Q., Baubec T., Rosa M., Lettner N. & Mittelsten Scheid O.(2010) Epigenetic regulation of repetitive elements is attenuated by pro-longed heat stress in Arabidopsis. The Plant Cell 22, 3118–3129.
Petesch S.J. & Lis J.T. (2012) Overcoming the nucleosome barrier duringtranscript elongation. Trends in Genetics 28, 285–294.
Popova O.V., Dinh H.Q., Aufsatz W. & Jonak C. (2013) The RdDM pathway isrequired for basal heat tolerance in Arabidopsis. Molecular Plant 6, 396–410.
Qu A.L., Ding Y.F., Jiang Q. & Zhu C. (2013) Molecular mechanisms of theplant heat stress response. Biochemical and Biophysical Research Commu-nications 432, 203–207.
Rufiange A., Jacques P.E., Bhat W., Robert F. & Nourani A. (2007) Genome-wide replication-independent histone H3 exchange occurs predominantly atpromoters and implicates H3 K56 acetylation and Asf1. Molecular Cell 27,393–405.
Saidi Y., Finka A. & Goloubinoff P. (2011) Heat perception and signalling inplants: a tortuous path to thermotolerance. The New Phytologist 190, 556–565.
Scharf K.D., Berberich T., Ebersberger I. & Nover L. (2012) The plant heatstress transcription factor (Hsf) family: structure, function and evolution.Biochimica et Biophysica Acta 1819, 104–119.
Schneider J., Bajwa P., Johnson F.C., Bhaumik S.R. & Shilatifard A. (2006)Rtt109 is required for proper H3K56 acetylation: a chromatin mark associ-ated with the elongating RNA polymerase II. The Journal of BiologicalChemistry 281, 37270–37274.
Swindell W.R., Huebner M. & Weber A.P. (2007) Transcriptional profiling ofArabidopsis heat shock proteins and transcription factors reveals extensive
AtASF1 in heat stress response 2137
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138

overlap between heat and non-heat stress response pathways. BMCGenomics 8, 125.
Tagami H., Ray-Gallet D., Almouzni G. & Nakatani Y. (2004) Histone H3.1and H3.3 complexes mediate nucleosome assembly pathways dependent orindependent of DNA synthesis. Cell 116, 51–61.
Teves S.S. & Henikoff S. (2011) Heat shock reduces stalled RNA polymeraseII and nucleosome turnover genome-wide. Genes & Development 25, 2387–2397.
Tittel-Elmer M., Bucher E., Broger L., Mathieu O., Paszkowski J. & Vaillant I.(2010) Stress-induced activation of heterochromatic transcription. PLoSGenetics 6, e1001175.
Volkov R.A., Panchuk I.I., Mullineaux P.M. & Schoffl F. (2006) Heat stress-induced H(2)O(2) is required for effective expression of heat shock genes inArabidopsis. Plant Molecular Biology 61, 733–746.
Watanabe S., Radman-Livaja M., Rando O.J. & Peterson C.L. (2013) A histoneacetylation switch regulates H2A.Z deposition by the SWR-C remodelingenzyme. Science 340, 195–199.
Williams S.K., Truong D. & Tyler J.K. (2008) Acetylation in the globular coreof histone H3 on lysine-56 promotes chromatin disassembly during tran-scriptional activation. Proceedings of the National Academy of Sciences ofthe United States of America 105, 9000–9005.
Xiao J., Zhang H., Xing L., Xu S., Liu H., Chong K. & Xu Y. (2013) Require-ment of histone acetyltransferases HAM1 and HAM2 for epigenetic modi-fication of FLC in regulating flowering in Arabidopsis. Journal of PlantPhysiology 170, 444–451.
Yeh C.H., Kaplinsky N.J., Hu C. & Charng Y.Y. (2012) Some like it hot, somelike it warm: phenotyping to explore thermotolerance diversity. PlantScience 195, 10–23.
Zhu Y., Dong A., Meyer D., Pichon O., Renou J.P., Cao K. & Shen W.H. (2006)Arabidopsis NRP1 and NRP2 encode histone chaperones and are requiredfor maintaining postembryonic root growth. The Plant Cell 18, 2879–2892.
Zhu Y., Weng M., Yang Y., Zhang C., Li Z., Shen W.H. & Dong A. (2011)Arabidopsis homologues of the histone chaperone ASF1 are crucial forchromatin replication and cell proliferation in plant development. The PlantJournal 66, 443–455.
Zhu Y., Dong A. & Shen W.H. (2012) Histone variants and chromatin assem-bly in plant abiotic stress responses. Biochimica et Biophysica Acta 1819,343–348.
Received 6 December 2013; received in revised form 29 January 2014;accepted for publication 29 January 2014
SUPPORTING INFORMATION
Additional Supporting Information may be found in theonline version of this article at the publisher’s web-site:
Figure S1. Distribution of genes perturbed in Atasf1ab on thefive Arabidopsis chromosomes.Figure S2. Relative expression levels of several key heatstress genes in Arabidopsis wild-type (WT) and Atasf1abmutant under standard growth conditions.Figure S3. Relative expression levels of AtASF1A/B duringheat treatment.Figure S4. Western blot analysis of Hsp101 and HSP17.6protein levels in Arabidopsis wild-type (WT) and Atasf1abmutant plants.Figure S5. Specificity control of anti-AtASF1A antibody onWestern blot.Figure S6. Western blot analysis of H3K56ac levels inArabidopsis wild-type (WT) and Atasf1ab mutant plants.Table S1. Primers used in this study.Table S2. List of up-regulated genes in Atasf1a fromtranscriptome analysis.Table S3. List of up-regulated genes in Atasf1b fromtranscriptome analysis.Table S4. List of up-regulated genes in Atasf1ab fromtranscriptome analysis.Table S5. List of down-regulated genes in Atasf1a fromtranscriptome analysis.Table S6. List of down-regulated genes in Atasf1b fromtranscriptome analysis.Table S7. List of down-regulated genes in Atasf1ab fromtranscriptome analysis.
2138 M. Weng et al.
© 2014 John Wiley & Sons Ltd, Plant, Cell and Environment, 37, 2128–2138





























