Embed Size (px)
Citation preview

BritishJournal ofOphthalmology 1992; 76: 486-490
Fibronectin synthesis in subretinal membranes ofproliferative vitreoretinopathy
P Hiscott, H A Waller, I Grierson, M G Butler, D L Scott, Z Gregor, I Morino
AbstractIn situ hybridisation and immunohistochemi-cal studies were conducted on six surgicallyexcised subretinal membranes of proliferativevitreoretinopathy to investigate whether dis-placement of retinal pigment epithelial andglial cells to subretinal membranes was
associated with fibronectin production by thesubretinal membrane cells. Fibronectinmessenger RNA (mRNA) and fibronectinimmunoreactivity were observed in some cellsin all of the subretinal membranes studied andup to 30% of the cells in individual specimensshowed intense labelling for fibronectinmRNA. The results support the concept thatthe cells in subretinal membranes producefibronectin. Locally produced fibronectin mayplay a role in subretinal membrane cohesion,and displacement of retinal pigment epithelialand glial cells from their normal location mayinduce the cells to manufacture fibronectin.Fibronectin production may be more promi-nent in migrating subretinal cells.(BrJ Ophthalmol 1992; 76: 486-490)
Institute ofOphthalmology,Department of PathologyP Hiscott
Department of ClinicalScienceP HiscottI Grierson
Departments ofImmunology andRheumatology, StBartholomew's HospitalH A WallerM G ButlerD L Scott
The Vitreoretinal Unit,Moorfields Eye HospitalP HiscottZ Gregor
Department ofOphthalmology, KobeUniversity, JapanI MorinoCorrespondence to:Dr P Hiscott, Institute ofOphthalmology, 17-25 CaytonStreet, London EC1V 9AT.Accepted for publication10 February 1992
Proliferative vitreoretinopathy (PVR) is a com-
plication of retinal detachment and is charac-terised by the development of contractile, scar-
like fibrocellular membranes on either side of thedetached neuroretina (epiretinal and subretinalmembranes).' The contractile epiretinal and sub-retinal membranes ofPVR have a similar cellularcomposition,2" although contractile subretinalmembranes appear to have a larger retinal pig-ment epithelial component and smaller glialcomponent than epiretinal membranes,356 andthe extracellular matrix of PVR membranes atboth sites contains collagens and glycoproteins,including fibronectin.7"' Fibronectin has a num-
ber of important biological functions such as arole in cell-cell and cell-substratum adhesion,""'3and the adhesive properties of fibronectin maycontribute to the cell-cell and cell-extracellularmatrix cohesion in early PVR membranes.
In PVR, epiretinal glial and retinal pigmentepithelial cells are replete with fibronectinmessenger RNA (mRNA) whereas cells in thesubjacent detached neuroretina and retinal pig-ment epithelial monolayer exhibit little or no
fibronectin message, suggesting that the ectopicretinal pigment epithelial and glial cells produceabundant glycoprotein while those in situ in theretina do not. 4 The 'up regulation' of fibronectinproduction by the displaced cells probably con-
tributes to the glycoprotein in epiretinalmembranes but the mechanism for the change ingene expression for fibronectin is obscure.One possible explanation for the activation of
fibronectin production in the ectopic cells is that
displacement of the retinal pigment epithelialand glial cells from their normal location stimu-lates, or removes inhibitions from, the cells toproduce fibronectin, in which case fibronectinshould be produced by the cells supplanted tosites other than the vitreoretinal junction. Onesuch location is within the contractile subretinalmembranes of PVR where retinal pigmentepithelial and glial cells are displaced into thedeveloping fibrocellular tissue.2-6 Therefore, toinvestigate further the hypothesis that displace-ment of pigment epithelial and glial retinal cellscauses them to produce fibronectin, we examinedthe cells in PVR subretinal membranes for thepresence of fibronectin mRNA employing an insitu hybridisation technique. The results werecorrelated with the distribution of fibronectinimmunoreactivity in the specimens.
Materials and methods
CONTROL TISSUEFormalin fixed, paraffin embedded synovial tis-sues, from patients with rheumatoid arthritisknown to contain abundant fibronectin mRNAand show strong fibronectin immunoreactivity,"were used as positive controls for the hybridisa-tion experiments and immunohistochemical pro-cedures.
SUBRETINAL MEMBRANESSix subretinal membranes removed during parsplana vitrectomy forPVR were used in the study.
PREPARATION OF SINGLE-STRANDED RNA PROBESA cDNA clone (pFHl) representing about onequarter of the estimated size of human cellularfibronectin mRNA6 17 was provided by Dr F EBaralle, Sir William Dunn School of Pathology,University of Oxford and subcloned into pGEMblue (Promega, WI, USA) to produce single-stranded RNA anti-sense and sense (control)probes, as previously described.'4 15
Transcription was conducted incorporatingdigoxigenin-l1-UTP (Boehringer Mannheim,GmbH) with the standard transcription reactionusing SP6 or T7 RNA polymerase (Boehringer).To achieve optimal probe length for tissue pene-tration alkaline hydrolysis was carried out to givean average probe length of 100 bases.'8 Toprevent digoxigenin loss into the organic phasephenol/chloroform extraction was omittedduring probe purification. 9 To ensure the correctsize of transcripts and the success of hydrolysisthe probes were checked by 2% denaturingagarose gel electrophoresis and northern blot-ting.20 Nitrocellulose filters were blocked with
486
on 9 June 2018 by guest. Protected by copyright.
http://bjo.bmj.com
/B
r J Ophthalm
ol: first published as 10.1136/bjo.76.8.486 on 1 August 1992. D
ownloaded from
Fibronectin synthesis in subretinal membranes ofproliferative vitreoretinopathy
3% normal sheep serum in phosphate bufferedsaline (PBS; pH 7-4) and subsequent immuno-chemical detection of probes was carried outusing sheep anti-digoxigenin alkaline phospha-tase conjugate (Boehringer) at 1:5000 in 3%normal sheep serum/PBS. Development waswith NBT/BCIP (Sigma, Poole, UK) in TRISbuffered saline, pH 9 5 (0-3 mg/ml NBT; 0-2mg/ml BCIP; 0-1 M TRIS; 0-1 M sodiumchloride; 0-005 M magnesium chloride). Probeswere stored at -20°C until used.
HYBRIDISATION HISTOCHEMISTRYSubretinal membranes were fixed for between 4and 48 h in 10% formol-saline, dehydrated ingraded concentrations of ethanol and embeddedin paraffin wax.
Sections (6 ,um thick) ofwax-embedded tissuewere cut, dewaxed, permeabilised, treated withproteinase K, post-fixed, dehydrated inmethanol and air-dried.'4 The anti-sense (SP6,fibronectin transcript) probe was then appliedat a concentration of 1:100 (approximately0-1 ,ug/ml) in hybridisation medium and thepreparations were hybridised overnight undercoverslips at 37°C in a humid chamber. Duringall the procedures up to and including hybridisa-tion, RNAse free conditions were employed.20The coverslips and unbound probe wereremoved as previously described. 14
Sites of probe binding were visualisedimmunohistochemically using the antibody todigoxigenin. Non-specific antibody binding wasblocked by a 20 minute incubation with 3%normal sheep serum/PBS, sheep anti-digoxigenin antibody-alkaline phosphatase con-jugate was applied to the sections for 30 minutesat a concentration of 1:200 in 3% normal sheepserum/PBS. The sections were washed in PBSand the reaction sites visualised blue/black incolour using NBT/BCIP (as described above)overnight. The preparations were washed,counterstained with haematoxylin and mountedin glycerol.
Procedural controls included (1) pre-treatmentof sections with 20 ,ug/ml RNAse in TRIS/EDTA buffer (0-5 M sodium chloride, 10 mMTRIS hydrochloride pH 8-0, 1 mM EDTA) for30 minutes at 37°C to remove all tissue RNA, and(2) incubation of sections with the sense (T7)probe at a concentration of 1:100 instead of theanti-sense probe.
FIBRONECTIN IMMUNOHISTOCHEMISTRYSerial sections (6 ,um thick) to those used for insitu hybridisation were immunostained forfibronectin with an antiserum raised in rabbitto human plasma fibronectin (Dako) or a mono-clonal antibody from Sigma (which gave similarstaining patterns). The immunostaining was con-ducted as previously described7 and the immu-noperoxidase technique was used throughoutthis investigation, employing 3-amino-9-ethylcarbazole as chromogen. Some sectionswere counterstained with Mayer's haematoxylin.Other sections were stained with haematoxylinand eosin. Controls were applied to the im-munostaining procedures as previously des-
cribed.7 In addition, where sufficient tissue wasavailable in the specimens the retinal origin ofthesubretinal cells was confirmed by immunotypingtheir intermediate filaments (using glial fibrillaryacidic protein to detect glial cells and cytokeratinto demonstrate retinal pigment epithelialcells).2' 22The hybridisation and immunohistochemical
preparations were examined and compared bybright field and differential interference contrastmicroscopy. Semi-quantitative estimates ofhybridisation and immunohistochemical labell-ing were made by comparing the 'test' sectionswith controls. Labelling equivalent to that seenin negative controls (for example, sense probetreated sections or immunohistochemical nega-tive control sections) was termed 'backgroundlabelling', while positive labelling at a levelequivalent to the strongest seen in positiverheumatoid synovium was designated 'intense
op* *<- s f +! b
4M' .
wf;::...i?v-w- X~:~~~ 0,$' +<_
r% v .4
C
'-~~~~~~~~~.*~ 4
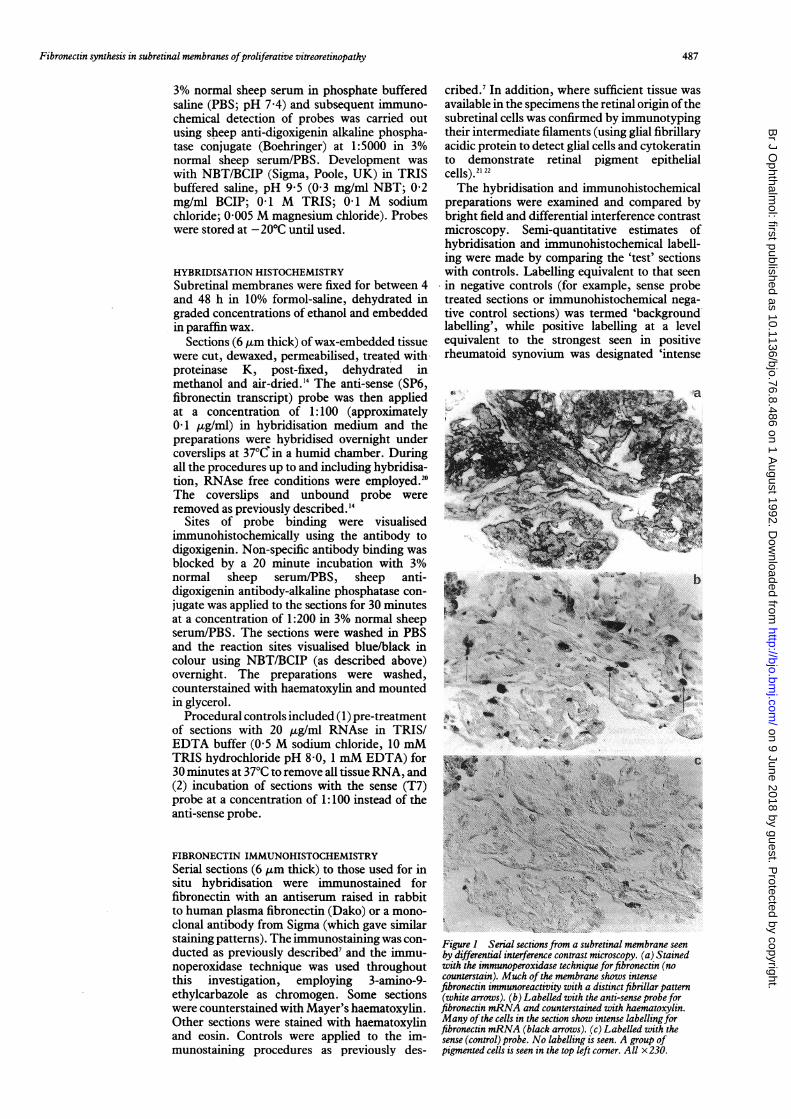
Figure 1 Serial sections from a subretinal membrane seenby differential interference contrast microscopy. (a) Stainedwith the immunoperoxidase technique for fibronectin (nocounterstain). Much ofthe membrane shows intensefibronectin immunoreactivity with a distinct fibrillar pattern(white arrows). (b) Labelled with the anti-sense probeforfibronectin mRNA and counterstained with haematoxylin.Many ofthe cells in the section show intense labellingforfibronectin mRNA (black arrows). (c) Labelled with thesense (control) probe. No labelling is seen. A group ofpigmented cells is seen in the top left corner. All x230.
487
on 9 June 2018 by guest. Protected by copyright.
http://bjo.bmj.com
/B
r J Ophthalm
ol: first published as 10.1136/bjo.76.8.486 on 1 August 1992. D
ownloaded from

Hiscott, Waller, Grierson, Butler, Scott, Gregor, Morino
labelling', Labelling between background aintense levels was called 'moderate labelling'.
Results
CONTROLSSections of rheumatoid synovium hybridiPwith the anti-sense probe showed moderateintense labelling in the synovial lining ccwhereas sections hybridised with the sense prcor pretreated with RNAse exhibited only ba4ground labelling. The synovium showed mo(rate or intense immunoreactivity for fibronecwhile no immunolabelling occurred in the neltive procedural controls. Subretinal membrasections labelled with the sense probe showonly background labelling levels (Fig 1).
.,~~~~~~~~~~~~~~~. ..... *
.~ ~ ~~~~~~w
Figure 2 Differentialinterference contrastmicrographs ofsectionsfroma subretinal membrane. (a)Stained by theimmunoperoxidase technique(no counterstain) for glialfibrillary acidic proteinreveals a small glialcomponent in the membrane(open arrows). Pigment ispresent in the tissue(asterisk). (b) Stained by theimmunoperoxidase technique(no counterstain) forcytokeratins showsprominent retinal pigmentepithelial layers (arrows).(c) Labelled with the anti-sense probe forfibronectinmRNA and counterstainedwith haematoxylin. Manyof the pigment epithelialcells show moderate orintense labelling forfibronectin message(arrows). (d) Labelled withthe sence (control) probe andcounterstained withhaemotoxylin. No labellingis seen. All x290.
nd
sedor
ells)beck-
Table I Fibronectin mRNA in the cells ofPVR subretinalmembranes
Proportion ofcells Proportion ofcellsshozving moderate showing intense labelling
Subretainal labellingforfibronectin forfibronectin mRNAmembrane no mRNA (%) (%)
1 20 02 20 303 10 104 50 05 30 206 20 10
ae; CELLS IN SUBRETINAL MEMBRANEStin All six subretinal membranes contained cellsga- which labelled with the anti-sense probe (Figs 1mne and 2). There was some variation between theved membrane specimens in the numbers of cells
which contained fibronectin mRNA. Between20% to 50% of cells showed levels of labelling
a above background with the anti-sense probe(Table 1). In f6ur membranes between 10% to30% of the total cells in the specimen showedintense labelling with the probe, the remainingpositive cells labelling at moderate levels. Label-ling was seen in both isolated cells and cellsaggregated in layers in the subretinal tissue,including cells in layers at apparent specimensurfaces (Fig 2). Moderate to strong fibronectinimmunoreactivity was also observed in all thesubretinal membranes (Fig 1). Enough tissuewas present in the specimens to confirm thatsome of the cells which contained fibronectin
b mRNA were retinal pigment epithelial (fourmembranes; Fig 2) or glial (one membrane) inorigin. It was not possible to detect a quantitativedifference between cell types in the intensity oflabelling for fibronectin mRNA.
Discussione Our results confirm that fibronectin contributes
to the extracellular matrix in the contractilesubretinal membranes ofPVR, demonstrate that
5ij; many subretinal cells contain mRNA for thec glycoprotein, and affirm that cells in subretinal
membranes produce fibronectin. Moreover, theresults are in keeping with the observation thatfibronectin is synthesised by retinal pigment
in 232epithelium in vitro, since retinal pigmentepithelial cells are a major component of sub-
> retinal membranes.'4s The production- of fibronectin by cells in
subretinal membranes represents a markedbehavioural change from the activities seen in thecells in situ in attached and even detached adult
K human retina, since retinal pigment epithelialand glial cells in situ lack appreciable fibronectin
d immunoreactivity and mRNA.'4 This 'up regula-tion' of fibronectin production by retinalpigment epithelial and glial cells in subretinalmembranes may be caused by one of severalmechanisms. One possibility is that inflamma-
4 tory mediators within the vitreous and subretinalfluids of eyes developing PVR subretinalmembranes, or factors derived from inflamma-tory cells in evolving membranes, may stimulatefibronectin production. It has been shown thathepatocytes manufacture fibronectin in the pres-ence of interleukin-6.2' However, the apparentabsence of fibronectin production in the adjacent
488
on 9 June 2018 by guest. Protected by copyright.
http://bjo.bmj.com
/B
r J Ophthalm
ol: first published as 10.1136/bjo.76.8.486 on 1 August 1992. D
ownloaded from

Fibronectin synthesis in subretinal membranes ofproliferative vitreoretinopathy
neuroretina and retinal pigment epithelial mono-layer is counter to this theory. Alternatively, theobservation of fibronectin synthesis in the sub-retinal, as well as the epiretinal,'4 component ofPVR is in keeping with the theory that it is thedisplacement of retinal pigment epithelial andglial cells from their normal location whichinduces, or removes constraints from, the cells toproduce fibronectin.Some of the fibronectin in subretinal
membranes may also be derived from plasmafibronectin, since fibronectin enters the vitreousand subretinal space following breakdown of theblood retina barrier in retinal detachment andPVR.26-28 The developing subretinal 'scars' maythen incorporate the plasma-derived fibronectin.Indeed, moderate to strong fibronectin immu-noreactivity was observed in all the subretinalmembranes studied irrespective of the propor-tion ofcells expressing fibronectin mRNA. How-ever, much of the fibronectin content of themembranes is likely to emanate from the subreti-nal cells themselves since cell surface fibronectinis thought to be insoluble'3 and therefore is likelyto accumulate in the tissue. Thus both plasmaand locally-produced fibronectin probably con-tribute to the tissue.The accumulation of both plasma and locally-
produced fibronectin has been described in repa-rative processes elsewhere in the body'3 29 and inthis respect subretinal membrane formation maybe likened to an evolving scar. However, in otherrespects subretinal membranes differ from heal-ing wounds - for example, the absence of aprominent vascular component,6 and it remainsto be seen whether the fibronectin is as importantin subretinal membrane formation as it is woundhealing in general.The production of fibronectin by subretinal
membrane cells does not imply that fibronectin isimportant in the pathobiology of the tissueper se. However, the abundance of message insome subretinal membranes and the recentdemonstration of fibronectin receptors on cellsdisplaced into PVR epiretinal membranes' is inkeeping with the concept that the glycoproteinplays a role in subretinal membrane develop-ment.
In early PVR, plasma-derived fibronectin islikely to be involved in chemotactic recruitmentof cells into the developing subretinal tissue. Asthe subretinal membrane cells themselves beginto produce fibronectin, the insoluble cell surfaceglycoprotein may contribute to the cohesion ofthe tissue and potentiate the chemotacticstimulus in the membrane. Indeed, work in ourlaboratory suggests that retinal pigment epithe-lial cells may be migrating into already wellestablished subretinal membranes.3' 32Migrating cells may not only be attracted into
developing subretinal membranes by fibronectinbut could specifically be producing the glycopro-tein since migratory cells in other pathologicaltissues can produce fibronectin.33 Interestingly,fibronectinmRNA was particularly noted in cellsat apparent surfaces ofthe subretinal membraneswhere migratory cells may be expected. Ifmigrating cells produce fibronectin in evolvingmembranes, it is possible that a single agentcould prevent both recruitment of cells into and
cohesion within developing membranes and pro-vide a new avenue of therapeutic intervention inthe process.
This work was funded by the Weilcome Trust (grant no17095/1.4R) and the Joint Research Board (St Bartholomew'sHospital). Mr R Howes provided photographic help. Dr Scott isMuir Hambro Fellow of the Royal College of Physicians.
1 The Retina Society Terminology Committee. The classifica-tion of retinal detachment with proliferative vitreo-retinopathy. Ophthalmology 1983; 90: 121-5.
2 Daicker B. Constricting retroretinal membranes associatedwith traumatic retinal detachments. Graefes Arch Clin ExpOphthalmol 1985; 222: 147-53.
3 Shirakawa H, Yoshimura N, Yamakawa R, Matasumura M,Okada M, Ogina N. Cell components in proliferativevitreoretinopathy: immunofluorescent double staining ofcultured cells from proliferative tissues. Ophthalmologica1987; 194: 56-62.
4 Schwartz D, de la Cruz ZC, Green WR, Michels RG.Proliferative vitreoretinopathy. Ultrastructural study of 20retroretinal membranes removed by vitreous surgery. Retina1988; 8:275-81.
5 Trese MT, Chandler DB, Machemer R. Subretinal strands:ultrastructural features. Graefes Arch Clin Exp Ophthalmol1985; 223: 35-40.
6 Hiscott P, Morino I, Alexander R, Grierson I, Gregor Z.Cellular components of subretinal membranes in prolifera-tive vitreoretinopathy. Eye 1989; 3: 606-10.
7 Hiscott PS, Grierson I, McLeod D. Natural history offibrocellular epiretinal membranes: a quantitative, auto-radiographic and immunohistochemical study. Br JOphthalmol 1984; 69: 810-23.
8 Scheiffarth OF, Kampik A, Guenther H, vd Mark K. Proteinsof the extracellular matrix in vitreoretinal membranes.GraefesArch ClinExpOphthalmol 1988: 226: 357-61.
9 Morino I, Hiscott P, McKechnie N, Grierson I. Variation inepiretinal membrane components with clinical duration ofthe proliferative tissue. BrJI Ophthalmol 1990; 74: 393-9.
10 Okada M, Matsumura M, Hosoda Y, Ogino N, Honda Y.Immunohistochemical identification of the cellular andextracellular components of subretinal proliferative tissues.Invest Ophthalmol VisSci 1991; 32(suppl): 857.
11 Yamada KM, Yamada SS, Pastan I. The major cell surfaceglycoprotein of chick embryo fibroblasts is an agglutinin.Proc Natl Acad Sci USA 1975; 72: 3158-62.
12 Pearlstein E, Gold LI, Garcio-Pardo A. Fibronectin: a reviewof its structure and biological activity. Mol Cell Biochem1980; 29: 103-28.
13 d'Ardenne AJ, McGee JO'D. Fibronectin in disease. J Pathol1984; 142:235-51.
14 Hiscott P, Waller HA, Butler MG, Grierson I, Scott D. Localproduction offibronectin by ectopic human retinal'cells. CellTissue Res 1992; 267: 185-92.
15 Waller HA, ButlerMG, McCleanJGB, Dowd GSE, Scott DL.The localisation of fibronectin mRNA in the rheumatoidsynovium by in situ hybridisation. BrJ Rheumatol 1991;29(suppl): 36.
16 Kornblihtt AR, Vibe-Pedersen K, Baralle FE. Isolation andcharacterization of cNDA clones for human and bovinefibronectins. Proc Natl Acad Sci USA 1983; 80: 3218-22.
17 Kornblihtt AR, Vibe-Pedersen K, Baralle FE. Humanfibronectin: molecular cloning evidence for two mRNAspecies differing by an internal segment coding for astructural domain. EMBOJ 1984; 3: 221-6.
18 Angerer LM, Stoler MH, Angerer RL. In situ hybridisationwith RNA probes: an annotated recipe. In: Valentino KL,Eberwise JH, Barchas JD, eds. In situ hybridisation applica-tions to neurobiology. Oxford: Oxford University Press, 1987,42-70.
19 Schmitz GG, Walter T, Seibl R, Kessler C. Non-radioactivelabelling of oligonucleotides in vitro with the haptendigoxigenin by tailing with terminal transferase. Anal Bio-chem 1991; 192: 222-31.
20 Sambrook J, Fritsch EF, Maniatis T. Molecular cloning - alaboratory manual, 2nd edn. New York: Cold Spring HarborLaboratory Press, 1989.
21 Hiscott PS, Grierson I, Trombetta CJ, Rahi AHS, Marshall J,McLeod D. Retinal and epiretinal glia - an immuno-histochemical study. Br J Ophthalmol 1984; 68: 698-707.
22 Hiscott PS, Grierson I, McLeod D. Retinal pigment epithelialcells in epiretinal membranes: an immunohistochemicalstudy. BrJ Ophthalmol 1984; 68: 708-15.
23 Campochiaro PA, Jerdan JA, Glaser BM. The extracellularmatrix of human retinal pigment epithelial cells in vivo andits synthesis in vitro. Invest Ophthalmol Vis Sci 1986; 27:1615-21.
24 Newsome DA, Pfeffer BA, Hewitt AT, Robey PG,Hassell JR.Detection of extracellular matrix molecules synthesized invitro by monkey and human retinal pigment epithelium:influence of donor age and multiple passages. Exp Eye Res1988; 46: 305-21.
25 Hagiwara T, Suzuki H, Kono I, Kashiwagi H, Akiyama Y,Onozaki K. Regulation of fibronectin synthesis by inter-leukin-1and interleukin-6 in rat hepatocytes. AmJI Pathol1990; 133: 39-47.
26 Campochiaro PA, Jerdan JA, Glaser BM. Serum containschemoattractants for human retinal pigment epithelial cells.Arch Ophthalmol 1984; 102: 1830-3.
27 Campochiaro PA, Jerdan JA, Glaser BM. Vitreous aspiratesfrom patients with proliferative vitreoretinopathy stimulate
489
on 9 June 2018 by guest. Protected by copyright.
http://bjo.bmj.com
/B
r J Ophthalm
ol: first published as 10.1136/bjo.76.8.486 on 1 August 1992. D
ownloaded from

Hiscott, Waller, Grierson, Butler, Scott, Gregor, Morino
retinal pigment epithelial cell migration. Arch Ophthalmol1885; 103: 403-5.
28 Campochiaro PA, Sen HA, Robertson TJ, Conway BP. Therole of the breakdown of the blood-retinal barrier inproliferative vitreoretinopathy. In: Heimann K, Wied-emann P, eds. Proliferative vitreoretinopathy. Heidelberg:Kaden, 1989, 45-9.
29 Casscells W, Kimura H, Sanchez JA, Yu Z, Ferraris VJ.Immunohistochemical study of fibronectin in experimentalmyocardial infarction. AmJ Pathol 1990;-137: 801-10.
30 Weller M, Wiedemann P, Bresgen M, Heimann K. Vitronec-tin and proliferative intraocular disorders. II. Expression ofcell surface receptors for fibronectin and vitronectin in
periretinal membranes. Int Ophthalmol 1991; 15: 103-8.31 Robey HL, Hiscott P, Grierson I. Cytoskeletal reorganisation
in mobilised migratory human retinal pigment epithelialcells in vitro. Invest Ophthalmol Vis Sci 1991; 32(suppl): 767.
32 Hiscott P, Morino I, McKechnie N, Grierson I, Gregor Z.Proliferating retinal pigment epithelial cells in subretinalmembranes. Proc XXVI Internat Cong Ophthalmol. Amster-dam: Elsevier, 1991.
33 Adachi K, Yamauchi K, BernaudinJ-F, Fouret P, Ferrans VJ,Crystal RG. Evaluation of fibronectin gene expression by insitu hybridisation: differential expression of the fibronectingene among populations of human alveolar macrophages.AmJ7 Pathol 1988; 133: 193-203.
490
on 9 June 2018 by guest. Protected by copyright.
http://bjo.bmj.com
/B
r J Ophthalm
ol: first published as 10.1136/bjo.76.8.486 on 1 August 1992. D
ownloaded from





![[Shinobi] Bleach 486](https://img.pdfslide.us/doc/110x75/568bd5611a28ab2034983901/shinobi-bleach-486.jpg)














![Second Thoughts[486]](https://img.pdfslide.us/doc/110x75/577d35651a28ab3a6b9052b5/second-thoughts486.jpg)







