Embed Size (px)
Citation preview

Functional synthetic Antennapedia genes and the dualroles of YPWM motif and linker size in transcriptionalactivation and repressionDimitrios K. Papadopoulosa,1, Diana Reséndez-Pérezb,1, Diana L. Cárdenas-Chávezb, Karina Villanueva-Segurab,Ricardo Canales-del-Castillob, Daniel A. Felixa,2, Raphael Fünfschillinga,3, and Walter J. Gehringa,4
aDepartment of Cell Biology, Biozentrum, University of Basel, 4056 Basel, Switzerland; and bFacultad de Ciencias Biológicas, Universidad Autónomade Nuevo Leon, Ciudad Universitaria, Monterrey 66450, NL, México
Contributed by Walter J. Gehring, June 2, 2011 (sent for review April 13, 2011)
Segmental identity along the anteroposterior axis of bilateralanimals is specified by Hox genes. These genes encode transcrip-tion factors, harboring the conserved homeodomain and, gener-ally, a YPWM motif, which binds Hox cofactors and increases Hoxtranscriptional specificity in vivo. Here we derive synthetic Dro-sophila Antennapedia genes, consisting only of the YPWM motifand homeodomain, and investigate their functional role through-out development. Synthetic peptides and full-length Antennapediaproteins cause head-to-thorax transformations in the embryo, aswell as antenna-to-tarsus and eye-to-wing transformations in theadult, thus converting the entire head to a mesothorax. This con-version is achieved by repression of genes required for head andantennal development and ectopic activation of genes promotingthoracic and tarsal fates, respectively. Synthetic Antennapedia pep-tides bind DNA specifically and interact with Extradenticle and Bric-à-brac interacting protein 2 cofactors in vitro and ex vivo. Substi-tution of the YPWM motif by alanines abolishes Antennapediahomeotic function, whereas substitution of YPWM by the WRPWrepressor motif, which binds the transcriptional corepressor Grou-cho, allows all proteins to act as repressors only. Finally, naturallyoccurring variations in the size of the linker between the homeo-domain and YPWM motif enhance Antennapedia repressive oractivating efficiency, emphasizing the importance of linker size,rather than sequence, for specificity. Our results clearly show thatsynthetic Antennapedia genes are functional in vivo and thereforeprovide powerful tools for synthetic biology. Moreover, the YPWMmotif is necessary—whereas the entire N terminus of the proteinis dispensable—for Antennapedia homeotic function, indicating itsdual role in transcriptional activation and repression by recruitingeither coactivators or corepressors.
Hox specificity | synthetic genes | homeotic transformations |antenna-to-tarsus transformation
Homeotic (Hox) genes encode transcription factors that specifybody segments along the anterior–posterior axis of bilaterians
(1–3). Hox transcription factors harbor the homeodomain (HD),a helix-turn-helix DNA-binding domain that has been highlyconserved throughout animal evolution and shares remarkablesequence similarity among different Hox paralogs (4–7). For Hoxproteins to confer segmental identity in the developing embryo,they need to regulate differentially—notwithstanding their se-quence similarity—a large number of target genes, most of thetime in a paralog-specific manner. Despite our current knowledgeof their DNA-binding properties (8), the identification of Hoxcofactors and “collaborators” that increase their target-selectionprecision (9), the possibility of performing genomewide analyses(10, 11), and our understanding of transcriptional regulationin vivo, we fall short of providing a complete explanation about thespecificity of Hox transcription factors that would allow us topredict or identify novel Hox functions.We have demonstrated previously that DNA-binding and the
functional specificity of Antennapedia (Antp) reside in the HDand its flexible N-terminal arm, respectively (8, 12–14), and that
the YPWM motif directly links Antp to the transcriptional ma-chinery (15). In the present study, we find that the YPWM motif,but not its flanking regions, is required forAntp homeotic functionin vivo. We derive synthetic Antp genes, which encode a smallportion of the full-length protein (spanning the region between theYPWMmotif and the beginning of the C terminus), and examinedtheir function in vivo, compared with their full-length counter-parts. We find that synthetic peptides and full-length Antp pro-teins featuring aYPWMmotif are able to generate head-to-thoraxtransformations in the embryo, to repress endogenous Sex combsreduced (Scr), and to activate Teashirt (Tsh) ectopically. They alsocause antenna-to-mesothoracic tarsus transformations throughrepression of Spalt major (Salm) and distal antenna (dan) and ec-topic activation of grain (grn) in the antennal disc. In a twin ofeyeless (toy)mutant background, which fails to launch the antennalmorphogenetic program (5), they additionally transform eyes towings. Moreover, they activate transcription and interact with theAntp cofactors Extradenticle (Exd) and Bric-à-brac interactingprotein 2 (Bip2) in vitro and inDrosophila tissue culture.When theYPWMmotif is replaced byWRPW, adomain interactingwith thegeneric transcriptional corepressor Groucho (Gro) (16), Antprepressive activity is retained in vivo. Finally, we evaluate the effectof either a long or a short linker between theYPWMmotif and theHD, comprising eight or four amino acids, respectively, whichoccur naturally in different Antp isoforms (17), and find that theyplay an important role in transcriptional potency and/or specificityin vivo.
ResultsThe Antp HD and YPWMMotifs Are Required for Homeotic Function inthe Embryo.We initially evaluated the requirement of the HD, theconserved YPWM motif, and its flanking regions for Antp func-tion in vivo. We constitutively expressed constructs that eitherlacked the entire HD and YPWM motif or contained alaninesubstitutions in either the YPWMmotif or in the four amino acidssituated N- or C-terminally to it (SI Appendix, Fig. S1), using thehsp70 promoter, as previously described (18). Abnormalities inthe development of the anterior (head) structures of the embryo,caused by transformation toward second thoracic segment iden-tity (T2), are caused upon ectopic expression of Antp in the seg-ments anterior to its normal expression domain (12, 19, 20). Thetransformation includes head-involution defects, repression of
Author contributions: D.K.P., D.R.-P., and W.J.G. designed research; D.K.P., D.R.-P., andD.L.C.-C. performed research; D.K.P., D.R.-P., D.L.C.-C., K.V.-S., R.C.-d.-C., D.A.F., and R.F.contributed new reagents/analytic tools; D.K.P., D.R.-P., D.L.C.-C., and W.J.G. analyzeddata; and D.K.P., D.R.-P., D.L.C.-C., and W.J.G. wrote the paper.
The authors declare no conflict of interest.1D.K.P. and D.R.-P. contributed equally to this work.2Present address: Institute of Genetics, Queen’s Medical Centre, University of Nottingham,Nottingham NG7 2UH, United Kingdom.
3Present address: Novartis Pharma AG, 4056 Basel, Switzerland.4To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1108686108/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1108686108 PNAS | July 19, 2011 | vol. 108 | no. 29 | 11959–11964
GEN
ETICS

prothoracic (T1) “beards,” and formation of ectopic denticlebands in the head segments (12, 20). Substitution of the YPWMmotif or deletion of both the HD and YPWM motif failed togenerate head-to-thorax transformations, whereas mutations ineither of the sequences flanking the YPWM motif generated aphenotype similar to the one caused by ectopic expression ofthe WT Antp protein (SI Appendix, Fig. S1). This result indicatesthat the HD and an intact YPWM motif are required for Antphomeotic function in vivo, whereas the amino acids flanking theYPWM motif are dispensable.
Synthetic Antp Peptides, Consisting of Only the HD and the YPWMMotif, Cause Homeotic Transformations in the Embryo. The func-tional requirement of the YPWMmotif prompted us to constructvarious versions of full-length and synthetic Antp genes to studythe functional specificity of each module and to derive the mini-mal requirements for Antp homeotic function in vivo. We usedconstructs with long and short linkers between the HD and theYPWM motif, replaced the YPWM motif by quadruple alanines(AAAA), fused additional repressor domains to the C terminusof the protein, replaced the YPWM motif with WRPW, and alsoconstructed synthetic Antp genes, comprising only the HD andYPWM motif but lacking the entire N terminus, to test whetherthey retain their homeotic function in vivo. For this purpose weused Upstream Activating Sequence (UAS) constructs and tag-ged the synthetic genes with eGFP (Fig. 1) to allow the visuali-zation and immunohistochemical detection of each peptide.Using the nullo-Gal4 driver (5) for strong expression of the Antpgenes at the preblastoderm and blastoderm stages of the embryo,we observed that all synthetic peptides and full-length Antpproteins featuring a YPWM motif cause head-involution defectsof variable strength and repression of the prothoracic beards (Fig.1 and SI Appendix, Fig. S2). In contrast, the full-length and syn-thetic AAAAAntp exhibited WT-like phenotypes with no re-pression of T1 beards (Fig. 1, lines 3 and 11 and SI Appendix, Fig.S2, line 12 and close-up). All synthetic and full-length constructsfeaturing a WRPW repressor motif (Fig. 1 and SI Appendix, Fig.S2) also caused head-to-thorax transformation and repression ofthe prothoracic beards. The most dramatic phenotype was dis-played by the full-length Antp with C-terminal fusions of WRPW(SI Appendix, Fig. S2, line 4 and 5), whereas the addition of theengrailed (en) repressor domain to the 3′ end of the full-lengthAntp (SI Appendix, Fig. S2, line 6) did not result in any phenotypeconsiderably different from the one caused by the WT Antp gene.In addition, we analyzed variations in the size of the linker regionbetween the YPWMmotif and the HD, consisting of four or eightamino acids that normally occur in different Antp isoforms (Fig.1, lines 1, 2, 9 and 10). Both full-length and synthetic YPWMAntpshort-linker peptides gave rise to a more pronounced head-in-volution defect with no detectable mouth parts in the cuticle.Therefore, the YPWM motif, but not the N terminus of Antp, isrequired for homeotic function in the embryo, with trans-formations being favored by short-linker isoforms. Replacing thetetrapeptide with WRPW retains part of its specificity in vivo.
The YPWM Motif of Antp Is Essential for Repression of EndogenousScr and Ectopic Activation of Tsh in the Embryo. Target genes re-pressed or activated by Antp in the embryo were then used asa system to determine the ability of synthetic peptides and full-length Antp proteins to act, directly or indirectly, as transcrip-tional repressors and activators. Such genes include Scr (21),normally expressed in the labial and T1 segments of the embryo,and Tsh, expressed in the trunk in parasegments 3–13 (22). Usingthe patched (ptc) enhancer (23) for ectopic Antp expression inthe anterior compartment of every parasegment, we observedpartial to complete repression of Scr by the full-length and syn-thetic Antp lines in the labial and T1 segments (Fig. 2A). Acomparison of the Scr protein levels among the different Antplines is shown in SI Appendix, Fig. S3. Interestingly, Antp full-length proteins and synthetic peptides bearing a YPWM-to-AAAA substitution did not repress Scr (Fig. 2A, lines 3 and 11and SI Appendix, Fig. S4A, line 12). A clear difference in re-pression potency was observed between the synthetic YPWMAntp
long- and short-linker genes, with the short-linker peptideexhibiting greater repressor activity than its long-linker counter-part (Fig. 2A, lines 9 and 10). However, the synthetic WRPWAntppeptides conferred only partial repression (SI Appendix, Fig. S4A,lines 13 and 14). The presence of a repressor motif (WRPW)seems to suffice for functionality of the transcription factor inthe repression of Scr. To ensure comparable expression levelsamong constructs and to compare the relative protein concen-tration of each line with the WT protein levels, we quantified therelative fluorescence intensity of each protein after staining withantibodies, as in ref. 24. Surprisingly, all constructs tested wereexpressed in approximately equal or lower amounts than theendogenous Antp protein, indicating that the observed pheno-types are not a result of overexpression (SI Appendix, Fig. S5).However, activation of Tsh by full-length proteins and syn-
thetic Antp peptides requires a functional YPWM motif (Fig. 2Band SI Appendix, Fig. S4B). Differences in the potency of ectopicTsh activation were also observed between the full-length (Fig.2B, lines 1 and 2) and synthetic YPWMAntp genes (Fig. 2B, lines 9and 10), but the trend was the opposite of that displayed inhomeotic transformations in the embryo or in the repression ofScr. In this case, the long-linker construct activated Tsh in a do-main as extensive as the full-length WT Antp, whereas the effect
Fig. 1. Full-length and synthetic Antp genes cause homeotic transfor-mations and head-involution defects in the embryo. (Upper) UAS Antpconstructs (full-length and synthetic) generated for the functional analysisof the YPWM motif and the N terminus. The short linker (SL) contains thesequence RSQF and the long linker (LL) contains the sequence RSQFGKCQ.The C terminus of synthetic peptides lacks the terminal 14 residues andcontains only the sequence TKGEPGS. All construct numbers are used forreference in the subsequent figures. (Lower) Comparative analysis of head-involution defects generated by constitutive expression of full-length andsynthetic Antp genes, using nullo-Gal4. Only proteins bearing a YPWM motif(Center and Right column) could induce transformation of head to mesothorax(T2). The size of thoracic segments is indicated by lines. The numbers 1–3 in-dicate segment identity: (1) prothoracic, (2) mesothoracic, or (3) metathoracic.Red arrows show the prothoracic beards, and blue asterisks indicate pro-nounced head-involution defects, where present. Note the mild head defects inthe synthetic AAAAAntp line, shown by the abnormal development of themouth hook.
11960 | www.pnas.org/cgi/doi/10.1073/pnas.1108686108 Papadopoulos et al.

of the short-linker construct was weaker. Therefore, the YPWMmotif is indispensable for transcriptional activation, but re-pression can be achieved at least partially by its substitution byWRPW. Also, the long linker between the HD and the YPWMmotif seems to favor Antp function in activation, whereas theshort linker favors repression of downstream genes.
Synthetic Antp Genes Featuring a Functional YPWM Motif CauseAntenna-to-Tarsus and Eye-to-Wing Transformations. We used theDistal-less (Dll) enhancer to generate tarsal transformations inthe antenna. Ectopic T2-legs bearing apical and/or preapicalbristles on the distal tibia were generated only by full-lengthproteins and synthetic Antp peptides featuring a functionalYPWM motif (Fig. 3A and SI Appendix, Fig. S6). As anticipated,no transformation or a mild thickening of the arista with a fewectopic leg bristles on the third antennal segment (A3) wereobserved with the full-length AAAA,LLAntp (Fig. 3A, line 3) andthe corresponding synthetic peptide (Fig. 3A, line 11), suggesting
mild transformation by the HD alone. The same rule applied toAAAA,LLAntpWRPW, where numerous tarsal bristles and absenceof the arista were identified on the antenna (SI Appendix, Fig. S6,line 5). Synthetic peptides and full-length Antp proteins withYPWM-to-WRPW substitutions caused overgrowth of the A3segment (SI Appendix, Fig. S6, lines 7, 8, 13 and 14), which oc-casionally displayed leg bristles. Finally, the antennal tarsi gen-erated by both the full-length and synthetic YPWMAntp lines alsoshowed a strong dependence on the size of the linker betweenthe HD and YPWM motif, with the long-linker constructs givingrise to more dramatic phenotypes than the short-linker ones(compare line 1 with 2 and line 9 with 10 in Fig. 3A). Thestrength of the transformation was assessed by (i) whether onlythe arista or both the arista and the third antennal segment weretransformed, (ii) the presence of claws on the distal tarsi, and(iii) whether tarsal and tibial or only tarsal structures could bedetected. Thus, a functional YPWM motif, but not the N ter-minus, is indispensable for Antp gain of function in the antenna.The WRPW cannot substitute for the tetrapeptide in this case;however, the linker size seems to play a considerable role, withthe long-linker lines favoring stronger transformations (SI Ap-pendix, Fig. S6, lines 7 and 8).However, Antp is responsible for specifying the entire meso-
thorax with a pair of legs (ventral appendages) and a pair of wings(dorsal appendages). We have shown previously that Antp caninduce ectopic wings in the dorsal head when misexpressed ina toymutant background (5). Because the toy gene is required forantennal morphogenesis, we expressed Antp ectopically in a hy-
Fig. 2. YPWM-dependent repression and activation of Antp target genes inthe embryo mediated by full-length and synthetic Antp genes. (A) Re-pression of Scr in the labial (lb, solid brackets) and prothoracic (T1, dashedbrackets) segments by full-length and synthetic Antp constructs. EctopicAntp is depicted in green and Scr in red. For each line a close-up of the re-gion of repression is shown, and below it an overview of an entire embryo isshown. Note the overlay of both gene products in the AAAA-substitutedlines, indicated by the yellow color. Staining in the WT embryo is for theendogenous proteins. (B) Ectopic activation of Tsh in the embryonic headsegments by full-length and synthetic Antp genes. Tsh is shown in red andectopic Antp in green. The upper image for each construct depicts Tsh, andthe lower image shows its overlay with ectopic Antp. The domain of ectopicTsh expression is outlined by dashed lines. Staining in the WT embryo is forthe endogenous genes. All UAS lines were induced using a ptc-Gal4 driver.
Fig. 3. Tarsal transformations caused by full-length proteins and syntheticAntp peptides. (A) The presence of the YPWM motif in Antp full-length andsynthetic peptides greatly enhances the transformation, but its absence insynthetic peptides does not completely abolish it. Note the leg bristles (blackarrows) that indicate weak transformation of the third antennal segment(A3) to a tarsus and the presence of apical/preapical bristles (red arrow-heads), indicating mesothoracic leg (T2) identity. Curved lines show antennalsegments. 2, second antennal segment; 3, third antennal segment; Ar, arista;transformed segments are identified in red type; tarsal claws are indicatedby blue asterisks. (B) Eye-to-wing transformations induced in a toyD2 mutantbackground. The synthetic genes behave comparably to the full-lengthAntp73b allele. W, wings; L, legs.
Papadopoulos et al. PNAS | July 19, 2011 | vol. 108 | no. 29 | 11961
GEN
ETICS
pomorphic toy mutant background (toyD2) using decapendaplegic(dppblink)-Gal4.Weobserved eye-to-wing transformationswith thesynthetic YPWM Antp constructs, comparable to the Antp73b gain-of-function allele (Fig. 3B), whereas the synthetic AAAAAntp linesdid not show this transformation. Again, as in the tarsal trans-formations described above, the long-linker construct conferreda more dramatic phenotype than the short-linker line (Fig. 3B,lines 9 and 10).
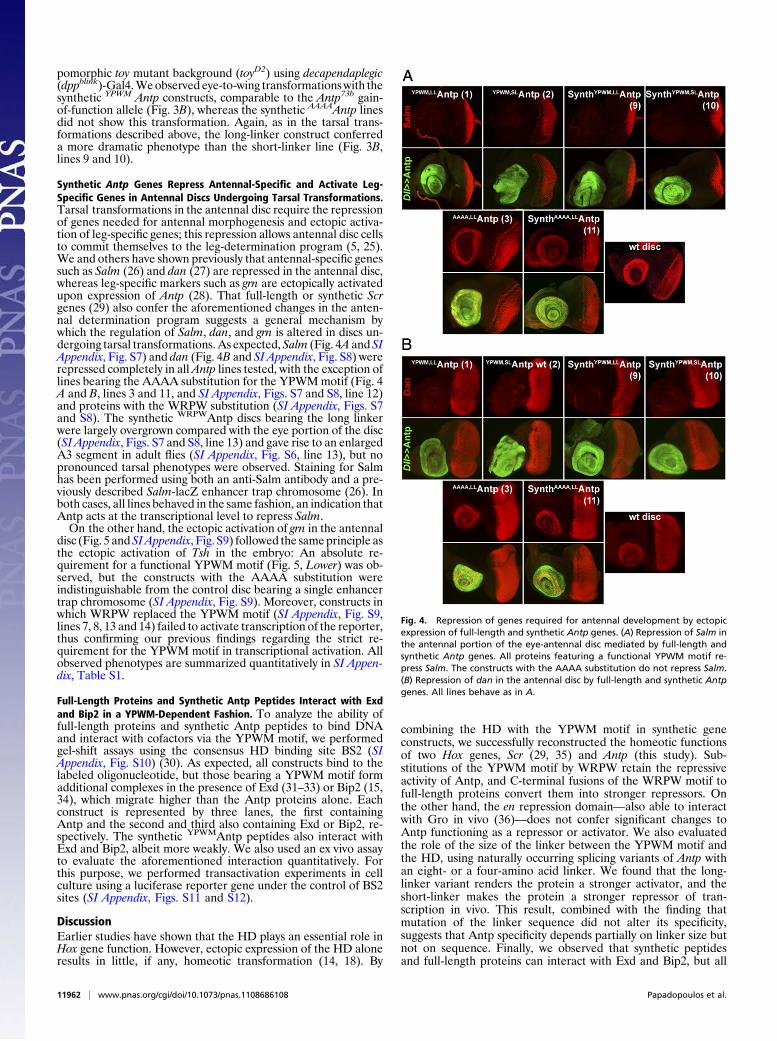
Synthetic Antp Genes Repress Antennal-Specific and Activate Leg-Specific Genes in Antennal Discs Undergoing Tarsal Transformations.Tarsal transformations in the antennal disc require the repressionof genes needed for antennal morphogenesis and ectopic activa-tion of leg-specific genes; this repression allows antennal disc cellsto commit themselves to the leg-determination program (5, 25).We and others have shown previously that antennal-specific genessuch as Salm (26) and dan (27) are repressed in the antennal disc,whereas leg-specific markers such as grn are ectopically activatedupon expression of Antp (28). That full-length or synthetic Scrgenes (29) also confer the aforementioned changes in the anten-nal determination program suggests a general mechanism bywhich the regulation of Salm, dan, and grn is altered in discs un-dergoing tarsal transformations. As expected, Salm (Fig. 4A andSIAppendix, Fig. S7) and dan (Fig. 4B and SI Appendix, Fig. S8) wererepressed completely in allAntp lines tested, with the exception oflines bearing the AAAA substitution for the YPWMmotif (Fig. 4A and B, lines 3 and 11, and SI Appendix, Figs. S7 and S8, line 12)and proteins with the WRPW substitution (SI Appendix, Figs. S7and S8). The synthetic WRPWAntp discs bearing the long linkerwere largely overgrown compared with the eye portion of the disc(SI Appendix, Figs. S7 and S8, line 13) and gave rise to an enlargedA3 segment in adult flies (SI Appendix, Fig. S6, line 13), but nopronounced tarsal phenotypes were observed. Staining for Salmhas been performed using both an anti-Salm antibody and a pre-viously described Salm-lacZ enhancer trap chromosome (26). Inboth cases, all lines behaved in the same fashion, an indication thatAntp acts at the transcriptional level to repress Salm.On the other hand, the ectopic activation of grn in the antennal
disc (Fig. 5 andSIAppendix, Fig. S9) followed the sameprinciple asthe ectopic activation of Tsh in the embryo: An absolute re-quirement for a functional YPWM motif (Fig. 5, Lower) was ob-served, but the constructs with the AAAA substitution wereindistinguishable from the control disc bearing a single enhancertrap chromosome (SI Appendix, Fig. S9). Moreover, constructs inwhich WRPW replaced the YPWM motif (SI Appendix, Fig. S9,lines 7, 8, 13 and 14) failed to activate transcription of the reporter,thus confirming our previous findings regarding the strict re-quirement for the YPWM motif in transcriptional activation. Allobserved phenotypes are summarized quantitatively in SI Appen-dix, Table S1.
Full-Length Proteins and Synthetic Antp Peptides Interact with Exdand Bip2 in a YPWM-Dependent Fashion. To analyze the ability offull-length proteins and synthetic Antp peptides to bind DNAand interact with cofactors via the YPWM motif, we performedgel-shift assays using the consensus HD binding site BS2 (SIAppendix, Fig. S10) (30). As expected, all constructs bind to thelabeled oligonucleotide, but those bearing a YPWM motif formadditional complexes in the presence of Exd (31–33) or Bip2 (15,34), which migrate higher than the Antp proteins alone. Eachconstruct is represented by three lanes, the first containingAntp and the second and third also containing Exd or Bip2, re-spectively. The synthetic YPWMAntp peptides also interact withExd and Bip2, albeit more weakly. We also used an ex vivo assayto evaluate the aforementioned interaction quantitatively. Forthis purpose, we performed transactivation experiments in cellculture using a luciferase reporter gene under the control of BS2sites (SI Appendix, Figs. S11 and S12).
DiscussionEarlier studies have shown that the HD plays an essential role inHox gene function. However, ectopic expression of the HD aloneresults in little, if any, homeotic transformation (14, 18). By
combining the HD with the YPWM motif in synthetic geneconstructs, we successfully reconstructed the homeotic functionsof two Hox genes, Scr (29, 35) and Antp (this study). Sub-stitutions of the YPWM motif by WRPW retain the repressiveactivity of Antp, and C-terminal fusions of the WRPW motif tofull-length proteins convert them into stronger repressors. Onthe other hand, the en repression domain—also able to interactwith Gro in vivo (36)—does not confer significant changes toAntp functioning as a repressor or activator. We also evaluatedthe role of the size of the linker between the YPWM motif andthe HD, using naturally occurring splicing variants of Antp withan eight- or a four-amino acid linker. We found that the long-linker variant renders the protein a stronger activator, and theshort-linker makes the protein a stronger repressor of tran-scription in vivo. This result, combined with the finding thatmutation of the linker sequence did not alter its specificity,suggests that Antp specificity depends partially on linker size butnot on sequence. Finally, we observed that synthetic peptidesand full-length proteins can interact with Exd and Bip2, but all
Fig. 4. Repression of genes required for antennal development by ectopicexpression of full-length and synthetic Antp genes. (A) Repression of Salm inthe antennal portion of the eye-antennal disc mediated by full-length andsynthetic Antp genes. All proteins featuring a functional YPWM motif re-press Salm. The constructs with the AAAA substitution do not repress Salm.(B) Repression of dan in the antennal disc by full-length and synthetic Antpgenes. All lines behave as in A.
11962 | www.pnas.org/cgi/doi/10.1073/pnas.1108686108 Papadopoulos et al.

require an intact YPWM motif. The WRPW motif cannot sub-stitute for YPWM in this case.
The Antp YPWM Motif Has a Dual Role in Transcriptional Activationand Repression. The YPWM motif is required for Antp homeoticfunction in the embryo, larva, and adult fly, but the flankingregions of YPWM have no effect on Antp function. The YPWMmotif has a dual role in Antp function, because it is necessaryand indispensable for activation and repression of all Antp targetgenes tested: repression of endogenous Scr, ectopic activation ofTsh, repression of Salm and dan, and ectopic activation of grn.These properties indicate the essential role of the YPWM motifin recruiting both corepressors and coactivators. The YPWMmotif is also required for ectopic induction of mesothoracic legstructures and for interaction with Exd and Bip2 cofactors ingel shift and transactivation assays. However, substitution byWRPW retains to a large extent its repressor activity in bothsynthetic peptides and full-length proteins and results in partialto complete repression of reporter genes.We have shown previously that the YPWM motif acts as an
activation domain in transactivation experiments (15). To ex-amine whether the YPWM motif also can act as a repressordomain, we fused it to the C terminus of the Gal4 protein andtested its binding to UAS and its transactivation efficiency. Weobserved that the Gal4YPWM protein decreased transcription ofthe reporter by about 60%, whereas a quadruple-alanine fusion(Gal4AAAA) did not (SI Appendix, Fig. S13). Therefore, theYPWMmotif, in addition to its role as an activation module, canact as a potent repressor module in vivo.The differential requirements for a functional YPWMmotif in
Antp support the notion that different rules may apply in Hox-mediated transcriptional repression versus activation. Sequencecomparisons among Hox binding preferences to direct targets (9)show that Pbc–Hox sites usually are found in enhancers of genesactivated by a Pbc–Hox complex, whereas regulatory elementsbound by Hox factors alone (without any Pbc input) are dis-tributed equally among the genes activated and repressed by Hoxfactors. This observation supports the idea that Hox-mediatedactivation depends largely on cofactors, whereas repression doesnot always require them.Our phenotypic observations in the embryo, in which both the
YPWM,LLAntpWRPW and the AAAA,LLAntpWRPW proteins acti-vated Tsh ectopically in the region of the embryonic head (Fig. 2B,lines 4 and 5), but the synthetic or full-length AAAAAntp proteinsdid not, suggest that this target gene is activated indirectly in theembryo (possibly through repression of a repressor). However,the finding that the YPWM,LLAntpWRPW construct can ectopicallyactivate grn (Fig. 5, line 4), whereas the AAAA,LLAntpWRPW linecannot (Fig. 5, line 5), indicates that Antp can act as a bona fideactivator and that the activation of grn may be direct.The substitution of the YPWM motif by alanines strongly
decreases but does not entirely abolish Antp homeotic functionin full-length and synthetic AAAAAntp constructs, because theystill exhibit aristal transformations and display ectopic leg bristles
on the A3 segment, suggesting weak transformations toward legidentity. Similar behavior of AAAA-substituted Antp has beenreported in flies for the full-length AAAA,LLAntp, coinduced witha constitutively active Notch (15), or the beetle fushi tarazu geneencoding a protein with a YPWM-to-AAAA substitution (37).Also, the phenotypes of AAAAAntp in the antenna are in line withthe head-involution defects in the larval cuticle early in de-velopment. Although the AAAA-substituted synthetic peptidesfail overall to exhibit significant phenotypes, they still infer mildmalformations in the larval head, including defects in the for-mation of the mouth hooks and, in these cases, lethality. How-ever, unlike their homeotic effect in the antenna, these linesretain T1 identity in the cuticle, as demonstrated by the presenceof prothoracic beards.
The Long and Short Linkers Between the YPWM Motif and the HDEnhance the Activation or Repression Capacity of Antp, Respectively.In the embryo, the antennal disc, and in Drosophila S2 cells thelong-linker Antp proteins behaved as potent activators, whereasthe short-linker constructs acted as strong repressors. The long-linker variant is expressed predominantly in embryonic stages,and equal amounts of both variants are detected in larval, pupal,and adult stages (17). Thus, different Antp transcripts might playdistinct, albeit similar, roles in development. Isoforms with dif-ferent linker sizes also are found in Ultrabithorax (Ubx) (38),and in Abdominal A mutations in the linker decrease its acti-vation capacity on the wingless enhancer (39). Recently, the sizeof the linker between the YPWM and the HD of Ubx was foundto play a regulatory role in Ubx function, with the long- andshort-linker variants not being interchangeable in vivo (40). Inthe same study, the short-linker variant was found to bind DNAmore strongly than its long-linker relative, in the presence ofExd. In line with these arguments, the Antp short-linker variantalso interacted more strongly with Exd and more weakly withBip2 than did its long-linker counterpart (SI Appendix, Fig. S12).
A C-Terminal Portion of Antp Containing the HD and YPWM MotifSuffices for Homeotic Function in Drosophila. One possible N-ter-minal domain to which potency could be attributed is the poly-glutamine (poly-Q) stretch (present in the N terminus of Antpand absent in synthetic Antp peptides), which has been proposedto function as an activator domain in Bicoid (41) and AbdominalB (39). Another conserved motif present in the N terminus ofmany homeoproteins (24) and reported to participate in tran-scriptional potency in the fly Scr (24) and the mouse Hoxa5 (42)is the TSYF motif (SSYF in Scr). An alignment of various Antporthologs showed that the N and C termini of all proteins arelargely variable between insects and mammals, with the latterdisplaying shorter N termini devoid of poly-Q stretches (SI Ap-pendix, Fig. S14). The residues flanking the YPWM motif areslightly conserved in arthropods but not between arthropodsand mammals, an observation that is in line with our findingthat their substitution by alanines does not affect the homeoticfunction of the protein (SI Appendix, Fig. S1). However, thelinker between the HD and the YPWM motif, differentiallyspliced in the transcripts of at least some Hox transcripts (Antp,Ubx), could account for increased specificity of target selectionin vivo. Recent evidence shows that the regions flanking the UbxYPWM motif (alternatively spliced in Ubx isoforms) are re-sponsible for Ubx specificity in vivo (40, 43). Moreover, theYPWM motif and the linker region (termed “the specificitymodule”) of Deformed and Scr result in their unique regulatorybehavior in vivo, and nonconserved domains N-terminal to thisregion allow subtle differences in Hox-DNA recognition prop-erties. These differences can have a considerable impact on thetranscriptional output—activation versus repression—of targetgenes (44). Finally, decisions on whether certain Hox factorsfunction as activators or repressors in a specific context can resultfrom the abundance of their isoforms rather than from theregulatory sequences to which they bind. The finding that thesame enhancer element upstream of the murine Six2 gene isactivated by Hox11 and repressed by Hoxa2 (45) substantiatesthis hypothesis.
Fig. 5. Ectopic activation of leg-specific genes by full-length and syntheticAntp genes in the antennal disc. Ectopic activation of grn in the antennaldisc by synthetic and full-length Antp constructs. Arrows show the ectopicexpression of β-galactosidase upon Antp gain of function in the antennalportion of the disc, as represented schematically. A Dll-Gal4 driver has beenused throughout.
Papadopoulos et al. PNAS | July 19, 2011 | vol. 108 | no. 29 | 11963
GEN
ETICS

The Power of Synthetic Genes. Our study has increased our un-derstanding of synthetic genes. Hypothesizing that protein func-tion is a sum of the functions of its protein domains, the latterbeing oriented in a precise 3D conformation that allows normalinteractions to take place, we may not be not far from attemptingto engineer and possibly predict the functions of novel syntheticpeptides on the basis of the structural architecture of their proteindomains. Inversely, functions of proteins might be scaled down tofractions of their sequences, with each domain retaining onlya subset of these functions. So far, we have reconstructed suc-cessfully, to a great extent, the function of two Hox proteins, Antpand Scr, using their synthetic-peptide versions, and have dem-onstrated that they act predictably in vivo. If biomedically rele-vant proteins also fell in this category, the development and use oftheir corresponding synthetic peptides might present advantagesover their full-length counterparts for application in vivo, becauseof their smaller size and more easily understood molecularproperties. Novel proteins with predictable functions have beenconstructed successfully, e.g., with the neuronal Wiskott–Aldrich
protein, N-WASP (46), and guanine nucleotide exchange factors(47), in which integrating artificial domains that respond tononnative inputs led to the precise control of physiologicalresponses, such as the formation of filopodia or lamellipodia.
Materials and MethodsPlasmids were generated using standard procedures, and fly transgenesis wasperformed as described (48). Preparation of embryos was performed as inref. 9, and imaginal discs were prepared as described (29). Embryonic cuticleswere prepared according to ref. 19. Gel-shift assays were performed asdescribed (49). Detailed methods are outlined in SI Appendix, Materialsand Methods.
ACKNOWLEDGMENTS. This work was supported by the kantons of Basel-Stadt and Basel-Landschaft, a grant from the Swiss National ScienceFoundation, the European Network of Excellence “Cells into Organs,” andthe Consejo Nacional de Ciencia y Tecnología of Mexico, Project 39479-Qand Scholarship 46659. D.K.P. also was supported by a European MolecularBiology Organization short-term fellowship (ASTF 499-2010).
1. Gehring WJ (1987) Homeo boxes in the study of development. Science 236:1245–1252.
2. Lewis EB (1978) A gene complex controlling segmentation in Drosophila. Nature 276:565–570.
3. McGinnis W, Krumlauf R (1992) Homeobox genes and axial patterning. Cell 68:283–302.
4. McGinnis W, Levine MS, Hafen E, Kuroiwa A, Gehring WJ (1984) A conserved DNAsequence in homoeotic genes of the Drosophila Antennapedia and bithoraxcomplexes. Nature 308:428–433.
5. Gehring WJ, Kloter U, Suga H (2009) Evolution of the Hox gene complex from anevolutionary ground state. Curr Top Dev Biol 88:35–61.
6. Kmita-Cunisse M, Loosli F, Bièrne J, Gehring WJ (1998) Homeobox genes in theribbonworm Lineus sanguineus: Evolutionary implications. Proc Natl Acad Sci USA 95:3030–3035.
7. Scott MP, Weiner AJ (1984) Structural relationships among genes that controldevelopment: Sequence homology between the Antennapedia, Ultrabithorax, andfushi tarazu loci of Drosophila. Proc Natl Acad Sci USA 81:4115–4119.
8. Gehring WJ, et al. (1994) Homeodomain-DNA recognition. Cell 78:211–223.9. Mann RS, Lelli KM, Joshi R (2009) Hox specificity unique roles for cofactors and
collaborators. Curr Top Dev Biol 88:63–101.10. Hueber SD, Lohmann I (2008) Shaping segments: Hox gene function in the genomic
age. Bioessays 30:965–979.11. Pearson JC, Lemons D, McGinnis W (2005) Modulating Hox gene functions during
animal body patterning. Nat Rev Genet 6:893–904.12. Furukubo-Tokunaga K, Flister S, Gehring WJ (1993) Functional specificity of the
Antennapedia homeodomain. Proc Natl Acad Sci USA 90:6360–6364.13. Qian YQ, Resendez-Perez D, Gehring WJ, Wüthrich K (1994) The des(1-6)
antennapedia homeodomain: Comparison of the NMR solution structure and theDNA-binding affinity with the intact Antennapedia homeodomain. Proc Natl Acad SciUSA 91:4091–4095.
14. Gibson G, Schier A, LeMotte P, Gehring WJ (1990) The specificities of Sex combsreduced and Antennapedia are defined by a distinct portion of each protein thatincludes the homeodomain. Cell 62:1087–1103.
15. Prince F, et al. (2008) The YPWMmotif links Antennapedia to the basal transcriptionalmachinery. Development 135:1669–1679.
16. Paroush Z, et al. (1994) Groucho is required for Drosophila neurogenesis,segmentation, and sex determination and interacts directly with hairy-related bHLHproteins. Cell 79:805–815.
17. Stroeher VL, Gaiser JC, Garber RL (1988) Alternative RNA splicing that is spatiallyregulatedGgeneration of transcripts from the Antennapedia gene of Drosophilamelanogaster with different protein-coding regions. Mol Cell Biol 8:4143–4154.
18. Bello B, Resendez-Perez D, Gehring WJ (1998) Spatial and temporal targeting of geneexpression in Drosophila by means of a tetracycline-dependent transactivator system.Development 125:2193–2202.
19. Gibson G, Gehring WJ (1988) Head and thoracic transformations caused by ectopicexpression of Antennapedia during Drosophila development. Development 102:657–675.
20. Zeng W, Andrew DJ, Mathies LD, Horner MA, Scott MP (1993) Ectopic expression andfunction of the Antp and Scr homeotic genes: The N terminus of the homeodomain iscritical to functional specificity. Development 118:339–352.
21. Andrew DJ, Horner MA, Petitt MG, Smolik SM, Scott MP (1994) Setting limits onhomeotic gene function: Restraint of Sex combs reduced activity by teashirt and otherhomeotic genes. EMBO J 13:1132–1144.
22. McCormick A, Coré N, Kerridge S, Scott MP (1995) Homeotic response elements aretightly linked to tissue-specific elements in a transcriptional enhancer of the teashirtgene. Development 121:2799–2812.
23. Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cellfates and generating dominant phenotypes. Development 118:401–415.
24. Tour E, Hittinger CT, McGinnis W (2005) Evolutionarily conserved domains requiredfor activation and repression functions of the Drosophila Hox protein Ultrabithorax.Development 132:5271–5281.
25. Struhl G (1981) A homoeotic mutation transforming leg to antenna in Drosophila.Nature 292:635–638.
26. Wagner-Bernholz JT, Wilson C, Gibson G, Schuh R, Gehring WJ (1991) Identification oftarget genes of the homeotic gene Antennapedia by enhancer detection. Genes Dev5(12B):2467–2480.
27. Emerald BS, Curtiss J, Mlodzik M, Cohen SM (2003) Distal antenna and distal antennarelated encode nuclear proteins containing pipsqueak motifs involved in antennadevelopment in Drosophila. Development 130:1171–1180.
28. Grieder NC, Morata G, Affolter M, Gehring WJ (2009) Spalt major controls thedevelopment of the notum and of wing hinge primordia of the Drosophilamelanogaster wing imaginal disc. Dev Biol 329:315–326.
29. Papadopoulos DK, et al. (2010) Function and specificity of synthetic Hox transcriptionfactors in vivo. Proc Natl Acad Sci USA 107:4087–4092.
30. Furukubo-Tokunaga K, et al. (1992) In vivo analysis of the helix-turn-helix motif of thefushi tarazu homeo domain of Drosophila melanogaster. Genes Dev 6:1082–1096.
31. Passner JM, Ryoo HD, Shen L, Mann RS, Aggarwal AK (1999) Structure of a DNA-bound Ultrabithorax-Extradenticle homeodomain complex. Nature 397:714–719.
32. Piper DE, Batchelor AH, Chang CP, Cleary ML, Wolberger C (1999) Structure ofa HoxB1-Pbx1 heterodimer bound to DNA: Role of the hexapeptide and a fourthhomeodomain helix in complex formation. Cell 96:587–597.
33. Ryoo HD, Mann RS (1999) The control of trunk Hox specificity and activity byExtradenticle. Genes Dev 13:1704–1716.
34. Gangloff YG, et al. (2001) The TFIID components human TAF(II)140 and DrosophilaBIP2 (TAF(II)155) are novel metazoan homologues of yeast TAF(II)47 containinga histone fold and a PHD finger. Mol Cell Biol 21:5109–5121.
35. Vukojevic V, Papadopoulos DK, Terenius L, Gehring WJ, Rigler R (2010) Quantitativestudy of synthetic Hox transcription factor-DNA interactions in live cells. Proc NatlAcad Sci USA 107:4093–4098.
36. Tolkunova EN, Fujioka M, Kobayashi M, Deka D, Jaynes JB (1998) Two distinct types ofrepression domain in engrailed: One interacts with the groucho corepressor and ispreferentially active on integrated target genes. Mol Cell Biol 18:2804–2814.
37. Löhr U, Pick L (2005) Cofactor-interaction motifs and the cooption of a homeotic Hoxprotein into the segmentation pathway of Drosophila melanogaster. Curr Biol 15:643–649.
38. O’Connor MB, Binari R, Perkins LA, Bender W (1988) Alternative RNA products fromthe Ultrabithorax domain of the bithorax complex. EMBO J 7:435–445.
39. Merabet S, et al. (2003) The hexapeptide and linker regions of the AbdA Hox proteinregulate its activating and repressive functions. Dev Cell 4:761–768.
40. Reed HC, et al. (2010) Alternative splicing modulates Ubx protein function inDrosophila melanogaster. Genetics 184:745–758.
41. Janody F, Sturny R, Schaeffer V, Azou Y, Dostatni N (2001) Two distinct domains ofBicoid mediate its transcriptional downregulation by the Torso pathway.Development 128:2281–2290.
42. Zhao JJ, Lazzarini RA, Pick L (1996) Functional dissection of the mouse Hox-a5 gene.EMBO J 15:1313–1322.
43. Liu Y, Matthews KS, Bondos SE (2009) Internal regulatory interactions determine DNAbinding specificity by a Hox transcription factor. J Mol Biol 390:760–774.
44. Joshi R, Sun L, Mann R (2010) Dissecting the functional specificities of two Hoxproteins. Genes Dev 24:1533–1545.
45. Yallowitz AR, Gong KQ, Swinehart IT, Nelson LT, Wellik DM (2009) Non-homeodomain regions of Hox proteins mediate activation versus repression of Six2via a single enhancer site in vivo. Dev Biol 335:156–165.
46. Dueber JE, Yeh BJ, Chak K, Lim WA (2003) Reprogramming control of an allostericsignaling switch through modular recombination. Science 301:1904–1908.
47. Yeh BJ, Rutigliano RJ, Deb A, Bar-Sagi D, Lim WA (2007) Rewiring cellular mor-phology pathways with synthetic guanine nucleotide exchange factors. Nature 447:596–600.
48. Spradling AC, Rubin GM (1982) Transposition of cloned P elements into Drosophilagerm line chromosomes. Science 218:341–347.
49. Plaza S, et al. (2001) Molecular basis for the inhibition of Drosophila eye developmentby Antennapedia. EMBO J 20:802–811.
11964 | www.pnas.org/cgi/doi/10.1073/pnas.1108686108 Papadopoulos et al.





















![GENES VII [Books biology genetics] [prentice.hall][genes](https://img.pdfslide.us/doc/110x75/55cce1ffbb61eb56508b46c3/genes-vii-books-biology-genetics-prenticehallgenes.jpg)







