Embed Size (px)
Citation preview

Extant Families Super family: Canoidea are Dog-like
• Canidae - (dogs, wolves, jackels, foxes) • Ursidae - (bears, giant panda) • Procyonidae - (raccoons, coatimundi) • Ailuridae - (red panda) • Mustelidae - (otter, weasels and badgers) • Mephitidae (skunks) note: formerly of Mustelidae Family • Pinnipeds – considered a clade with the following families:
i. Phocidae (earless or true seals) ii. Otariidae (eared seals - fur seals & sealions) iii. Odobenidae (walrus)
Super Family: Feloidea are cat-like 1. Felidae - (cats) 2. Viverridea – (civets & genets) 3. Herpestidae - (meerkats, mongooses, fossa) 4. Hyaenidae - (hyenas)
The Order Carnivora is distributed nearly world-wide. Antarctica remains to extreme to support this group while others may be found in, but are not native to the Australasian region. Bears are found in the New World and Eurasia but not in Africa where as the Hyenas are found only in Africa. Procyonids are restricted to the New World while Viverrids and Herpistids are found throughout the Old World. Canids and Felids have managed to become more wide spread than all other Carnivora families. The Pinnipeds appear to have arisen from Mustelids and Canids, they have been considered to be in a separate Order due to their specialized adaptations to aquatic life. See Pinniped Study Guide in the docent notebook. Carnivorous refers to an animal (or plant) that primarily eats meat, which can be vertebrates (fish, birds, mammals, etc.) or invertebrates (crustaceans, mollusks, etc.) but not insects (insectivorous). While most of the species we classify as carnivorous are within the Order Carnivora, there are some species outside of the order that have also evolved to eat primarily meat. These include the Cetacea (whales and dolphins), some Marsupialia (Tasmanian wolf and devil et al) and Artyodactyla (peccaries and swine etc). Carnivora translates to meat-eater, but it refers to the order or an animal within the taxonomic order (species may or may not actually eat meat). Most Carnivora do eat meat, but many are omnivorous (have a diet that consists of both meat and plant material), and some few species have switched back through the course of their evolution to specialize in eating plant material (secondary herbivorous). Carnivore refers to any carnivorous animal, but especially to members of the order Carnivora. For simplicity, within this study guide, the word carnivore is generally used to refer to a member of the order. EVOLUTION

Mammals as a group have been around for a very long time first appearing during the Permian some 250 mya. These were the so-called mammal-like reptiles now referred to as synapsids. Carnivores, herbivores and insectivores were all present but only a small handful made their way through the Permian extinction event. All were small during the Mesozoic period (245 - 65 mya. Today’s carnivores evolved from small, forest dwelling insectivores that lived during the Paleocene Epoch (64 -54 mya). These early hunters probably looked a lot like the modern Viverrids, forest dwellers that fed on rodents, reptiles and birds. At that time, the largest land predators were giant flightless birds (e.g. the 2 m Diatryma). During the Eocene (54 - 34 mya) at least two groups shared the niche of mammalian predators. The archaic dog - jaguar sized creodonts (60 - 30 mya) were the first large carnivore to fill the terrestrial hunting niche. While the smaller weasel-like Miacids were confined to hunting in the woodlands. Yet they were the first to have “true” carnassial teeth whereas the creodonts were less refined. Though the creodonts were specialized to meat-eating alone, the Miacids retained grinding molars, allowing them to continue to exploit other feeding opportunities such as seeds, fruits, and insects. By mid Eocene, herbivores had evolved speedier and brainier species, effectively dooming the Creodonts who not keep pace. The Miacids however, had evolved proportionally larger brains, and it is probable that a combination of these traits (especially intelligence and opportunistic feeding) allowed them to survive when the Creodonts became extinct. With the Creodonts gone, the Miacids came out from the trees to explore new niches and became the precursors to the modern Carnivora. The familiar Carnivora families began to appear toward the end of the late Eocene. The primary features of carnivores include: the development of carnassial dentition (specialized cutting teeth), retention of grinding molars while reducing the number of teeth, enlargement of the canines, digitigrades stance, enlarged brains and, for some, cursorial locomotion, were the keys to their evolutionary success and remains the distinction of this order. It is important to note that the evolution of the carnivores paralleled the evolution of their prey species, whose development they influenced and were influenced by. As prey became larger and swifter, so then did the predators. All evolved within the larger picture of changes in vegetation types, changing climates, and drifting continents. No system is static, least of all an ecological one. What we see today is a moment on a continuum that moves into the future. When you observe an animal, ask yourself how it may have come to its present form. Mesozoic mammals were by and large small nocturnal creatures with long noses, acute hearing and whiskers for feeling their way about in the dark. Bare scaly tails are in part temperature-regulation devices; through such tails mammals can radiate excess body heat. This attribute may descend from pre-mammalian days.

Evolution of families Canoidea (dog-like forms) - typically possess a long snout and non-retractile claws:
Canidae (dogs, wolves, jackels, foxes) – currently not at the zoo This family has a natural distribution that is nearly worldwide. All continents have indigenous representatives except Australia and Antarctica (the dingo being introduced to Australia by humans prior to European exploration). The dog was not the first domesticated animal (sheep & goats preceded them) but was the first domesticated carnivore, and the first animal in Europe to be domesticated. Evidence of canine domestication begins nearly 10,000 years ago. Canidae have adapted to occupy an enormous range of environments, from Arctic tundra to equatorial desert. Evolution of the canids within Carnivora is somewhat controversial. Historically, the Canids were once viewed as the oldest group of carnivores, but recent evidence actually points to them as the youngest. Certainly they reached their apex as a group within the recent past. Originating in North America, the early dogs could not compete in the niches already occupied by the large successful cats. The dogs were more generalist hunters, and evolved long legs well adapted for walking long distances and for endurance running. These adaptations allowed them to range far and survive in marginal environments, which could not support a big cat. Perhaps to help them deal with raising young in these less friendly areas, the dogs also began to develop the social necessary to live cooperatively in packs. About 4 mya, the global environment changed. The world became drier, and grasslands expanded while forest retreated. Dogs were well positioned to take advantage of hunting the more open spaces without cover. Dogs had the stamina and group teamwork needed to hunt fast running prey in the open allowing them to become the most successful and widespread hunters on Earth. It is only within the last few thousand years that mankind became numerous enough to spread out and displace them as the world’s top predator. Ursidae (bears) Widely distributed the Ursidae family is found in the Arctic southward into both Eurasia and North and South America, and until recently in the Atlas Mountains of North Africa. The Ursidae family emerged as a separate Carnivora family during the Oligocene (34 - 23 mya). Early fossils, once thought to be ancestral to the Canidae because of similar dentition, were reclassified as Ursidae with new and more complete findings. The bears specialized early into the niche of large opportunistic omnivores. There are presently three subfamilies of bears two of which appear in the early in the Pleistocene (2 mya), the Tremarctinae (New World bears and the Ursinae (Old World bears). The third subfamily consists of the Giant Panda that was once thought to be a member of the Procyonidae (Raccoons) family. It is thought that the Panda diverged from the Ursidae sometime during the Miocene (23 - 5 mya). Of the Tremarctinae bears only the spectacled bear survives, though a number of genera existed in North America during the last Ice Age. The Ursinae subfamily originally evolved in Eurasia but subsequently expanded into North America. Today bears occupy a wide range of environments, from Arctic ice floes to tropical forests. Procvonidae (raccoons, coatimundi) There are two subfamilies that are traditionally found; Ailurinae (the lesser panda) found in the Himalayas and nearby areas in eastern Asia; and Procyoninae (all others) found in the temperate and tropical areas of the Western Hemisphere. At one time, the giant panda was thought to belong to Ailurinae, but recently genetic studies show that it actually belongs to the Ursidae family.

Ailuridae (red panda) The red panda is the only living species of the genus Ailurus and the family Ailuridae. The taxonomic classification of the red panda has been controversial since it was discovered. It has been previously placed in the raccoon and bear families. Mustelidae (otters, ferrets, weasels, badgers) This large family has a natural distribution that is nearly worldwide with indigenous representatives on all continents except Australia and Antarctica. Mustelidae was probably one of the first modern carnivore families to appear, but poor fossil evidence clouds the earliest origins of this group. Mustelids are certainly separate and evident by the mid Oligocene (30 mya).all five subfamilies had diverged from each other by mid Miocene (15 mya). Found at that time throughout the northern hemisphere, they expanded into Africa and South America during the Pliocene (5 - 1.8 mya). Mephitidae (skunks, stink badgers) note: formerly of Mustelidae Family Three of the four genera of skunks inhabit the New World, collectively ranging from Canada to central South America; the exception are stink badgers, which occur on islands in Indonesia and the Philippines. Mephitids, meaning "stench", are mainly omnivorous. They often eat vegetation, insects and other small invertebrates, and smaller vertebrates such as snakes, birds and rodents. Mephitids are nocturnal, and inhabit a range of habitats that includes woodlands, deserts, grasslands, and rocky montane areas. Pinnipeds [Phocidae (earless or true seals), Otariidae (eared seals - fur seals & sealions), and Odobenidae (walrus)] See the Pennipedia Study Guide. The pinnipeds have specialized adaptations to aquatic life.
Feloidea (cat-like forms, obligate carnivores) typically possess a shortened snout and retractable claws:
Viverridae (civets, genets) – currently not at the zoo This family is found in southwestern Europe, southern Asia, and the East Indies, Africa and Madagascar. Certain genera have been introduced to areas in which the family does not naturally occur. At one time, it was thought that the Herpistidae were part of this family. However, scientists have concluded that the Herpistidae have sufficient anatomical differences so they are now in the new family. The geological range of this family is Oligocene to recent in Europe, Miocene to recent in Asia and Africa, and Pleistocene to recent in Madagascar. Herpestidae (meerkat, fossa, mongoose) – currently not at the zoo This family is found in southern Asia, the East Indies, Afreica and Madagascar. One genus (Herpestes) occurs in Spain and Portugal, probably brought there by people in ancient times, and also has been introduced in many parts of Europe and on many islands around the world. These species were once part of the Viverridae family, but now warrant full familial distinction, although there is still some disagreement over the genus Cryptoprocta (fossa). The geological range of this family is Oligocene to recent in Europe, Miocene to recent in Asia and Africa, and Pleistocene to recent in Madagascar. Hyaenidae (hyena, aardwolves) – currently not at the zoo Hyenas first arose in Eurasia during the Miocene period from viverrid-like ancestors, and developed into two distinct branches; the lightly built dog-like hyenas and the robust bone-crushing hyenas. Although the dog-like hyenas thrived 15 million years ago they died out after

a change in climate along with the arrival of canids into Eurasia. Of the dog-like hyena lineage, only the insectivorous aardwolf survived, while the bone-crushing hyenas became the undisputed top scavengers of Eurasia and Africa. Felidae (cats) This family has a natural distribution that is nearly worldwide with indigenous representatives on all continents except Australia and Antarctica. Last of the domesticated predators, the house cat has a long and distinguished pedigree. However, many of its behaviors still reflect the evolutionary development of a family of animals considered the most highly developed of all carnivores. While this may be argued on several points, all cats are superbly adapted to overpowering and killing strong agile prey. The first cats to appear during the Miocene were the size of jungle cats, and were forest dwellers. These earliest cat fossils are found in Africa. As with the earlier Creodonts, the cats gave up their grinding teeth and developed their carnassial teeth for a meat-eating diet. Felidae hunters, with their stalk and ambush techniques, were more intelligent than their ancient predecessors. These traits combined with their fast sprinting ability and powerful forelimbs consequently proved more successful. The ancient Felidae evolved not only the modern cats we know today, but also a subfamily of saber-toothed cats. These cats were common in Eurasia and North America, and even extended into South America during the glacial period. These cats became extinct about 10,000 years ago, possibly because the large prey species they relied on (mastodonts, mammoths, ground sloth, and many large perissodactyls) also died out during this period. While this is considered, in geologic terms, a ‘minor extinction’, many other large successful predators also disappeared around the same time, including the North American Lion that closely resembled today’s African Lion but was 35% larger. For the most part the cats have retained the tree climbing abilities of their Miacid ancestors.
Physical & Behavioral Adaptations For mammals the digestion of cellulose is a formidable physiological problem, but the digestion of flesh is easy. One cannot, however, have it both ways; flesh may be easily digested but is hard to come by. Moreover, prey species cannot support an equal number of predators, so the business of the killer is a highly competitive as well as a highly skilled occupation. Much of their fascination lies in the fact that they are so clearly adapted to their mode of life - the otter in pursuit of a fish; the cheetah on a Thompson’s gazelle; the lightning-quick pounce of the fox on a vole; even the domestic cat stalking a mouse - each in its own way, seems to display a grace and perfection of movement which we both envy and admire. (Ewer, 1973) Carnivora means meat-eater but, as explained in the introduction, this trait is not unique to the Carnivora. A better distinction might be their food capturing (or predatory) abilities, the range of physical and behavioral adaptations, which help them sense, seize, and kill their prey. Predation was an early Carnivora adaptive strategy, and most modern members of the order remain predators. Nearly all Carnivora species are more or less carnivorous. Most species are very efficient killers, with a range of specific adaptations, which assist them in hunting. As mentioned above meat is more easily digested than plant material, so Carnivora have evolved simple stomachs and short intestinal tracks.

Generally, carnivores bear altricial young, which undergo a long period of learning and physical development. This is a “high-cost” reproductive strategy (few offspring that require significant parental time investment for their survival). A large advantage of this strategy is that is has allowed Carnivora offspring to have brains with well developed cerebral hemispheres and great potential for learning, providing them a certain adaptive flexibility. (This type of reproductive strategy has been exploited to its current apex in the Primates). Carnivores tend to be intelligent and mentally alert, abilities that assist them in the complex business of outwitting and capturing their prey. The carnivores adapted, originally, to a general predatory way of life and each of the seven families have specialized further, developing certain physical features to the detriment of others. All species are more or less carnivorous (except a few which have become secondarily herbivorous), are efficient killers, and show specific adaptations toward this end. Generally, carnivores bear altricial young, which undergo a long period of learning and physical development. Their brains have well developed cerebral hemispheres and great potential for learning. It is this period of training, the physical traits of each species, and the learned skills that typify the carnivores. When studying or observing these species, keep in mind how each animal has specialized toward efficient predation.
A. SKELETON Carnivora have generalized mammalian skeletons, except for the limbs and head, which accommodate a wide variety of motions: the complex movements of tracking and killing including running, swimming and climbing. Clavicles tend to be reduced (compared to most mammals) providing a greater range of mobility for forelimbs. These structural adaptations help support the complex movements Carnivora require for tracking and killing, which include running, swimming and climbing. (All Carnivora species, if need be, are good to excellent swimmers). The length of the tail is the most variable form on the skeleton, lengthy ones are used for balance (for running on the ground or in the trees) or in the case of the kinkajou hanging in the trees. Differences in the skull of carnivores lie in the elongation of the snout and the development in some species of prominent sagittial and occipital crests for the attachment of neck and jaw muscles (e.g. tiger). Carnivora range in size from that of the Least weasel’s mere 35 grams (½ oz.) to the Polar / Grizzly bear’s 800 kg (1,760 + lbs.) Limbs can very widely in length from the short, low slung weasel to the long legged, erect wolf. There is also a major difference in methods of walking from the plantigrade walk of bears (ursids) to digitigrade found among the canids and felids. All species are clawed and all have at least four digits on each foot though the first is sometimes reduced or absent. Claws very widely in there ability to retract: fully retractable and sharp in all cats except the cheetah, and fully exposed and worn in dogs and hyenas. Some animals are better adapted to climbing (e.g. Black bears, S. American Grey fox, Leopard) by having the ability to rotate the hind and fore limbs. Others without this feature are better suited for running. In many there is a strong tendency toward the reduction in the size of the clavicle. All species are clawed and all have at least four digits on each foot. The first digit is sometimes reduced or absent. Claws vary in their ability to retract: fully retractile and sharp (e.g. most cats), or exposed and worn (e.g. dogs, bears, and hyenas). The one shared characteristic that distinguishes Carnivora from other carnivorous orders is in their ancestral line all possessed four carnassial teeth. When referring to the skull diagrams in

the Handbook, recall the foods of each species and the need for strong chewing muscles and/or killing bite. B. Dentition The generalized, “typical” carnivore dental formula is: (i3/3, c1/1, pm 4/4, m2/2) x 2 = 40. Canidae dentition matches this norm though the canines are large but not necessarily very sharp. Incisors are moderately large and used with the canines for gripping and tearing. The canines are often used as a killing device, either to penetrate the spinal cord of the prey at the neck or close off the windpipe. Carnassial teeth (the last upper pre-molar and the first lower molar) are specially adapted to cutting flesh with a scissor-like action. Their occlusal surfaces are sloped to allow them to slide past one another when the jaw closes. The teeth behind these molars are designed for crushing, similar to ours. The lower canines are set a little in advance of the uppers with some gaps (called the diastimatic gap) allowing the jaw to interlock when it closes. Jaw articulation allows open and shut movements, but not side to side. The Felids have the most specialized carnassials whereas the Ursids have the more specialized crushing molars. Specifics of skeleton and dentition by family Canidae: digitigrade and highly cursorial; limbs typically long; mostly meat-eaters (except for the raccoon dog - vegetation and insects and the Bat-eared fox - primarily insectivorous). Jaws are long with typical cutting carnassials and crushing molars. The African hunting dog, and Bush dog all have 42 teeth (i3/3, c1/1, pm 4/4, m2/3) x 2 = 42) Dhole [Asian wild dog] is (i3/3, c1/1, pm 4/4, m2/2) x 2 = 40) while the Bat-eared fox has 48 and the Bush dog with 38. Ursidae: plantigrade, large heavily built animals with long non-retractile claws and reduced tail. These omnivorous animals have carnassials that crush rather than slice. They are molar-like in the Giant panda and spectacled bear. By losing the premolar with age their dental formula reduces from 40 to 38. Procyonidae: plantigrade to semi-plantigrade with long tails, most of these animals are good climbers. These omnivorous members have the more crushing type carnassials (e.g. Red panda). Most have 40 teeth while the kinkajou has only 36. Ailuridae: Red Pandas are generally solitary. They are well adapted for climbing and spend much of their time off the ground. They are plantigrade; this affords greater stability and weight-bearing ability and is helpful when standing your ground or trying to balance in trees. Mustelidae: are typically long-bodied animals with short legs with either a semi-plantigrade or digitigrade stance. With short and powerful jaws they are capable of accurately placing a death bite like cats. The dental formula varies from 28 - 38; most have sharp canines and cutting carnassials while other; skunks are omnivorous, otters eat fish, frogs, crayfish and crabs. Mephitidae: are recognized by their striking color patters. Coloration in skunks and stink badgers serves as an aposematic signal to would-be predators. All mephitids have extremely well-developed anal scent glands with which they produce noxious odors to deter threats. Pinnipeds: aquatic animals. See the Pinnipedia Study Guide.

Felidae: digitigrade, typically with sharp retractile claws generally protected by lobes of skin, forming sheaths; braincase large and shaped for maximum jaw attachment (sagittial and occipital crests); jaws short and well muscled; carnassials and canines highly developed; most specialized and strictly carnivorous predators with a d f of (i3/3, c1/1, pm 3/2, m1/1) x 2 = 30). Viverridae: are small animals with relatively short limbs, semi-plantigrade to digitigrade stance. Claws typically are non-retractile, though there are exceptions. Head is rather long and low, with jaws not shortened; carnassials not highly specialized with dentition varying from 32 - 40. Herpestidae: resemble the Viverridae somewhat, but are smaller; fur is more uniform in color than the viverrids, although some have stripes; the body is long, limbs rather short, and tail is relatively short. The exception to is the fossa, which has an extremely long tail. Most genera have five toes on each feet; claws are usually not retractile. Skull commonly shorter and broader than viverrids; commonly 36-40 teeth. Hyaenidae: are medium to large animals with relatively heads and forelimbs longer and heavier than hindlimbs. With digitigrade stance they are not as specialized as Felidae they have the ability to crush bones to a greater degree than any other carnivore. Their dental formula is (i3/3, c1/1, pm 4/3, m1/1) x 2 = 34) while the aardwolf that feeds mainly on termites, not only has reduced tooth size, but the number may be as few as 24 in adults.
C. Pelage (Fur) Fur is of course, a general mammalian adaptation for temperature regulation. Many carnivores in northern climates stay active year round and are protected mainly by their fur. A Siberian husky can sleep at - 49° F and an Arctic Fox at even colder temperatures with no change in metabolic rate. Each animal may be able to vary the effectiveness of its fur by wetting or fluffing, curling up to protect limbs and nose, and controlling peripheral blood circulation. To adapt to temperature changes that otherwise have prevented their movement into northern habitats, Arctic species often shed their very heavy coats in spring and shed again in autumn to restore the winter coat. The primary function of various colors and patterns of fur is concealment, either defensive or offensive. In general, the super-family Canoidea has uniform coloration, while the super-family Feloidea has a varied coloration, which may suggest a divergence in their early evolution into open country hunters. (Canoidea), and forest predators (Feloidea). Desert species tend toward light, sandy colors while forest or woodland species are darker, often with mottling to match light and shade patterns. Northern carnivores are often whitish or have seasonal variations. Some colorations are for warning, as in some Mustelids which use anal gland secretions for defense (i.e. skunks). Coloration, especially around the face, ears and tail, is also often a factor in complex intra-specific communication. It may be used for warning or in everyday socialization. D. Skin Glands Skin glands assist in thermoregulation and provide suitable dressing for the hairs. The majority of carnivores have also secondarily developed some of these glands for olfactory communication. Gland locations vary: cheeks (mongoose), abdomen (Mustelids), throat (Procyonids), root of tail (badger, otter, and cat) dorsal surface of tail and between paws (canids), and the ano-genital region, which has the most complex odor producing glands. E. Sense Organs

Since it is necessary for predators to have highly effective sense organs, each species has developed at least one superb faculty of the three main senses: hearing, sight and smell. Specific sense development is most often in response to the carnivore species’ interaction with their environment and their typical prey. Predators whose game is small or who hunt in dense cover at night, often have well developed olfactory or hearing. Those that run down their prey over long distance often have well developed vision. Depth perception is well developed in most carnivores. Their eye placement (toward the front of the head) allows for binocular stereoscopic vision (though this is not as highly developed as it is in primates). Many carnivores have adapted to see well both at night and by day. Felids and other nocturnal carnivores have very good night vision due to a well developed tapetum lucidum. As light passes into the lens of the eye, it is focused on the retina layer, which forms a membrane at the back of the eye. Behind this membrane lies the tapetum lucidum, which reflects light back out past the retina again. This allows the small amount of night light received to stimulate the receptors (rods and cones) in the retina twice. While they are sensitive to very small amounts of light at night, during the day they can close their pupils almost completely so as not to be blinded in bright sunlight. By comparison humans cannot do this easily or effectively because we have a different type of iris. Even with this eye sensitivity most carnivores do not have the ability to distinguish colors.
F. Behavioral Generally, Carnivores are opportunistic feeders, relying to a large degree on what is available, and are quite adaptable in their feeding habits. Many of the large carnivores are further adapted to scavenging other’s kill or eating carrion. Yet most are able to eat a great deal of plant material as well as meat. The diets of Carnivores that eat significant quantities of vegetable matter may vary seasonally. What a carnivore kills is dependent on several factors relating to their possible prey species:
• Availability: (abundance in the home range due to seasonal or other variations) • Vulnerability: (ease of kill depending on age, health, coloration, habitat, and herd
verses solitary existence) • Palatability: (unqualified but obvious) • Experience of the individual hunter: Carnivora have the capacity to learn, and adjust
their hunting behavior from experience. Carnivora exhibit two primary types of hunting strategies: social and non-social.
1. Social hunters (exemplified by the large Canids, spotted hyena and the lion): • They are highly predacious and kill large prey by running it down. • Group pursuit provides for greater kill success, sometimes able to catch surplus
food for short or long periods up to a season. • A second type of social hunter is not highly predacious but forages on small prey
(largely invertebrates), and varied types of food (omnivorous), and is of small size. Examples include the coati (Procyonidae) and the mongoose (Herpestidae). Here, the group provides the advantages of increased vigilance and defense.
2. Non-Social hunters are not necessarily “solitary” all year round or at all stages of their
lives (breeding and family rearing being typical exceptions). Examples of this type of predator include the felids, otters, weasels, foxes, skunks and raccoons. There is no typical food finding pattern in this group.

Large carnivores primarily hunt individuals of their prey species who are young, elderly, sick or injured. Most large prey species have evolved sufficient defenses so that attacking an individual in its prime can be costly to the predator (especially when one considers that the predator must do so, frequently to survive). The species in Carnivora kill their prey in various ways. Members of the families Viverridae and Mustelidae could be referred to as “occipital cruncher” since they smash the back of their prey’s brain case with their teeth. After the stalk and ambush of a cat hunt, the prey is typically killed with a bite to the nape of the neck. Placing a canine tooth between two vertebrae breaks the spinal cord. Sometimes they also kill by strangulation or by crushing the back of the skull. Dogs, on the other hand, bring their prey down from the rear and open the belly to feed, or execute the “mouse jump” while hunting alone or in pairs - pouncing on smaller prey (typically lagomorphs or rodents). They pin the prey down with fore paws and dislocate the prey’s head by biting the nape of the neck and shake it vigorously. Members of the bear family, with exception of the polar bear, are the least predacious family of Carnivora. Their diet consists mainly of plant material, making their food habits also dependent on the season.
G. Locomotion Most Carnivora are terrestrial, but methods of walking vary from plantigrade (ursids) to digitigrade (canids and felids). Cursorial locomotion (fast running speed) is characteristic of many of the carnivores. However a few are better adapted to climbing (e.g. bears, gray fox and ocelots) by the ability to rotate the leg and arm, hand and foot. Tails are often used as an aid in locomotion: for example lengthy tails are used for balance (in running on the ground or in the trees), or for hanging (e.g. kinkajou). All Carnivora can swim if necessary but polar bears and river otters are semi-aquatic while sea otters live most of their lives in the ocean. H. Communication & Other Behavior Scent plays a role in individual recognition, assessment of sexual status, and in interactions between mother and young. Scent marking is a long-lived signal which most species mainly use to define territory. Urine and feces are frequently used, especially with the admixture of certain glands secretions to aid in their effectiveness. These may be deposited on standing or piled up objects. Tree scratching, biting and rubbing are practiced by certain species (bears, cats et al.) sometimes with the deposition of scent or saliva. Scent can indicate several things.
• Scent most certainly shows the sex of the individual and probably its exact identity. • It may reveal time since deposition (allowing two individuals to use the same path and
not encounter one another), size and/or strength of the donor, and readiness for sexual contact.
• There is no doubt that the depositing of scent and its reception both elicit psychological responses. For example, scent plays a major roll in bringing about reproductive behavior, stimulating physiological changes in possible mates and readying each for the other. Anxiety and aggression can be alleviated or increased, and various behaviors can be altered. All this to the species’ advantage, assuring biological continuity (reproduction, proper spacing, etc.).
Visual, vocal and olfactory signals are used by social and solitary mammals for purpose of group cohesion and peaceful coexistence, or to avoid contact or notify of short term readiness for contact. Some carnivores are such efficient killers (e.g. the Felidae); the complex signals probably play an essential role in the continuation of the species. Visual signals may be expressed by face, bared teeth, twitching tail, laid back ears, crouching, inflating or back up

posture, raised mane of neck and shoulders, sudden barks, hisses or spitting. Dominance systems (hierarchies) of the more social species often rely on these signals to prevent fighting. Sounds also play a role in social interactions, frequently accompanying visual cues, and are quite numerous in carnivores. Auditory signals may at one and the same time serve to locate companions and discourage the approach of strangers (i.e. lion roar). The carnivores can often afford to be noisier than other animals because there is usually less fear of predation. I. Courtship & Young While the reproductive processes are broadly similar in the placental mammals, the predatory life has required some adaptations: because predators are generally solitary and range over wide areas, the early stages of the reproductive cycle may be more drawn out. Males and females must locate one another and come to terms when they previously may have been hostile and will soon be again. Therefore, in most species, the female estrus period is long, being over several days with repeated copulation over this period. Also helpful is the phenomenon of induced ovulation, not exclusive to carnivores but found more commonly in this order (and in the Pinnipeds). Here, ovulation, the release of the egg from an ovary, is triggered by copulation. Most carnivores may be induced ovulators, but it has only been shown in house cats, weasels, martens, stoats, spotted skunks, raccoons, American river otters, dogs, and foxes. Delayed implantation appears to be an adaptation, which permits the timing of mating and the timing of birth to be independently adjusted to the environmental situation (i.e. for timing births in seasonal environments so that the young are born at periods when food is most abundant or when weather is the mildest). In most mammals, the egg is fertilized after ovulation as it travels down the oviduct from the ovary. It then implants in the uterine wall to complete development. With delayed implantation, cell division begins in the fertilized egg and the blastocyst reaches the uterus but does not implant. It becomes inactive for varying periods of time, up to several months, until reactivated. The process is poorly understood, though no doubt it will be found to be a combination of psychological and physiological factors. Amongst carnivores, delayed implantation has been found to occur in a number of the Mustelids and ursids. One to two litters per year is typical, except in the larger species which tend to have a longer cycle of one every two years. Most carnivores are born furred but with the eyes closed. Although they are capable of crawling, locomotory powers are limited. Some smaller Mustelids and Viverrids, and some bears, are even more altricial, and are protected by a den or burrow. On the other hand, precocial young are found in the sea otter, spotted hyena and fossa. Animals that do not have precise breeding and birthing cycles may show year round activity but usually have peak periods and these often reflect some environmental changes (prey movements, plants maturing, etc.). There is a great deal to learn from experience as a carnivore juvenile (relating to food and killing), so even after weaning, the young stay with the mother and she does not mate again until they are independent. A major activity of most young carnivores is to play at hunting and fighting, developing adult skills and often establishing dominance.
Status & Conservation Predators are often the first animals to noticeably disappear from a threatened ecosystem. They are disproportionally highly represented on lists of endangered species. Carnivores are often considered “keystone” species, playing a greater role in the ecosystem than their numbers might

indicate, and their loss often leads to much greater ecological disruptions. Conversely, they are often “umbrella” species for conservation efforts. Large and easily recognizable, they can serve as a rallying point for ecosystem protection efforts. Threats to Carnivora Carnivores are high on the food chain, and as such must always be less numerous than their prey species. This also means that carnivore species have much higher space and resource requirements for the maintenance of viable populations. For this reason, habitat destruction and fragmentation are particularly devastating to Carnivora. Unfortunately, habitat fragmentation is often the first effect of human encroachment, lowering the carrying capacity of an area and creating possible barriers within a population. Such fragmentation tends to reduce the genetic diversity of a population, increasing its vulnerability to secondary adaptive pressures such as disease. For example, expansion of human settlements with domestic dogs have brought a variety of canine diseases which, in some cases, have eradicated indigenous canids from their historic ranges. The “high cost” strategy of reproduction (few offspring that require significant parental time investment for their survival), which most carnivores employ in order to raise successful hunters, reduces the rate at which a population can rebound once reduced. Members of the Ursidae family are an extreme example of this. Historically, carnivores have been specifically hunted by humans for many reasons, particularly the large carnivores. Wolves and bears were “cleared” from Europe long ago. In North America and much of the rest of the world this trend continues. Reasons for this include:
a) Carnivores were perceived as threats to humans or domestic livestock. b) They were seen as competitors for prey species c) They were hunted for the status associated with killing a “killer” (i.e., trophies) d) Individual and later consumer, demand for carnivore body parts, particularly fur.
In fact nearly all Mustelids are hunted (or raised and killed specifically) for their pelts. The sea otter was nearly exterminated for this reason (the California population remains about 2000 individuals in spite of being under international protection since the 1920s). Conservation efforts While still a small percentage of the order, there are many Carnivora species with conservation breeding programs. Besides the fact that carnivores are often particularly threatened, carnivore intelligence, their ability to adapt to new situations and their natural propensity for social solitude assist them in their candidacy for both captive breeding programs and re-release efforts. Many species have done well in captive breeding programs, once established, but a number of species or subspecies have become endangered too fast for programs to be set up. Another major problem for captive breeding programs has been the legal road blocks put up by conservation groups that view these programs as cruel and ineffective. Unfortunately, habitat destruction, fragmentation and hunting continue to take their toll while court battles are fought. The Florida panther, black-footed ferret and the California condor are all examples of efforts that faced these difficulties. Helping to reduce this reluctance through education should be one of our main priorities.

There are a number of Carnivora Species Survival Plans (SSPs), representing both species and subspecies. Canids Others Felids Red wolf Spectacled bear Tigers (Siberian, Sumatran)* Maned wolf Lesser or Red panda Asian lion African hunting dog Black footed ferret Cheetah Asian small-clawed otter Clouded leopard Snow leopard* * animals at the San Francisco Zoo CARNIVORA BIBLIOGRAPHY updated: 10/01, 3/14 Grzimick., Encyclopedia of Mammals, © 1991 Walker, Mammals of the World, © 1991 Ewer, R.F., The Carnivores, Cornell University Press, NY. © 1973 Rosevear, D.R. Carnivores of West Africa, British Museum, London ©1974 Kingdom, J., East African Mammals, Vol. I & II, Academy Press ©1982 Eisenberg, J.F., The Mammalian Radiation, University of Chicago Press © 1981 Halstead, L.B., The Evolution of the Mammals, Hippocrene Books © 1981 Prothero, D.R., After the Dinosaurs: the Age of Mammals, Indiana University Press © 2006
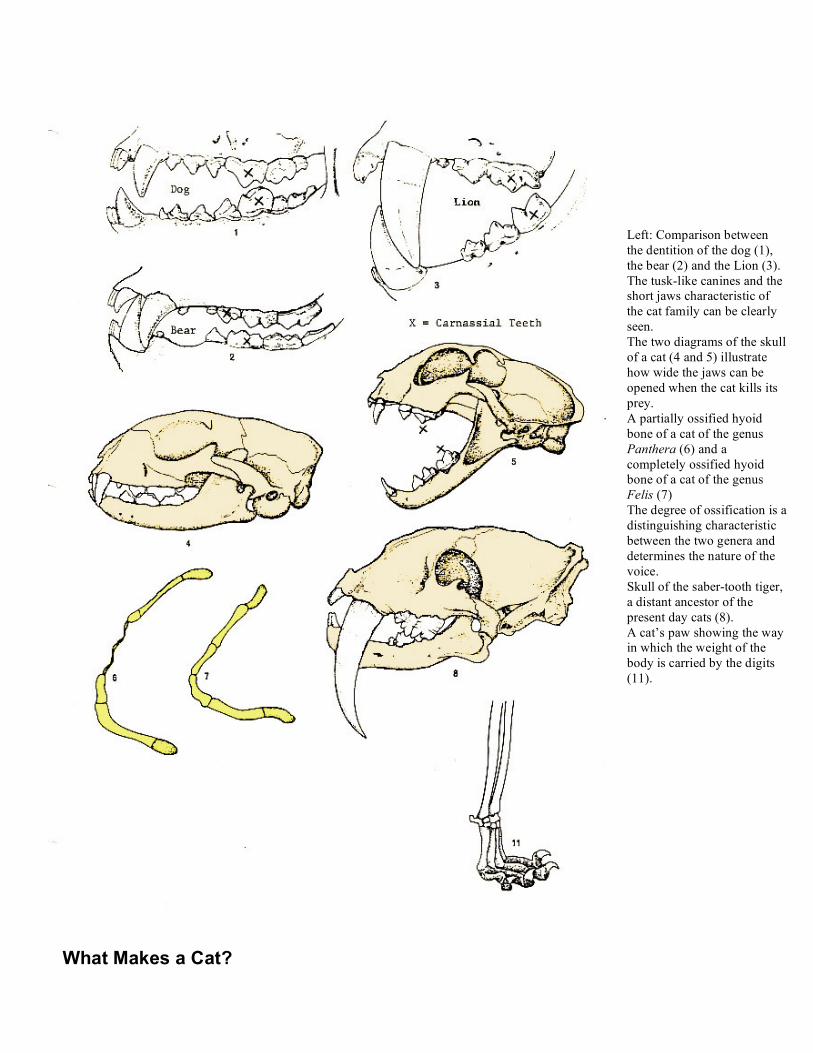
What Makes a Cat?
Left: Comparison between the dentition of the dog (1), the bear (2) and the Lion (3). The tusk-like canines and the short jaws characteristic of the cat family can be clearly seen. The two diagrams of the skull of a cat (4 and 5) illustrate how wide the jaws can be opened when the cat kills its prey. A partially ossified hyoid bone of a cat of the genus Panthera (6) and a completely ossified hyoid bone of a cat of the genus Felis (7) The degree of ossification is a distinguishing characteristic between the two genera and determines the nature of the voice. Skull of the saber-tooth tiger, a distant ancestor of the present day cats (8). A cat’s paw showing the way in which the weight of the body is carried by the digits (11).
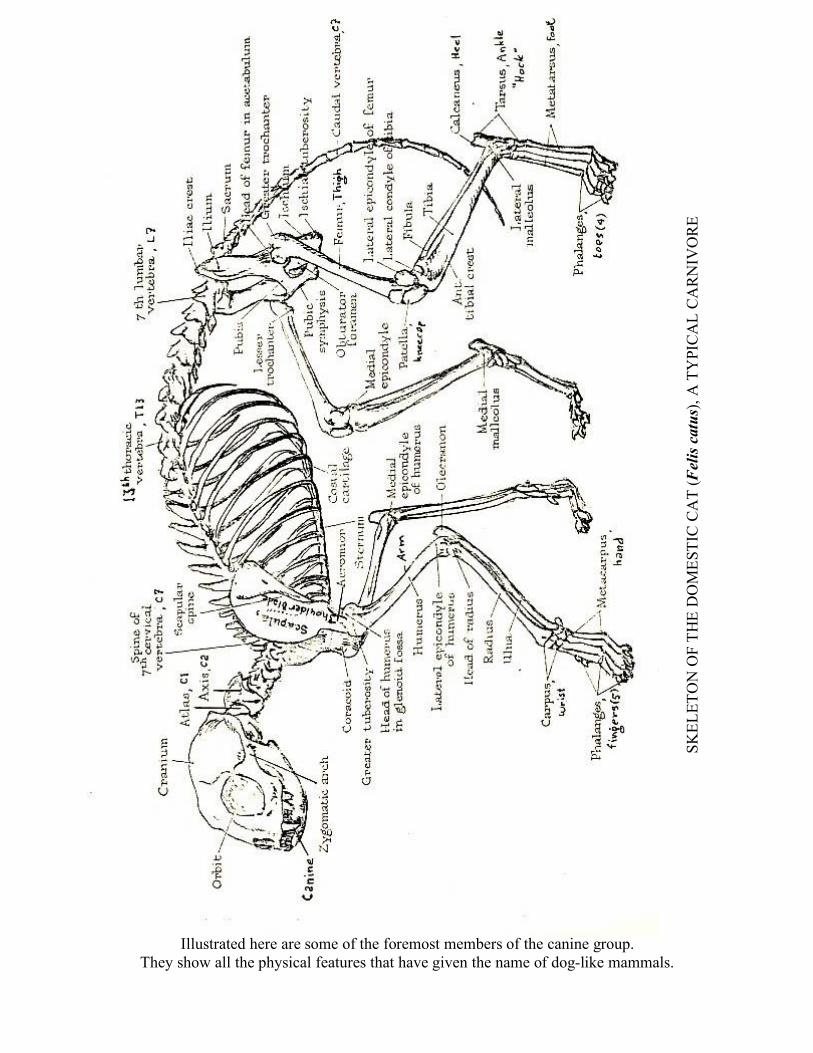
Illustrated here are some of the foremost members of the canine group. They show all the physical features that have given the name of dog-like mammals.
SKEL
ETO
N O
F TH
E D
OM
ESTI
C C
AT
(Fel
is ca
tus)
, A T
YPI
CA
L C
AR
NIV
OR
E
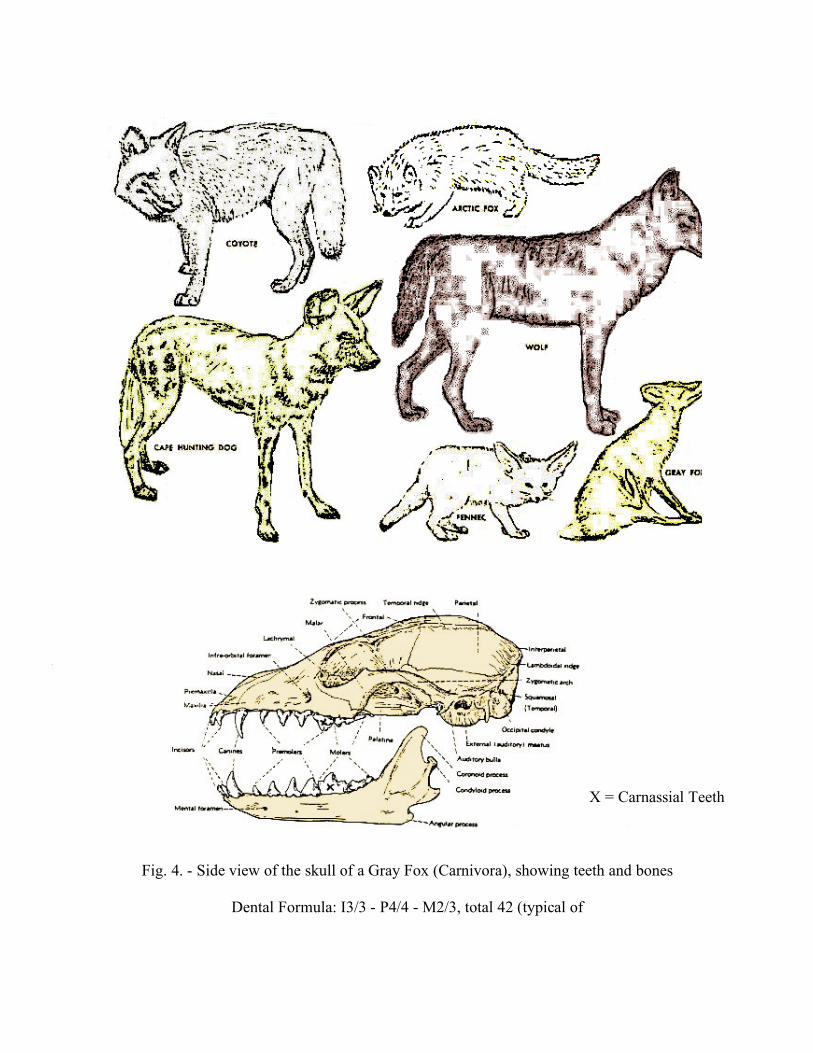
X = Carnassial Teeth
Fig. 4. - Side view of the skull of a Gray Fox (Carnivora), showing teeth and bones
Dental Formula: I3/3 - P4/4 - M2/3, total 42 (typical of





























