Embed Size (px)
Citation preview

Glutamate Receptors at BipolarSynapses in the Inner Plexiform
Layer of Primate Retina:Light Microscopic Analysis
ULRIKE GRUNERT,1,2* BIN LIN,1AND PAUL R. MARTIN1,2
1Department of Physiology F13 and Institute for Biomedical Research,The University of Sydney, NSW 2006, Australia
2National Vision Research Institute and Department of Optometry and Vision Sciences,The University of Melbourne, Carlton, VIC 3053, Australia
ABSTRACTAt least 10 different types of bipolar cells have been distinguished in the primate retina.
The axon terminals of these cells stratify in distinct strata in the inner plexiform layer andare involved in parallel pathways to distinct types of ganglion cells. Ionotropic glutamatereceptor (GluR) subunits also show a stratified distribution in the inner plexiform layer.Here, we investigated whether different types of bipolar cells are associated with differenttypes of ionotropic glutamate receptors in the inner retina of a New World primate, thecommon marmoset Callithrix jacchus. Vertical cryostat sections through central retina weredouble labeled with immunohistochemical markers for bipolar cell types and with antibodiesto �-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptor subunits GluR1 to 4,kainate receptor subunits GluR6/7, and the NR1C2� subunit of the N-methyl-D-aspartate(NMDA) receptor. The axon terminals of bipolar cell types were reconstructed from confocalsections, and the colocalized immunoreactive puncta were quantified. For all bipolar celltypes, immunoreactive puncta for the AMPA receptor subunits GluR2, 2/3, and 4 werecolocalized at highest densities, whereas GluR1-immunoreactive puncta were expressed atvery low densities. The kainate receptor subunits GluR6/7 were predominantly associatedwith diffuse bipolar (DB6) and rod bipolar cells. The NMDA receptor subunit NR1C2� wasspecifically colocalized with flat midget and DB3 axons. These findings suggest that rod andcone bipolar cell types contribute to multiple but distinct glutamate receptor pathways inprimate retina. J. Comp. Neurol. 466:136–147, 2003. © 2003 Wiley-Liss, Inc.
Indexing terms: parallel pathways; CD15 immunohistochemistry; midget bipolar cells; diffuse
bipolar cells; rod bipolar cells
The existence of parallel pathways in the primate visualsystem has long been established (for reviews, see Lennieet al., 1990; Silveira and De Mello, 1998; Boycott andWassle, 1999; Wassle, 1999; Dacey, 2000). It is also wellestablished that these parallel pathways originate at thefirst synapse in the retina made by photoreceptors ontobipolar cells. And it is already at this synapse that diver-sity of glutamate receptors has been described. For exam-ple, ON-bipolar cell types express the metabotropic gluta-mate receptor mGluR6 (Nomura et al., 1994; Vardi et al.,2000), whereas OFF-bipolar and horizontal cells expressAMPA and/or kainate receptors (Haverkamp et al., 2000,2001a,b).
Parallel signals in primate retina are transmitted to theinner plexiform layer (IPL) by at least 10 morphologically
different types of bipolar cells (Boycott and Wassle, 1991;Kolb et al., 1992; Chan et al., 2001a). The axon terminalsof different bipolar cell types stratify in distinct layers of
Grant sponsor: National Health & Medical Research Council; Grantnumber: 153823; Grant sponsor: University of Sydney International Post-graduate Research Scholarship (B.L.).
*Correspondence to: Ulrike Grunert, National Vision Research Institute,The University of Melbourne, Cnr Keppel & Cardigan Streets, Carlton,VIC 3053 Australia. E-mail: [email protected]
Received 15 April 2003; Revised 3 June 2003; Accepted 6 June 2003DOI 10.1002/cne.10862Published online the week of September 15, 2003 in Wiley InterScience
(www.interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 466:136–147 (2003)
© 2003 WILEY-LISS, INC.

the IPL. Some of the bipolar types, including rod bipolar(RB) cells, blue cone bipolar cells, diffuse bipolar (DB)cells, and flat midget bipolar (FMB) cells, can be stainedselectively by immunohistochemical markers (Marshak etal., 1990; Grunert and Martin, 1991; Kouyama and Mar-shak, 1992; Milam et al., 1993; Grunert et al., 1994; Millsand Massey, 1999; Chan et al., 2001a,b). All bipolar celltypes probably contain glutamate and, thus, provide exci-tatory input to amacrine and ganglion cell processes in theIPL (Davanger et al., 1991; Crooks and Kolb, 1992; Martinand Grunert, 1992).
Ionotropic glutamate receptors are oligomeric ligand-gated ion channels and can be further subdivided into threemain groups: �-amino-3-hydroxy-5-methyl-4-isoxazole-propionate (AMPA), kainate (KA), and N-methyl-D-aspartate (NMDA) receptor channels. Receptors in eachclass are composed of several subunits: AMPA (GluR1,GluR2, GluR3, GluR4), kainate (GluR5, GluR6, GluR7,KA-1, KA-2), NMDA (NR1, NR2A, NR2B, NR2C, NR2D).
Several recent immunohistochemical studies have dem-onstrated that all major subgroups of ionotropic gluta-mate receptors (GluR), i.e., AMPA, kainate, and NMDAreceptors, are present in mammalian retinae (Hartveit etal., 1994; Qin and Pourcho, 1996, 1999a,b, 2001; Brand-statter et al., 1997; Morigiwa and Vardi, 1999; Fletcher etal., 2000; Haverkamp and Wassle, 2000; Hack et al., 2001;Pourcho et al., 2001; Grunert et al., 2002). These studiesalso showed that immunoreactivity is distinct for eachGluR subunit, i.e., different bands in the IPL show differ-ent staining intensities. Thus, it has been suggested thatthe laminar staining pattern seen for ionotropic GluRs inthe IPL corresponds to the stratification pattern of bipolarcell axons (Boycott and Wassle, 1999). Consistent withthis idea, recent studies of ionotropic GluRs demonstratedthat the AMPA receptor subunits GluR2/3 and GluR4, thekainate receptor subunits GluR6/7 and the � 1/2 subunitsare involved with the rod pathway (Qin and Pourcho,1999a; Ghosh et al., 2001; Grunert et al., 2002; Li et al.,2002) but NMDA receptors are not (Fletcher et al., 2000).In the present study, we identified the ionotropic GluRsubunits postsynaptic to the axon terminals of three typesof cone bipolar cell and the RB cell.
MATERIALS AND METHODS
Animals and tissue preparation
The retinae studied were from adult marmosets, Cal-lithrix jacchus, that were killed after electrophysiologicalexperiments unrelated to those described here. All proce-dures conformed to the provisions of the Australian Na-tional & Medical Research Council code of practice of thecare and use of animals. The marmosets were overdosedwith sodium pentobarbitone (80–150 mg/kg, i.v.), and theeyes were quickly removed. The posterior eyecup wasimmersion-fixed in 2% paraformaldehyde in 0.1 M phos-phate buffer (PB, pH 7.4) for 30 minutes. After fixation,the retina was dissected from the eyecup and cryopro-tected in 30% sucrose in 0.1 M phosphate buffer. Retinalpieces were sectioned vertically at 12 �m with a cryostatand collected onto gelatinized slides.
Immunofluorescence
The antibodies used to identify bipolar cell types inprimate retina have been described in detail previously. A
mouse monoclonal antibody to protein kinase C� (Amer-sham, diluted 1:100) was used to stain RB cells (Greferathet al., 1990). The mouse monoclonal antibody against thecalcium binding protein calbindin (CaBP D-28K) was ob-tained from Sigma (St. Louis, MO). It was used at adilution of 1:20,000 to stain DB3 cells (Martin andGrunert, 1992; Grunert et al., 1994; Luo et al., 1999). Themouse monoclonal antibody against the carbohydrateepitope CD15 (diluted 1:6) was a generous gift from Prof.Mai (Universitat Dusseldorf, Germany) and has been de-scribed previously (Andressen and Mai, 1997). It stainsFMB and DB6 cells in marmoset retina (Chan et al.,2001a).
Several antibodies against glutamate receptor subunitswere used. They were the same as used in our previousstudies on glutamate receptors in primate retina(Haverkamp et al., 2000, 2001a,b; Grunert et al., 2002;Lin et al., 2002). The rabbit anti-GluR1 receptor anti-serum (AB1504, diluted 1:50), the rabbit anti-GluR2 an-tiserum (AB1768, diluted 1:100), the rabbit anti-GluR2 &3 antiserum (AB1506, diluted 1:400), the rabbit anti-GluR4 antiserum (AB1508, diluted 1:400), and the rabbitanti-NMDAR1 C2� splice variant antiserum (AB5050P,which recognizes four of the eight splice variants of theNR1 subunit, diluted 1:1,000) were purchased fromChemicon (Temecula, CA). The rabbit anti GluR6/7 anti-serum (06-309, diluted 1:1,000) was purchased from Up-state Biotechnology, Inc. (Lake Placid, NY). In double-labeling experiments, sections were incubated in amixture of primary antibodies followed by a mixture ofsecondary antibodies. The following secondary antibodieswere used (purchased from Molecular Probes): goat-antirabbit IgG coupled to Alexa 594 (diluted 1:250), goat anti-mouse IgG coupled to Alexa 488 (1:250).
Confocal analysis
Fluorescent specimens were examined with a Leica TCSSP confocal laser-scanning microscope equipped with akrypton-argon laser. Laser lines and emission were opti-mized with the Leica TCS power scan software. High-resolution scanning was performed with a 100�1.4 NA oilPlanApo objective. Scans were collected with a z-axis stepsize of 0.3 or 0.4 �m for each frame with 1,024 � 1,024pixels, using a zoom of 4� to give a final magnification of40.0 pixels/�m. The brightness and the contrast of thefinal images were adjusted by using Adobe Photoshop 6.0.
The axon terminals and colocalized immunoreactive(IR) puncta were marked directly from the monitor usingimage display and graphic overlay routines, written inMatLab (Image processing toolbox, Mathworks) by theauthors and available on request. The (X, Y) coordinates ofcolocalized IR puncta and of points on the dendritic treewere stored as a connectivity matrix, so each IR punctumwas assigned to one point on a two-dimensional represen-tation of the axon terminal.
In a complementary analysis, colocalized IR punctawere identified by using semiautomated image processingroutines (image processing toolbox; Mathworks, Natick,MA). The (X,Y) position and (Z) focal plane of each fluo-rescent punctum in the image stack was determined byfinding the local intensity maximum in three-dimensional(image voxel) space. The region enclosing a single bipolaraxon terminal was defined by operator-assisted imagemask and threshold operations. Threshold values close to1/2 maximum intensity were chosen as they gave the best
137GLUTAMATE RECEPTORS IN PRIMATE RETINA
match with the observers’ judgment of axonal borders inmanual analyses. These procedures yield two sets of coor-dinates: the (X,Y,Z) center position of fluorescent punctaand the (Y,Y,Z) coordinates of the borders of a single axonterminal. Estimates of colocalized puncta density thenproceeded automatically by Boolean logic operations onthese data.
RESULTS
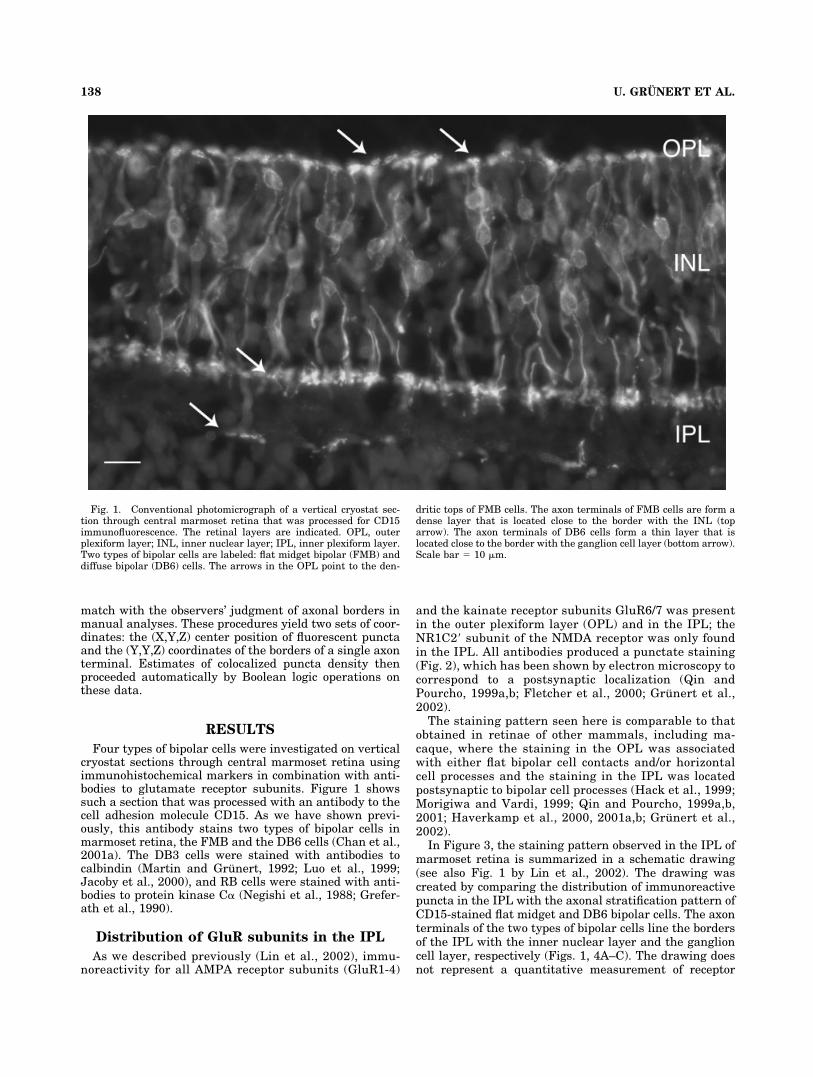
Four types of bipolar cells were investigated on verticalcryostat sections through central marmoset retina usingimmunohistochemical markers in combination with anti-bodies to glutamate receptor subunits. Figure 1 showssuch a section that was processed with an antibody to thecell adhesion molecule CD15. As we have shown previ-ously, this antibody stains two types of bipolar cells inmarmoset retina, the FMB and the DB6 cells (Chan et al.,2001a). The DB3 cells were stained with antibodies tocalbindin (Martin and Grunert, 1992; Luo et al., 1999;Jacoby et al., 2000), and RB cells were stained with anti-bodies to protein kinase C� (Negishi et al., 1988; Grefer-ath et al., 1990).
Distribution of GluR subunits in the IPL
As we described previously (Lin et al., 2002), immu-noreactivity for all AMPA receptor subunits (GluR1-4)
and the kainate receptor subunits GluR6/7 was presentin the outer plexiform layer (OPL) and in the IPL; theNR1C2� subunit of the NMDA receptor was only foundin the IPL. All antibodies produced a punctate staining(Fig. 2), which has been shown by electron microscopy tocorrespond to a postsynaptic localization (Qin andPourcho, 1999a,b; Fletcher et al., 2000; Grunert et al.,2002).
The staining pattern seen here is comparable to thatobtained in retinae of other mammals, including ma-caque, where the staining in the OPL was associatedwith either flat bipolar cell contacts and/or horizontalcell processes and the staining in the IPL was locatedpostsynaptic to bipolar cell processes (Hack et al., 1999;Morigiwa and Vardi, 1999; Qin and Pourcho, 1999a,b,2001; Haverkamp et al., 2000, 2001a,b; Grunert et al.,2002).
In Figure 3, the staining pattern observed in the IPL ofmarmoset retina is summarized in a schematic drawing(see also Fig. 1 by Lin et al., 2002). The drawing wascreated by comparing the distribution of immunoreactivepuncta in the IPL with the axonal stratification pattern ofCD15-stained flat midget and DB6 bipolar cells. The axonterminals of the two types of bipolar cells line the bordersof the IPL with the inner nuclear layer and the ganglioncell layer, respectively (Figs. 1, 4A–C). The drawing doesnot represent a quantitative measurement of receptor
Fig. 1. Conventional photomicrograph of a vertical cryostat sec-tion through central marmoset retina that was processed for CD15immunofluorescence. The retinal layers are indicated. OPL, outerplexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer.Two types of bipolar cells are labeled: flat midget bipolar (FMB) anddiffuse bipolar (DB6) cells. The arrows in the OPL point to the den-
dritic tops of FMB cells. The axon terminals of FMB cells are form adense layer that is located close to the border with the INL (toparrow). The axon terminals of DB6 cells form a thin layer that islocated close to the border with the ganglion cell layer (bottom arrow).Scale bar � 10 �m.
138 U. GRUNERT ET AL.
staining. The staining pattern summarized here is consis-tent with our previous results of GluR staining in ma-caque and marmoset retinae (Grunert et al., 2002; Lin etal., 2002).
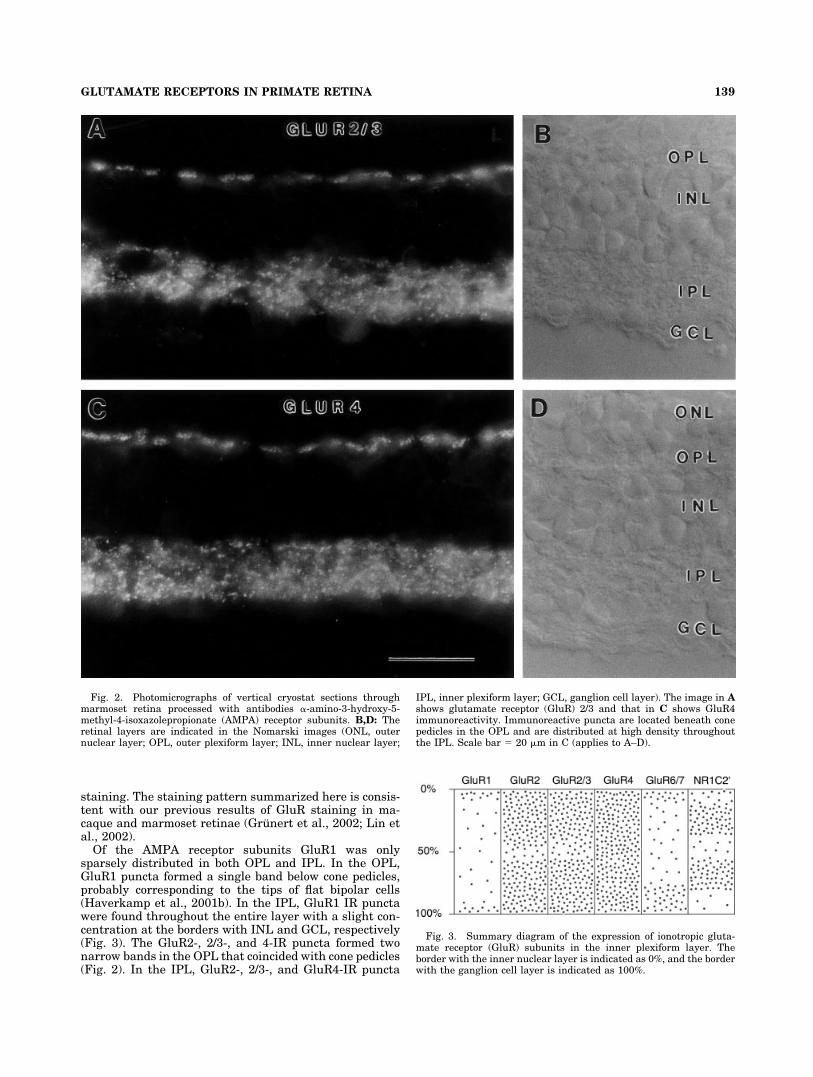
Of the AMPA receptor subunits GluR1 was onlysparsely distributed in both OPL and IPL. In the OPL,GluR1 puncta formed a single band below cone pedicles,probably corresponding to the tips of flat bipolar cells(Haverkamp et al., 2001b). In the IPL, GluR1 IR punctawere found throughout the entire layer with a slight con-centration at the borders with INL and GCL, respectively(Fig. 3). The GluR2-, 2/3-, and 4-IR puncta formed twonarrow bands in the OPL that coincided with cone pedicles(Fig. 2). In the IPL, GluR2-, 2/3-, and GluR4-IR puncta
Fig. 2. Photomicrographs of vertical cryostat sections throughmarmoset retina processed with antibodies �-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptor subunits. B,D: Theretinal layers are indicated in the Nomarski images (ONL, outernuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer;
IPL, inner plexiform layer; GCL, ganglion cell layer). The image in Ashows glutamate receptor (GluR) 2/3 and that in C shows GluR4immunoreactivity. Immunoreactive puncta are located beneath conepedicles in the OPL and are distributed at high density throughoutthe IPL. Scale bar � 20 �m in C (applies to A–D).
Fig. 3. Summary diagram of the expression of ionotropic gluta-mate receptor (GluR) subunits in the inner plexiform layer. Theborder with the inner nuclear layer is indicated as 0%, and the borderwith the ganglion cell layer is indicated as 100%.
139GLUTAMATE RECEPTORS IN PRIMATE RETINA
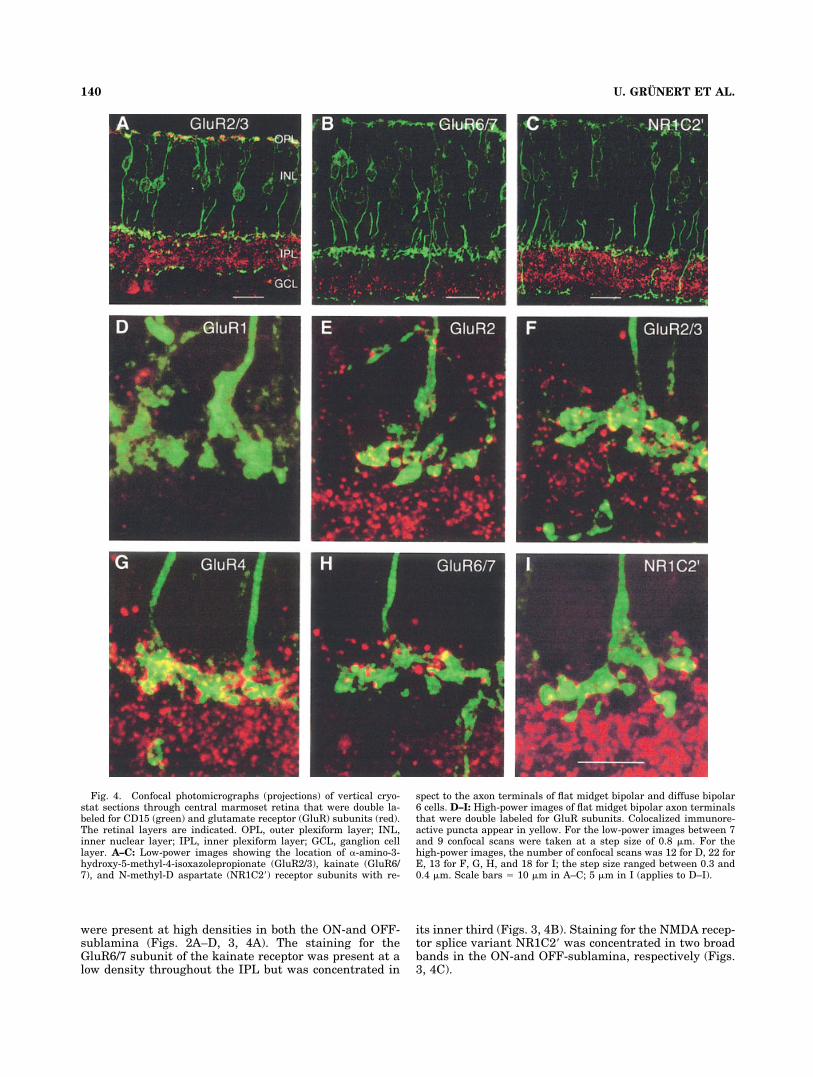
were present at high densities in both the ON-and OFF-sublamina (Figs. 2A–D, 3, 4A). The staining for theGluR6/7 subunit of the kainate receptor was present at alow density throughout the IPL but was concentrated in
its inner third (Figs. 3, 4B). Staining for the NMDA recep-tor splice variant NR1C2� was concentrated in two broadbands in the ON-and OFF-sublamina, respectively (Figs.3, 4C).
Fig. 4. Confocal photomicrographs (projections) of vertical cryo-stat sections through central marmoset retina that were double la-beled for CD15 (green) and glutamate receptor (GluR) subunits (red).The retinal layers are indicated. OPL, outer plexiform layer; INL,inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion celllayer. A–C: Low-power images showing the location of �-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (GluR2/3), kainate (GluR6/7), and N-methyl-D aspartate (NR1C2�) receptor subunits with re-
spect to the axon terminals of flat midget bipolar and diffuse bipolar6 cells. D–I: High-power images of flat midget bipolar axon terminalsthat were double labeled for GluR subunits. Colocalized immunore-active puncta appear in yellow. For the low-power images between 7and 9 confocal scans were taken at a step size of 0.8 �m. For thehigh-power images, the number of confocal scans was 12 for D, 22 forE, 13 for F, G, H, and 18 for I; the step size ranged between 0.3 and0.4 �m. Scale bars � 10 �m in A–C; 5 �m in I (applies to D–I).
140 U. GRUNERT ET AL.

Glutamate receptors associatedwith FMB axons
Figure 4D–I shows high-power confocal images (projec-tions) of axon terminals of CD15-labeled FMB cells. Im-munoreactive puncta for the GluR2, 2/3, and 4 subunits ofthe AMPA receptor and the NR1C2� subunit of the NMDAreceptor are densely colocalized with the axon terminals ofFMB cells (Fig. 4E–G,I). In comparison, there are only fewcolocalized IR puncta for the GluR1 and GluR6/7 subunits(Fig. 4D,H), indicating that these subunits are not ex-pressed postsynaptically to FMB cells.
Glutamate receptors associatedwith DB3 axons
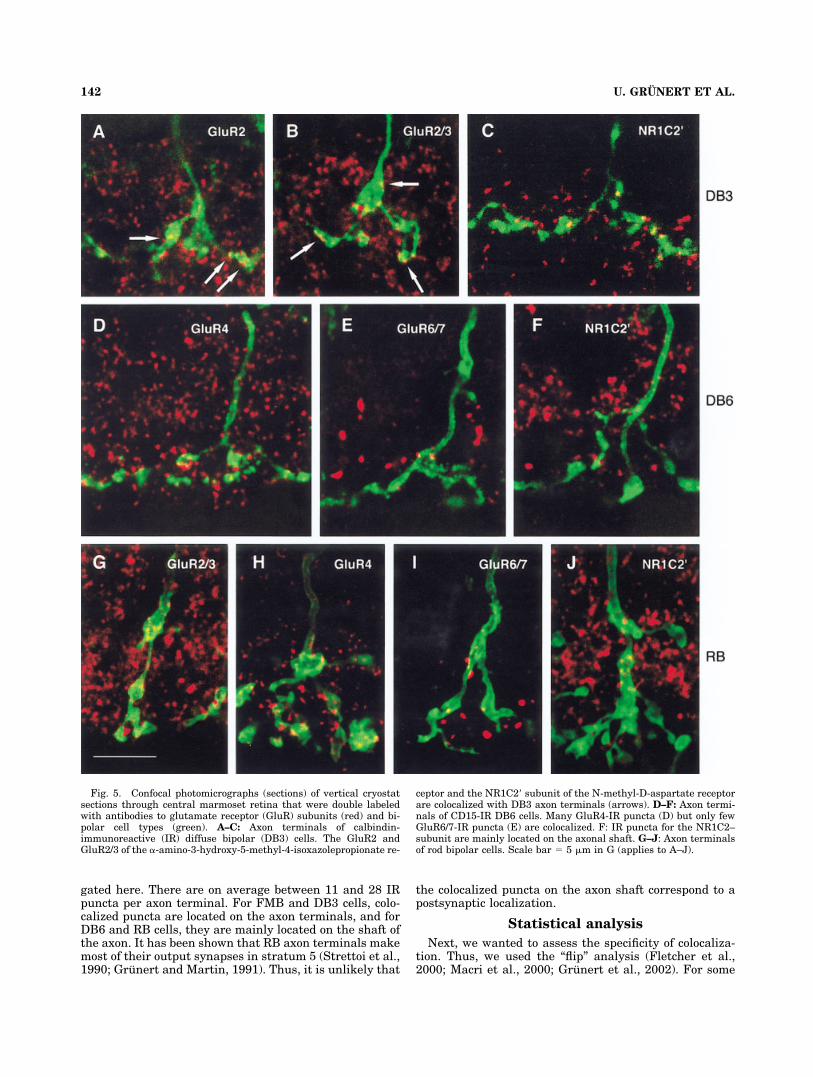
Figure 5A–C shows high-power confocal images ofdouble-labeled axon terminals of DB3 cells. The axon ter-minals are shown in combination with the GluR2 andGluR2/3 subunits of the AMPA receptor and the NR1C2�subunit of the NMDA receptor. All subunits are colocal-ized with DB3 axon terminals. Similar results were ob-tained with antibodies to the GluR4 subunit of the AMPAreceptor, but the GluR1 subunit of the AMPA receptor andthe GluR6/7 subunits of the kainate receptor showed ba-sically no colocalization.
Glutamate receptors associatedwith DB6 axons
Figure 5D–F shows DB6 axon terminals double-labeledwith antibodies to CD15 and antibodies to GluR subunits.The AMPA receptor subunit GluR4 is strongly associatedwith the axon and axon terminals of DB6 cells (Fig. 5D).Similar results were obtained for the AMPA subunitsGluR2 and GluR2/3 but the GluR1 subunit showed onlylow numbers of colocalized IR puncta.
Consistent with previous studies, we found that immu-noreactivity for the GluR6/7 subunits of the kainate re-ceptor produces large and small puncta. The large punctaare concentrated in strata 4 and 5 of the IPL (Grunert etal., 2002; Li et al., 2002; Lin et al., 2002). Puncta of bothsizes are colocalized with DB6 axon terminals with thesmall puncta predominating (Fig. 5E). However, the colo-calized IR puncta formed only a small percentage of thetotal number of GluR6/7 puncta.
The NMDA receptor subunits NR1C2� are absent fromstratum 5 (Figs. 3, 4C); thus, colocalized IR puncta are notfound on the axon terminals of DB6 cells but some werelocated on the shaft of DB6 axons (Figs. 5F).
Glutamate receptors associatedwith RB axons
Figure 5G–J shows sections of RB axon terminals pro-cessed with antibodies to GluR subunits. Consistent withprevious studies in macaque and rabbit retinae (Ghosh et al.,2001; Li et al., 2002), the IR puncta for the AMPA receptorsubunits GluR2/3 and GluR4 are colocalized with RB axonterminals (Fig. 5G,H), and there is also some colocalizationwith the kainate receptor subunits GluR6/7 (Fig. 5I).
As described above, the NMDA receptor subunitsNR1C2� are absent from stratum 5 (Figs. 3, 4C), andconsistently it has been reported previously that NMDAreceptor subunits are not postsynaptic to RB cells(Fletcher et al., 2000). However, by using confocal micros-copy at high resolution, we noted that there were someimmunoreactive puncta colocalized with the axon shaft of
RB terminals (Figs. 5J, 7). Whether this colocalization ofIR puncta exceeds a random distribution will be discussedbelow.
Quantitative Analysis
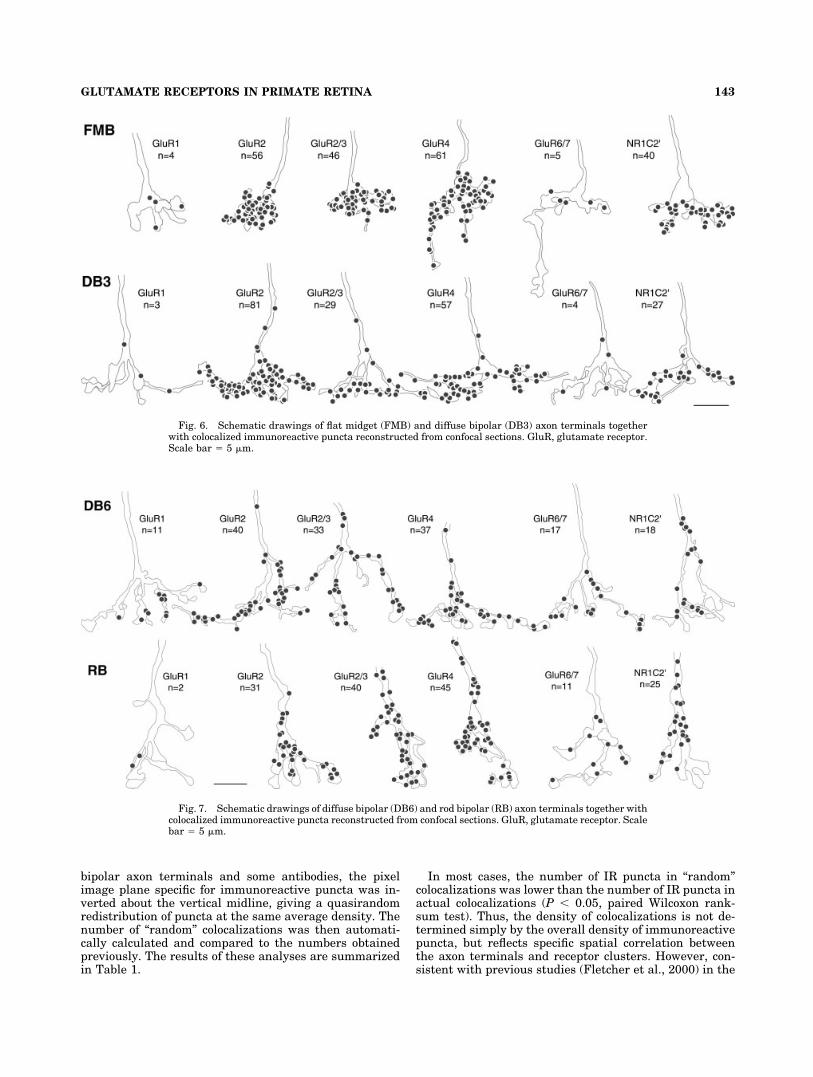
Bipolar axon terminals were reconstructed from confo-cal slices and the colocalized IR puncta were counted foreach axon terminal. The diffuse bipolar types (DB3 andDB6) have larger axonal tree diameters than FMB and RBcells and thus at the magnification used here the axonalprocesses of DB3 and DB6 cells were usually cut off attheir distal ends. For each cell type and GluR subunit, atleast 3 and up to 10 terminals were analyzed quantita-tively yielding in a total of 108 bipolar axon terminals. Allcells were from central retina, i.e., between 1 and 2 mmeccentricity.
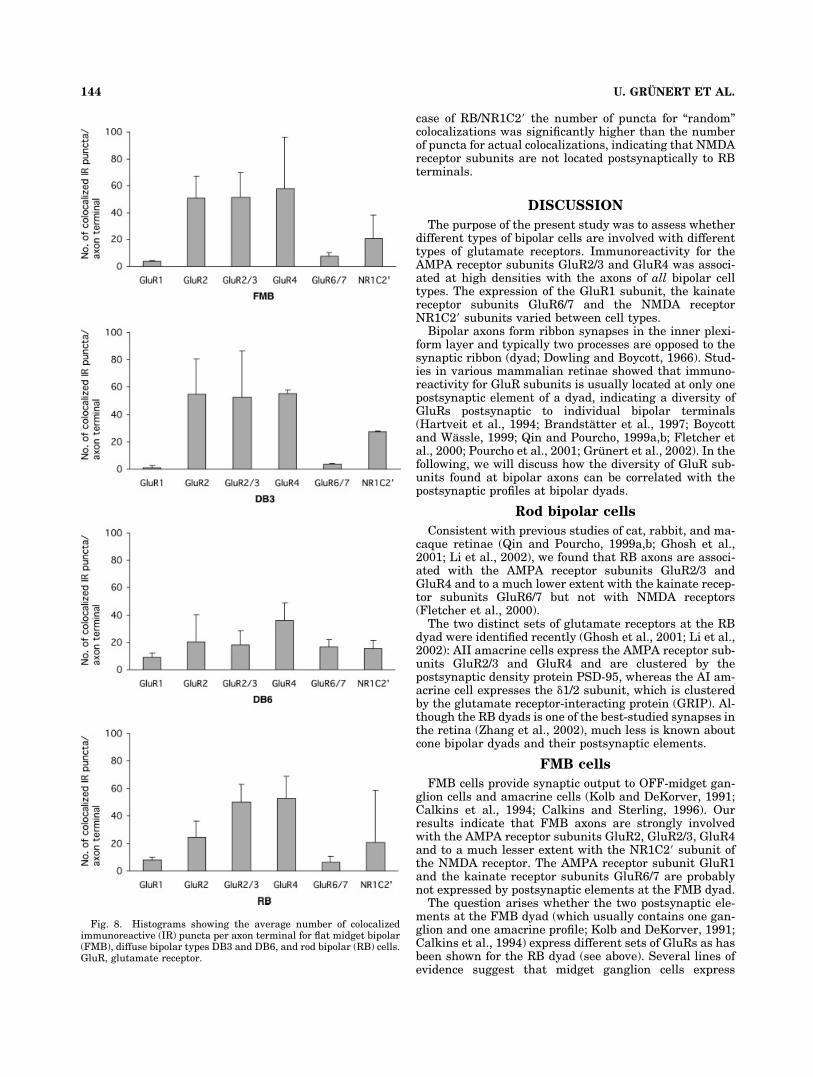
In Figures 6 and 7, examples of reconstructed FMB,DB3, DB6, and RB axon terminals are shown togetherwith colocalized IR puncta for different GluR subunits.The numbers given next to an axon terminal indicate thenumbers of colocalized IR puncta for this terminal. Theresults of the quantitative analysis are summarized in thehistograms of Figure 8. The average number of colocalizedIR puncta per bipolar axon terminal is given with thestandard deviation.
For some bipolar axon terminals we also calculated thedensities of colocalized puncta as puncta per �m2. Thevalues are given in Table 1. The AMPA receptor subunitsGluR2/3 and GluR4 are colocalized at high densities,whereas GluR6/7 and NR1C2� subunits only show lowdensities. There is good correlation between puncta densityand the absolute number of puncta. This finding is expected,as the different bipolar cell types do not show dramaticvariation in the size of their respective axon terminals. Weconclude that, although the number of colocalized punctamay not be as meaningful a measure as puncta density, it isnevertheless a good measure of the functional output ofbipolar cells to distinct receptor pathways.
AMPA receptor subunits
The GluR subunits 2, 2/3, and 4 are present at a relativehigh densities for all bipolar cell types analyzed here. Onaverage, there were between 20 and 55 IR puncta per axonterminal. Thus, these AMPA receptor subunits are prob-ably involved with cone as well as rod pathways.
The GluR 1 subunit is only expressed at low numbers.Consistent with their overall low density, the IR punctafor the GluR1 subunit show very little colocalization withbipolar axons (average: between 1 and 9 IR puncta peraxon terminal). The highest density of colocalized GluR1IR puncta is found on axon terminals of DB6 cells.
Kainate receptor subunits
The IR puncta for the GluR6/7 subunit of the kainatereceptor are predominantly associated with the axon ter-minals of the two ON-bipolar types investigated here, DB6and RB cells (average: between 7 and 17 IR puncta peraxon terminal). The low number of colocalized punctafound on FMB axon terminals was probably not specific asmany terminals did not show any colocalization (these arenot included in the quantitative analysis).
NMDA receptor subunits
The IR puncta for the NR1C2� subunit of the NMDAreceptor are associated with all bipolar terminals investi-
141GLUTAMATE RECEPTORS IN PRIMATE RETINA
gated here. There are on average between 11 and 28 IRpuncta per axon terminal. For FMB and DB3 cells, colo-calized puncta are located on the axon terminals, and forDB6 and RB cells, they are mainly located on the shaft ofthe axon. It has been shown that RB axon terminals makemost of their output synapses in stratum 5 (Strettoi et al.,1990; Grunert and Martin, 1991). Thus, it is unlikely that
the colocalized puncta on the axon shaft correspond to apostsynaptic localization.
Statistical analysis
Next, we wanted to assess the specificity of colocaliza-tion. Thus, we used the “flip” analysis (Fletcher et al.,2000; Macri et al., 2000; Grunert et al., 2002). For some
Fig. 5. Confocal photomicrographs (sections) of vertical cryostatsections through central marmoset retina that were double labeledwith antibodies to glutamate receptor (GluR) subunits (red) and bi-polar cell types (green). A–C: Axon terminals of calbindin-immunoreactive (IR) diffuse bipolar (DB3) cells. The GluR2 andGluR2/3 of the �-amino-3-hydroxy-5-methyl-4-isoxazolepropionate re-
ceptor and the NR1C2� subunit of the N-methyl-D-aspartate receptorare colocalized with DB3 axon terminals (arrows). D–F: Axon termi-nals of CD15-IR DB6 cells. Many GluR4-IR puncta (D) but only fewGluR6/7-IR puncta (E) are colocalized. F: IR puncta for the NR1C2–subunit are mainly located on the axonal shaft. G–J: Axon terminalsof rod bipolar cells. Scale bar � 5 �m in G (applies to A–J).
142 U. GRUNERT ET AL.
bipolar axon terminals and some antibodies, the pixelimage plane specific for immunoreactive puncta was in-verted about the vertical midline, giving a quasirandomredistribution of puncta at the same average density. Thenumber of “random” colocalizations was then automati-cally calculated and compared to the numbers obtainedpreviously. The results of these analyses are summarizedin Table 1.
In most cases, the number of IR puncta in “random”colocalizations was lower than the number of IR puncta inactual colocalizations (P � 0.05, paired Wilcoxon rank-sum test). Thus, the density of colocalizations is not de-termined simply by the overall density of immunoreactivepuncta, but reflects specific spatial correlation betweenthe axon terminals and receptor clusters. However, con-sistent with previous studies (Fletcher et al., 2000) in the
Fig. 6. Schematic drawings of flat midget (FMB) and diffuse bipolar (DB3) axon terminals togetherwith colocalized immunoreactive puncta reconstructed from confocal sections. GluR, glutamate receptor.Scale bar � 5 �m.
Fig. 7. Schematic drawings of diffuse bipolar (DB6) and rod bipolar (RB) axon terminals together withcolocalized immunoreactive puncta reconstructed from confocal sections. GluR, glutamate receptor. Scalebar � 5 �m.
143GLUTAMATE RECEPTORS IN PRIMATE RETINA
case of RB/NR1C2� the number of puncta for “random”colocalizations was significantly higher than the numberof puncta for actual colocalizations, indicating that NMDAreceptor subunits are not located postsynaptically to RBterminals.
DISCUSSION
The purpose of the present study was to assess whetherdifferent types of bipolar cells are involved with differenttypes of glutamate receptors. Immunoreactivity for theAMPA receptor subunits GluR2/3 and GluR4 was associ-ated at high densities with the axons of all bipolar celltypes. The expression of the GluR1 subunit, the kainatereceptor subunits GluR6/7 and the NMDA receptorNR1C2� subunits varied between cell types.
Bipolar axons form ribbon synapses in the inner plexi-form layer and typically two processes are opposed to thesynaptic ribbon (dyad; Dowling and Boycott, 1966). Stud-ies in various mammalian retinae showed that immuno-reactivity for GluR subunits is usually located at only onepostsynaptic element of a dyad, indicating a diversity ofGluRs postsynaptic to individual bipolar terminals(Hartveit et al., 1994; Brandstatter et al., 1997; Boycottand Wassle, 1999; Qin and Pourcho, 1999a,b; Fletcher etal., 2000; Pourcho et al., 2001; Grunert et al., 2002). In thefollowing, we will discuss how the diversity of GluR sub-units found at bipolar axons can be correlated with thepostsynaptic profiles at bipolar dyads.
Rod bipolar cells
Consistent with previous studies of cat, rabbit, and ma-caque retinae (Qin and Pourcho, 1999a,b; Ghosh et al.,2001; Li et al., 2002), we found that RB axons are associ-ated with the AMPA receptor subunits GluR2/3 andGluR4 and to a much lower extent with the kainate recep-tor subunits GluR6/7 but not with NMDA receptors(Fletcher et al., 2000).
The two distinct sets of glutamate receptors at the RBdyad were identified recently (Ghosh et al., 2001; Li et al.,2002): AII amacrine cells express the AMPA receptor sub-units GluR2/3 and GluR4 and are clustered by thepostsynaptic density protein PSD-95, whereas the AI am-acrine cell expresses the �1/2 subunit, which is clusteredby the glutamate receptor-interacting protein (GRIP). Al-though the RB dyads is one of the best-studied synapses inthe retina (Zhang et al., 2002), much less is known aboutcone bipolar dyads and their postsynaptic elements.
FMB cells
FMB cells provide synaptic output to OFF-midget gan-glion cells and amacrine cells (Kolb and DeKorver, 1991;Calkins et al., 1994; Calkins and Sterling, 1996). Ourresults indicate that FMB axons are strongly involvedwith the AMPA receptor subunits GluR2, GluR2/3, GluR4and to a much lesser extent with the NR1C2� subunit ofthe NMDA receptor. The AMPA receptor subunit GluR1and the kainate receptor subunits GluR6/7 are probablynot expressed by postsynaptic elements at the FMB dyad.
The question arises whether the two postsynaptic ele-ments at the FMB dyad (which usually contains one gan-glion and one amacrine profile; Kolb and DeKorver, 1991;Calkins et al., 1994) express different sets of GluRs as hasbeen shown for the RB dyad (see above). Several lines ofevidence suggest that midget ganglion cells express
Fig. 8. Histograms showing the average number of colocalizedimmunoreactive (IR) puncta per axon terminal for flat midget bipolar(FMB), diffuse bipolar types DB3 and DB6, and rod bipolar (RB) cells.GluR, glutamate receptor.
144 U. GRUNERT ET AL.

AMPA receptors. Electrophysiological recordings frommacaque and baboon ganglion (including midget) cellsfound that responses were dominated by non-NMDA re-ceptors (Cohen and Miller, 1994; Zhou et al., 1994). Morerecently, Jacoby and Wu (2001) found that the non-NMDAreceptor responses in primate ganglion cells are mediatedby AMPA-preferring receptors with little if any involve-ment of kainate receptor. Consistent with this idea, ourresults from preembedding immunoelectron microscopyindicated that AMPA receptors in the IPL are predomi-nantly located on nonamacrine profiles (Grunert et al.,2002).
It possible that midget ganglion cells in addition toAMPA receptors also express NMDA receptors for thefollowing reasons. The electrophysiological studies men-tioned above also found some responses to NMDA, andNMDA receptors are located in the region of the IPLwhere midget bipolar and midget ganglion cells stratify(this study; Grunert et al., 2002). In situ hybridization andimmunohistochemical studies in other mammalian reti-nae, including primate have shown that ganglion cellsexpress AMPA and NMDA receptor subunits (Hamassaki-Britto et al., 1993; Brandstatter et al., 1994; Hof et al.,1998; Lagreze et al., 2000). In addition, double-labelingimmunocytochemistry has shown that AMPA and NMDAreceptor subunits can be coexpressed in primate retina(Grunert et al., 2002). Thus, it is possible that AMPA andNMDA receptor subunits are expressed at the samepostsynaptic profile (i.e., the midget ganglion cell) of theFMB dyad.
The number of AMPA receptor IR puncta colocalizedwith FMB cells is higher than the number of NR1C2� IRpuncta, suggesting that not all AMPA receptor subunitsare colocalized with NMDA receptors. The pure AMPAreceptors could be localized to the amacrine cell(s)postsynaptic at the FMB dyad, one of which is the AIIamacrine cell (Grunert, 1997). Consistently, electrophysi-ological recordings in slice preparations of other mamma-lian retinae have shown that AII amacrine cells expressAMPA receptors (Boos et al., 1993; Mørkve et al., 2002),whereas the question whether AII cells express NMDAreceptors is still controversial (Hartveit and Veruki, 1997;Zhou and Dacheux, 2001). It is also unclear whether thelobular appendages of AII cells express AMPA receptors atOFF cone bipolar dyads. A calcium imaging study ofmouse AII amacrine cells reported that ionotropic non-NMDA receptor agonists evoked a calcium increase in thearboreal dendrites but not in the lobular appendages(Habermann et al., 2000), suggesting that AMPA recep-tors are absent from lobular appendages. Double-labelingexperiments using markers for AII cells in combination
with antibodies to GluR receptor subunits could addressthis question anatomically.
The AII cell is only one of presumably a number ofamacrine cell types postsynaptic to FMB cells but theother types have not yet been identified. Previous studiesof the rod pathway found that the �1/2 subunits of theorphan receptor are localized postsynaptic at the RB dyadon amacrine cells that play a modulatory role (Ghosh etal., 2001; Li et al., 2002). As �1/2 subunits are foundthroughout the entire IPL (Grunert et al., 2002), theycould also be involved with the amacrine profiles postsyn-aptic to FMB terminals.
Diffuse bipolar cell DB3
Our results indicate that DB3 axons are presynaptic tothe AMPA receptor subunits GluR2, GluR2/3, and GluR4and to the NR1C2� subunit of the NMDA receptor. Thesefindings are consistent with the following anatomical andelectrophysiological data. The axons of DB3 cells in mar-moset and macaque retinae stratify in the same region asOFF-parasol cells and DB3 cells provide ribbon synapsesto OFF-parasol cells (Ghosh et al., 1997; Jacoby et al.,2000). Electrophysiological recordings (Cohen and Miller,1994; Zhou et al., 1994; Jacoby and Wu, 2001) and ana-tomic studies (Lin et al., 2002) suggest that parasol cellsexpress AMPA as well as NMDA receptors. Thus, ourresults support the notion that DB3 cells provide gluta-matergic input to parasol cells by means of AMPA andNMDA receptors.
Electron microscopy of calbindin-labeled DB3 cells inmarmoset and macaque retinae found that the postsynap-tic profiles of most DB3 dyads derive from an amacrineand a ganglion cell, with amacrine–amacrine andganglion–ganglion profiles being less frequent (Luo et al.,1999; Jacoby and Marshak, 2000). Jacoby and Marshak(2002) also found that AII amacrine cells are postsynapticto DB3 axons. Thus, AMPA receptors could be expressedby AII cells at DB3 dyads (but see Discussion above). Inconclusion, DB3 cells are presynaptic to AMPA andNMDA receptors but the specific arrangement of thesesubunits at the dyad needs to be investigated in moredetail.
Diffuse bipolar cell DB6
The axon terminals of DB6 cells were associated with allAMPA receptor subunits (GluR1-4) and the kainate recep-tor subunits GluR6/7 but not significantly with NMDAreceptors. The axons of DB6 cells stratify close to theganglion cell layer in the same region as RB cells (Boycottand Wassle, 1991; Chan et al., 2001a,b). Our recent anal-ysis of the synaptic output of this cell type revealed that
TABLE 1. Number and Density of Colocalized IR Puncta on Bipolar Cell Terminals: Comparison of Colocalized IR Punctain the Original and “Flipped” Sections
Cell type/GluRsubunit
Number ofconfocal sections
Number of colocalizedpuncta, before flip
Number of colocalizedpuncta, after flip
Pvalue Puncta/�m2
Number of axonterminals
FMB/GluR4 37 289 218 *** 0.32 5FMB/GluR2/3 30 168 100 *** 0.16 3FMB/NR1C2� 28 63 43 ** 0.1 3DB6/GluR6/7 24 54 25 ** 0.1 3DB6/NR1C2� 32 48 29 0.02 0.08 3DB3/NR1C2� 25 25 12 0.03 0.27 2FMB/GluR6/7 36 32 17 0.05 0.06 5RB/NR1C2� 38 162 148 0.45 0.12 4
IR, immunoreactive; GluR, glutamate receptor; FMB, flat midget bipolar; DB, diffuse bipolar; RB, rod bipolar. **P � 0.01, ***P � 0.001.
145GLUTAMATE RECEPTORS IN PRIMATE RETINA

the vast majority of synapses goes to amacrine cells withonly a small percentage going to a still unidentified gan-glion cell type (Jusuf et al., 2002).
Where are the different GluR subunits located at theDB6 dyad? As discussed above, AMPA receptor subunitsmight be predominantly associated with ganglion cellsand with the arboreal dendrites of AII amacrine cells.Thus, the lower number of colocalized IR puncta for theAMPA receptor subunits GluR2-4 on DB6 axon terminalscompared with FMB and DB3 axon terminals might beexplained by the finding that DB6 cells provide only littleoutput to ganglion cells.
The GluR1 subunit of the AMPA receptor was expressedat low density throughout the IPL with a concentration atthe inner and outer borders of the IPL (Grunert et al.,2002; Lin et al., 2002), indicating that only specific celltypes express this subunit. In situ hybridization studiesfound GluR1 expression to be stronger in the inner nu-clear layer than in the ganglion cell layer (Hamassaki-Britto et al., 1993). Thus, the GluR1 subunit might pref-erentially be expressed by amacrine cells and, thus, couldbe located postsynaptic at the DB6 dyad.
Small and large GluR6/7 IR puncta were also colocal-ized with DB6 axon terminals. We suggested previouslythat small puncta are located postsynaptic to synapticribbons, whereas large GluR6/7 IR puncta are aggregatedat unusual contacts postsynaptic to bipolar axons at “con-ventional” synapses not involving synaptic ribbons(Grunert et al., 2002). Our previous study also indicatedthat the large GluR 6/7 IR puncta were located on gan-glion cells rather than on amacrine cells and that RB cellsmight form the presynaptic elements to these largepuncta. The present study indicates that DB6 axon termi-nals might also be presynaptic at these unusual contacts.
ACKNOWLEDGMENTS
We thank Ana Lara and Petra Mross for excellent tech-nical assistance. We also thank Dr. Dale Wilson for assis-tance with software development.
LITERATURE CITED
Andressen C, Mai JK. 1997. Localization of the CD15 carbohydrate epitopein the vertebrate retina. Vis Neurosci 14:253–262.
Boos R, Schneider H, Wassle H. 1993. Voltage-and transmitter-gated cur-rents of AII-amacrine cells in a slice preparation of the rat retina.J Neurosci 13:2874–2888.
Boycott BB, Wassle H. 1991. Morphological classification of bipolar cells ofthe primate retina. Eur J Neurosci 3:1069–1088.
Boycott B, Wassle H. 1999. Parallel processing in the mammalian retina.Invest Ophthalmol Vis Sci 40:1313–1327.
Brandstatter JH, Hartveit E, Sassoe-Pognetto M, Wassle H. 1994. Expres-sion of NMDA and high-affinity kainate receptor subunit mRNAs inthe adult rat retina. Eur J Neurosci 6:1100–1112.
Brandstatter JH, Koulen P, Wassle H. 1997. Selective synaptic distribu-tion of kainate receptor subunits in the two plexiform layers of the ratretina. J Neurosci 17:9298–9307.
Calkins DJ, Sterling P. 1996. Absence of spectrally specific lateral inputs tomidget ganglion cells in primate retina. Nature 381:613–615.
Calkins DJ, Schein SJ, Tsukamoto Y, Sterling P. 1994. M and L cones inmacaque fovea connect to midget ganglion cells by different numbers ofexcitatory synapses. Nature 371:70–72.
Chan TL, Martin PR, Clunas N, Grunert U. 2001a. Bipolar cell diversity inthe primate retina: morphologic and immunocytochemical analysis of aNew World monkey, the marmoset Callithrix jacchus. J Comp Neurol437:219–239.
Chan TL, Martin PR, Grunert U. 2001b. Immunocytochemical identifica-tion and analysis of the diffuse bipolar cell type DB6 in macaquemonkey retina. Eur J Neurosci 13:829–832.
Cohen ED, Miller RF. 1994. The role of NMDA and non-NMDA excitatoryamino acid receptors in the functional organization of primate retinalganglion cells. Vis Neurosci 11:317–332.
Crooks J, Kolb H. 1992. Localization of GABA, glycine, glutamate andtyrosine hydroxylase in the human retina. J Comp Neurol 315:287–302.
Dacey DM. 2000. Parallel pathways for spectral coding in primate retina.Annu Rev Neurosci 23:743–775.
Davanger S, Ottersen OP, Storm-Mathisen J. 1991. Glutamate, GABA,and glycine in the human retina: an immunocytochemical investiga-tion. J Comp Neurol 311:483–494.
Dowling JE, Boycott BB. 1966. Organization of the primate retina: electronmicroscopy. Proc R Soc Lond B 166:80–111.
Fletcher EL, Hack I, Brandstatter JH, Wassle H. 2000. Synaptic localiza-tion of NMDA receptor subunits in the rat retina. J Comp Neurol420:98–112.
Ghosh KK, Martin PR, Grunert U. 1997. Morphological analysis of the bluecone pathway in the retina of a New World monkey, the marmosetCallithrix jacchus. J Comp Neurol 379:211–225.
Ghosh KK, Haverkamp S, Wassle H. 2001. Glutamate receptors in the rodpathway of the mammalian retina. J Neurosci 21:8636–8647.
Greferath U, Grunert U, Wassle H. 1990. Rod bipolar cells in the mam-malian retina show protein kinase C-like immunoreactivity. J CompNeurol 301:433–442.
Grunert U. 1997. Anatomical evidence for rod input to the parvocellularpathway in the visual system of the primate. Eur J Neurosci 9:617–621.
Grunert U, Martin PR. 1991. Rod bipolar cells in the macaque monkeyretina: immunoreactivity and connectivity. J Neurosci 11:2742–2758.
Grunert U, Martin PR, Wassle H. 1994. Immunocytochemical analysis ofbipolar cells in the macaque monkey retina. J Comp Neurol 348:607–627.
Grunert U, Haverkamp S, Fletcher EL, Wassle H. 2002. Synaptic distri-bution of ionotropic glutamate receptors in the inner plexiform layer ofthe primate retina. J Comp Neurol 447:138–151.
Habermann CJ, Wassle H, Protti DA. 2000. Ionotropic glutamate receptorsand voltage-dependent calcium channels in mouse AII amacrine cells.Soc Neurosci Abstr 26:248.
Hack I, Peichl L, Brandstatter JH. 1999. An alternative pathway for rodsignals in the rodent retina: rod photoreceptors, cone bipolar cells, andthe localization of glutamate receptors. Proc Natl Acad Sci U S A96:14130–14135.
Hack I, Frech M, Dick O, Peichl L, Brandstatter JH. 2001. Heterogeneousdistribution of AMPA glutamate receptor subunits at the photoreceptorsynapses of rodent retina. Eur J Neurosci 13:15–24.
Hamassaki-Britto DE, Hermans-Borgmeyer I, Heinemann S, Hughes TE.1993. Expression of glutamate receptor genes in the mammalian ret-ina: the localization of GluR1 through GluR7 mRNAs. J Neurosci13:1888–1898.
Hartveit E, Veruki ML. 1997. AII amacrine cells express functional NMDAreceptors. Neuroreport 8:1219–1223.
Hartveit E, Brandstatter JH, Sassoe-Pognetto M, Laurie DJ, Seeburg PH,Wassle H. 1994. Localization and developmental expression of theNMDA receptor subunit NR2A in the mammalian retina. J CompNeurol 348:570–582.
Haverkamp S, Wassle H. 2000. Immunocytochemical analysis of the mouseretina. J Comp Neurol 424:1–23.
Haverkamp S, Grunert U, Wassle H. 2000. The cone pedicle, a complexsynapse in the retina. Neuron 27:85–95.
Haverkamp S, Grunert U, Wassle H. 2001a. Localization of kainate recep-tors at the cone pedicles of the primate retina. J Comp Neurol 436:471–486.
Haverkamp S, Grunert U, Wassle H. 2001b. The synaptic architecture ofAMPA receptors at the cone pedicle of the primate retina. J Neurosci21:2488–2500.
Hof PR, Lee P-Y, Yeung G, Wang R-F, Podos SM, Morrison JH. 1998.Glutamate receptor subunit GluR2 and NMDAR1 immunoreactivity inthe retina of macaque monkeys with experimental glaucoma does notidentify vulnerable neurons. Exp Neurol 153:234–241.
Jacoby RA, Marshak DW. 2000. Synaptic connections of DB3 diffuse bipo-lar cell axons in macaque retina. J Comp Neurol 416:19–29.
146 U. GRUNERT ET AL.

Jacoby RA, Wu SM. 2001. AMPA-preferring receptors mediate excitatorynon-NMDA responses of primate ganglion cells. Vis Neurosci 18:703–710.
Jacoby RA, Wiechmann AF, Amara SG, Leighton BH, Marshak DW. 2000.Diffuse bipolar cells provide input to OFF parasol ganglion cells in themacaque retina. J Comp Neurol 416:6–18.
Jusuf PR, Lee SCS, Grunert U. 2002. Light and electron microscopicanalysis of a diffuse cone bipolar cell type (DB6) in primate retina. ProcAust Neurosci Soc 13:220.
Kolb H, DeKorver L. 1991. Midget ganglion cells of the parafovea of thehuman retina: a study by electron microscopy and serial section recon-structions. J Comp Neurol 303:617–636.
Kolb H, Linberg KA, Fisher SK. 1992. Neurons of the human retina: aGolgi study. J Comp Neurol 318:146–187.
Kouyama N, Marshak DW. 1992. Bipolar cells specific for blue cones in themacaque retina. J Neurosci 12:1233–1252.
Lagreze WA, Darstein M, Feuerstein TJ, Otto T, Landwehrmeyer GB.2000. N-methyl-D-aspartate receptor subunit mRNA expression in hu-man retinal ganglion cells. Graefes Arch Clin Exp Ophthalmol 238:486–490.
Lennie P, Trevarthen C, Van Essen D, Wassle H. 1990. Parallel processingof visual information. In: Visual perception: the neurophysiologicalfoundations. San Diego: Academic Press. p 103–128.
Li W, Trexler EB, Massey SC. 2002. Glutamate receptors at rod bipolarribbon synapses in the rabbit retina. J Comp Neurol 448:230–248.
Lin B, Martin PR, Grunert U. 2002. Expression and distribution of iono-tropic glutamate receptor subunits on parasol ganglion cells in theprimate retina. Vis Neurosci 19:453–465.
Luo X, Ghosh KK, Martin PR, Grunert U. 1999. Analysis of two types ofcone bipolar cells in the retina of a New World monkey, the marmoset,Callithrix jacchus. Vis Neurosci 16:709–719.
Macri J, Martin PR, Grunert U. 2000. Distribution of the �1 subunit of theGABAA receptor on midget and parasol ganglion cells in the retina ofthe common marmoset Callithrix jacchus. Vis Neurosci 17:437–448.
Marshak DW, Aldrich LB, Del Valle J, Yamada T. 1990. Localization ofimmunoreactive cholecystokinin precursor to amacrine cells and bipo-lar cells of the macaque monkey retina. J Neurosci 10:3045–3055.
Martin PR, Grunert U. 1992. Spatial density and immunoreactivity ofbipolar cells in the macaque monkey retina. J Comp Neurol 323:269–287.
Milam AH, Dacey DM, Dizhoor AM. 1993. Recoverin immunoreactivity inmammalian cone bipolar cells. Vis Neurosci 10:1–12.
Mills SL, Massey SC. 1999. AII amacrine cells limit scotopic acuity incentral macaque retina: a confocal analysis of calretinin labeling.J Comp Neurol 411:19–34.
Morigiwa K, Vardi N. 1999. Differential expression of ionotropic glutamatereceptor subunits in the outer retina. J Comp Neurol 405:173–184.
Mørkve SH, Veruki ML, Hartveit E. 2002. Functional characteristics ofnon-NMDA-type ionotropic glutamate receptor channels in AII ama-crine cells in rat retina. J Physiol 542:147–165.
Negishi K, Kato S, Teranishi T. 1988. Dopamine cells and rod bipolar cellscontain protein kinase C-like immunoreactivity in some vertebrateretinas. Neurosci Lett 94:247–252.
Nomura A, Shigemoto R, Nakamura Y, Okamoto N, Mizuno N, NakanishiS. 1994. Developmentally regulated postsynaptic localization of ametabotropic glutamate receptor in rat rod bipolar cells. Cell 77:361–369.
Pourcho RG, Qin P, Goebel DJ. 2001. Cellular and subcellular distributionof NMDA receptor subunit NR2B in the retina. J Comp Neurol 433:75–85.
Qin P, Pourcho RG. 1996. Distribution of AMPA-selective glutamate re-ceptor subunits in the cat retina. Brain Res 710:303–307.
Qin P, Pourcho RG. 1999a. AMPA-selective glutamate receptor subunitsGluR2 and GluR4 in the cat retina: an immunocytochemical study. VisNeurosci 16:1105–1114.
Qin P, Pourcho RG. 1999b. Localization of AMPA-selective glutamatereceptor subunits in the cat retina: a light-and electron-microscopicstudy. Vis Neurosci 16:169–177.
Qin P, Pourcho RG. 2001. Immunocytochemical localization of kainate-selective glutamate receptor subunits GluR5, GluR6, and GluR7 in thecat retina. Brain Res 890:211–221.
Silveira LCL, De Mello Jr., HD. 1998. Parallel pathways of primate vision.In: Chalupa LM, Finlay BL, editors. Development and organization ofthe retina. New York: Plenum Press. p 173–199.
Strettoi E, Dacheux RF, Raviola E. 1990. Synaptic connections of rodbipolar cells in the inner plexiform layer of the rabbit retina. J CompNeurol 295:449–466.
Vardi N, Duvoisin R, Wu G, Sterling P. 2000. Localization of mGluR6 todendrites of ON bipolar cells in primate retina. J Comp Neurol 423:402–412.
Wassle H. 1999. Parallel pathways from the outer to the inner retina inprimates. In: Gegenfurtner KR, Sharpe LT, editors. Color vision: fromgenes to perception. Cambridge: Cambridge University Press. p 145–162.
Zhang J, Li W, Trexler EB, Massey SC. 2002. Confocal analysis of recip-rocal feedback at rod bipolar terminals in the rabbit retina. J Neurosci22:10871–10882.
Zhou C, Dacheux RF. 2001. Voltage-and ligand gated currents of AIIamacrine cells in the rabbit retina. ARVO abstracts Invest OphthalmolVis Sci 42:3625.
Zhou ZJ, Marshak DW, Fain GL. 1994. Amino acid receptors of midget andparasol cells in primate retina. Proc Natl Acad Sci U S A 91:4907–4911.
147GLUTAMATE RECEPTORS IN PRIMATE RETINA
![Solitary Intraparotid Facial Nerve Plexiform Neurofibroma · peripheral nerve sheath tumor, which occurs in 2% - 5% of patients with plexiform neurofibroma [8]. Malignat peripheral](https://img.pdfslide.us/doc/110x75/5f7de695ec881b64331afe7f/solitary-intraparotid-facial-nerve-plexiform-neurofibroma-peripheral-nerve-sheath.jpg)

![Neuromyelitis optica spectrum disorder: a pediatric case report · 2017-09-15 · [4]. AQP4 is responsible for glutamate and potassium regulation in the BBB, synapses, and paranodes](https://img.pdfslide.us/doc/110x75/5f59b906dac5f12477718358/neuromyelitis-optica-spectrum-disorder-a-pediatric-case-2017-09-15-4-aqp4.jpg)
![Optical monitoring of glutamate release at multiple synapses ......of glutamate release at identified synapses in organised brain tissue [20, 21], including in vivo [22]. In the present](https://img.pdfslide.us/doc/110x75/60b6f5df1a6bec75153fd1b4/optical-monitoring-of-glutamate-release-at-multiple-synapses-of-glutamate.jpg)
























