Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1986 by The American SDciety of Biological Chemists, Inc.
Vol. 261, No. 27, Issue of September 25, PP. 12850-12855,1986 Printed in U.S.A.
Characterization of the Subunit Structure of the Maize Tonoplast ATPase IMMUNOLOGICAL AND INHIBITOR BINDING STUDIES*
(Received for publication, June 9, 1986)
Suzanne Mandala$ and Lincoln Taizg From the Biology Department, Thimann Laboratories, University of California at Santa Cruz, Santa Cruz, California 95064
Gradient purified preparations of the maize 400-kDa tonoplast ATPase are enriched in two major polypep- tides, 72 and 62 kDa. Polyclonal antibodies were pre- pared against these two putative subunits after elution from sodium dodecyl sulfate-polyacrylamide gel elec- trophoresis gel slices and against the solubilized native enzyme. Antibodies to both the 72- and 62-kDa poly- peptides cross-reacted with similar bands on immuno- blots of a tonoplast-enriched fraction from barley, while only the 72-kDa antibodies cross-reacted with tonoplast and tonoplast ATPase preparations from Neurospora. Antibodies to the 72-kDa polypeptide and the native enzyme both strongly inhibited enzyme ac- tivity, but the 62-kDa antibody was without effect.
The identity and function of the subunits was further probed using radiolabeled covalent inhibitors of the tonoplast ATPase, 7-chloro-4-nitro[’4C]benzo-2-oxa- 1,3-diazole ([‘4C]NBD-Cl) and N,N’-[’4C]dicyclohex- ylcarbodiimide ([14C]DCCD). [14C]NBD-C1 preferen- tially labeled the 72-kDa polypeptide, and labeling was prevented by ATP. [14C]DCCD, an inhibitor of the proton channel portion of the mitochondrial ATPase, bound to a 16-kDa polypeptide. Venturicidin blocked binding to the mitochondrial 8-kDa polypeptide but did not affect binding to the tonoplast 16-kDa polypep- tide. Taken together, the results implicate the 72-kDa polypeptide as the catalytic subunit of the tonoplast ATPase. The DCCD-binding 16-kDa polypeptide may comprise the proton channel. The presence of nucleo- tide-binding sites on the 62-kDa polypeptide suggests that it may function as a regulatory subunit.
Plant vacuoles are acidic organelles in which ions, sugars, organic acids, and hydrolytic enzymes are stored (1). Studies with isolated vacuoles and tonoplast (plant vacuolar mem- brane) vesicles have indicated that a proton-translocating ATPase present on the tonoplast generates an electrochemical gradient, which may be responsible for the observed accu- mulation of ions and solutes (reviewed in Ref. 2 ) . A similar ATP-dependent proton pump is also present on the Golgi of maize coleoptiles (3). Several recent reports have described the partial purification of a novel ATPase from plant and
~~
* This research was supported by Grant PCM-8301995 from the National Science Foundation and Grant DE-FG03-84ER13245 from the Department of Energy. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Present address: Dept. of Human Genetics and Physiology, Yale University School of Medicine, New Haven, CT 06510.
To whom correspondence should be addressed.
fungal vacuolar membranes (4-12). The partially purified ATPases retain the same characteristics as the tonoplast proton pumps but appear to differ significantly from the other two types of H+-ATPases known to occur in these cells, the FoF1 type found in mitochondria and chloroplasts and the ElE2 type located on plasma membranes. The plant and fungal vacuolar membrane ATPases are not inhibited by the plasma membrane ATPase inhibitor, vanadate, or the mitochondrial ATPase inhibitor, azide, but are sensitive to KN03 and KSCN (4-12). The subunit composition of the vacuolar membrane ATPases also appears to be novel. Partial purifications have indicated enrichment in two major polypeptides, one of M , 66,000-89,000 and the second of Mr 54,000-64,000 (4, 5, 7, 8, IO, 12,13). In beet root, Neurospora, and yeast, a potypeptide of M, 15,000-19,500 has been shown to bind to [14C]-DCCD1 (5-7). The presence of multiple subunits resembles the FoF,- type ATPases rather than E1E2-type ATPases. However, suf- ficient differences are observed in the subunit composition and inhibitor effects to suggest that the tonoplast proton pumps may represent a third type of ATPase.
In animal cells, ATP-dependent proton-pumping activity has been measured in many endomembrane vesicles, including lysosomes (13), chromaffin granules (14), Golgi (15), coated vesicles (16,17), synaptosomes (18), and multivesicular bodies (19). The characteristics of proton-pumping activity in the animal hydrolytic or storage organelles are similar to those of plant and fungal vacuoles, suggesting possible homology. An early report from Cidon and Nelson (14) postulated a novel subunit composition for the chromaffin granule ATPase. They reported enrichment of polypeptides of M, 125,000, 80,000, 40,000, and 20,000. More recently, Percy et al. (20) reported the presence of 5 polypeptides in preparations of chromaffin granule ATPase I of apparent M , 70,000, 57,000, 41,000, 33,000, and 16,000. The smallest polypeptide bound radioactive DCCD. Thus, there appears to be some similarity among plant, fungal, and animal endomembrane ATPases.
Previously we have shown that an antibody produced to the M , 72,000 protein from maize cross-reacts to a similar poly- peptide in Neurospora vacuolar membranes (5). In the present study, we have examined the effects of antibodies made to the M, 72,000 and 62,000 proteins on ATPase activity and have analyzed their cross-reactivity to barley tonoplast mem- branes. In addition, radioactively labeled inhibitors of the tonoplast ATPase were used as probes to characterize putative subunits on SDS gels.
MATERIALS AND METHODS
Enzyme Purification-Solubilization and purification of the tono- plast ATPase was conducted as previously described (4). Briefly,
The abbrevations used are: DCCD, N,N,’-dicyclohexylcarbodii- mide; NBD-C1,7-chloro-4-nitrobenzo-2-oxa-1,3-diazole; SDS, sodium dodecyl sulfate; Mes, 4-morpholineethanesulfonic acid.
12850

2 Immunological and Binding Studies on Maize Tonoplast ATPase 12851
coleoptiles were collected from 5-day-old maize seedlings (Zea mays L. cv Golden Cross Bantam), and tonoplast-enriched membranes were isolated from 1/8% dextran step gradients and stored in resus- pension buffer (0.25 M sorbitol, 1 mM EDTA, 1 mM dithiothreitol, 10 mM Tris-Mes, pH 7.5) a t -70 "C. We have previously shown that the H+-translocating ATPase enriched in this preparation is indistin- guishable from the H+-translocating ATPase associated with isolated vacuoles purified from corn coleoptile protoplasts (21). Treatment of the membranes with 0.15% deoxycholate removed loosely bound protein and increased the specific activity of the tonoplast ATPase 5-fold to approximately 1 pmol/min/mg of protein. The ATPase was solubilized with 40 mM octyl-P-D-glucopyranoside and partially pu- rified by centrifugation into a 4-ml10-25% linear sucrose gradient in 1 mM EDTA, 1 mM dithiothreitol, 20 mM octyl-P-D-glucopyranoside, and 10 mM Tris-Mes, pH 7.5. The gradient was centrifuged in a Beckman SW 65 rotor a t 200,000 X g for 5 h a t 4 "C, and 0.5-ml fractions were collected. The peak fractions of ATPase were enriched in the M, 72,000 and M, 62,000 bands on SDS-polyacrylamide gel electrophoresis gels, as previously reported (4).
ATPase Assay-Five to 20 pl of enzyme were assayed in a 0.5-ml volume containing 2.5 mM ATP, 2.5 mM MgS04, 50 mM KCl, 25 mM Tris-Mes, pH 6.5. After a 20-30-min incubation a t 37 "C, the reaction was terminated, and released Pi was determined by the method of Fiske and Subbarow (22).
Inhibitor Binding-To label tonoplast proteins with [l4C]NBD-C1, 50 pl of deoxycholate-washed membranes (100 pg of protein) were treated with 0.5 pCi of [l4C]NBD-C1 (109 mCi/mmol) to give a final concentration of 75 pM. Following a 1-h incubation on ice in the presence or absence of 5 mM MgATP, the membranes were diluted with 4 ml of 50 mM Tris/HCl, pH 8.0, and centrifuged at 120,000 X g for 1 h in a Beckman SW 60 rotor. The pellet was resuspended in 40 pl of dilution buffer and prepared for SDS-gel electrophoresis. Labeling of partially purified ATPase from the sucrose gradient was performed similarly except that 60 pl of ATPase (7 pg of protein, specific activity, 2.5 pmol/min/mg) were incubated with 0.3 pCi of ["CINBD-Cl to give a final concentration of 35 p ~ . After the 1 h of incubation, samples were prepared for gel electrophoresis without washing.
The method of Sussman and Slayman (23) was used to label maize proteins with [14C]DCCD. Mitochondrial membranes (150 pg of pro- tein collected from the 35/50% interface of a sucrose step gradient) and deoxycholate-washed tonoplast membranes (80 pg) were pre- treated in the presence or absence of 50 pg/ml venturicidin for 10 min on ice. A 90-min incubation with 0.375 pCi of[14C]DCCD at a concentration of 12 p~ was terminated by dilution into 4 ml of resuspension buffer and centrifugation a t 120,000 X g for 45 min. Pellets were resuspended in 40 p1 of buffer and prepared for gel electrophoresis. Gradient-purified ATPase was labeled with 0.25 pCi of [14C]DCCD a t a final concentration of 67 p~ and was prepared directly for electrophoresis without washing.
Antibody Production-The M, 62,000 and 72,000 antigens were prepared by cutting out bands from 8% acrylamide gels that had been loaded with 900 pg of deoxycholate-washed tonoplast membranes. Coomassie-stained gel slices (40 pg of estimated protein) were mixed with 1 ml of 0.15 M NaCl and thoroughly crushed with a glass homogenizer. Antigen was vortexed with 1 ml of complete Freund's adjuvant and 5 mg of tubercle bacilli. Twenty intradermal injections were given by the method of Vaitukaitis et al. (24) on the shaved backs of New Zealand White rabbits. Subsequent injections were given subcutaneously a t 1-3-month intervals for a total of 6 injections. Antigen for boosting was prepared by homogenizing gel slices with incomplete Freund's adjuvant.
Antisera to the native ATPase was prepared by subcutaneously injecting rabbits with 1 ml (-100 pg) of partially purified ATPase from the sucrose gradient mixed with 1 ml of complete Freund's adjuvant. Two boosts using the same amount of ATPase with incom- plete Freund's adjuvant were given at 1-month intervals. Approxi- mately 40 ml of blood were collected from the ear 10 days after the last injections. Antiserum was precipitated with 40% ammonium sulfate and resuspended in 50 mM Tris/HCl, pH 7.4, 150 mM NaC1. Specific cross-reaction to the M, 72,000 protein could be detected by the immunoblot procedure after 2 injections, and after 3 injections for the antibodies to the M. 62,000 protein.
SDS-Polyacrylamide Gel Electrophoresis-Slab gels were 1.5-mm thick and consisted of 10% acrylamide, 0.27% bisacrylamide, 25% urea, and 0.37 M Tris-HC1, pH 8.8. Polymerization was catalyzed by the addition of 0.02% N,N,N',N'-tetramethylethylenediamine and 0.72 mg/ml ammonium persulfate. A stacking gel was composed of
3% acrylamide, 0.08% bisacrylamide, 0.12 M Tris-HC1, pH 6.8. The running buffer was 0.2 M glycine, 0.25 M Tris, and 0.1% SDS. Proteins were mixed with sample preparation buffer to final concentrations of 0.1 mM phenylmethylsulfonyl fluoride, 1.7% SDS, 1% 8-mercaptoeth- anol, 8% glycerol, 42 mM Tris-HC1, pH 6.8, and incubated for 20 min at 70 "C. The gels were stained and destained by the method of Fairbanks et al. (25).
Fluorography-Gels of 14C-labeled material were incubated in 80 ml of ENHANCE (New England Nuclear) for 1 h and then rinsed with cold water for 1 h before drying. Intensifying screens (Du Pont) were used with 32P- and '251-treated material. Radioactive bands were detected with Kodak x-ray (XAR-5) film after 1-24-day storage in the dark at -70 "C.
Electrotransfer-Proteins from SDS gels were electrotransferred to nitrocellulose, and the blots were washed, incubated with antibody, and treated with 9-Pro te in A, as described by Rott and Nelson (26). For the overnight incubation with antisera, a concentration of 20 pl in 40 ml of buffer was used for the antisera to the M, 62,000 and 72,000 proteins and 40 pl of antisera to gradient-purified ATPase.
Protein Assay-Protein was determined by the method of Lowry et al. (27) after trichloroacetic acid precipitation.
Neurospora and Barley Membranes-The Neurospora vacuolar membranes and ATPase fractions, prepared as described in Ref. 5, were provided by Dr. E. J. Bowman, University of California, Santa Cruz. The barley membranes were a gift from Dr. F. DuPont (United States Department of Agriculture, Albany, CA) and were obtained from a 22/30% interface of a sucrose step gradient using a modifica- tion of the procedure described in DuPont and Hurkman (28).
Reagent~-['~ClDCCD was from Amersham Corp., and [14C]NBD- C1 was from Research Products International Corp. Adjuvant and tubercle bacilli (Mycobacterium tuberculosis H37 RA) were obtained from Difco. Venturicidin came from BDH Chemicals, Ltd., Great Britain. All other chemicals were obtained as previously described (4).
RESULTS
Immunological Characterization-Antibodies to the M, 62,000 and 72,000 polypeptides were produced by immunizing rabbits against bands cut out from SDS gels. Specific cross- reaction to the appropriate proteins was detected by immu- noblotting as shown in Fig. 1. Antibodies to the M, 62,000 polypeptide cross-reacted strongly to the M, 62,000 protein from deoxycholate-washed maize tonoplasts and the partially
A. Anti-62K B. Anti-72K c. Anti- ATPase
1 2 3 4 5 1 2 3 4 5 -_"- 1
R
92K-
66K*
45K.
31 K-
*62K 1- 72K -&
i I .
i
L1
FIG. 1. Immunoblot analysis of three antibodies cross-re- acted to plant and fungal proteins. Antibodies were produced against the M, = 62,000 subunit cut out from SDS gels (A), the M, 72,000 subunit cut out from SDS gels (B), and gradient-purified ATPase fractions (C). Lanes: I , gradient-purified maize tonoplast ATPase (-7 pg); 2, deoxycholate-washed maize tonoplasts (-20 pg); 3, Neurospora vascuolar membranes (-50 pg); 4, Neurospora gradient- purified vacuolar ATPase (-5 pg); 5, barley tonoplast-enriched mem- branes (-50 pg) Molecular mass standards are given on the left.
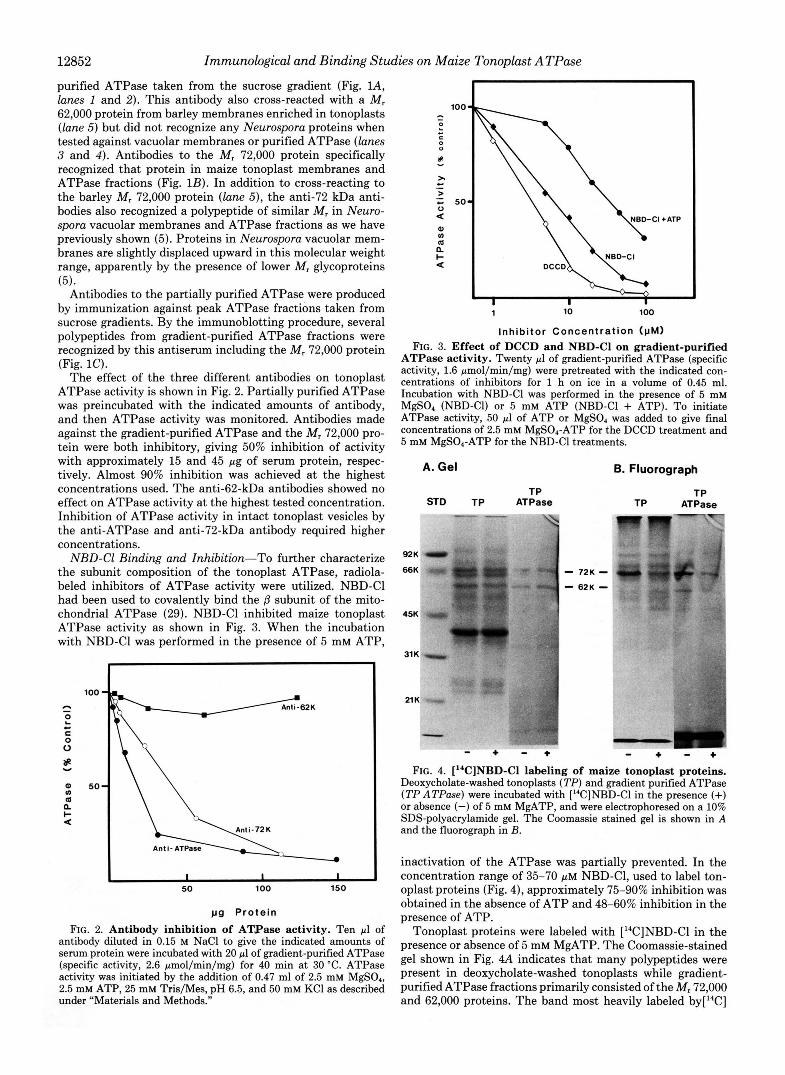
12852 Immunological and Binding Studies on Maize Tonoplast ATPase
purified ATPase taken from the sucrose gradient (Fig. lA, lanes 1 and 2). This antibody also cross-reacted with a M, 62,000 protein from barley membranes enriched in tonoplasts (lane 5 ) but did not recognize any Neurospora proteins when tested against vacuolar membranes or purified ATPase (lanes 3 and 4) . Antibodies to the M, 72,000 protein specifically recognized that protein in maize tonoplast membranes and ATPase fractions (Fig. 1B). In addition to cross-reacting to the barley MI 72,000 protein (lane 5 ) , the anti-72 kDa anti- bodies also recognized a polypeptide of similar M, in Neuro- spora vacuolar membranes and ATPase fractions as we have previously shown (5). Proteins in Neurospora vacuolar mem- branes are slightly displaced upward in this molecular weight range, apparently by the presence of lower MI glycoproteins (5).
Antibodies to the partially purified ATPase were produced by immunization against peak ATPase fractions taken from sucrose gradients. By the immunoblotting procedure, several polypeptides from gradient-purified ATPase fractions were recognized by this antiserum including the M, 72,000 protein (Fig. IC).
The effect of the three different antibodies on tonoplast ATPase activity is shown in Fig. 2. Partially purified ATPase was preincubated with the indicated amounts of antibody, and then ATPase activity was monitored. Antibodies made against the gradient-purified ATPase and the M, 72,000 pro- tein were both inhibitory, giving 50% inhibition of activity with approximately 15 and 45 pg of serum protein, respec- tively. Almost 90% inhibition was achieved at the highest concentrations used. The anti-62-kDa antibodies showed no effect on ATPase activity at the highest tested concentration. Inhibition of ATPase activity in intact tonoplast vesicles by the anti-ATPase and anti-72-kDa antibody required higher concentrations.
NBD-C1 Binding and Inhibition-To further characterize the subunit composition of the tonoplast ATPase, radiola- beled inhibitors of ATPase activity were utilized. NBD-C1 had been used to covalently bind the /3 subunit of the mito- chondrial ATPase (29). NBD-C1 inhibited maize tonoplast ATPase activity as shown in Fig. 3. When the incubation with NBD-Cl was performed in the presence of 5 mM ATP,
I I I 50 100 150
pg Protein FIG. 2. Antibody inhibition of ATPase activity. Ten pl of
antibody diluted in 0.15 M NaCl to give the indicated amounts of serum protein were incubated with 20 pl of gradient-purified ATPase (specific activity, 2.6 pmol/min/mg) for 40 min at 30 "C. ATPase activity was initiated by the addition of 0.47 ml of 2.5 mM MgSO,, 2.5 mM ATP, 25 mM Tris/Mes, pH 6.5, and 50 mM KC1 as described under "Materials and Methods."
1 10 100
Inhibitor Concentration (pM) FIG. 3. Effect of DCCD and NBD-Cl on gradient-purified
ATPase activity. Twenty pl of gradient-purified ATPase (specific activity, 1.6 pmol/min/mg) were pretreated with the indicated con- centrations of inhibitors for 1 h on ice in a volume of 0.45 ml. Incubation with NBD-Cl was performed in the presence of 5 mM MgSO, (NBD-C1) or 5 mM ATP (NBD-Cl + ATP). To initiate ATPase activity, 50 pl of ATP or MgSO, was added to give final concentrations of 2.5 mM MgS04-ATP for the DCCD treatment and 5 mM MgS04-ATP for the NBD-Cl treatments.
A. Gel B. Fluorograph
TP STD TP ATPase TP
TP ATPase
'W > Wm-Y 92K
6 6 K
45K
31 K
21 n
- + - + FIG. 4. I"C1NBD-Cl labelin
I n
- - + - - +
aize tonODlast Droteins. f m Deoxycholate-washed tonoplasts (TP) and gradient purifie'd ATPase (TPATPase) were incubated with ['4C]NBD-Cl in the presence (+) or absence (-) of 5 mM MgATP, and were electrophoresed on a 10% SDS-polyacrylamide gel. The Coomassie stained gel is shown in A and the fluorograph in B.
kg 0:
inactivation of the ATPase was partially prevented. In the concentration range of 35-70 p~ NBD-C1, used to label ton- oplast proteins (Fig. 4), approximately 75-90% inhibition was obtained in the absence of ATP and 48-60% inhibition in the presence of ATP.
Tonoplast proteins were labeled with ['4C]NBD-Cl in the presence or absence of 5 mM MgATP. The Coomassie-stained gel shown in Fig. 44 indicates that many polypeptides were present in deoxycholate-washed tonoplasts while gradient- purified ATPase fractions primarily consisted of the M, 72,000 and 62,000 proteins. The band most heavily labeled by[14C]

Immunological and Binding Studies on Maize Tonoplast ATPase 12853
NBD-C1 in the fluorograph (Fig. 4B) was the M , 72,000 polypeptide. The M, 72,000 protein was also the only one to show marked protection from labeling by MgATP.
DCCD Binding and Inhibition-The presence of a separate polypeptide ( M , - 8,000) that is involved in proton conduct- ance distinguishes FoF1- from E,E,-type ATPases. [14C]DCCD has been used to identify the proton channel of both types of ATPases (23, 30), since DCCD is thought to inhibit H'- ATPases by blocking proton conductance (30). Tonoplast ATPase activity was also inhibited by DCCD. Fifty percent inactivation of ATPase was measured at a concentration less than 10 ~ L M DCCD (Fig. 3). [14C]DCCD was used to compare labeling of maize mitochondrial and tonoplast proteins. Fig. 5 shows that [14C]DCCD-labeled a polypeptide in mitochon- drial membranes at a M, of approximately 8,000, expected for the Fo proteolipid. Binding to this subunit was prevented by venturicidin, as has been previously demonstrated (11). An- other protein of M, - 25,000 was also labeled in mitochondrial membranes. This may be explained by the existence of several other mitochondrial proteins, such as subunits of cytochrome oxidase and cytochrome bcl, that are sensitive to DCCD (30). In tonoplast membranes,[14C]DCCD also labeled a low molec- ular weight polypeptide, which differed from the mitochon- drial proteolipid by having twice the relative mobility on the gel (Mr 16,000) and by not showing sensitivity to venturicidin. The same polypeptide was also identified as the major ["C] DCCD binding protein in purified tonoplast ATPase fractions from the sucrose gradient. The low level of binding to the M, 72,000 and 62,000 proteins in the gradient fraction may be nonspecific binding due to the method of directly loading the sample on the gel or may represent specific binding as has been reported for the @ subunit of the mitochondrial ATPase (31). When mitochondrial and tonoplast membranes were treated the same way (i.e. without dilution and pelleting) these subunits also showed preferential labeling.
DISCUSSION
Studies on partially purified vacuolar membrane ATPases from plants and fungi have provided evidence for an enzyme that differs kinetically and in subunit composition from FoF1- type and EIEz-type ATPases (4-12). Although the complete
A. Gel
921
881
451
31 I
21 I
141 - 1 6 K -
- 8 K -
- + - + - - + - + -
subunit composition of the vacuolar ATPase is not known, purification from several species reveals substantial agree- ment on the major polypeptides (Table I). In all species examined so far, two major polypeptides, M, 66,000-89,000 and MI 54,000-64,000, have been detected as well as a smaller DCCD-binding polypeptide of M, 13,000-19,500. In Heuea (lo), and Saccharomyces carlsbergensis (8) other bands also co-purify with ATPase activity, as indicated in Table I. The apparent subunit composition of the animal chromaffin gran- ule ATPase I bears a striking resemblance to that of the vacuolar membrane ATPases. These data are consistent with a growing consensus that the vacuolar membrane ATPase represents a new class of proton-pumping ATPases associated with a variety of single membrane-bound acidic organelles in eukaryotes (e.g. Refs. 5,6, 11,32, and 33). The functional size of the tonoplast ATPase has been measured by radiation inactivation and shown to be large, approximately 400 kDa for the maize enzyme (4) and 520 kDa for the ATPase of Neurospora (5). A dichloromethane-solubilized vacuolar ATPase of Heuea has a molecular mass of about 200 kDa, determined by gel filtration (10).
The presence of multiple subunits, including a low M, polypeptide that binds [14C]DCCD, is reminiscent of FoFI- type ATPases. The soluble F, portion is composed of 5 or more subunits with molecular weights ranging between 7,500 and 62,000 Da (34). The catalytic polypeptide is thought to be the p subunit of M, - 55,000. The membrane-bound Fo portion contains the M , 8,000 proton channel that binds ["C] DCCD and 2 other subunits. In contrast, the proton-translo- cating EIEz ATPase found on plant and fungal plasma mem- branes consists of a single type of polypeptide of MI 100,000. This subunit functions catalytically and also binds ["C] DCCD (23). With respect to sensitivity to inhibitors, the tonoplast ATPase appears to more closely resemble the FoF1- type than the EIEz-type enzyme (11, 32). The tonoplast and FoF1-type ATPases are not inhibited by vanadate, suggesting that they do not form phosphorylated intermediates charac- teristic of the reaction cycle of E1E2-type enzymes (4, 7, 11, 32). In comparisons of the effectiveness of a large variety of inhibitors on all three types of ATPases in Neurospora (11) and between the mitochondrial and tonoplast ATPases in oat roots (32), it was demonstrated that many inhibitors had strikingly similar effects on the mitochondrial and tonoplast enzymes. However, the tonoplast ATPase is resistant to the mitochondrial inhibitor, azide (4, 7,9, 32).
This study was undertaken in an effort to further charac- terize the subunits and to identify the catalytic site of the maize tonoplast ATPase using immunological and inhibitor binding methods. Previously we have shown that the tono- plast ATPase represents a distinct immunological class based on cross-reactivity studies (5). Antibodies to the Neurospora plasma membrane ATPase and the p subunit of the E. coli FoFl ATPase do not cross-react with the vacuolar membrane ATPase of Neurospora, and the antibody to the maize M, 72,000 tonoplast polypeptide does not recognize mitochon- drial or plasma membrane polypeptides (5). Uchida et al. (7) have shown that antiserum to the F1 ATPase of S. cerevisiae inhibits mitochondrial but not the vacuolar membrane ATP- ase. In the present study, we have shown that antibodies to the maize MI 72,000 and MI 62,000 polypeptides both cross- reacted with barley, while only the antibody to the maize M, 72,000 subunit cross-reacted with Neurospora (Fig. 1). This indicates that the larger subunit is more highly conserved. In a study on the immunological properties of FoFl ATPases, Rott and Nelson (26) demonstrated that only the catalytic
FIG. 5. ['4C]DCCD labeling of maize proteins. Mitochondrial membranes (Mito), deoxycholate-washed tonoplasts (TP), and gra- dient-purified tonODlast ATPase (TP ATPase) were incubated with [14C]DCCD in the presence (+) or absence (-) of 5 pg/ml venturicidin. Following electrophoresis on an 8% acrylamide gel (A) , radioactive bands were detected by fluorography (B) . subunit (p) of mitochondrial, chloroplast, and E. coli ATPases

12854 Immunological and Binding Studies on Maize Tonoplast ATPase TABLE I
Subunit composition of vacuolar membrane H'-ATPases
Source M , range
66,000-89,000 54.000-64,000 23,000-45,000 13,000-19,500 Ref.
Plant Zeu mays 72,000" 62,000 16,000' 4 Hevea brasiliensis 66,000 54,000 23,000 13,000 10 Beta vulgaris 67,000 57,000 16,000b 6 Avena sativa 74,000 64,000 16,000* 12
Neurospora crassa 70,000" 62,000 15,000* 5, 11 Saccharomyces cerevisiae 89,000 64,000 19,500' 7 Saccharomyces carlsbergensis 75,000' 62,000 16,000, 14,000, 12,000,9,000' 8
Chromaffin granule 70,000 57,000 41,000, 33,000 16,000' 20
Fungal
Animal
Indicates the polypeptide NBD-CI and that binding is substrate protectable. - " . - ' Indicates the polypeptide binds DCCD. e Indicates the polypeptide binds diethylstilbestrol.
consistently cross-reacted to antibodies made to the 0 subunit from a different species. Antibodies made to a or y subunits cross-reacted weakly or not at all to subunits from other species.
The importance of the M , 72,000 protein for the ATPase was also evident from measuring antibody effects on activity (Fig. 2). ATPase activity was inhibited only by the two anti- bodies that recognized the M , 72,000 protein in immunoblots: the antiserum made against gradient-purified ATPase frac- tions and the 72-kDa polypeptide. Thus, the immunological data support the M , 72,000 polypeptide as the catalytic sub- unit.
Studies with radioactively labeled inhibitors are consistent with this conclusion. NBD-Cl was chosen as a potentially useful label of the catalytic site since Ferguson et al. (29) have found that NBD-Cl reacts exclusively with the p subunit of the mitochondrial ATPase. NBD-C1 inhibited the tonoplast ATPase in an ATP-protectable manner (Fig. 3). Fig. 4 shows that [14C]NBD-C1 primarily labeled the M , 72,000 polypeptide in maize tonoplast membranes. The M , 72,000 polypeptide was also the only band to show significant protection from labeling by ATP. Similar results have been obtained for the vacuolar ATPase of Neurospora (5 ) . Manolson et al. (6) re- ported that the radioactively labeled substrate analog ["PI-3- (0)-benzoy1)benzoyl ATP preferentially labeled the 57-kDa polypeptide of beet tonoplast ATPase, rather than the 67- kDa polypeptide. However, Manolson et al. (6) concluded that the binding site on the 57-kDa subunit is probably a regulatory site since BzATP was not a simple competitive inhibitor of the ATPase. In contrast, we have observed that [3'P]8-azido- ATP binds to both the 72- and 62-kDa polypeptides of maize tonoplast ATPase,' suggesting that both catalytic and regu- latory sites are being labeled. By analogy to the F, ATPase, which has as many as three nucleotide-binding sites on the a and @ subunits (34, 35), nucleotide-binding sites may be present on both of the major subunits of the tonoplast ATP- ase. Since other evidence strongly supports the 72-kDa poly- peptide as the catalytic subunit, we concur with Manolson et al. (6) that the 62-kDa polypeptide probably represents a regulatory subunit.
In conclusion, further characterization of the subunit com- position of the maize tonoplast ATPase has provided evidence for the oligomeric nature of the enzyme. Results from the immunoIogica1 and inhibitor binding studies have suggested that the M , 72,000 polypeptide is the catalytic subunit of the ATPase. The M , 62,000 protein may be a regulatory subunit
S. Mandala and L. Taiz, unpublished data.
with nucleotide-binding sites, while the M , 16,000 polypeptide that binds DCCD is most likely the proton channel of the ATPase. Similarities between FoF1-type ATPases and the vacuolar ATPases from plants and fungi are evident from this and other studies (11, 32). As previously noted ( 5 ) , the FoF, ATPase that most closely resembles the tonoplast ATPase is one from the anaerobic bacterium, Clostridium pasteurianum (36). This enzyme has a simpler subunit composition ( M , = 66,000, 58,000, 43,000, and 15,000) than that found in mito- chondria, chloroplasts, and aerobic bacteria (36). The possi- bility that this enzyme is a primitive form from which more complex F,F,-type ATPases has evolved has been proposed (33).
Acknowledgments-We thank Dr. E. J. Bowman and Dr. B. J. Bowman for stimulating and helpful discussions throughout this study and for providing the Neurospora vacuolar membranes and ATPase fractions. We also thank Dr. Francis DuPont of the United States Department of Agriculture in Albany, CA for preparing the barley membranes used in this study.
REFERENCES 1. Wagner, G. J. (1982) in Recent Advances in Phytochemistry
(Creasy, L., and Hrazdina, G., eds) pp. 1-45, Plenum Publishing Corp., New York
2. Sze, H. (1985) Annu. Rev. Plant Physiol. 36, 175-208 3. Chanson, A. & Taiz, L. (1985) Plant Physiol. 78, 232-240 4. Mandala, S. & Taiz, L. (1985) Plant Physwl. 78, 327-333 5. Bowman, E. J., Mandala, S., Taiz, L. & Bowman, B. J. (1986)
6. Manolson, M. F., Rea, P. A. & Poole, R. J. (1985) J. Biol. Chem.
7. Uchida, E., Ohsumi, Y. & Anraku, Y. (1985) J. Biol. Chem. 2 6 0 ,
8. Lichko, L. P. & Okorokov, L. A. (1985) FEBS Lett. 187 , 349-
9. Tognoli, L. (1985) Eur. J. Biochem. 146,581-588
Proc. Natl. Acad. Sei. U. S. A. 83, 48-52
260,12273-12279
1090-1095
353
10. Marin, B., Preisser, J. & Komor, E. (1985) Eur. J. Biochem. 151 ,
11. Bowman, E. J. (1983) J. Biol. Chem. 258, 15238-15244 12. Randall, S. K. & Sze, H. (1986) J. Bwl. Chem. 261,1364-1371 13. Okhuma, S., Moriyama, Y. & Takano, T. (1982) Proc. Natl. Acad.
14. Cidon, S. & Nelson, N. (1983) J. Biol. Chem. 258, 2892-2898 15. Glickman, J., Croen, K., Kelly, S. & Al-Awqati, Q . (1983) J. Cell
Biol. 97,1303-1308 16. Forgac, M., Cantley, L., Wiedernmann, B., Altstiel, L. & Branton,
D. (1983) Proc. Natl. Acad. Sci. U. S. A. 80,1300-1303 17. Xie, X.-S., Stone, D. K. & Racker, E. (1984) J. Biol. Chem. 259,
18. Russell, J. T. & Holz, R. W . (1981) J. Biol. Chem. 256 , 5950-
19. Van Dyke, R. W., Hornick, C. A., Belcher, J., Scharschmidt, B.
131-140
Sci. U. S. A. 79,2758-2762
11676-11678
5953

20.
21. 22. 23.
24.
25.
26. 27.
Immunological and Binding Studies on Maize Tonoplast ATPase 12855
F. & Havel, R. J. (1985) J. Biol. Chem. 260, 11021-11026
557-564 Percy, J. M., Pryde, J. G. & Apps, D. K. (1985) Biochern. J. 231,
Mandala, S. & Taiz, L. (1985) Plant Physiol. 78, 104-109 Fiske, C. H. & Subbarow, Y. (1925) J. Biol. Chem. 66,375-400 Sussman, M. R. & Slayman, C. W. (1983) J. Biol. Chern. 258,
Vaitukaitis, J., Robbin, J. B., Nieschlag, E. & Ross, G. T. (1971)
Fairbanks, G., Steck, T. L. & Wallach, D. F. H. (1971) Biochem-
Rott, R. & Nelson, N. (1981) J. Biol. Chem. 256, 9224-9228 Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J.
1839-1843
J. Clin. Endocrinol. Metab. 33,958-991
istry 10.2606-2617
(1951) J. Biol. Chem. 193, 265-275
28.
29.
30. 31.
32. 33. 34.
35.
36.
DuPont, F. M. & Hurkman, W. J. (1985) Plant Physiol. 77,857-
Ferguson, S. J., Lloyd, W. J. & Radda, G. K. (1975) Eur. J.
Solioz, M. (1984) Trends Biochem. Sci. 9, 309-312 Khananshvili, D. & Gromet-Elhanan, 2. (1983) J. Biol. Chern.
Wang, Y. & Sze, H. (1985) J. Biol. Chem. 260,10434-10443 Maloney, P. C . & Wilson, T. H. (1985) Bioscience 35, 43-48 Khananshvili, D. & Gromet-Elhanan, Z. (1985) Proc. Natl. Acad.
Amzel, L. M. & Pedersen, P. L. (1983) Annu. Reu. Biochern. 52,
Clarke, D. J., Fuller, F. M. & Morris, J. G. (1979) Eur. J. Biochem.
862
Biochern. 54, 127-133
258,3720-3725
Sei. U. S. A. 82,1886-1890
801-824
98,597-612





























