Embed Size (px)
Citation preview

REVIEW
Genome-wide comparative analysis of tonoplast intrinsicprotein (TIP) genes in plants
Preetom Regon & Piyalee Panda & Erina Kshetrimayum &
Sanjib Kumar Panda
Received: 6 January 2014 /Revised: 13 July 2014 /Accepted: 21 July 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Tonoplast intrinsic proteins (TIPs) play a vital rolein water transport across membranes. In the present study, weperformed a comparative analysis of TIP genes in ten plantspecies including both monocots and dicots. A total of 100TIP aquaporin genes were identified, and their relationshipsamong the plant species were analyzed. Phylogenetic analysiswas performed to evaluate the relationship of these geneswithin the plant species. Based on the phylogenetic analysisresults, TIPs were classified into five distinct arbitrary groups(group I to group V), which represented TIP2, TIP5, TIP4,TIP1, and TIP3, respectively. Group I represented the largestarbitrary group, followed by group IV, in the phylogenetictree. The result clearly indicates that TIP2 and TIP1 areabundant aquaporins and highly related among the species.In the present review, a comparative study of gene structureanalysis between dicots and monocots has been performed toanalyze their structural variation. Most of the predicted motifsare conserved among the species, signifying an evolutionaryrelationship. The gene expression analysis indicated that theexpression of TIP genes varies during different developmentalstages and also during stressed conditions. The results indi-cated a great degree of evolutionary relationship and variationin the expression levels of TIPs in plants.
Keywords Aquaporin . Genome-wide analysis . Plants .
Tonoplast intrinsic protein
AbbreviationsAPQ AquaporinTIPs Tonoplast intrinsic proteinsCDS Coding DNA sequenceNPA Asparagine-Proline-Alanine
Introduction
Plant’s response to abiotic stress depends on several physio-logical factors. These responses are regulated by a wide arrayof genes, which are either upregulated or downregulated inresponse to stresses. Maintenance of water balance in cells iscritically important for plants to sustain cellular and functionalhomeostasis during stressed conditions. This process is regu-lated by a large number of aquaporin genes in plants.Aquaporins (AQPs) are water channels/major intrinsic pro-teins (MIPs), which facilitate the passive movement of waterand down the water potential gradient (Kruse et al. 2006;Forrest and Bhave 2008). In plants, AQPs regulate the cellvolume and internal osmotic pressure by facilitating highlyselective passive water transport. Both plasma membrane andtonoplast AQPs are vital for maintenance of the proper cyto-solic osmolarity in plants (Kaldenhoff and Fischer 2006). TheMIPs have a common molecular weight of 23–31 kDa, sixtransmembrane helices, asparagine-proline-alanine (NPA)motifs, and cytosolic C- and N-terminal. NPA motifs arewidespread in all the aquaporins (Fig. 1).
AQPs are abundant in plants, and based on localiza-tion, they are divided in several distinct groups (Maurelet al. 2008; Gomes et al. 2009). These include plasmamembrane intrinsic proteins (PIPs), tonoplast intrinsicproteins (TIPs), nodulin 26-like intrinsic proteins(NIPs), and small basic intrinsic proteins (SIPs)(Johanson et al. 2001). Further, three other groups of
Electronic supplementary material The online version of this article(doi:10.1007/s10142-014-0389-9) contains supplementary material,which is available to authorized users.
P. Regon (*) : P. Panda : E. Kshetrimayum : S. K. PandaPlant Molecular Biotechnology Laboratory, Department of LifeScience andBioinformatics, AssamUniversity, Silchar 788011, Indiae-mail: [email protected]
Funct Integr GenomicsDOI 10.1007/s10142-014-0389-9

the MIP subfamily which includes GlpF-like intrinsicproteins (GIPs) (Gustavsson et al. 2005), X-intrinsicproteins (XIPs), and hybrid intrinsic proteins (HIPs)(Danielson and Johanson 2008) were reported. In plants,TIPs are abundant and facilitate high water permeabilityfor the tonoplast. Based on their sequence homology,TIPs were further classified into five distinct groups,which include TIP1 (TIP1;1, TIP1;2, and TIP1;3),TIP2 (TIP2;1, TIP2;2, and TIP2;3), TIP 3 (TIP3;1 andTIP3;2), TIP4 (TIP4;1), and TIP5 (TIP5;1) (Johansonet al. 2001). In addition to their role as water channelproteins, TIPs also transport hydrogen peroxide (H2O2)and glycerol (Maeshima 2001). Studies have shown that TIPsregulate water flow in response to drought and salinity stressin Arabidopsis thaliana (Kaldenhoff and Fischer 2006;Alexandersson et al. 2005; Boursiac et al. 2005).
The localization of TIP aquaporins is in the tonoplastof the vacuolar membrane which is extremely permeableto water with up to 100-fold higher osmotic permeabil-ities than those of the plasma membrane (Maurel et al.1997). The phenomenon can make up as much as 40 %of the total tonoplast function (Higuchi et al. 1998).Overall, understanding the greater water permeabilityand its abundances, the present study is mainly focusedon the TIP aquaporins and the availability of genomesequences of a variety of plant species, providing us theopportunity to carry out various in silico experiments atthe sequence level. This comparative study is taken upto develop the basic phylogenetic and structural rela-tionship of TIP aquaporin genes among the plant
species. In addition, in silico expression analysis willhelp us to understand the basic expression pattern ofTIP aquaporin genes in various developmental stagesand environmental stress conditions.
Analytical approach
The coding sequences of A. thaliana were downloaded fromthe TAIR (http://www.arabidopsis.org/) (Lamesch et al. 2012)database. The Phytozome (http://www.phytozome.net/)(Goodstein et al. 2012) database was used to download theCDS of Brachypodium distachyon, Citrus sinensis, Ricinuscommunis, and Sorghum bicolor. The Plaza Bioinformatics(http://bioinformatics.psb.ugent.be/plaza/) (Proost et al.2009) database was used to download the CDS of Medicagotruncatula, Zea mays, Oryza sativa, and Fragaria vescawhereas the CocoaGenDB (http://cocoagendb.cirad.fr)database was used to obtain the CDS of Theobroma cacao.The aquaporin genes were identified by performing keyword“aquaporin” search in the abovementioned databases. Usingthe resulting genes as a query, stand-alone BLASTn searchwas performed against the downloaded CDS sequences bysetting up the E-value to 1e−6. The resulting and nonredun-dant sequences were annotated using the Blast2GO (Conesaet al. 2005) functional annotation suite. During the stand-alone BLAST (Altschul et al. 1997) search, the well-annotated aquaporin genes of Arabidopsis were used as acommon query for all. From the identified aquaporin genes,the TIP aquaporins were taken for our study.
Fig. 1 An overview of the MIP structure. a The secondary structure of MIPs with their helices. b The 3-D structure of MIPs with two NPA boxes(PDBID 1IH5)
Funct Integr Genomics

To perform the phylogenetic analysis, the amino acid se-quences were aligned with Clustal X 2.0 (Larkin et al. 2007).Using the aligned dataset, a maximum likelihood phylogenet-ic tree was reconstructed through raxmlGUI v. 1.3 (Silvestroand Michalak 2011) under the PROTGAMMA distributionmodel and Jones-Taylor-Thornton (JJT) substitutionmatrix. Atotal of 100 rapid bootstrap replicates were conducted forsupport estimation. FigTree (ver. 1.4) was used to visualizethe resulted phylogenetic tree (Rambaut 2012).
The localization of TIP aquaporins was determined basedon their position on chromosomes corresponding to their locusnumbers and regions in the TAIR genome database forA. thaliana and Plaza Bioinformatics for Z. mays,M. truncatula, and F. vesca. The nature of the duplication ofTIP aquaporin genes of Z. mays, M. truncatula, and F. vescaalong with A. thaliana was also determined using theWorkbench of Plaza Bioinformatics and TAIR database,respectively.
The structure of TIP aquaporin genes of the plant specieswas constructed by comparing the CDS sequences and fullgenomic sequences of the genes. During the analysis, intronphases in between the exon-intron junction were obtained byusing Gene Structure Display Server (http://gsds.cbi.pku.edu.cn) (Jami et al. 2012).
The InterPro (www.ebi.ac.uk/interpro) domainanalysis tool was used to identify the domain of theprotein sequences. For motif analysis, the amino acidsequences were aligned through Clustal X 2.0. Theover-represented motifs were also identified throughthe MEME Suite by setting up the E-value of 0.01and motif length up to 20–60 (Bailey and Elkan1994). Resulted motifs were also analyzed throughSMART (http://smart.embl-heidelber.de) to obtain theirdomain information (Schultz et al. 1998).
In silico gene expression analysis of A. thaliana, Z. mays,and O. sativa was carried out through GENEVESTIGATOR
v.3 (https://www.genevestigator.com/) (Hruz et al. 2008). Toobtain the expression data of Z. mays, a BLASTp search wasperformed in theNCBI database. Reference sequence accessionIDs were used to retrieve the expression data for Z. mays.
Identification of TIP aquaporin genes
With the discovery of AQPs and extensive progress ingenome sequencing, we are now able to identify a largenumber of aquaporins at the DNA sequence level. Thefirst AQP-like sequences that were identified in plantsinclude a representative of the TIP subfamily (Johnsonet al. 1990). In the present study, we found importantAQPs in C. sinensis, F. vesca, M. truncatula, andA. thaliana from their respective genome. From theseAQP genes, TIP aquaporin subfamilies were identifiedand assorted for further analysis. Results show a largenumber of TIP aquaporin subfamily in C. sinensis,F. vesca, A. thaliana, Z. mays, and S. bicolor as com-pared to other plant species. Previous reports show thatthere are 35 aquaporins present in Arabidopsis(Johanson et al. 2001) and 31 in Z. mays (Chaumontet al. 2001). Another report also shows that theArabidopsis genome contains 38 aquaporins including11 TIPs (Quigley et al. 2002). In this investigation,we have identified 38 aquaporin genes in Arabidopsis(Supplementary material 1). The highest numbers ofaquaporins were observed in C. sinensis (57), Z. mays(43), and F. vesca (42). Among them, the highest num-ber of TIPs was found in S. bicolor (13) and the lowestnumber of TIPs was identified in R. communis (7).Overall, a total of 398 AQP genes and 100 TIP aqua-porin genes were identified from these ten differentplants including both monocots and dicots (Table 1).
Table 1 Distribution of TIPaquaporin genes in different plantspecies
Sl. no. Organism name Aquaporin genes Total no. of TIPs Source
1 Arabidopsis thaliana 38 11 TAIR
2 Brachypodium distachyon 33 10 Phytozome
3 Citrus sinensis 57 11 Phytozome
4 Fagaria vesca 42 10 Plaza Bioinformatics
5 Medicago trunculata 36 8 Plaza Bioinformatics
6 Oryza sativa (Japonica group) 39 10 Plaza Bioinformatics
7 Ricinus communis 37 7 Phytozome
8 Sorghum bicolor 40 13 Phytozome
9 Theobroma cacao 33 8 CocoaGenDB
10 Zea mays 43 12 Plaza Bioinformatics
Total 398 100
Funct Integr Genomics
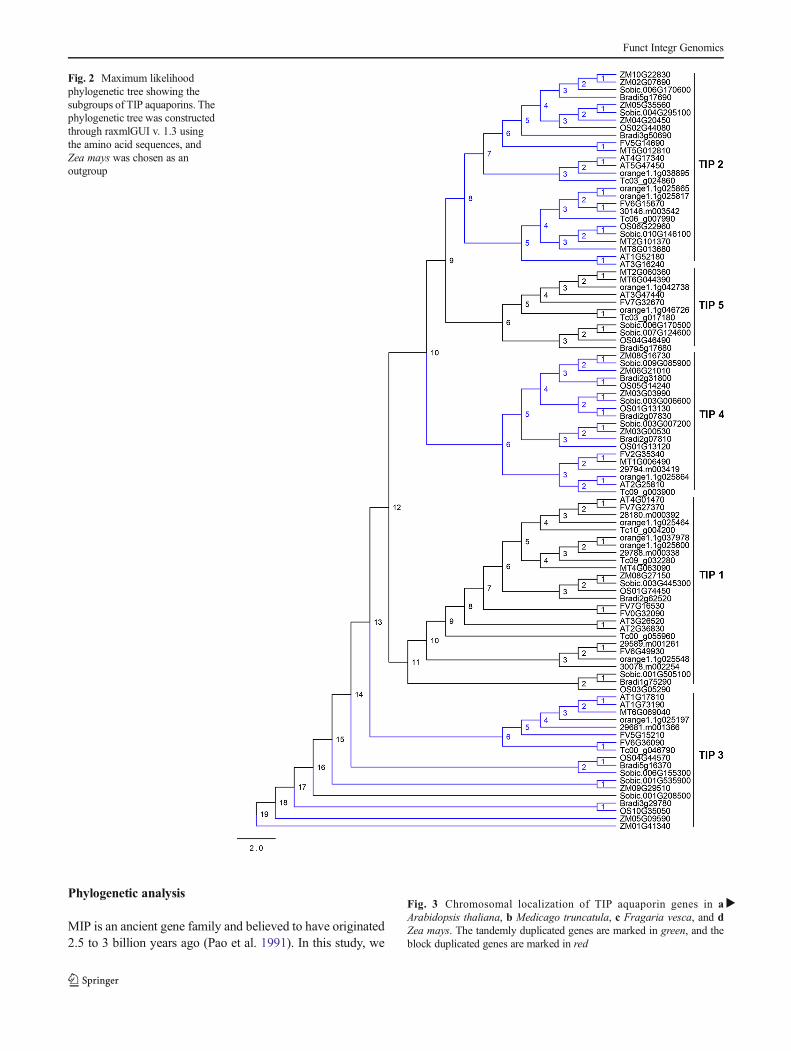
Phylogenetic analysis
MIP is an ancient gene family and believed to have originated2.5 to 3 billion years ago (Pao et al. 1991). In this study, we
Fig. 2 Maximum likelihoodphylogenetic tree showing thesubgroups of TIP aquaporins. Thephylogenetic tree was constructedthrough raxmlGUI v. 1.3 usingthe amino acid sequences, andZea mays was chosen as anoutgroup
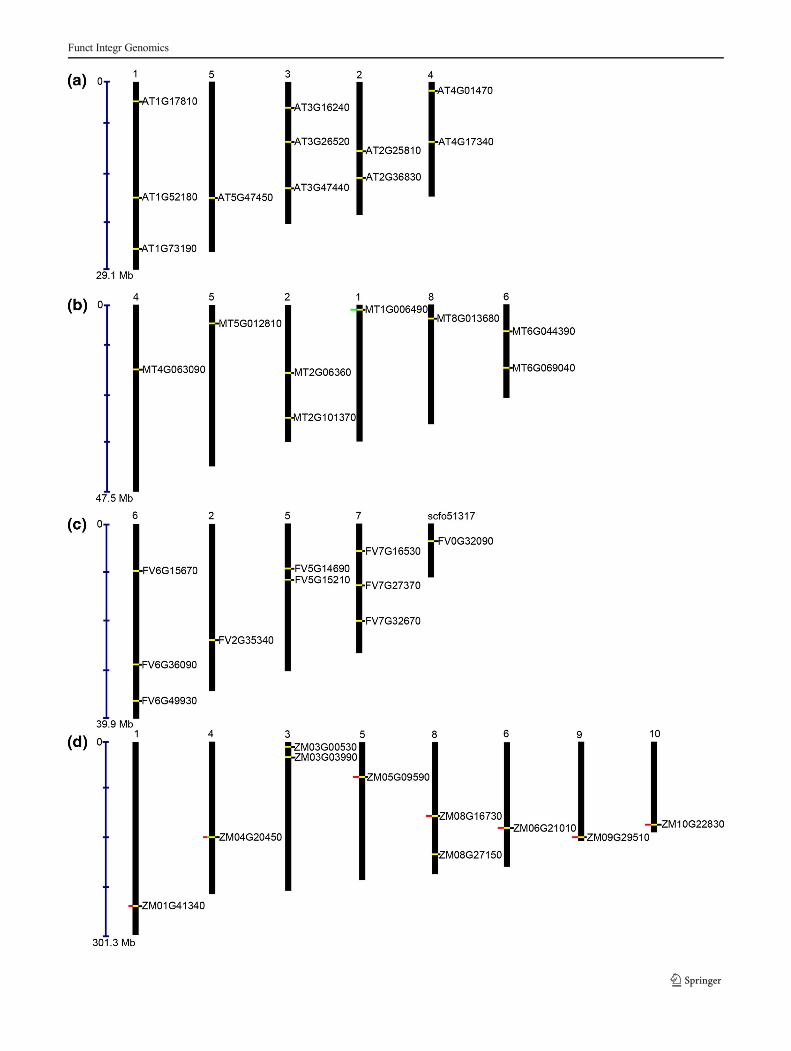
�Fig. 3 Chromosomal localization of TIP aquaporin genes in aArabidopsis thaliana, b Medicago truncatula, c Fragaria vesca, and dZea mays. The tandemly duplicated genes are marked in green, and theblock duplicated genes are marked in red
Funct Integr Genomics
Funct Integr Genomics

performed a phylogenetic study to assist the relationship ofTIP aquaporins among the plant species. In order to infer theevolution of a gene, it is necessary to relate it to the evolutionof plants (Danielson and Johanson 2010). In this study, themain prospective of the phylogenetic investigation was onlyto find out the relationship of TIP aquaporins among thespecies. We reevaluated the relationship of TIPs among theplant species at the amino acid sequence level. The resultedtree is composed of five distinct arbitrary groups. Group I isthe largest arbitrary group in the phylogenetic treerepresenting the whole TIP2 isoforms. The second arbitrarygroup represents the TIP5 isoform of aquaporins, and it is thesmallest arbitrary group in the phylogenetic tree, followed bygroup III, group IV, and group V which represent the TIP4isoform, TIP1 isoform, and TIP3 isoform of aquaporins, re-spectively. A comprehensive study of all the arbitrary groupsshowed that the TIP2 class of aquaporin genes is more closelyrelated to the TIP5 class of aquaporin genes. Further, the TIP5aquaporin is also related with TIP4 rather than TIP1 and TIP3.The phylogenetic tree also revealed that TIP3 and TIP1 aqua-porins are closely related and more diverged as compared toTIP2 and TIP5. As the TIP2 and TIP1 isoforms produced thelargest arbitrary groups in the phylogenetic tree, these twotypes of aquaporins are found to be present in greater numberin the genome as compared to others. On the other hand, theTIP5 isoform is found to be rare in number in each of the plantspecies (Fig. 2).
Chromosomal distribution and gene duplication
For the chromosomal distribution of the TIP family inA. thaliana, Z. mays, M. truncatula, and F. vesca, the 5′-3′coordinates of each of the genes were obtained from theirrespective genome databases. Eleven TIPs were distributedalong the entire (1–5) chromosome of A. thaliana, while threeTIPs were equally located on chromosomes 1 and 3. InM. truncatula, eight TIPs were distributed over six chromo-somes (chromosome numbers 1, 2, 4, 5, 6, and 8). Maximumof two TIPs were equally distributed on chromosome 2 and6. InF. vesca, TIPs were distributed on four chromosomes andone gene on a small scaffold. A maximum of three TIPs wereequally distributed on chromosome 6 and 7 while two TIPs
were distributed on chromosomes 5. In Z. mays, a total of 10TIPs were distributed among eight chromosomes, two each onchromosomes 3, and 8 while one each on chromosomes 1, 4,5, 6, 9, and 10. The gene duplication analysis showed that onegene of M. truncatula has undergone tandem duplication onchromosome 1. In the case of Z. mays, 7 out of 10 genesundertook a block duplication event and one gene,ZM04G20450, participated in tandem duplication as well asin the block duplication event and is located on chromosome 4in Z. mays (Fig. 3).
Analysis of gene structure
In order to investigate the structural relationship of TIP aqua-porins between monocots and dicots, the gene structures wereanalyzed by comparing the CDS and genomic sequences offour plant species. The analysis of exon-intron organizationshows similarities among the species. The number of intronsvaried from 0 to 3, and depending on the exon-intron numbersand positions, the gene structure pattern was distinguishedinto five distinct patterns (Fig. 4).
Pattern I was observed only in the monocots, which con-sists of a single intron. Therefore, pattern I was adopted as anideal pattern in monocotyledonous plants. Pattern II was ob-served in both monocots and dicots, which is composed oftwo introns. Most of the dicot gene organizations fall in thiscategory that indicates the pattern that is common amongdicots. Very few numbers of monocot genes were found inthis splicing pattern which is found to be participating in theblock duplication event in the earlier gene duplication study.Therefore, it was assumed that the splicing pattern in mono-cots is possibly acquired through the gene duplication eventduring the course of evolution. Two intronless genes wereobserved in Arabidopsis (AT4G01470) and Brachypodium(Bradi2g07810) which is a characteristic of primitive plants.It was reported that intronless genes may occur due to thesequential loss of introns during evolution (Roy and Penny2007). Following this assumption, pattern III might evolvedue to the sequential loss of introns. This sequential loss ofintrons was observed only in dicots. Two other types ofsplicing patterns were observed only in M. truncatula.Pattern IV was seen only in a single gene of Medicago.
Fig. 4 Exon-intron patterns ofTIP aquaporins found inmonocots and dicots
Funct Integr Genomics
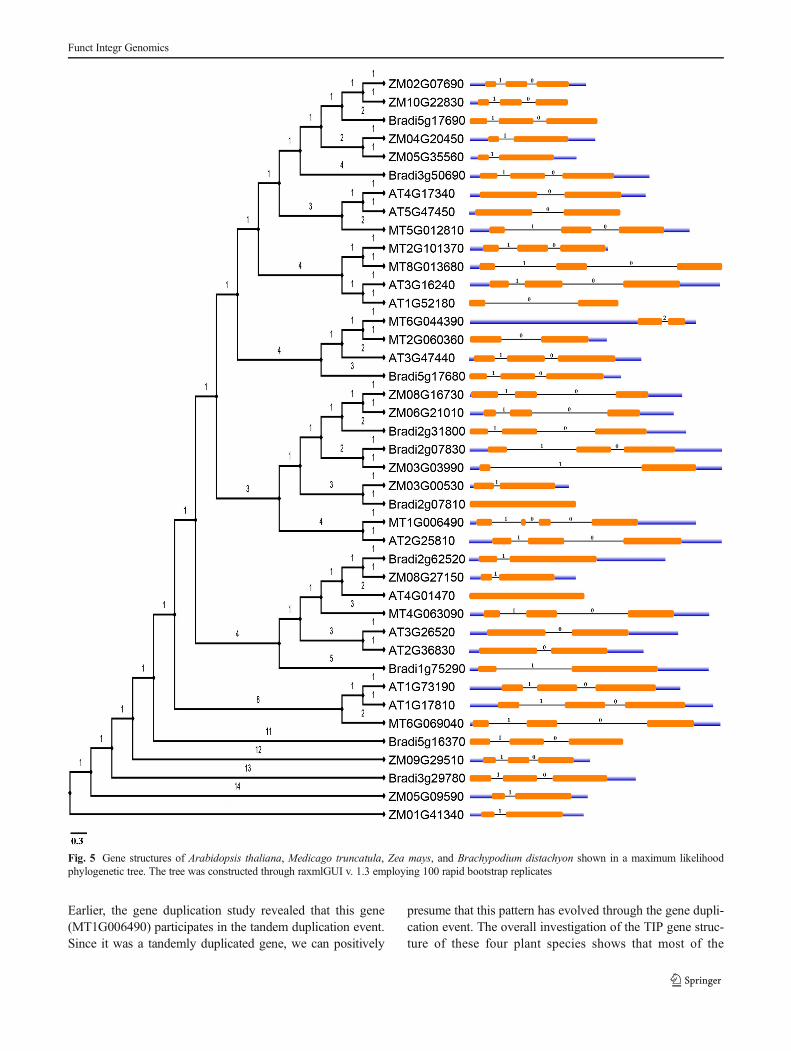
Earlier, the gene duplication study revealed that this gene(MT1G006490) participates in the tandem duplication event.Since it was a tandemly duplicated gene, we can positively
presume that this pattern has evolved through the gene dupli-cation event. The overall investigation of the TIP gene struc-ture of these four plant species shows that most of the
Fig. 5 Gene structures of Arabidopsis thaliana, Medicago truncatula, Zea mays, and Brachypodium distachyon shown in a maximum likelihoodphylogenetic tree. The tree was constructed through raxmlGUI v. 1.3 employing 100 rapid bootstrap replicates
Funct Integr Genomics

monocots have a single intron in their genes and dicots have aminimum of two introns, although intron variation was ob-served in dicots ranging from 0 to 3. As compared to the otherplants,Medicago has the highest numbers of intron variations;as a result, it produced the maximum number of splicingpattern. Therefore, Medicago is the most advanced andevolved plant species among these four plants. It was alsoobserved that the gene structure of the monocots also resem-bled that of the dicots, but no single gene of dicots wasobserved to fall under the splicing pattern of the monocots.Therefore, it is suggested that the monocots are more primitivethan the dicots and they may have evolved from a commonancestor (Fig. 5).
Domain and motif identification
The domain represents the sequential feature, which acts as adistinct structural and functional unit of a protein. The domainorganization was predicted through the InterPro domain anal-ysis tool for the identified TIP aquaporin. Our results indicatethat all the TIPs belong to the sameMIP domain (IPR000425).NPA is the common motif that is present in the AQP genefamily. Most of the aquaporin genes have either one or twoNPA motifs in their sequences. We have also observed thevariation of the NPA motif among the protein sequences. Wefurther observed four aquaporin genes without the NPAmotif,e.g., AT1G52180 (NCBI ref seq. NP_175629) in Arabidopsis,FV6G36090 in F. vesca, MT6G044390 (NCBI ref seq.XP_003619188) in M. truncatula, and 007G124600 inS. bicolor (Supplementary material 2). This suggests thatalthough the NPA motif is common, it may not necessarilyoccur among all the aquaporin genes. Using the MEME Suite,we identified the over-represented motif in the 100 TIP aqua-porin protein sequences among ten plant species along withfive different motifs among the protein sequences (Table 2).
Motifs 2 and 3 were found to be the most common andconserved motifs among the genes, and the conserved motifscould be the essential elements to determine the TIP aquaporinfamily and share a common molecular function among theplant species (Supplementary material 3). As detected bySignalP, motifs 1, 2, and 4 are the part of the signal peptidewhich are present at the N-terminus of the protein, and those
regions of a protein are the significant factor for determiningtheir transport system. Motifs 3 and 5 are found to be the partof transmembrane helix regions. The presence of similarmotifs among the sequences also suggested that the genesare evolutionary conserved and are related to each other.
Gene expression analysis
TIP aquaporin genes regulate greater water transport,which also implicates their involvement in the regula-tion of water flow in response to osmotic changes(Boursiac et al. 2005). Though aquaporins are believedto be the main water transporter protein in plants, theirpresence and expression may vary from species to spe-cies. In silico gene expression analysis of A. thaliana,O. sativa, and Z. mays were performed by using theGENEVESTIGATOR tool (www.genevestigator.com/gv/)(Hruz et al. 2008; Ye et al. 2013). In the case of Z.mays, for each of the proteins, a reference sequenceaccession ID was obtained by performing the BLASTpprogram of NCBI and the corresponding reference sequenceID was used to view their expression level throughGENEVESTIGATOR (Table 3).
Table 2 Identified motifs in TIP aquaporins through the MEME Suite
Motif Width Best possible match1 60 MEIVITFALVYTVYATAADPKKGSLGTIAPIAIGFIVGANILAAGPFSGGSMNPARSFGP2 50 VRTIGPLLTGLIVGANSLAGGNFTGASMNPARSFGPAMATGVWTNHWVYW3 60 FEIVMTFGLVYTVYATAVDPKKGNIGIIAPIAIGFIVGANILAGGAFDGASMNPAVSFGP4 57 FEIVMTFGLVYTVYATAVDPKKGSLGTIAPIAIGFIVGANILAGGAFDGASMNPAVS5 57 FEIIITFGLVYTVYATAADPKKGSLGTIAPIAIGFIVGANILAAGPFSGGSMNPARS
Table 3 Reference sequence IDs of TIP aquaporins of Zea mays thatwere used to invest igate thei r gene express ion throughGENEVESTIGATOR
Gene accession RefSeq ID E-value Identity (%)
ZM01G41340 NM_001254792.1 0.0 100
ZM03G00530 NM_001112171.1 5e−162 100
ZM03G03990 NM_001111565.1 3e−171 98
ZM04G20450 NM_001111560.1 5e−171 100
ZM05G09590 NM_001111562.1 0.0 100
ZM05G35560 NM_001111561.1 6e−173 100
ZM06G21010 NM_001111563.1 1e−81 64
ZM08G16730 NM_001111564.1 9e−180 99
ZM08G27150 NM_001111559.1 7e−175 99
ZM09G29510 NM_001153458.1 0.0 100
ZM10G22830 NM_001111437.1 2e−173 100
Funct Integr Genomics

The genes AT2G36830 (TIP1;1), AT3G16240 (TIP2;1),and AT3G26520 (TIP1;2) were highly expressed in the earlystages of growth development in A. thaliana. These threegenes were observed to express highly until the floweringstage and were gradually downregulated. AT1G17810(TIP3;2) and AT1G73190 (TIP3;2) were highly expressed
during the period of senescence, while other TIP aquaporingenes were expressed almost at the same level during eachdevelopmental stage. In O. sativa, the genes OS6G22960(TIP2;2), OS03G05290 (TIP1;1), and OS05G14240(TIP4;1) were found to be highly expressed throughout itsdevelopmental stages while the genes OS01G74450 (TIP1;2),
Fig. 6 Expression of TIP aquaporin genes in various developmental stages in a Arabidopsis thaliana, b Zea mays, and c Oryza sativa
Funct Integr Genomics
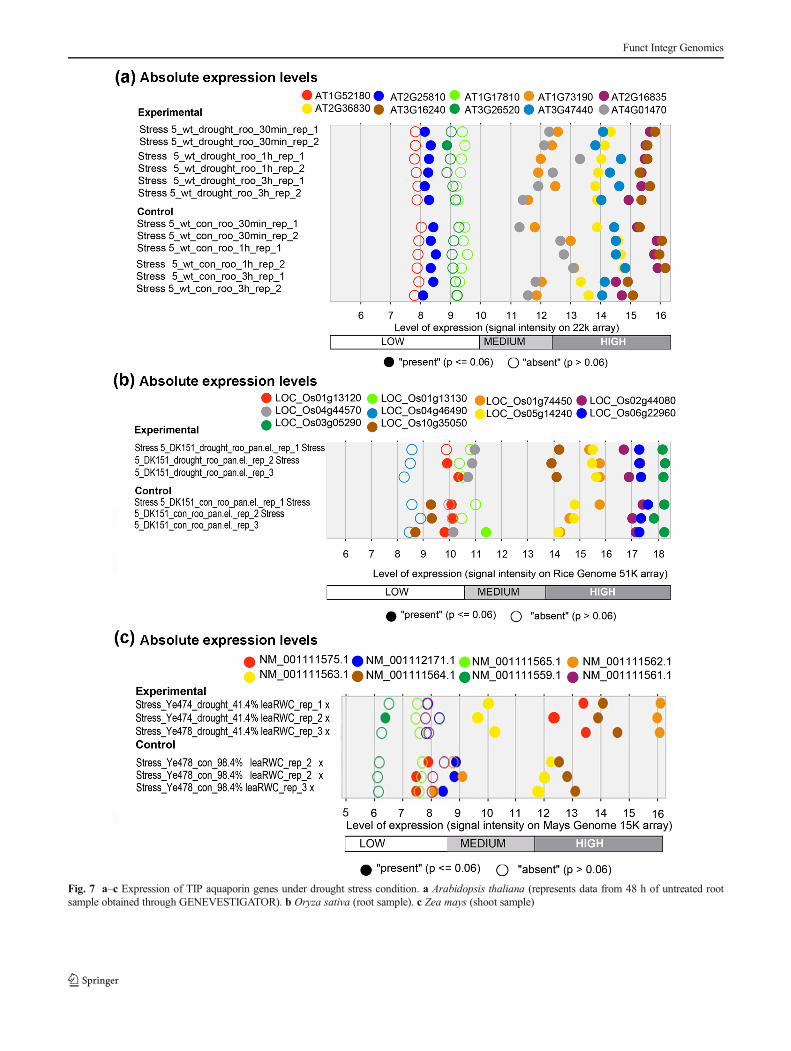
Fig. 7 a–c Expression of TIP aquaporin genes under drought stress condition. a Arabidopsis thaliana (represents data from 48 h of untreated rootsample obtained through GENEVESTIGATOR). b Oryza sativa (root sample). c Zea mays (shoot sample)
Funct Integr Genomics

OS01G13120 (TIP4;1), and OS01G13130 (TIP4;1) weremodera te in express ion. In Z. mays , the genesZM03G00530 /NM_0011121171 .1 (TIP4 ;4 ) andZM08G16730/NM_001111564.1 (TIP4;2) were progressive-ly upregulated throughout the developmental stages andZM05G09590/NM_001111562.1 (TIP3;1) was highlyexpressed during the dough stage of the development (Fig. 6).
Under different environmental conditions, developmentof plant growth is affected during their life cycle. Theexpression of the aquaporin gene, being the main watertransporter, is crucial under environmental stress conditions.Analysis of TIP aquaporins under drought conditions inA. thaliana showed that AT2G36830 (TIP1;1), AT3G16240(TIP2;1), AT4G17340 (TIP2;2), and AT3G26520 (TIP1;2)were highly expressed while AT2G25810 (TIP4;1) andAT5G47450 (TIP2;3) were moderate in expression. Thesesame genes were found to be highly expressed even undersalt stress condition. In O. sativa, OS03G05290 (TIP1;1),OS6G22960 (TIP2 ;2 ) , OS02G44080 (TIP2 ;1 ) ,OS01G74450 (TIP1;2), OS05G14240 (TIP4;1), andOS10G35050 (TIP3;1) were found to be highly expressed
under drought condition. In this condition, the geneOS10G35050 (TIP3;1) was found to be high in expressionas compared to its normal expression level. Generally, theexpression of the gene OS01G74450 (TIP1;2) was very low,but under drought condition, it seemed to be highlyexpressed. This indicated that this gene is particularly induc-ible under drought condition (Fig. 7).
In salt stress condition, the same genes were found to behighly expressed in O. sativa. But the gene OS01G74450,which was particularly expressed in drought condition, wasfound to be very low in expression under salt stress condition.A better expression under drought condition was observed inZ. mays. The genes ZM05G09590/NM_001111562.1 (TIP3;1)and ZM08G16730/NM_001111564.1 (TIP4;2) were foundto be highly expressed only in drought condition, whereasthe gene ZM06G21010/NM_001111563.1 (TIP4;1) wasfound to be downregulated in this condition (Fig. 8).
The tissue anatomical expression pattern of TIP aquaporinsin Arabidopsis shows that the root cortex protoplast, rootendodermis, and root phloem protoplast are the suitable re-gions for expression. The genes AT2G36830 (TIP1;1),
Fig. 8 a–b Expression of TIP aquaporins under salt stress condition. a Arabidopsis thaliana (represents data from the root epidermis and quiescentcenter protoplast sample). b Oryza sativa (root sample)
Funct Integr Genomics

AT3G16240 (TIP2;1), and AT3G26520 (TIP2;1) were foundto be highly expressed in most of the tissues except theendosperm, suspensor, testa, and seed coat. The genesAT3G47440 (TIP5;1) and AT4G01470 (TIP1;3) were foundto be well expressed only in the sperm cell, stamen, and pollenparts of tissue, whereas the genes AT1G17810 (TIP4;1) andAT1G73190 (TIP3;1) were found to be expressed only in theendosperm region (Supplementary material 4). TIP isoformsTIP1;1, TIP2;1, and TIP2;2 are well known to be expressed inseeds (Gattolin et al. 2011), roots, and aerial parts of the plant(Ludevid et al. 1992; Beebo et al. 2009) but not in embryos(Gattolin et al. 2009). In the case of O. sativa, the roots, roottip, peduncle, sheath, and leaf are the suitable areas of geneexpression and the genes OS03G05290 (TIP1;1),OS06G22960 (TIP2;2), and OS05G14240 (TIP4;1) werefound to be well expressed in most of the tissues butnot in the embryos (Supplementary material 5). In con-t r a s t , t h e g en e s OS10G35050 (T IP3 ; 1 ) andOS04G44570 (TIP3;2) were found to be only expressedin the embryo, endosperm, caryopsis, spikelet, and cal-lus. In Z. mays, the root tip, lateral root, foliar leaf,glume, and pollen parts are found to be the suitableareas of gene expression (Supplementary material 6) andthe genes ZM08G16730/NM_001111564.1 (Tip4;2) andZM06G21010/NM_001111563.1 (TIP4;1) were wellexpressed in most of the tissues. In addition, the geneZM05G09590/NM_001111562.1 (TIP3;1) was found tobe expressed only in the embryo and caryopsis. Acomparative study shows that the TIP3 isoform of thesethree species was expressed only in the embryo andcaryopsis but not in others. This suggests that theTIP3 isoform is embryo and caryopsis specific in ex-pression. It was also only detected in maturing andmature seeds (Gattolin et al. 2011).
Conclusion
We performed a comprehensive analysis of TIP aquaporingenes in different plant species covering phylogeny, chromo-somal localization, gene structures, and in silico expression.The exon-intron structure and motif compositions are found tobe relatively conserved in each subgroup. The highest numberof splicing pattern was observed in M. truncatula which is acharacteristic of evolving plant species. The gene expressionanalysis of TIPs in Arabidopsis, Z. mays, and O. sativashowed differential expression during different developmentalstages and under environmental stress. The roots, pericycle,vascular bundles, and embryo were found to be the predom-inant areas for high gene expression. Gene expression underdrought stress in Z. mays also showed that some genes aremore inducible in response to their environmental conditions.
References
Alexandersson E, Fraysse L, Sjovall-Larsen S, Gustavsson S, Fellert M,Karlsson M, Johanson U, Kjellbom P (2005) Whole gene familyexpression and drought stress regulation of aquaporins. Plant MolBiol 59:469–484
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new gener-ation of protein database search programs. Nucleic Acids Res 25:3389–3402
Bailey TL, Elkan C (1994) Fitting a mixture model by expectationmaximization to discover motifs in biopolymers. Proceedings ofthe second international conference on 132 intelligent systems formolecular biology. AAAI Press, Menlo Park, pp 28–36
Beebo A et al (2009) Life with and without AtTIP1;1, an Arabidopsisaquaporin preferentially localized in the apposing tonoplasts ofadjacent vacuoles. Plant Mol Biol 70:193–209. doi:10.1007/s11103-009-9465-2
Boursiac Y, Chen S, Luu DT, Sorieul M, van den Dries N, Maurel C(2005) Early effects of salinity on water transport in Arabidopsisroots. Molecular and cellular features of aquaporin expression. PlantPhysiol 139:790–805
Chaumont F, Barrieu F, Wojcik E, Chrispeels MJ, Jung R (2001)Aquaporins constitute a large and highly divergent protein familyin maize. Plant Physiol 125:1206–1215
Conesa A, Gotz S, Garcia-Gomez JM, Terol J, TalonM, RoblesM (2005)Blast2GO: a universal tool for annotation, visualization and analysisin functional genomics research. Bioinformatics 21(18):3674–3676
Danielson JAH, Johanson U (2008) Unexpected complexity of the aqua-porin gene family in the moss Physcomitrella patens. BMC PlantBiol 8:45
Danielson JAH, Johanson U (2010) Phylogeny of major intrinsic pro-teins. Adv Exp Med Biol 679:19–31
Forrest KL, BhaveM (2008) The PIP and TIP aquaporins in wheat form alarge and diverse family with unique gene structures and function-ally important features. Funct Integr Genom 8:115–133
Gattolin S, Sorieul M, Hunter PR, Khonsari RH, Frigerio L (2009) Invivo imaging of the tonoplast intrinsic protein family in Arabidopsisroots. BMC Plant Biol 9(1):133. doi:10.1186/1471-2229-9-133
Gattolin S, Sorieul M, Frigerio L (2011) Mapping of tonoplast intrinsicproteins in maturing and germinating Arabidopsis seeds reveals duallocalization of embryonic TIPs to the tonoplast and plasma mem-brane. Mol Plant 4(1):180–189. doi:10.1093/mp/ssq051
Gomes D, Agasse A, Thiébaud P, Delrot S, Delrot S, Chaumont F (2009)Aquaporins are multifunctional water and solute transporters highlydivergent in living organisms. Biochim Biophys Acta 1788:1213–1228
Goodstein DM et al (2012) Phytozome: a comparative platform for greenplant genomics. Nucleic Acids Res 40(D1):D1178–D1186. doi:10.1093/nar/gkr944
Gustavsson S, Lebrun AS, Norden K, Chaumont F, Johanson U (2005) Anovel plant major intrinsic protein in Physcomitrella patens mostsimilar to bacterial glycerol channels. Plant Physiol 139:287–295
Higuchi T, Suga S, Tsuchiya T, Hisada H, Morishima S, Okada Y,Maeshima M (1998) Molecular cloning, water channel activityand tissue specific expression of two isoforms of radish vacuolaraquaporin. Plant Cell Physiol 39:905–913
Hruz T, Laule O, Szabo G, Wessendorp F, Bleuler S, Oertle L,Zimmermann P (2008) Genevestigator v3: a reference expressiondatabase for the meta-analysis of transcriptomes. Adv Bioinforma2008:420747. doi:10.1155/2008/420747
Jami SK, Clark GB, Ayele BT, Ashe P, Kirti PB (2012) Genome-widecomparative analysis of annexin superfamily in plants. PLoS ONE7:e47801
Johanson U, Karlsson M, Johansson I, Gustavsson S, Sjovall S, FraysseL,Weig AR, Kjellbom P (2001) The complete set of genes encoding
Funct Integr Genomics

major intrinsic proteins in Arabidopsis provides a framework for anew nomenclature for major intrinsic proteins in plants. PlantPhysiol 126:1358–1369
Johnson KD, Hofte H, Chrispeels MJ (1990) An intrinsic tonoplastprotein of protein storage vacuoles in seeds is structurally relatedto a bacterial solute transporter (GIpF). Plant Cell 2:525–532
Kaldenhoff R, Fischer M (2006) Functional aquaporin diversity in plants.Biochim Biophys Acta 1758(8):1134–1141. doi:10.1016/j.bbamem.2006.03.012
Kruse E, Uehlein N, Kaldenhoff R (2006) The aquaporins. Genome Biol7(2):206. doi:10.1186/gb-2006-7-2-206
Lamesch P et al (2012) The Arabidopsis Information Resource (TAIR):improved gene annotation and new tools. Nucleic Acids Res 40:D1202–D1210. doi:10.1093/nar/gkr1090
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA,McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R,Thompson JD, Gibson TJ, Higgins DG (2007) Clustal W andClustal X version 2.0. Bioinformatics 23:2947–2948
Ludevid D, Hofte H, Himelblau E, Chrispeels MJ (1992) The expressionpattern of the tonoplast intrinsic protein gamma-TIP in Arabidopsisthaliana is correlated with cell enlargement. Plant Physiol 100:1633–1639
Maeshima M (2001) Tonoplast transporters: organization and function.Annu Rev Plant Physiol Plant Mol Biol 52:469–497. doi:10.1146/annurev.arplant.52.1.469
Maurel C, Tacnet F, Guclu J, Guern J, Ripoche P (1997) Purified vesiclesof tobacco cell vacuolar and plasmamembranes exhibit dramatically
different water permeability and water channel activity. Proc NatlAcad Sci U S A 94:7103–7108
Maurel C, Verdoucq L, Luu DT, Santoni V (2008) Plant aquaporins:membrane channels with multiple integrated functions. Annu RevPlant Biol 59:595–624
Pao GM,Wu LF, Johnson KD, Hofte H, Chrispeels MJ, Sweet G, SandalNN, Saier MH Jr (1991) Evolution of the MIP family of integralmembrane transport proteins. Mol Microbiol 5:33–37
Proost S, Van Bel M, Sterck L, Billiau K, Van Parys T, Van de Peer Y,Vandepoele K (2009) PLAZA: a comparative genomics resource tostudy gene and genome evolution in plants. Plant Cell 21:3718–3731
Quigley F, Rosenberg JM, Shachar-Hill Y, Bohnert HJ (2002) Fromgenome to function: the Arabidopsis aquaporins. Genome Biol3(1):RESEARCH0001
Rambaut A (2012) FigTree v1.4. Institute of Evolutionary Biology,University of Edinburgh, Edinburgh
Roy SW, Penny D (2007) Patterns of intron loss and gains in plants:intron loss-dominated evolution and genome-wide comparison of O.sativa and A. thaliana. Mol Biol Evol 24(1):171–181
Schultz J et al (1998) SMART, a simple modular architecture researchtool: identification of signaling domains. Proc Natl Acad Sci U S A95:5857–5864
Silvestro D, Michalak I (2011) raxmlGUI: a graphical front-end forRAxML. Org Divers Evol 12(4):335–337. doi:10.1007/s13127-011-0056-0
Ye CY, Yang X, Xia X, Yin W (2013) Comparative analysis of cation/proton antiporter superfamily in plants. Gene 521:245–251
Funct Integr Genomics





























