Embed Size (px)
Citation preview

Functional Type 1 Secretion System Involved in Legionellapneumophila Virulence
Fabien Fuche,a,b,c,d,e Anne Vianney,a,b,c,d,e Claire Andrea,a,b,c,d,e Patricia Doublet,a,b,c,d,e Christophe Gilberta,b,c,d,e
CIRI, International Center for Infectiology Research, Legionella Pathogenesis Group, Université de Lyon, Lyon, Francea; Inserm, U1111, Lyon, Franceb; Ecole NormaleSupérieure de Lyon, Lyon, Francec; Université Lyon 1, Centre International de Recherche en Infectiologie, Lyon, Franced; CNRS, UMR5308, Lyon, Francee
Legionella pneumophila is a Gram-negative pathogen found mainly in water, either in a free-living form or within infected pro-tozoans, where it replicates. This bacterium can also infect humans by inhalation of contaminated aerosols, causing a severeform of pneumonia called legionellosis or Legionnaires’ disease. The involvement of type II and IV secretion systems in the viru-lence of L. pneumophila is now well documented. Despite bioinformatic studies showing that a type I secretion system (T1SS)could be present in this pathogen, the functionality of this system based on the LssB, LssD, and TolC proteins has never beenestablished. Here, we report the demonstration of the functionality of the T1SS, as well as its role in the infectious cycle of L.pneumophila. Using deletion mutants and fusion proteins, we demonstrated that the repeats-in-toxin protein RtxA is secretedthrough an LssB-LssD-TolC-dependent mechanism. Moreover, fluorescence monitoring and confocal microscopy showed thatthis T1SS is required for entry into the host cell, although it seems dispensable to the intracellular cycle. Together, these resultsunderline the active participation of L. pneumophila, via its T1SS, in its internalization into host cells.
Legionella pneumophila is a Gram-negative pathogen that colo-nizes aquatic environments, where it survives by infecting wa-
ter protozoans, especially amoebas. Pathogenic strains of L. pneu-mophila emerge from the environment after replication in suchprotozoans, are disseminated through aerosols of contaminatedwater, and reach human alveolar macrophages, where they begin anew infectious cycle, causing a severe form of pneumonia calledLegionnaires’ disease or legionellosis. The development of air-conditioning systems, cooling towers, and other aerosol-generat-ing systems is frequently reported as the reason for the spread ofthis pathogen (1). Indeed, L. pneumophila is the second etiologicalagent of pneumonia requiring hospitalization in intensive careunits, after Streptococcus pneumoniae (2). Moreover, the mortalityrate of Legionnaires’ disease, even with appropriate antibiotictreatment, ranges from 7 to 25%, thus making legionellosis a pub-lic health concern (3). Moreover, L. pneumophila is a scientificallyrelevant model to study intracellular pathogens (4).
Among pathogenic bacteria, secretion systems play a crucialrole in virulence, whether by damaging the host or by being essen-tial for bacterial replication, for example, by hijacking cellularpathways or promoting escape from the immune system (5). Bac-terial protein secretion systems have been extensively studied, andto date, seven of them have been identified within two categories:those that address their substrates to the extracellular environ-ment, such as type I secretion systems (T1SSs), T2SSs, T5SSs, andT7SSs, and those whose substrates are injected through the hostmembrane into its cytoplasm, such as T3SSs, T4SSs, and T5SSs(6). In L. pneumophila, a T2SS and a T4SS were identified severalyears ago (7–9) and their implication in the virulence of this bac-terium has been extensively studied (10, 11). T4SS Dot/Icm hasbeen particularly well investigated, as it is responsible for thetranslocation of more than 275 effector proteins into the cyto-plasm of infected cells (12, 13). Those effectors are required for theentire intracellular cycle of L. pneumophila, as they are involved inthe creation of a replicative niche inside the host cell, called aLegionella-containing vacuole (LCV), that is suitable for bacterialreplication. A decade ago, bioinformatic studies predicted a set of
genes that code for a putative T1SS in Legionella, but the function-ality of such a system has never been demonstrated (14). Theimplication of T1SSs in the virulence of pathogenic bacteria, suchas uropathogenic Escherichia coli (UPEC), Bordetella pertussis, oreven the entomopathogen Photorhabdus, was demonstrated manyyears ago (15–17). To date, the most extensively studied examplesof T1SSs are the Hly system in UPEC (18), the Apr system inPseudomonas aeruginosa (19), and the Prt system in Dickeya da-dantii (20). For example, in the case of UPEC, the system respon-sible for the secretion of the hemolytic toxin HlyA is based onHlyB, an inner membrane ABC transporter involved in substraterecognition and translocation, the periplasmic membrane fusionprotein HlyD, and the outer membrane-spanning porin TolC(21). Together, these three components form a channel that spansboth membranes to export substrates into the extracellular envi-ronment. The most commonly found substrates are toxins such aslipases and proteases, adhesins, and hemophores (22). Of theT1SS-secreted toxins, the repeats-in-toxin (RTX) family is themost extensively studied. RTX proteins are large, multidomainproteins that have diverse functions, depending on the embeddedfunctional domains. HlyA exhibits a pore-forming activity, CyaAof Bordetella pertussis has an adenylate cyclase activity, and LapA
Received 30 July 2014 Accepted 14 November 2014
Accepted manuscript posted online 24 November 2014
Citation Fuche F, Vianney A, Andrea C, Doublet P, Gilbert C. 2015. Functional type1 secretion system involved in Legionella pneumophila virulence. J Bacteriol 197:563–571. doi:10.1128/JB.02164-14.
Editor: J. S. Parkinson
Address correspondence to Christophe Gilbert,[email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.02164-14.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JB.02164-14
February 2015 Volume 197 Number 3 jb.asm.org 563Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

and LapF of Pseudomonas putida are adhesins involved in cellsurface or cell-cell interactions, respectively (23). They also con-tain several repeats of the glycine-rich nonapeptide motifGGXGXDXXX, which is the signature of RTX proteins (24). TheRtxA protein of L. pneumophila also contains a high number oftandem repeats, whose exact number and nature vary amongstrains (25). In this pathogen, RtxA has been linked to the abilitiesof the bacterium to invade and replicate within amoebas and mac-rophages (26, 27). However, the mechanism of the potential se-cretion of this protein has never been investigated.
To date, there is no available study of the functionality of theT1SS in L. pneumophila or its role in virulence. In this paper, wereport that LssD-LssD-TolC is a functional T1SS and that RtxA isa substrate of this T1SS. Moreover, we show that it is involved invirulence for amoeba and macrophages, precisely in early steps ofinfection, such as entry into the host cell.
MATERIALS AND METHODSStrains and growth conditions. All of the strains used in these study arelisted in Table S1 in the supplemental material. L. pneumophila strainsLens and Paris were grown on buffered charcoal yeast extract (BCYE) agaror in liquid BYE medium. Unless otherwise mentioned, all cultures weregrown at 30°C since we observed in our lab that virulence for amoebas wasoptimal at this temperature. Kanamycin (10 �g ml�1), chloramphenicol(5 �g ml�1), or gentamicin (5 �g ml�1) was added when appropriate. E.coli strains were grown at 37°C in LB medium supplemented with chlor-amphenicol (5 �g ml�1) or ampicillin (100 �g ml�1) when appropriate.Axenic Acanthamoeba castellanii cells were grown on proteose-yeast extract-glucose medium at 30°C and split once a week. Dictyostelium discoideumDd04 expressing calnexin-green fluorescent protein (GFP) (DBS0236184)was obtained from the Dicty Stock Center (http://dictybase.org/StockCenter/StockCenter.html; depositor, A. Muller-Taubenberger). D. discoideumcells were grown axenically at 22°C in HL5 medium supplemented withG418 at 20 �g ml�1 when necessary. Human acute monocytic leukemia(THP1) cells and U937 cells were maintained at 37°C and 5% CO2 inRPMI 1640 medium supplemented with 10% heat-inactivated fetal calfserum. Differentiation into macrophages was triggered by the addition ofphorbol 12-myristate 13-acetate at a final concentration of 100 ng ml�1.
Plasmid constructions. All DNA constructions were made by using E.coli XL1 Blue or DH5� as the host. L. pneumophila Lens and Paris genomicDNAs were used as the templates for PCRs with the primers listed in TableS2 in the supplemental material. Plasmids pXDC50 (28) and pXDC61(29) were obtained from Xavier Charpentier (see Table S1 in the supple-mental material). All recombinant plasmids were systematically verifiedby enzymatic digestion before electroporation into L. pneumophila (2,400V, 200 �, 25 �F).
Gene inactivation in L. pneumophila. Gene-specific knockout strainswere constructed by the homologous-recombination method. Briefly,2-kb flanking regions located upstream and downstream of the gene to beinactivated were amplified by PCR. A kanamycin resistance cassette wasinserted between the two regions by a double-joint PCR. To obtain mu-tant strains derived from wild-type (WT) Lens, the final PCR fragmentwas cloned into the pLaw344 plasmid (30), which can be selected onchloramphenicol and counterselected on sucrose-supplemented me-dium. After electroporation and selection, Cmr clones containing theplasmid were grown for 24 h in liquid medium without antibiotic andspread onto BCYE agar plates containing kanamycin (10 �g ml�1) andsucrose (5% final concentration). Recombinant clones were then ana-lyzed to confirm the deletion of the target gene(s) and the loss of pLaw344.To knock out genes in strain Paris, bacteria were grown overnight at 37°Cto an optical density at 600 nm (OD600) of 1 to 1.2. One to two micro-grams of a purified PCR DNA fragment containing the resistance cassetteflanked by the two 2-kb homologous regions were added to the bacteria.The tubes were incubated overnight at 30°C without shaking. Bacteria
were then plated on BCYE agar containing 20 �g ml�1 kanamycin. Re-combinant clones were verified by PCR.
Intracellular growth. A. castellanii cells were seeded into a 96-wellmicroplate at 1 � 105/well and allowed to adhere for at least 2 h at 30°C.After washing with peptone-yeast extract medium, amoebas were infectedat a multiplicity of infection (MOI) of 10 with bacterial suspensions madeby dilution of late-stationary-phase cultures of L. pneumophila strainsexpressing mCherry (pXDC50). Plates were then centrifuged for 10 min at600 � g to phase bacterium-cell contact and incubated for 1 h at 30°C.Amoeba were washed three times to remove nonadherent bacteria, andplates were incubated at 30°C for 48 h.
D. discoideum cells were seeded at 1 � 105/well and allowed to adherefor 16 h at 22°C. Infections were performed in the same way by using MBmedium (yeast extract, 7.15 g liter�1; peptone, 14.3 g liter�1; MES [mor-pholineethanesulfonic acid], 3.9 g liter�1; pH 6.9) at 25°C.
Intracellular growth was monitored by measuring the intensity of flu-orescence emitted by mCherry (excitation wavelength, 580 nm; emissionwavelength, 620 nm) with a TECAN InfinitePro plate reader. The platereader was thermostat regulated at 25 or 30°C for D. discoideum or A.castellanii, respectively.
Cytotoxicity assays. Cell viability, hence, bacterium-induced celldeath or cytotoxicity, was measured by using the alamarBlue viabilityindicator according to the manufacturer’s protocol (Life Technologies).Briefly, after 24 or 48 h of incubation, infected cells were washed threetimes in the appropriate medium and 100 �l of 10% alamarBlue diluted inmedium was added to each well. Plates were incubated at the temperaturesuitable for the cells considered for 4 to 16 h, and absorbances at 570 and600 nm were recorded with a BioTek Instruments �Quant plate reader.Cell viability was estimated by comparing the 570-nm/600-nm absor-bance ratio with the ratio obtained with noninfected cells.
Recruitment of the endoplasmic reticulum (ER) to the LCV in D.discoideum. D. discoideum cells producing calnexin-GFP were seededinto sterile glass coverslips on six-well plates at 5 � 106/well in HL5 me-dium and allowed to adhere overnight. Monolayers were infected at anMOI of 100 with mCherry-producing L. pneumophila grown for 4 days at30°C. The plates were centrifuged at 880 � g for 10 min and incubated for1 h at 25°C. The medium was then carefully removed, and the monolayerswere fixed with 3.7% paraformaldehyde (30 min, 4°C). Coverslips wereexamined with an inverted confocal microscope (Axiovert 200M; Zeiss,Thornwood, NJ).
Pore-forming activity assays. The pore-forming activity assay usedwas previously described by Kirby et al. (31). Briefly, A. castellanii cellswere used to seed coverslips placed on a six-well plate and infected at anMOI of 500 for 1 h at 30°C. The coverslips were then inverted onto a 5-�ldrop of phosphate-buffered saline containing 25 �g ml�1 ethidium bro-mide and 5 �g ml�1 acridine orange placed on a glass side. Coverslipswere immediately observed by using appropriate filters with a Zeiss fluo-rescence microscope.
�-Lactamase fusion construction and enzymatic-activity assay. Thelast 162 or 731 amino acids of the coding sequence of the L. pneumophilaParis rtxA gene were PCR amplified with the primers listed in Table S2 inthe supplemental material. The resulting PCR fragments were digestedand cloned into pXDC61 in frame with the 3= terminus of the blaM gene(with the sequence encoding the signal peptide deleted), creating transla-tional fusions Bla-RtxA162 and Bla-RtxA731, respectively.
L. pneumophila strains carrying pblaM-rtxA162, pblaM-rtxA731, orpblaM-fabI (no-secretion control) were grown in BYE liquid medium tostationary phase. Cells were then pelleted by centrifugation, adjusted to anOD600 of 20 in BYE medium supplemented with 1 mM isopropyl-�-D-thiogalactopyranoside (IPTG) to induce the production of the hybridproteins, and incubated for 3 h at 30°C without shaking. Bacterial suspen-sions were then added to a sterile paper disk placed onto an E. coli MG1655layer made on an ampicillin-supplemented LB agar plate, and the growthof E. coli around the disk was monitored after a 16-h incubation at 37°C.Supernatants were also collected after centrifugation and dropped onto an
Fuche et al.
564 jb.asm.org February 2015 Volume 197 Number 3Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

E. coli MG1655 layer made on an ampicillin-supplemented LB agar plate.Growth of E. coli was monitored as previously mentioned.
mCherry fusion construction and secretion assay. The blaM-con-taining fragments of plasmids pblaM-rtxA162 and pblaM-rtxA731 were ex-cised (NdeI/KpnI), and the PCR-amplified mCherry-encoding gene wascloned into the same sites, generating pmCherry-rtxA162 and pmCherry-rtxA731, respectively. Plasmids plssBDh, ptolCh, and plssBD-tolCh wereconstructed by cloning the lssBD, tolC, or lssBD and tolC genes into thep15A-derived plasmid pACYC184kan under the control of the constitu-tive pKan promoter. For the secretion assay, E. coli MG1655 strains car-rying one fusion-expressing plasmid and one T1SS gene-expressing plas-mid were grown to late exponential phase, centrifuged, and resuspendedin fresh LB medium supplemented with 1 mM IPTG. Induction of hybridprotein expression was carried out at 37°C for 30 min, the supernatant wascollected, and mCherry fluorescence was monitored in a 96-well micro-plate for 16 h at 30°C in a TECAN InfinitePro plate reader.
RESULTSL. pneumophila encodes two putative members of a T1SS. Jacobiand Heuner identified two genes, lssB and lssD, of L. pneumophilaas part of a putative T1SS, based on similarity with toxin trans-porter systems of Vibrio cholerae, Salmonella typhi, and E. coli (14).Moreover, we showed in a previous study that a tolC gene is pres-ent in the L. pneumophila genome and that it is involved in earlysteps of host invasion, as a �tolC mutant exhibits impaired viru-lence for amoebas and macrophages (32). A similar phenotype hasalso been observed in a �rtxA mutant strain (26, 27). Carefullyanalyzing the LssB protein sequence, and also the classical ABC
transporter domains (a transmembrane domain and a nucleotidebinding domain), we noticed the presence of a C39 peptidase-likemotif (from position 16 to position 137) that has been recentlyreported as a signature of T1SSs that export RTX proteins (33). Inthe case of a functional C39 peptidase, the T1SS is involved in theexport of bacteriocins, and the corresponding activity (C39 pep-tidase) is essential for the maturation of the secreted bacteriocins(cleavage downstream of a GG pattern of a prodomain). At thedifference, the C39 peptidase-like motif corresponds to a degen-erated catalytic site of a C39 peptidase and the exported proteinsall belong to the RTX family and are not matured during export.
We therefore hypothesized that LssB-LssD-TolC acts as a func-tional T1SS and has a role in the virulence of this bacterium, pre-sumably by allowing L. pneumophila to secrete the RtxA protein.Therefore, we inactivated the lssB, lssD, and rtxA genes in theepidemic strain Lens, as well as in the endemic strain Paris, byhomologous recombination and studied the roles of these genes inthe physiology and virulence of the bacteria. It should be notedthat lssB and lssD have been found to be the last genes of a six-geneoperon by studying mRNA synthesis (14), therefore reducing apossible polar effect of inactivation. The roles of the other genesproducts within this operon have yet to be documented. The rtxAgene is annotated on genomes with its own transcript unit.
The Legionella lssB and lssD genes are required for intracel-lular replication. As a preliminary test, the growth of each strainwas followed by measuring absorbance at 600 nm. No growth
FIG 1 Intracellular replication of L. pneumophila Lens and mutant derivatives in eucaryotic host cells. Intracellular replication of the �lssBD mutant in A.castellanii (A) and D. discoideum (B) was monitored by measuring fluorescence intensity with a TECAN plate reader (excitation wavelength, 580 nm; emissionwavelength, 620 nm). All bacteria expressed the mCherry fluorescent protein under the control of an IPTG-inducible promoter (plasmid pXDC50). The resultsare representative of at least four infection experiments (MOI 10) performed in triplicate (errors bars are indicated). RLU, relative fluorescence units.
T1SS Involved in L. pneumophila Virulence
February 2015 Volume 197 Number 3 jb.asm.org 565Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
difference between the WT and mutant strains could be observed,indicating that lssB and lssD are not essential for optimal growth inthe laboratory (see Fig. S1 in the supplemental material). Theability to replicate within different hosts was then investigated byusing the strains carrying pXDC50. The intensity of mCherry flu-orescence was monitored for 2 to 6 days after contact with twomodels of environmental hosts, A. castellanii and D. discoideum(Fig. 1A and B). As expected, a �dotA mutant cannot replicatewithin amoebas. This mutant is unable to assemble a functionalT4SS, which is necessary for the translocation of many effectors inthe cytosol of infected cells and for intracellular growth (34). WTstrain Lens showed detectable replication after 8 h in A. castellaniiand 40 h in D. discoideum (Fig. 1A and B), the difference in timebeing attributed to the difference in the temperatures used in theseinfection experiments. For A. castellanii, the optimal temperatureis 30°C, while D. discoideum infections were carried out at 25°C(closer to the optimum temperature of growth for these amoebas,22°C). A delay in the appearance of �lssBD mutant strain replica-tion in both hosts was clearly observed (38 and 105 h, respec-tively). A �lssBD mutant strain carrying a plasmid with the WTcopy of the native lssB and lssD genes exhibits an intracellularreplication profile in A. castellanii that is similar to the WT strainprofile. This complementation is only partial in D. discoideum, butthe difference from the �lssBD mutant strain profile is still signif-icant.
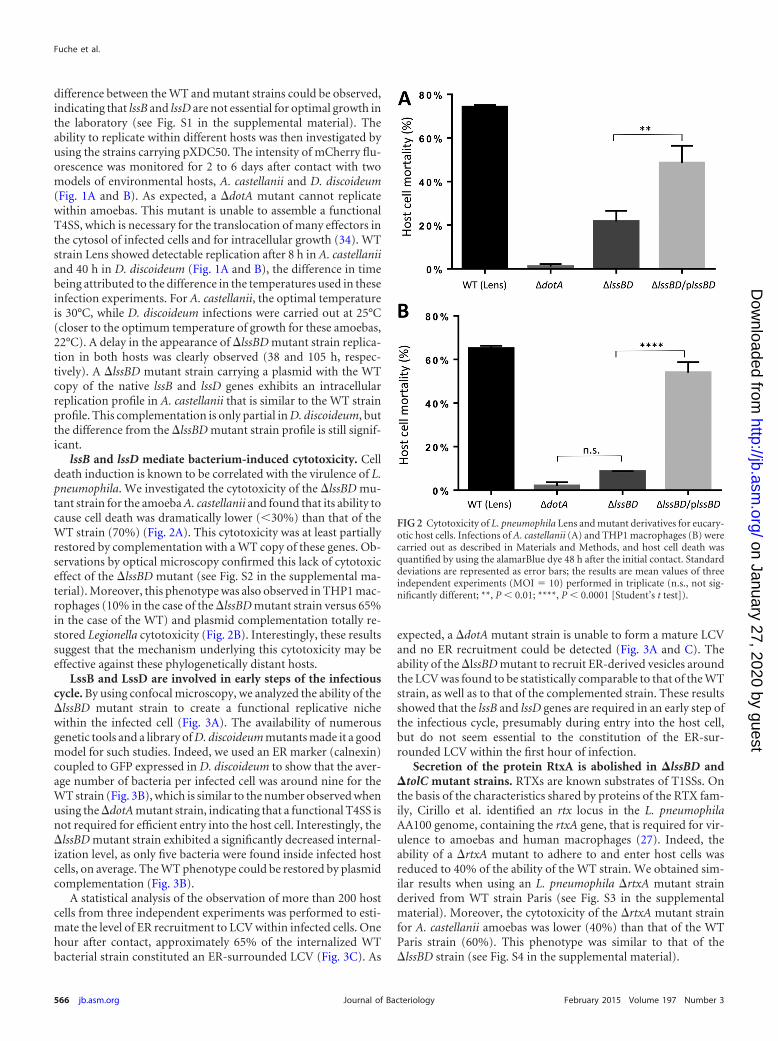
lssB and lssD mediate bacterium-induced cytotoxicity. Celldeath induction is known to be correlated with the virulence of L.pneumophila. We investigated the cytotoxicity of the �lssBD mu-tant strain for the amoeba A. castellanii and found that its ability tocause cell death was dramatically lower (30%) than that of theWT strain (70%) (Fig. 2A). This cytotoxicity was at least partiallyrestored by complementation with a WT copy of these genes. Ob-servations by optical microscopy confirmed this lack of cytotoxiceffect of the �lssBD mutant (see Fig. S2 in the supplemental ma-terial). Moreover, this phenotype was also observed in THP1 mac-rophages (10% in the case of the �lssBD mutant strain versus 65%in the case of the WT) and plasmid complementation totally re-stored Legionella cytotoxicity (Fig. 2B). Interestingly, these resultssuggest that the mechanism underlying this cytotoxicity may beeffective against these phylogenetically distant hosts.
LssB and LssD are involved in early steps of the infectiouscycle. By using confocal microscopy, we analyzed the ability of the�lssBD mutant strain to create a functional replicative nichewithin the infected cell (Fig. 3A). The availability of numerousgenetic tools and a library of D. discoideum mutants made it a goodmodel for such studies. Indeed, we used an ER marker (calnexin)coupled to GFP expressed in D. discoideum to show that the aver-age number of bacteria per infected cell was around nine for theWT strain (Fig. 3B), which is similar to the number observed whenusing the �dotA mutant strain, indicating that a functional T4SS isnot required for efficient entry into the host cell. Interestingly, the�lssBD mutant strain exhibited a significantly decreased internal-ization level, as only five bacteria were found inside infected hostcells, on average. The WT phenotype could be restored by plasmidcomplementation (Fig. 3B).
A statistical analysis of the observation of more than 200 hostcells from three independent experiments was performed to esti-mate the level of ER recruitment to LCV within infected cells. Onehour after contact, approximately 65% of the internalized WTbacterial strain constituted an ER-surrounded LCV (Fig. 3C). As
expected, a �dotA mutant strain is unable to form a mature LCVand no ER recruitment could be detected (Fig. 3A and C). Theability of the �lssBD mutant to recruit ER-derived vesicles aroundthe LCV was found to be statistically comparable to that of the WTstrain, as well as to that of the complemented strain. These resultsshowed that the lssB and lssD genes are required in an early step ofthe infectious cycle, presumably during entry into the host cell,but do not seem essential to the constitution of the ER-sur-rounded LCV within the first hour of infection.
Secretion of the protein RtxA is abolished in �lssBD and�tolC mutant strains. RTXs are known substrates of T1SSs. Onthe basis of the characteristics shared by proteins of the RTX fam-ily, Cirillo et al. identified an rtx locus in the L. pneumophilaAA100 genome, containing the rtxA gene, that is required for vir-ulence to amoebas and human macrophages (27). Indeed, theability of a �rtxA mutant to adhere to and enter host cells wasreduced to 40% of the ability of the WT strain. We obtained sim-ilar results when using an L. pneumophila �rtxA mutant strainderived from WT strain Paris (see Fig. S3 in the supplementalmaterial). Moreover, the cytotoxicity of the �rtxA mutant strainfor A. castellanii amoebas was lower (40%) than that of the WTParis strain (60%). This phenotype was similar to that of the�lssBD strain (see Fig. S4 in the supplemental material).
FIG 2 Cytotoxicity of L. pneumophila Lens and mutant derivatives for eucary-otic host cells. Infections of A. castellanii (A) and THP1 macrophages (B) werecarried out as described in Materials and Methods, and host cell death wasquantified by using the alamarBlue dye 48 h after the initial contact. Standarddeviations are represented as error bars; the results are mean values of threeindependent experiments (MOI 10) performed in triplicate (n.s., not sig-nificantly different; **, P 0.01; ****, P 0.0001 [Student’s t test]).
Fuche et al.
566 jb.asm.org February 2015 Volume 197 Number 3Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

Although the secretion signal for T1SS substrates is not con-served, it has been shown to be uncleaved and located in the last 50to 60 C-terminal residues of the substrate proteins (21). To assesswhether LssB, LssD, and TolC are able to export Legionella RtxAprotein, we constructed two different translational fusions of thegene encoding the �-lactamase (blaM) with its signal sequencedeleted and 3= portions of the rtxA gene respectively encoding 162and 731 amino acids of the RtxA C-terminal region. A blaM-fabIfusion was used as a negative control for secretion, as FabI is an L.pneumophila cytoplasmic protein (29). The production of all threehybrid proteins was assessed by Western blotting with anti-BlaMantibodies (see Fig. 5). Comparable amounts of protein were de-tected with BlaM, BlaM-RtxA731, and BlaM-FabI, but it seems thatsmaller amounts were produced in the case of BlaM-RtxA162.Moreover, two bands can be seen for hybrid proteins, certainlybecause of the instability of the fusion products. The secretion ofthese hybrid proteins was then studied by monitoring the growthof ampicillin-sensitive E. coli in the presence of L. pneumophilaParis and mutant derivatives producing these hybrid proteins.This test and the time of study were designed to limit the back-ground, resulting in the detection of hybrid proteins released after
cell lysis, which was followed with BlaM-FabI fusion protein.Strain Paris and its �lssBD derivative mutant were used for theseexperiments, and the results presented in Fig. 4A showed that thesecretion of the two BlaM-RtxA162 and BlaM-RtxA731 hybrid pro-teins was abolished in a �lssBD genetic background (no growth ofE. coli around the central disk containing the Legionella strain)compared to the active secretion in a WT genetic background. TheBlaM-FabI hybrid did not allow good growth of the E. coli ampi-cillin-sensitive strain, indicating that there was no secretion of thehybrid protein and no lysis (WT) or very little lysis (�lssBD) of theL. pneumophila strain during the time of the experiment. Asimilar experiment was performed with cell-free culture super-natant of hybrid-producing L. pneumophila and showed thatthe TolC protein is also involved in the secretion of the BlaM-RtxA731 hybrid protein as ampicillin was degraded and allowedthe ampicillin-sensitive E. coli strain to grow around the super-natant drop (Fig. 4B).
RtxA is secreted in a LssB-LssD-TolC-dependent manner. Toensure that the secretion of the hybrid proteins observed in an L.pneumophila WT background is specific to the presence of LssB,LssD, and TolC, the corresponding genes, as well as the genes
FIG 3 L. pneumophila entry into host cells and ER recruitment to the LCV. D. discoideum cells expressing calnexin-GFP were used to seed glass coverslips andinfected with L. pneumophila WT Lens and mutant derivatives expressing mCherry (MOI 100). After 1 h, the coverslips were fixed, mounted. and observed byconfocal microscopy (A). Statistical analysis was performed by observing at least 200 host cells in three independent experiments, and the average number ofbacteria in each cell (B) and the proportion of internalized bacteria actually recruiting ER to their LCV (C) are shown (n.s., not significantly different; ****, P 0.0001 [Student’s t test]). The symbol ‡ indicates that no statistical analysis could be performed because the �dotA mutant is, by definition, unable to recruit ER.
T1SS Involved in L. pneumophila Virulence
February 2015 Volume 197 Number 3 jb.asm.org 567Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

encoding new hybrid proteins mCherry-RtxA162 and mCherry-RtxA731, were expressed in an E. coli background (WT strainMG1655). The mCherry fluorescent protein was chosen becauseof its low folding rate (35), as it was shown that rapid folding of thesubstrate prior to secretion by the T1SS could block its export(36). The secretion of mCherry-RtxA hybrid proteins can be eval-uated by monitoring the fluorescence intensity at 620 nm in theculture supernatant (excitation wavelength, 580 nm). The resultsshowed that the three proteins LssB, LssD, and TolC are requiredfor efficient secretion of the two hybrid proteins tested in thisheterologous background (Fig. 5). We also noticed a higher levelof secretion (more than twice as high) when using the shortest
RtxA end (162 amino acid residues). Since those two hybrid pro-teins were produced at similar levels (data not shown), we as-sumed that this is due to inappropriate folding of the larger hybridprotein. The expression of the LssB and LssD proteins in the ab-sence of the Legionella TolC protein led to a low level of fusionprotein secretion (increase by a factor of up to five compared tothe control strain) that might result from E. coli TolC proteininvolvement but does not appear statistically significant. It isworth noting that expression of the Legionella TolC protein alonedid not result in fusion protein secretion by E. coli.
The lssB, lssD, and tolC genes are required for the RtxA-me-diated pore-forming activity of L. pneumophila. In addition to
FIG 4 Secretion assay. The secretion of BlaM-RtxA hybrid proteins in the L. pneumophila Paris WT or �lssBD or �tolC mutant background was assayed bymonitoring the growth of an ampicillin-sensitive E. coli strain plated on ampicillin-containing medium. Growth can be restored if the ampicillin on the agar plateis degraded by the secreted �-lactamase activity. L. pneumophila strains expressing hybrid proteins were dropped onto a sterile paper disk, and production ofhybrid proteins by bacteria prior to the test was verified by Western blotting of whole-cells extracts (anti-BlaM) (A), or cell-free culture supernatants weredropped directly onto the E. coli monolayer (B).
Fuche et al.
568 jb.asm.org February 2015 Volume 197 Number 3Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
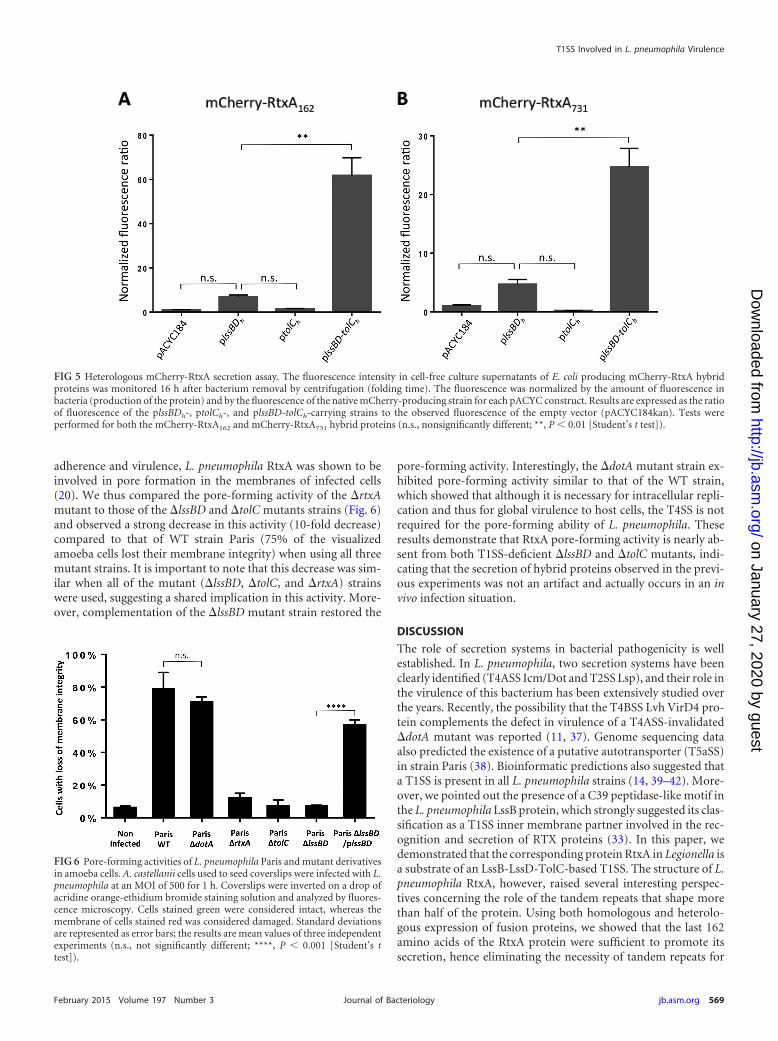
adherence and virulence, L. pneumophila RtxA was shown to beinvolved in pore formation in the membranes of infected cells(20). We thus compared the pore-forming activity of the �rtxAmutant to those of the �lssBD and �tolC mutants strains (Fig. 6)and observed a strong decrease in this activity (10-fold decrease)compared to that of WT strain Paris (75% of the visualizedamoeba cells lost their membrane integrity) when using all threemutant strains. It is important to note that this decrease was sim-ilar when all of the mutant (�lssBD, �tolC, and �rtxA) strainswere used, suggesting a shared implication in this activity. More-over, complementation of the �lssBD mutant strain restored the
pore-forming activity. Interestingly, the �dotA mutant strain ex-hibited pore-forming activity similar to that of the WT strain,which showed that although it is necessary for intracellular repli-cation and thus for global virulence to host cells, the T4SS is notrequired for the pore-forming ability of L. pneumophila. Theseresults demonstrate that RtxA pore-forming activity is nearly ab-sent from both T1SS-deficient �lssBD and �tolC mutants, indi-cating that the secretion of hybrid proteins observed in the previ-ous experiments was not an artifact and actually occurs in an invivo infection situation.
DISCUSSION
The role of secretion systems in bacterial pathogenicity is wellestablished. In L. pneumophila, two secretion systems have beenclearly identified (T4ASS Icm/Dot and T2SS Lsp), and their role inthe virulence of this bacterium has been extensively studied overthe years. Recently, the possibility that the T4BSS Lvh VirD4 pro-tein complements the defect in virulence of a T4ASS-invalidated�dotA mutant was reported (11, 37). Genome sequencing dataalso predicted the existence of a putative autotransporter (T5aSS)in strain Paris (38). Bioinformatic predictions also suggested thata T1SS is present in all L. pneumophila strains (14, 39–42). More-over, we pointed out the presence of a C39 peptidase-like motif inthe L. pneumophila LssB protein, which strongly suggested its clas-sification as a T1SS inner membrane partner involved in the rec-ognition and secretion of RTX proteins (33). In this paper, wedemonstrated that the corresponding protein RtxA in Legionella isa substrate of an LssB-LssD-TolC-based T1SS. The structure of L.pneumophila RtxA, however, raised several interesting perspec-tives concerning the role of the tandem repeats that shape morethan half of the protein. Using both homologous and heterolo-gous expression of fusion proteins, we showed that the last 162amino acids of the RtxA protein were sufficient to promote itssecretion, hence eliminating the necessity of tandem repeats for
FIG 5 Heterologous mCherry-RtxA secretion assay. The fluorescence intensity in cell-free culture supernatants of E. coli producing mCherry-RtxA hybridproteins was monitored 16 h after bacterium removal by centrifugation (folding time). The fluorescence was normalized by the amount of fluorescence inbacteria (production of the protein) and by the fluorescence of the native mCherry-producing strain for each pACYC construct. Results are expressed as the ratioof fluorescence of the plssBDh-, ptolCh-, and plssBD-tolCh-carrying strains to the observed fluorescence of the empty vector (pACYC184kan). Tests wereperformed for both the mCherry-RtxA162 and mCherry-RtxA731 hybrid proteins (n.s., nonsignificantly different; **, P 0.01 [Student’s t test]).
FIG 6 Pore-forming activities of L. pneumophila Paris and mutant derivativesin amoeba cells. A. castellanii cells used to seed coverslips were infected with L.pneumophila at an MOI of 500 for 1 h. Coverslips were inverted on a drop ofacridine orange-ethidium bromide staining solution and analyzed by fluores-cence microscopy. Cells stained green were considered intact, whereas themembrane of cells stained red was considered damaged. Standard deviationsare represented as error bars; the results are mean values of three independentexperiments (n.s., not significantly different; ****, P 0.001 [Student’s ttest]).
T1SS Involved in L. pneumophila Virulence
February 2015 Volume 197 Number 3 jb.asm.org 569Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

recognition and/or translocation through the T1SS. We are cur-rently investigating the localization of RtxA during the infection ofhost cells by L. pneumophila, as it was shown in Pseudomonasfluorescens that the RtxA homolog LapA was a surface-associatedadhesin (43, 44). Although the roles of the different regions ofsuch large RTX proteins remain to be experimentally assessed, thevariability in the number and nature of the tandem repeats be-tween the RtxA proteins of different strains of L. pneumophila maysuggest a modulation of the virulence based on these repeats.
Additionally, we documented the phenotype of an L. pneumo-phila strain defective for this T1SS. The �lssBD mutant straindisplayed strongly attenuated virulence for A. castellanii and D.discoideum. Moreover, preliminary results showed moderately at-tenuated virulence for U937 macrophages (data not shown). Wewere able to study the early steps of the infectious cycle, demon-strating that a �lssBD mutant strain is defective for entry into hostcells, which is consistent with the observations of Cirillo et al., whoused a �rtxA mutant strain (26). The absence of a T1SS did notseem to cause a defect in the creation of the replicative LCV, as therecruitment of ER-derived vacuoles to the surface of the phago-some is not altered after 1 h of infection. This recruitment is con-sidered a good marker of the constitution of the LCV and of suc-cessful hijacking of host vesicular trafficking (28). Therefore, wehypothesized that the initial entry into the host cell was responsi-ble for the general intracellular replication defect observed. How-ever, the attenuation of entry may not to be sufficient to explainthe quasiabolished virulence for amoebas, suggesting that the in-creased phagosome-lysosome fusion observed in the �rtxA mu-tant by Cirillo and coworkers might also be involved in this phe-notype.
Altogether, these results demonstrate for the first time thefunctionality of a T1SS in L. pneumophila and underline its im-portance in the virulence of this pathogen. Indeed, the lss locusseems to be present in the majority of the 217 L. pneumophilaisolates analyzed by Cazalet and coworkers (25). The rtxA geneencoding the T1SS substrate is also present in all of the isolatestested, though its structure is different between several straingroups. These results underline the active participation of L. pneu-mophila in its internalization into host cells via its T1SS and itscognate substrate RtxA.
ACKNOWLEDGMENTS
Fabien Fuche was supported by a fellowship from the Ministère del’Enseignement Supérieur et de la Recherche (France).
We are grateful to Nathalie Bailo for technical assistance, especially inmicroscopy. We also thank Xavier Charpentier (University of Lyon) forkindly providing the pXDC50 and pXDC61 plasmids.
REFERENCES1. Shelton BG, Kerbel W, Witherell L, Millar JD. 2000. Review of Legion-
naires’ disease. AIHAJ 61:738 –742. http://dx.doi.org/10.1202/0002-8894(2000)0610738:ROLD�2.0.CO;2.
2. Sharpe BA, Flanders SA. 2006. Community-acquired pneumonia: apractical approach to management for the hospitalist. J Hosp Med 1:177–190. http://dx.doi.org/10.1002/jhm.95.
3. Stout JE, Muder RR, Mietzner S, Wagener MM, Perri MB, DeRoos K,Goodrich D, Arnold W, Williamson T, Ruark O, Treadway C, EcksteinEC, Marshall D, Rafferty ME, Sarro K, Page J, Jenkins R, Oda G,Shimoda KJ, Zervos MJ, Bittner M, Camhi SL, Panwalker AP, DonskeyCJ, Nguyen MH, Holodniy M, Yu VL, Legionella Study G. 2007. Role ofenvironmental surveillance in determining the risk of hospital-acquiredlegionellosis: a national surveillance study with clinical correlations. InfectControl Hosp Epidemiol 28:818 – 824. http://dx.doi.org/10.1086/518754.
4. Fields BS, Benson RF, Besser RE. 2002. Legionella and Legionnaires’disease: 25 years of investigation. Clin Microbiol Rev 15:506 –526. http://dx.doi.org/10.1128/CMR.15.3.506-526.2002.
5. Xu L, Luo Z-Q. 2013. Cell biology of infection by Legionella pneumophila.Microbes Infect 15:157–167. http://dx.doi.org/10.1016/j.micinf.2012.11.001.
6. Tseng T-T, Tyler BM, Setubal JC. 2009. Protein secretion systems inbacterial-host associations, and their description in the Gene Ontol-ogy. BMC Microbiol 9(Suppl 1):S2. http://dx.doi.org/10.1186/1471-2180-9-S1-S2.
7. Liles MR, Viswanathan VK, Cianciotto NP. 1998. Identification andtemperature regulation of Legionella pneumophila genes involved in typeIV pilus biogenesis and type II protein secretion. Infect Immun 66:1776 –1782.
8. Berger KH, Merriam JJ, Isberg RR. 1994. Altered intracellular targetingproperties associated with mutations in the Legionella-pneumophila dotAgene. Mol Microbiol 14:809 – 822. http://dx.doi.org/10.1111/j.1365-2958.1994.tb01317.x.
9. Brand BC, Sadosky AB, Shuman HA. 1994. The Legionella-pneumophila-icm locus—a set of genes required for intracellular multiplication in hu-man macrophages. Mol Microbiol 14:797– 808. http://dx.doi.org/10.1111/j.1365-2958.1994.tb01316.x.
10. Nagai H, Kubori T. 2011. Type IVB secretion systems of Legionella andother Gram-negative bacteria. Front Microbiol 2:136. http://dx.doi.org/10.3389/fmicb.2011.00136.
11. Bandyopadhyay P, Lang EAS, Rasaputra KS, Steinman HM. 2013.Implication of the VirD4 coupling protein of the Lvh type 4 secretionsystem in virulence phenotypes of Legionella pneumophila. J Bacteriol 195:3468 –3475. http://dx.doi.org/10.1128/JB.00430-13.
12. Zhu W, Banga S, Tan Y, Zheng C, Stephenson R, Gately J, Luo Z-Q.2011. Comprehensive identification of protein substrates of the Dot/Icmtype IV transporter of Legionella pneumophila. PLoS One 6(3):e17638.http://dx.doi.org/10.1371/journal.pone.0017638.
13. Allombert J, Fuche F, Michard C, Doublet P. 2013. Molecular mimicryand original biochemical strategies for the biogenesis of a Legionella pneu-mophila replicative niche in phagocytic cells. Microbes Infect 15:981–988.http://dx.doi.org/10.1016/j.micinf.2013.09.007.
14. Jacobi S, Heuner K. 2003. Description of a putative type I secretionsystem in Legionella pneumophila. Int J Med Microbiol 293:349 –358. http://dx.doi.org/10.1078/1438-4221-00276.
15. Brooks HJL, Ogrady F, McSherry MA, Cattell WR. 1980. Uropathogenicproperties of Escherichia-coli in recurrent urinary-tract infection. J MedMicrobiol 13:57– 68. http://dx.doi.org/10.1099/00222615-13-1-57.
16. Shrivastava R, Miller JF. 2009. Virulence factor secretion and transloca-tion by Bordetella species. Curr Opin Microbiol 12:88 –93. http://dx.doi.org/10.1016/j.mib.2009.01.001.
17. Bowen DJ, Rocheleau TA, Grutzmacher CK, Meslet L, Valens M,Marble D, Dowling A, Ffrench-Constant R, Blight MA. 2003. Geneticand biochemical characterization of PrtA, an RTX-like metalloproteasefrom Photorhabdus. Microbiology 149:1581–1591. http://dx.doi.org/10.1099/mic.0.26171-0.
18. Mackman N, Nicaud JM, Gray L, Holland IB. 1986. Secretion of hemo-lysin by Escherichia coli. Curr Top Microbiol Immunol 125:159 –181.
19. Lazdunski A, Guzzo J, Filloux A, Bally M, Murgier M. 1990. Secretionof extracellular proteins by Pseudomonas aeruginosa. Biochimie 72:147–156. http://dx.doi.org/10.1016/0300-9084(90)90140-C.
20. Létoffé S, Delepelaire P, Wandersman C. 1990. Protease secretion byErwinia chrysanthemi—the specific secretion functions are analogous tothose of Escherichia coli alpha-hemolysin. EMBO J 9:1375–1382.
21. Koronakis V, Eswaran J, Hughes C. 2004. Structure and function of tolC:The bacterial exit duct for proteins and drugs. Annu Rev Biochem 73:467–489. http://dx.doi.org/10.1146/annurev.biochem.73.011303.074104.
22. Masi M, Wandersman C. 2010. Multiple signals direct the assembly andfunction of a type 1 secretion system. J Bacteriol 192:3861–3869. http://dx.doi.org/10.1128/JB.00178-10.
23. Satchell KJF. 2011. Structure and function of MARTX toxins and otherlarge repetitive RTX proteins. Annu Rev Microbiol 65:71–90. http://dx.doi.org/10.1146/annurev-micro-090110-102943.
24. Delepelaire P. 2004. Type I secretion in Gram-negative bacteria. BiochimBiophys Acta 1694:149 –161. http://dx.doi.org/10.1016/j.bbamcr.2004.05.001.
25. Cazalet C, Jarraud S, Ghavi-Helm Y, Kunst F, Glaser P, Etienne J,Buchrieser C. 2008. Multigenome analysis identifies a worldwide distrib-
Fuche et al.
570 jb.asm.org February 2015 Volume 197 Number 3Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

uted epidemic Legionella pneumophila clone that emerged within a highlydiverse species. Genome Res 18:431– 441. http://dx.doi.org/10.1101/gr.7229808.
26. Cirillo SLG, Bermudez LE, El-Etr SH, Duhamel GE, Cirillo JD. 2001.Legionella pneumophila entry gene rtxA is involved in virulence. InfectImmun 69:508 –517. http://dx.doi.org/10.1128/IAI.69.1.508-517.2001.
27. Cirillo SLG, Yan L, Littman M, Samrakandi MM, Cirillo JD. 2002. Roleof the Legionella pneumophila rtxA gene in amoebae. Microbiology 148(Pt6):1667–1677.
28. Hervet E, Charpentier X, Vianney A, Lazzaroni JC, Gilbert C, Atlan D,Doublet P. 2011. Protein kinase LegK2 is a type IV secretion systemeffector involved in endoplasmic reticulum recruitment and intracellularreplication of Legionella pneumophila. Infect Immun 79:1936 –1950. http://dx.doi.org/10.1128/IAI.00805-10.
29. de Felipe KS, Glover RT, Charpentier X, Anderson OR, Reyes M,Pericone CD, Shuman HA. 2008. Legionella eukaryotic-like type IV sub-strates interfere with organelle trafficking. PLoS Pathog 4(8):e1000117.http://dx.doi.org/10.1371/journal.ppat.1000117.
30. Wiater LA, Sadosky AB, Shuman HA. 1994. Mutagenesis of Legionellapneumophila using Tn903 dIIlacz: identification of a growth-phase-regulated pigmentation gene. Mol Microbiol 11:641– 653. http://dx.doi.org/10.1111/j.1365-2958.1994.tb00343.x.
31. Kirby JE, Vogel JP, Andrews HL, Isberg RR. 1998. Evidence for pore-forming ability by Legionella pneumophila. Mol Microbiol 27:323–336.http://dx.doi.org/10.1046/j.1365-2958.1998.00680.x.
32. Ferhat M, Atlan D, Vianney A, Lazzaroni JC, Doublet P, Gilbert C.2009. The TolC protein of Legionella pneumophila plays a major role inmulti-drug resistance and the early steps of host invasion. PLoS One 4(11):e7732. http://dx.doi.org/10.1371/journal.pone.0007732.
33. Kanonenberg K, Schwarz CKW, Schmitt L. 2013. Type I secretion sys-tems—a story of appendices. Res Microbiol 164:596 – 604. http://dx.doi.org/10.1016/j.resmic.2013.03.011.
34. Roy CR, Berger KH, Isberg RR. 1998. Legionella pneumophila DotAprotein is required for early phagosome trafficking decisions that occurwithin minutes of bacterial uptake. Mol Microbiol 28:663– 674. http://dx.doi.org/10.1046/j.1365-2958.1998.00841.x.
35. Kelkar DA, Khushoo A, Yang Z, Skach WR. 2012. Kinetic analysis ofribosome-bound fluorescent proteins reveals an early, stable, cotransla-tional folding intermediate. J Biol Chem 287:2568 –2578. http://dx.doi.org/10.1074/jbc.M111.318766.
36. Debarbieux L, Wandersman C. 2001. Folded HasA inhibits its own se-
cretion through its ABC exporter. EMBO J 20:4657– 4663. http://dx.doi.org/10.1093/emboj/20.17.4657.
37. Bandyopadhyay P, Liu SQ, Gabbai CB, Venitelli Z, Steinman HM.2007. Environmental mimics and the Lvh type IVA secretion system con-tribute to virulence-related phenotypes of Legionella pneumophila. InfectImmun 75:723–735. http://dx.doi.org/10.1128/IAI.00956-06.
38. Cazalet C, Rusniok C, Bruggemann H, Zidane N, Magnier A, Ma L,Tichit M, Jarraud S, Bouchier C, Vandenesch F, Kunst F, Etienne J,Glaser P, Buchrieser C. 2004. Evidence in the Legionella pneumophilagenome for exploitation of host cell functions and high genome plasticity.Nat Genet 36:1165–1173. http://dx.doi.org/10.1038/ng1447.
39. Chien MC, Morozova I, Shi SD, Sheng HT, Chen J, Gomez SM,Asamani G, Hill K, Nuara J, Feder M, Rineer J, Greenberg JJ, SteshenkoV, Park SH, Zhao BH, Teplitskaya E, Edwards JR, Pampou S,Georghiou A, Chou IC, Iannuccilli W, Ulz ME, Kim DH, Geringer-Sameth A, Goldsberry C, Morozov P, Fischer SG, Segal G, Qu XY,Rzhetsky A, Zhang PS, Cayanis E, De Jong PJ, Ju JY, Kalachikov S,Shuman HA, Russo JJ. 2004. The genomic sequence of the accidentalpathogen Legionella pneumophila. Science 305:1966 –1968. http://dx.doi.org/10.1126/science.1099776.
40. Gomez-Valero L, Rusniok C, Jarraud S, Vacherie B, Rouy Z, Barbe V,Medigue C, Etienne J, Buchrieser C. 2011. Extensive recombinationevents and horizontal gene transfer shaped the Legionella pneumophilagenomes. BMC Genomics 12:536. http://dx.doi.org/10.1186/1471-2164-12-536.
41. Steinert M, Heuner K, Buchrieser C, Albert-Weissenberger C, GlocnerG. 2007. Legionella pathogenicity: genome structure, regulatory networksand the host cell response. Int J Med Microbiol 297:577–587. http://dx.doi.org/10.1016/j.ijmm.2007.03.009.
42. D’Auria G, Jimenez-Hernandez N, Peris-Bondia F, Moya A, Latorre A.2010. Legionella pneumophila pangenome reveals strain-specific virulencefactors. BMC Genomics 11:181. http://dx.doi.org/10.1186/1471-2164-11-181.
43. Newell PD, Boyd CD, Sondermann H, O’Toole GA. 2011. A c-di-GMPeffector system controls cell adhesion by inside-out signaling and surfaceprotein cleavage. PLoS Biol 9(2):e1000587. http://dx.doi.org/10.1371/journal.pbio.1000587.
44. Ivanov IE, Boyd CD, Newell PD, Schwartz ME, Turnbull L, JohnsonMS, Whitchurch CB, O’Toole GA, Camesano TA. 2012. Atomic forceand super-resolution microscopy support a role for LapA as a cell-surfacebiofilm adhesin of Pseudomonas fluorescens. Res Microbiol 163:685– 691.http://dx.doi.org/10.1016/j.resmic.2012.10.001.
T1SS Involved in L. pneumophila Virulence
February 2015 Volume 197 Number 3 jb.asm.org 571Journal of Bacteriology
on January 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from





























